Effect of Heat Stress on Growth and Physiological Traits of Alfalfa (Medicago sativa L.) and a Comprehensive Evaluation for Heat Tolerance

Abstract
1. Introduction
2. Materials and Methods
2.1. Plant Materials, Growth Conditions, and Heat Treatment
2.2. Plant Biomass Measurement
TDB = SDW + RDW
2.3. Physiological Traits Analysis
2.4. Relative Water Content (RWC) Measurement
2.5. Chlorophyll Content Determination
2.6. Electrolyte Linkage (EL) Measurement
2.7. Soluble Sugar Content Determination
2.8. Preparation of Crude Enzyme Extract
2.9. Determination of MDA Content
2.10. Soluble Protein Content Determination
2.11. A Comprehensive Evaluation of Alfalfa for Heat Tolerance
2.12. Chlorophyll a (Chl a) Fluorescence Analysis
2.13. Statistical Analysis
3. Results
3.1. Effect of Heat Stress on Alfalfa Plant Biomass
3.2. Heat Stress Reduced the Relative Water Content (RWC) of Alfalfa
3.3. Heat Stress Increased the Electrolyte Leakage (EL) of Alfalfa
3.4. Effect of Heat Stress on Lipid Peroxidation
3.5. Heat Stress Decreased the Chlorophyll Content of Alfalfa
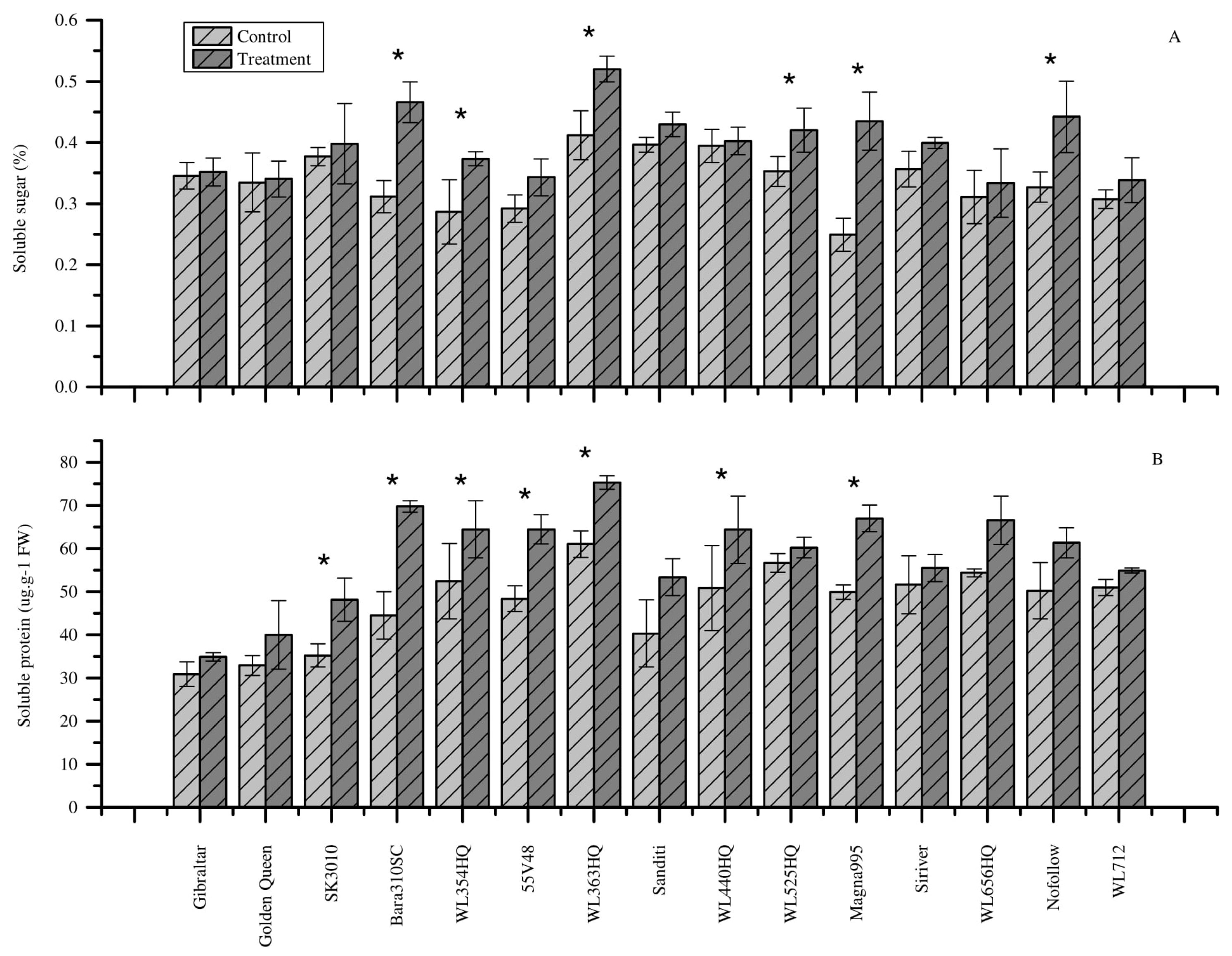
3.6. Effect of Heat Stress on Soluble Sugar Content
3.7. Effect of Heat Stress on Soluble Protein Content
3.8. A Comprehensive Evaluation of the Heat Tolerance of Alfalfa Cultivars
3.9. Alteration of Chlorophyll a Fluorescence under Heat Stress
3.9.1. OJIP Transient Curve
3.9.2. Basic Photosynthetic Parameters (F0, Fj, Fi, Fm, F300 μs, and Fv/Fm)
3.9.3. Specific Energy Fluxes (TP0/RC, ETO/RC, RE0/RC, and ABS/RC)
3.9.4. Quantum Yield and Efficiency (φpo, φEo, δRo, and RC/ABS)
3.9.5. Performance Indexes (PIABS and PItotal)
4. Discussion
5. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Ahuja, I.; de Vos, R.C.H.; Bones, A.M.; Hall, R.D. Plant molecular stress responses face climate change. Trends Plant Sci. 2010, 15, 664–674. [Google Scholar] [CrossRef] [PubMed]
- Hasanuzzaman, M.; Nahar, K.; Alam, M.; Roychowdhury, R.; Fujita, M. Physiological, biochemical, and molecular mechanisms of heat stress tolerance in plants. Int. J. Mol. Sci. 2013, 14, 9643–9684. [Google Scholar] [CrossRef] [PubMed]
- Bañon, S.; Fernandez, J.A.; Franco, J.A.; Torrecillas, A.; Alarcón, J.J.; Sánchez-Blanco, M.J. Effects of water stress and night temperature preconditioning on water relations and morphological and anatomical changes of Lotus creticus plants. Sci. Hortic. 2004, 101, 333–342. [Google Scholar] [CrossRef]
- Wahid, A.; Gelani, S.; Ashraf, M.; Foolad, M.R. Heat tolerance in plants: An overview. Environ. Exp. Bot. 2007, 61, 199–223. [Google Scholar] [CrossRef]
- Li, S.; Li, F.; Wang, J.; Zhang, W.E.N.; Meng, Q.; Chen, T.H.H.; Lycopersicon, T. Glycinebetaine enhances the tolerance of tomato plants to high temperature during germination of seeds and growth of seedlings. Plant Cell Environ. 2011, 34, 1931–1943. [Google Scholar] [CrossRef] [PubMed]
- Vollenweider, P.; Gu, M.S. Diagnosis of abiotic and biotic stress factors using the visible symptoms in foliage. Environ. Pollut. 2005, 137. [Google Scholar] [CrossRef] [PubMed]
- Bi, A.; Fan, J.; Hu, Z.; Wang, G.; Amombo, E.; Fu, J.; Hu, T. Differential Acclimation of Enzymatic Antioxidant Metabolism and Photosystem II Photochemistry in Tall Fescue under Drought and Heat and the Combined Stresses. Front. Plant Sci. 2016, 7. [Google Scholar] [CrossRef]
- Hu, T.; Liu, S.; Amombo, E.; Fu, J. Stress memory induced rearrangements of HSP transcription, photosystem II photochemistry and metabolism of tall fescue (Festuca arundinacea Schreb) in response to high-temperature stress. Front. Plant Sci. 2015, 6, 1–13. [Google Scholar] [CrossRef]
- Xu, W.; Cai, S.; Zhang, Y.; Wang, Y.; Ahammed, G.J.; Xia, X.; Campus, Z. Melatonin enhances thermotolerance by promoting cellular protein protection in tomato plants. Front. Plant Sci. 2016. [Google Scholar] [CrossRef]
- Potters, G.; Pasternak, T.P.; Guisez, Y.; Palme, K.J.; Jansen, M.A.K. Stress-induced morphogenic responses: Growing out of trouble? Trends Plant Sci. 2007, 12. [Google Scholar] [CrossRef]
- Sade, B.; Soylu, S.; Yetim, E. Drought and oxidative stress. Afr. J. Biotechnol. 2011, 10, 11102–11109. [Google Scholar] [CrossRef]
- Barnabás, B.; Jäger, K.; Fehér, A. The effect of drought and heat stress on reproductive. Plant Cell Environ. 2008, 31, 11–38. [Google Scholar] [CrossRef] [PubMed]
- Mo, Y.; Liang, G.; Shi, W.; Xie, J. Metabolic responses of alfalfa (Medicago Sativa L.) leaves to low and high temperature induced stresses. Afr. J. Biotechnol. 2011, 10, 1117–1124. [Google Scholar] [CrossRef]
- Song, Y.; Lv, J.; Ma, Z.; Dong, W. The mechanism of alfalfa (Medicago sativa L.) response to abiotic stress. Plant Growth Regul. 2019. [Google Scholar] [CrossRef]
- Aranjuelo, I.; Irigoyen, J.J.; Sánchez-Díaz, M. Effect of increased temperature and drought associated to climate change on productivity of nodulated alfalfa. Quality in lucerne and medics for animal production. In Proceedings of the XIV Eucarpia Medicago spp. Group Meeting, Zaragoza and Lleida, Lleida, Spain, 12–15 January 2001. [Google Scholar]
- Aranjuelo, I.; Irigoyen, J.J.; Sanchez-Diaz, M. Effect of elevated temperature and water availability on CO2 exchange and nitrogen fixation of nodulated alfalfa plants. Environ. Exp. Bot. 2007, 59, 99–108. [Google Scholar] [CrossRef]
- Erice, G.; Irigoyen, J.J.; Sánchez-Díaz, M.; Avice, J.C.; Ourry, A. Effect of drought, elevated CO2 and temperature on accumulation of N and vegetative storage proteins (VSP) in taproot of nodulated alfalfa before and after cutting. Plant Sci. 2007, 172, 903–912. [Google Scholar] [CrossRef]
- Li, W.; Wei, Z.; Qiao, Z.; Wu, Z.; Cheng, L.; Wang, Y. Proteomics analysis of alfalfa response to heat stress. PLoS ONE 2013, 8, e82725. [Google Scholar] [CrossRef]
- Zhang, Q.; Liu, X.; Zhang, Z.; Liu, N.; Li, D.; Hu, L. Melatonin Improved Waterlogging Tolerance in Alfalfa (Medicago sativa) by Reprogramming Polyamine and Ethylene Metabolism. Front. Plant Sci. 2019, 10, 1–14. [Google Scholar] [CrossRef]
- Huang, X.; Shi, H.; Hu, Z.; Liu, A.; Amombo, E.; Chen, L.; Fu, J. ABA Is Involved in Regulation of Cold Stress Response in Bermudagrass. Front. Plant Sci. 2017, 8, 1–10. [Google Scholar] [CrossRef]
- Hu, L.; Bi, A.; Hu, Z.; Amombo, E.; Li, H.; Fu, J. Antioxidant Metabolism, Photosystem II, and Fatty Acid Composition of Two Tall Fescue Genotypes with Different Heat Tolerance Under High Temperature Stress. Front. Plant Sci. 2018, 9, 1–13. [Google Scholar] [CrossRef]
- Bradford, M. A rapid and sensitive method for the quantification of microgram quantities of protein utilizing the principle of protein-dye binding. Anal. Biochem. 1976, 254, 248–254. [Google Scholar] [CrossRef]
- Tian, Z.; Yang, Y.; Wang, F. A comprehensive evaluation of heat tolerance in nine cultivars of marigold. Hortic. Environ. Biotechnol. 2015, 56, 749–755. [Google Scholar] [CrossRef]
- Bita, C.E.; Gerats, T. Plant tolerance to high temperature in a changing environment: Scientific fundamentals and production of heat stress-tolerant crops. Front. Plant Sci. 2013, 4, 1–18. [Google Scholar] [CrossRef] [PubMed]
- Siddiqui, M.H.; Al-Khaishany, M.Y.; Al-Qutami, M.A.; Al-Whaibi, M.H.; Grover, A.; Ali, H.M.; Al-Wahibi, M.S. Morphological and physiological characterization of different genotypes of faba bean under heat stress. Saudi J. Biol. Sci. 2015, 22, 656–663. [Google Scholar] [CrossRef] [PubMed]
- Ashraf, M.; Hafeez, M. Thermotolerance of pearl millet and maize at early growth stages: Growth and nutrient relations. Biol. Plant. 2004, 48, 81–86. [Google Scholar] [CrossRef]
- Wahid, A.; Close, T.J. Expression of dehydrins under heat stress and their relationship with water relations of sugarcane leaves. Biol. Plant. 2007, 51, 104–109. [Google Scholar] [CrossRef]
- Hameed, A.; Goher, M.; Iqbal, N. Heat Stress-Induced Cell Death, Changes in Antioxidants, Lipid Peroxidation, and Protease Activity in Wheat Leaves. J. Plant Growth Regul. 2012, 31, 283–291. [Google Scholar] [CrossRef]
- Fahad, S.; Hussain, S.; Saud, S.; Hassan, S.; Tanveer, M. A combined application of biochar and phosphorus alleviates heat-induced adversities on physiological, agronomical and quality attributes of rice. Plant Physiol. Biochem. 2016, 103, 191–198. [Google Scholar] [CrossRef]
- Sita, K.; Sehgal, A.; Kumar, J.; Kumar, S.; Singh, S.; Siddique, K.H.M.; Nayyar, H. Identification of High-Temperature Tolerant Lentil (Lens culinaris Medik.) Genotypes through Leaf and Pollen Traits. Front. Plant Sci. 2017, 8, 1–27. [Google Scholar] [CrossRef]
- Dias, A.S.; Barreiro, M.G.; Campos, P.S.; Ramalho, J.C.; Lidon, F.C. Wheat cellular membrane thermotolerance under heat stress. J. Agron. Crop Sci. 2010, 196, 100–108. [Google Scholar] [CrossRef]
- Kumar, S.; Thakur, P.; Kaushal, N.; Malik, J.A.; Gaur, P.; Nayyar, H. Effect of varying high temperatures during reproductive growth on reproductive function, oxidative stress and seed yield in chickpea genotypes differing in heat sensitivity. Arch. Agron. Soil Sci. 2013, 59, 823–843. [Google Scholar] [CrossRef]
- Awasthi, R.; Bhandari, K.; Nayyar, H. Temperature stress and redox homeostasis in agricultural crops. Front. Environ. Sci. 2015, 3. [Google Scholar] [CrossRef]
- Chakraborty, U.; Pradhan, B. Drought stress-induced oxidative stress and antioxidative responses in four wheat (Triticum aestivum L.) varieties. Arch. Agron. Soil Sci. 2012, 58, 617–630. [Google Scholar] [CrossRef]
- Zhang, H.; Liu, X.-L.; Zhang, R.-X.; Yuan, H.-Y.; Wang, M.-M.; Yang, H.-Y.; Liang, Z.-W. Root Damage under Alkaline Stress Is Associated with Reactive Oxygen Species Accumulation in Rice (Oryza sativa L.). Front. Plant Sci. 2017, 8, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Bajji, M.; Kinet, J.M.; Lutts, S. The use of the electrolyte leakage method for assessing cell membrane stability as a water stress tolerance test in durum wheat. Plant Growth Regul. 2002, 36, 61–70. [Google Scholar] [CrossRef]
- Chen, J.; Wang, P.; Mi, H.L.; Chen, G.Y.; Xu, D.Q. Reversible association of ribulose-1, 5-bisphosphate carboxylase/oxygenase activase with the thylakoid membrane depends upon the ATP level and pH in rice without heat stress. J. Exp. Bot. 2010, 61, 2939–2950. [Google Scholar] [CrossRef] [PubMed]
- Awasthi, R.; Kaushal, N.; Vadez, V.; Turner, N.C.; Berger, J.; Siddique, K.H.M.; Nayyar, H. Individual and combined effects of transient drought and heat stress on carbon assimilation and seed filling in chickpea. Funct. Plant Biol. 2014, 41, 1148–1167. [Google Scholar] [CrossRef]
- Dutta, S.; Mohanty, S.; Tripathy, B.C. Role of Temperature Stress on Chloroplast Biogenesis and Protein Import in Pea. Plant Physiol. 2009, 150, 1050–1061. [Google Scholar] [CrossRef]
- Janska, A.; Marsik, P.; Zelenkova, S.; Ovesna, J. Cold stress and acclimation: What is important for metabolic adjustment? Plant Biol. 2010, 12, 395–405. [Google Scholar] [CrossRef]
- Gupta, N.K.; Agarwal, S.; Agarwal, V.P.; Nathawat, N.S.; Gupta, S.; Singh, G. Effect of short-term heat stress on growth, physiology and antioxidative defence system in wheat seedlings. Acta Physiol. Plant. 2013, 35, 1837–1842. [Google Scholar] [CrossRef]
- Han, Y.; Fan, S.; Zhang, Q.; Wang, Y. Effect of heat stress on the MDA, proline and soluble sugar content in leaf lettuce seedlings. Agric. Sci. 2013, 4, 112–115. [Google Scholar] [CrossRef]
- Harsh, A.; Sharma, Y.K.; Joshi, U.; Rampuria, S.; Singh, G.; Kumar, S.; Sharma, R. Effect of short-term heat stress on total sugars, proline and some antioxidant enzymes in moth bean (Vigna aconitifolia). Ann. Agric. Sci. 2016, 61, 57–64. [Google Scholar] [CrossRef]
- Lang-Mladek, C.; Popova, O.; Kiok, K.; Berlinger, M.; Rakic, B.; Aufsatz, W.; Luschnig, C. Transgenerational inheritance and resetting of stress-induced loss of epigenetic gene silencing in arabidopsis. Mol. Plant 2010, 3, 594–602. [Google Scholar] [CrossRef] [PubMed]
- Khan, S.U.; Din, J.U.; Qayyum, A.; Jan, N.E.; Jenks, M.A. Heat tolerance indicators in Pakistani wheat (Triticum aestivum L.) genotypes. Acta Bot. Croat. 2015, 74, 109–121. [Google Scholar] [CrossRef]
- Kumar, S.; Beena, A.S.; Awana, M.; Singh, A. Physiological, Biochemical, Epigenetic and Molecular Analyses of Wheat (Triticum aestivum) Genotypes with Contrasting Salt Tolerance. Front. Plant Sci. 2017, 8, 1–20. [Google Scholar] [CrossRef] [PubMed]
- Porcar-Castell, A.; Tyystjärvi, E.; Atherton, J.; Van Der Tol, C.; Flexas, J.; Pfündel, E.E.; Berry, J.A. Linking chlorophyll a fluorescence to photosynthesis for remote sensing applications: Mechanisms and challenges. J. Exp. Bot. 2014, 65, 4065–4095. [Google Scholar] [CrossRef] [PubMed]
- Krause, G.H.; Winter, K.; Krause, B.; Virgo, A. Light-stimulated heat tolerance in leaves of two neotropical tree species, Ficus insipida and Calophyllum longifolium. Funct. Plant Biol. 2015, 42, 42–51. [Google Scholar] [CrossRef]
- Tan, W.; wei Meng, Q.; Brestic, M.; Olsovska, K.; Yang, X. Photosynthesis is improved by exogenous calcium in heat-stressed tobacco plants. J. Plant Physiol. 2011, 168, 2063–2071. [Google Scholar] [CrossRef] [PubMed]
- Maxwell, K.; Johnson, G.N. Chlorophyll fluorescence—A practical guide. J. Exp. Bot. 2000, 51, 659–668. [Google Scholar] [CrossRef] [PubMed]
- Zushi, K.; Kajiwara, S.; Matsuzoe, N. Chlorophyll a fluorescence OJIP transient as a tool to characterize and evaluate response to heat and chilling stress in tomato leaf and fruit. Sci. Hortic. 2012, 148, 39–46. [Google Scholar] [CrossRef]
- Zhao, B.; Wang, J.; Gong, H.; Wen, X.; Ren, H.; Lu, C. Effects of heat stress on PSII photochemistry in a cyanobacterium Spirulina platensis. Plant Sci. 2008, 175, 556–564. [Google Scholar] [CrossRef]
- Stefanov, D.; Petkova, V.; Denev, I.D. Screening for heat tolerance in common bean (Phaseolus vulgaris L.) lines and cultivars using JIP-test. Sci. Hortic. 2011, 128, 1–6. [Google Scholar] [CrossRef]
- Apostolova, E.L.; Dobrikova, A.G. Effect of high temperature and UV-A radiation on photosystem II. In Handbook of Plant and Crop Stress; Pessarakli, M., Ed.; CRC Press: Boca Raton, FL, USA, 2011; pp. 577–591. [Google Scholar]
- Wen, X.; Qiu, N.; Lu, Q.; Lu, C. Enhanced thermotolerance of photosystem II in salt-adapted plants of the halophyte Artemisia anethifolia. Planta 2005, 220, 486–497. [Google Scholar] [CrossRef] [PubMed]
- Yusuf, M.A.; Kumar, D.; Rajwanshi, R.; Strasser, R.J.; Tsimilli-Michael, M.; Govindjee; Sarin, N.B. Overexpression of γ-tocopherol methyl transferase gene in transgenic Brassica juncea plants alleviates abiotic stress: Physiological and chlorophyll a fluorescence measurements. Biochim. Biophys. Acta Bioenerg. 2010, 1797, 1428–1438. [Google Scholar] [CrossRef]
- Kalaji, H.M.; Jajoo, A.; Oukarroum, A.; Brestic, M.; Zivcak, M.; Samborska, I.A.; Ladle, R.J. Chlorophyll a fluorescence as a tool to monitor physiological status of plants under abiotic stress conditions. Acta Physiol. Plant. 2016, 38. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| S. No | Cultivars | Origin | Dormancy Rate | S. No | Cultivars | Origin | Dormancy Rate |
|---|---|---|---|---|---|---|---|
| 1 | Gibraltar | USA | 2 | 9 | WL440HQ | USA | 6 |
| 2 | Golden Queen | USA | 2.5 | 10 | WL525HQ | USA | 8 |
| 3 | SK3010 | Canada | 2.5 | 11 | Magna995 | USA | 9 |
| 4 | Bara310SC | USA | 3 | 12 | Siriver | Australia | 9 |
| 5 | WL354HQ | USA | 3.9 | 13 | WL656HQ | USA | 9.3 |
| 6 | 55V48 | USA | 5 | 14 | Nofollow | China | 9.6 |
| 7 | WL363HQ | USA | 5 | 15 | WL712 | USA | 10.2 |
| 8 | Sanditi | France | 5 |
| SFW | SDW | RFW | RDW | TFB | TDB | |||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Cultivars | CK | HT | CK | HT | CK | HT | CK | HT | CK | HT | CK | HT |
| Gibraltar | 1.87 | 1.28 * | 0.37 | 0.24 * | 0.88 | 0.66 * | 0.16 | 0.08 * | 2.75 | 1.94 * | 0.53 | 0.32 * |
| Golden queen | 1.88 | 1.02 * | 0.44 | 0.24 * | 0.92 | 0.63 * | 0.19 | 0.11 | 2.8 | 1.64 * | 0.62 | 0.35 |
| SK3010 | 2.09 | 2.07 | 0.66 | 0.54 * | 0.94 | 0.65 * | 0.18 | 0.12 * | 3.03 | 2.72 * | 0.84 | 0.77 |
| Bara310SC | 1.85 | 1.71 | 0.46 | 0.4 | 0.88 | 0.84 | 0.16 | 0.15 | 2.73 | 2.55 | 0.62 | 0.55 |
| WL354HQ | 2.39 | 1.59 * | 0.49 | 0.22 * | 0.87 | 0.69 * | 0.17 | 0.12 * | 3.26 | 2.28 * | 0.66 | 0.34 * |
| 55V48 | 2.26 | 1.49 | 0.52 | 0.35 * | 0.9 | 0.74 * | 0.18 | 0.10 * | 3.16 | 2.22 * | 0.7 | 0.45 * |
| WL363HQ | 2 | 1.56 | 0.6 | 0.5 * | 0.86 | 0.71 | 0.16 | 0.13 | 2.86 | 2.27 | 0.76 | 0.63 |
| Sanditi | 2.13 | 1.97 | 0.59 | 0.37 * | 0.94 | 0.63 * | 0.19 | 0.10 * | 3.06 | 2.59 | 0.78 | 0.47 * |
| WL440HQ | 2.01 | 1.66 | 0.54 | 0.49 | 0.91 | 0.81 * | 0.18 | 0.10 * | 2.92 | 2.47 | 0.72 | 0.59 |
| WL525HQ | 1.92 | 1.45 | 0.55 | 0.34 | 0.94 | 0.77 * | 0.18 | 0.12 * | 2.86 | 2.21 | 0.73 | 0.46 |
| Magna995 | 2.02 | 1.61 * | 0.54 | 0.44 | 0.93 | 0.91 | 0.19 | 0.18 | 2.96 | 2.52 * | 0.72 | 0.63 |
| Siriver | 1.93 | 1.58 * | 0.52 | 0.37 | 0.87 | 0.72 * | 0.14 | 0.11 | 2.8 | 2.31 * | 0.66 | 0.48 |
| WL656HQ | 2.07 | 1.69 * | 0.52 | 0.41 * | 0.91 | 0.69 | 0.15 | 0.09 * | 2.98 | 2.38 * | 0.67 | 0.5 * |
| Nofollow | 1.93 | 1.85 | 0.53 | 0.49 | 0.88 | 0.83 | 0.18 | 0.12 * | 2.81 | 2.69 * | 0.71 | 0.61 |
| WL712 | 1.82 | 1.3 * | 0.47 | 0.28 * | 0.88 | 0.75 * | 0.19 | 0.09 * | 2.7 | 2.05 * | 0.65 | 0.38 * |
| Cultivars | SFW | SDW | RFW | RDW | TFB | TDB | RWC | EL | MDA | Chla | Chlb | Tchl | SS | SP |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Gibraltar | 68.76 | 66.16 | 74.29 | 47.05 | 70.54 | 60.31 | 86.97 | 236.10 | 155.76 | 59.70 | 77.41 | 66.89 | 111.38 | 112.94 |
| Golden Queen | 61.11 | 71.02 | 68.11 | 64.66 | 63.20 | 69.12 | 90.56 | 328.84 | 134.05 | 64.94 | 75.31 | 68.95 | 101.66 | 121.71 |
| SK3010 | 76.67 | 63.64 | 68.60 | 70.24 | 74.27 | 64.99 | 91.55 | 186.63 | 176.98 | 78.96 | 82.25 | 80.25 | 105.62 | 136.49 |
| Bara310SC | 92.45 | 87.84 | 94.75 | 92.48 | 93.19 | 89.04 | 96.45 | 131.69 | 102.06 | 91.66 | 89.28 | 98.91 | 149.40 | 156.68 |
| WL354HQ | 66.53 | 44.90 | 79.13 | 70.10 | 69.88 | 51.31 | 84.71 | 208.66 | 133.16 | 74.50 | 65.56 | 70.98 | 130.37 | 122.89 |
| 55V48 | 65.71 | 68.58 | 82.10 | 54.55 | 70.36 | 64.98 | 88.57 | 299.32 | 132.07 | 76.00 | 85.49 | 79.60 | 117.63 | 133.17 |
| WL363HQ | 77.93 | 83.33 | 82.76 | 82.45 | 79.38 | 83.15 | 94.50 | 122.85 | 105.07 | 95.16 | 95.91 | 98.72 | 126.29 | 123.32 |
| Sanditi | 92.43 | 63.66 | 67.36 | 53.47 | 84.77 | 61.15 | 87.46 | 219.36 | 150.00 | 68.89 | 90.13 | 76.60 | 108.49 | 132.35 |
| WL440HQ | 82.50 | 89.59 | 89.08 | 55.13 | 84.55 | 80.96 | 87.58 | 254.19 | 107.80 | 85.60 | 68.47 | 78.13 | 102.01 | 126.59 |
| WL525HQ | 75.57 | 61.38 | 81.18 | 68.15 | 77.42 | 63.07 | 96.13 | 184.42 | 123.20 | 79.23 | 86.02 | 81.64 | 119.07 | 106.21 |
| Magna995 | 79.58 | 82.14 | 97.58 | 101.63 | 85.27 | 87.12 | 96.28 | 137.04 | 113.47 | 97.79 | 91.58 | 95.22 | 174.57 | 134.26 |
| Siriver | 81.89 | 72.23 | 83.15 | 75.71 | 82.29 | 73.00 | 89.99 | 214.81 | 166.06 | 66.59 | 98.87 | 75.87 | 112.05 | 107.56 |
| WL656HQ | 81.74 | 78.14 | 76.08 | 61.50 | 80.01 | 74.39 | 90.99 | 157.36 | 103.87 | 66.04 | 87.96 | 98.17 | 107.40 | 122.42 |
| Nofollow | 84.02 | 73.24 | 91.47 | 69.00 | 83.68 | 71.75 | 90.08 | 150.95 | 103.87 | 92.76 | 98.88 | 95.23 | 135.21 | 122.09 |
| WL712 | 71.23 | 60.85 | 84.97 | 50.25 | 75.73 | 57.81 | 86.72 | 213.66 | 128.23 | 68.72 | 55.86 | 63.43 | 110.16 | 107.78 |
| SFW | SDW | RFW | RDW | TFB | TDB | RWC | EL | MDA | CHLa | CHLb | Tchl | SS | SP | |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| SFW | 1.00 | 0.49 | 0.33 | 0.33 | 0.94 ** | 0.52 * | 0.34 | −0.61 * | −0.24 | 0.58 * | 0.34 | 0.41 | 0.26 | 0.42 |
| SDW | 1.00 | 0.51 * | 0.40 | 0.60 * | 0.94 ** | 0.53 * | −0.29 | −0.55 * | 0.70 ** | 0.72 ** | 0.73 ** | 0.26 | 0.41 | |
| RFW | 1.00 | 0.56 * | 0.62 * | 0.61 * | 0.40 | −0.48 | −0.64 * | 0.26 | 0.68 ** | 0.58 * | 0.74 ** | 0.21 | ||
| RDW | 1.00 | 0.48 | 0.68 ** | 0.77 ** | −0.64 * | −0.31 | 0.37 | 0.64 * | 0.61 * | 0.81 ** | 0.45 | |||
| TFB | 1.00 | 0.66 ** | 0.44 | −0.67 ** | −0.42 | 0.55 * | 0.51 | 0.53 * | 0.47 | 0.42 | ||||
| TDB | 1.00 | 0.70 ** | −0.46 | −0.56 * | 0.70 ** | 0.80 ** | 0.80 ** | 0.49 | 0.49 | |||||
| RWC | 1.00 | −0.57 * | −0.37 | 0.41 | 0.56 * | 0.57 * | 0.55 * | 0.32 | ||||||
| EL | 1.00 | 0.40 | −0.38 | −0.62 * | −0.58 * | −0.61 * | −0.23 | |||||||
| MDA | 1.00 | −0.38 | −0.53 * | −0.61 * | −0.44 | −0.22 | ||||||||
| CHLa | 1.00 | 0.63 * | 0.79 ** | 0.33 | 0.65 ** | |||||||||
| CHLb | 1.00 | 0.94 ** | 0.57 * | 0.42 | ||||||||||
| Tchl | 1.00 | 0.54 * | 0.59 * | |||||||||||
| SS | 1.00 | 0.43 | ||||||||||||
| SP | 1.00 |
| Cultivars | SFW | SDW | RFW | RDW | TFB | TDB | RWC | EL | MDA | Chla | Chlb | Tchl | SS | TSP | Mean | Rank |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Gibraltar | 0.24 | 0.48 | 0.23 | 0.00 | 0.24 | 0.24 | 0.19 | 0.81 | 0.28 | 0.00 | 0.50 | 0.10 | 0.13 | 0.13 | 0.26 | 14 |
| Golden Queen | 0.00 | 0.58 | 0.02 | 0.32 | 0.00 | 0.47 | 0.50 | 0.50 | 0.57 | 0.14 | 0.45 | 0.16 | 0.00 | 0.31 | 0.29 | 13 |
| SK3010 | 0.50 | 0.42 | 0.04 | 0.42 | 0.37 | 0.36 | 0.58 | 0.42 | 0.00 | 0.51 | 0.61 | 0.47 | 0.05 | 0.60 | 0.38 | 9 |
| Bara310SC | 1.00 | 0.96 | 0.91 | 0.83 | 1.00 | 1.00 | 1.00 | 0.00 | 1.00 | 0.84 | 0.78 | 1.00 | 0.65 | 1.00 | 0.86 | 1 |
| WL354HQ | 0.17 | 0.00 | 0.39 | 0.42 | 0.22 | 0.00 | 0.00 | 1.00 | 0.58 | 0.39 | 0.23 | 0.21 | 0.39 | 0.33 | 0.31 | 12 |
| 55V48 | 0.15 | 0.53 | 0.49 | 0.14 | 0.24 | 0.36 | 0.33 | 0.67 | 0.60 | 0.43 | 0.69 | 0.46 | 0.22 | 0.53 | 0.42 | 7 |
| WL363HQ | 0.54 | 0.86 | 0.51 | 0.65 | 0.54 | 0.84 | 0.83 | 0.17 | 0.96 | 0.93 | 0.93 | 0.99 | 0.34 | 0.34 | 0.67 | 3 |
| Sanditi | 1.00 | 0.42 | 0.00 | 0.12 | 0.72 | 0.26 | 0.23 | 0.77 | 0.36 | 0.24 | 0.80 | 0.37 | 0.09 | 0.52 | 0.42 | 10 |
| WL440HQ | 0.68 | 1.00 | 0.72 | 0.15 | 0.71 | 0.79 | 0.24 | 0.76 | 0.92 | 0.68 | 0.29 | 0.41 | 0.00 | 0.40 | 0.55 | 5 |
| WL525HQ | 0.46 | 0.37 | 0.46 | 0.39 | 0.47 | 0.31 | 0.97 | 0.03 | 0.72 | 0.51 | 0.70 | 0.51 | 0.24 | 0.00 | 0.44 | 11 |
| Magna995 | 0.59 | 0.83 | 1.00 | 1.00 | 0.74 | 0.95 | 0.99 | 0.01 | 0.85 | 1.00 | 0.83 | 0.90 | 1.00 | 0.56 | 0.80 | 2 |
| Siriver | 0.66 | 0.61 | 0.52 | 0.53 | 0.64 | 0.57 | 0.45 | 0.55 | 0.15 | 0.18 | 1.00 | 0.35 | 0.14 | 0.03 | 0.46 | 8 |
| WL656HQ | 0.66 | 0.74 | 0.29 | 0.26 | 0.56 | 0.61 | 0.53 | 0.47 | 0.98 | 0.17 | 0.75 | 0.98 | 0.08 | 0.32 | 0.53 | 6 |
| Nofollow | 0.73 | 0.63 | 0.80 | 0.40 | 0.68 | 0.54 | 0.46 | 0.54 | 0.98 | 0.87 | 1.00 | 0.90 | 0.46 | 0.31 | 0.67 | 4 |
| WL712 | 0.32 | 0.36 | 0.58 | 0.06 | 0.42 | 0.17 | 0.17 | 0.73 | 0.45 | 0.24 | 0.00 | 0.00 | 0.11 | 0.03 | 0.26 | 14 |
| Gibraltar | Bara310SC | Magna995 | WL712 | |||||
|---|---|---|---|---|---|---|---|---|
| CK | HT | CK | HT | CK | HT | CK | HT | |
| Data extracted from OJIP fluorescence transient curves | ||||||||
| F0 | 0.15 | 0.19 * | 0.16 | 0.18 | 0.18 | 0.19 | 0.18 | 0.22 * |
| Fj | 0.50 | 0.53 | 0.49 | 0.50 | 0.51 | 0.58 * | 0.53 | 0.60 * |
| Fi | 0.72 | 0.73 | 0.72 | 0.70 | 0.74 | 0.77 | 0.76 | 0.77 |
| Fm | 0.84 | 0.85 | 0.81 | 0.84 | 0.86 | 0.89 | 0.87 | 0.87 |
| F300μs | 0.37 | 0.43 | 0.37 | 0.36 | 0.36 | 0.43 | 0.35 | 0.50 * |
| Fv/Fm | 0.82 | 0.77 | 0.80 | 0.79 | 0.80 | 0.78 | 0.80 | 0.75 * |
| Specific energy fluxes (per active PS II reaction center) | ||||||||
| TPo/RC | 2.53 | 2.75 | 2.41 | 2.44 | 2.22 | 2.47 | 1.92 | 2.95 |
| ETo/RC | 1.25 | 1.31 | 1.27 | 1.17 | 1.14 | 1.08 | 0.95 | 1.24 |
| REo/RC | 0.42 | 0.50 | 0.43 | 0.39 | 0.40 | 0.41 | 0.32 | 0.50 |
| ABS/RC | 3.11 | 3.55 | 3.04 | 3.04 | 2.79 | 3.16 | 2.41 | 3.93 |
| Quantum yields and efficiencies | ||||||||
| ϕpo | 0.82 | 0.77 | 0.79 | 0.80 | 0.80 | 0.78 | 0.80 | 0.75 * |
| φEo | 0.41 | 0.37 | 0.41 | 0.38 | 0.41 | 0.34 * | 0.39 | 0.32 * |
| δRo | 0.34 | 0.38 | 0.34 | 0.34 | 0.35 | 0.38 | 0.34 | 0.39 |
| RC/ABS | 0.33 | 0.28 | 0.33 | 0.33 | 0.36 | 0.32 | 0.42 | 0.26 * |
| Performance indexes | ||||||||
| PIABS | 1.08 | 0.60 * | 0.90 | 0.86 | 1.01 | 0.65 * | 1.10 | 0.37 * |
| PItotal | 0.63 | 0.36 | 0.48 | 0.44 | 0.55 | 0.38 | 0.57 | 0.25 * |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wassie, M.; Zhang, W.; Zhang, Q.; Ji, K.; Chen, L. Effect of Heat Stress on Growth and Physiological Traits of Alfalfa (Medicago sativa L.) and a Comprehensive Evaluation for Heat Tolerance. Agronomy 2019, 9, 597. https://doi.org/10.3390/agronomy9100597
Wassie M, Zhang W, Zhang Q, Ji K, Chen L. Effect of Heat Stress on Growth and Physiological Traits of Alfalfa (Medicago sativa L.) and a Comprehensive Evaluation for Heat Tolerance. Agronomy. 2019; 9(10):597. https://doi.org/10.3390/agronomy9100597
Chicago/Turabian StyleWassie, Misganaw, Weihong Zhang, Qiang Zhang, Kang Ji, and Liang Chen. 2019. "Effect of Heat Stress on Growth and Physiological Traits of Alfalfa (Medicago sativa L.) and a Comprehensive Evaluation for Heat Tolerance" Agronomy 9, no. 10: 597. https://doi.org/10.3390/agronomy9100597
APA StyleWassie, M., Zhang, W., Zhang, Q., Ji, K., & Chen, L. (2019). Effect of Heat Stress on Growth and Physiological Traits of Alfalfa (Medicago sativa L.) and a Comprehensive Evaluation for Heat Tolerance. Agronomy, 9(10), 597. https://doi.org/10.3390/agronomy9100597