
Storage Temperature and Grain Moisture Effects on Phenolic Compounds as a Driver of Seed Coat Darkening in Red Lentil

 , , ,
, , ,  , , and
, , and {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Materials and Methods
2.1. Estimation of Phenolic Compounds
2.2. Statistical Analysis
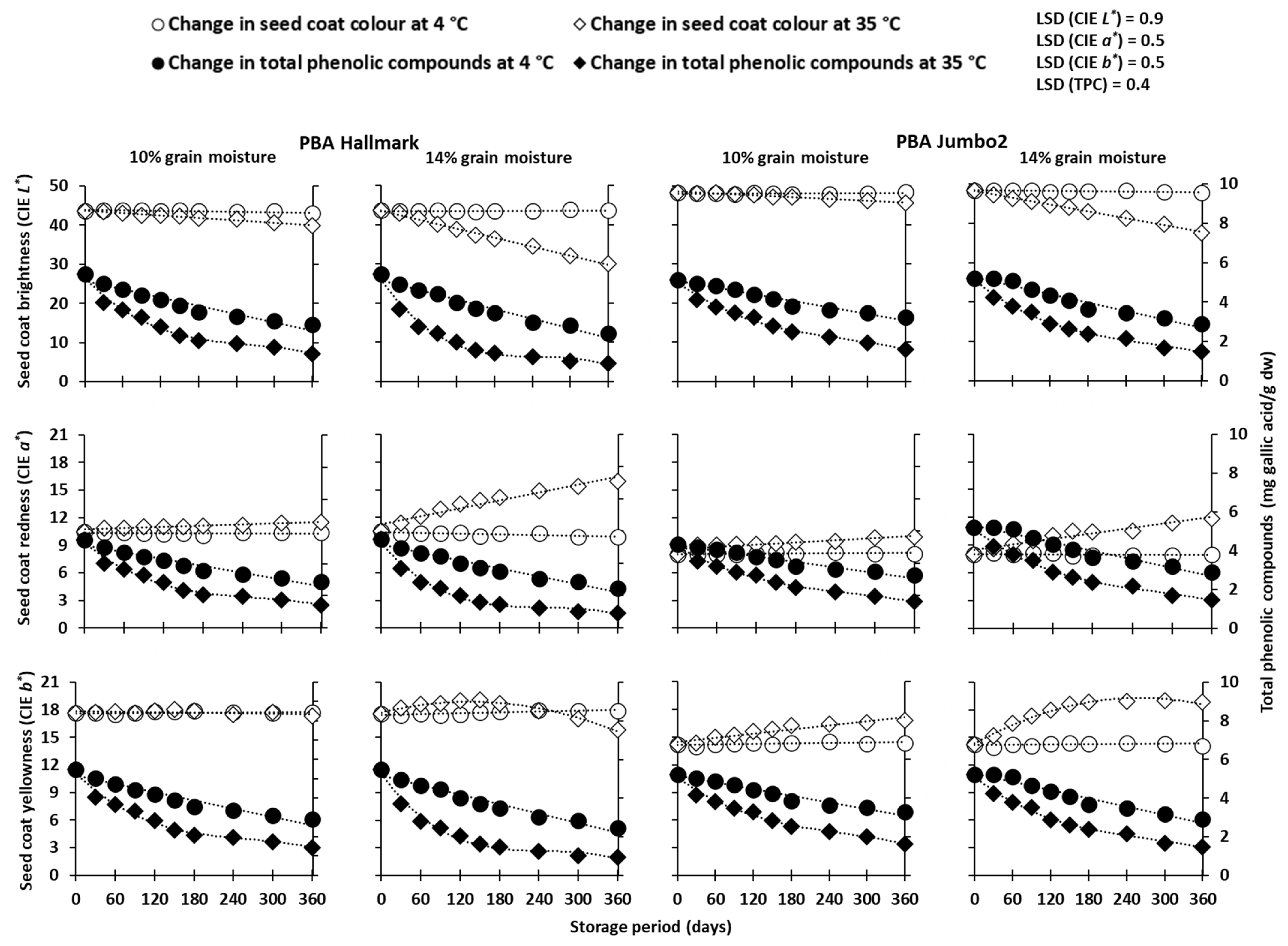
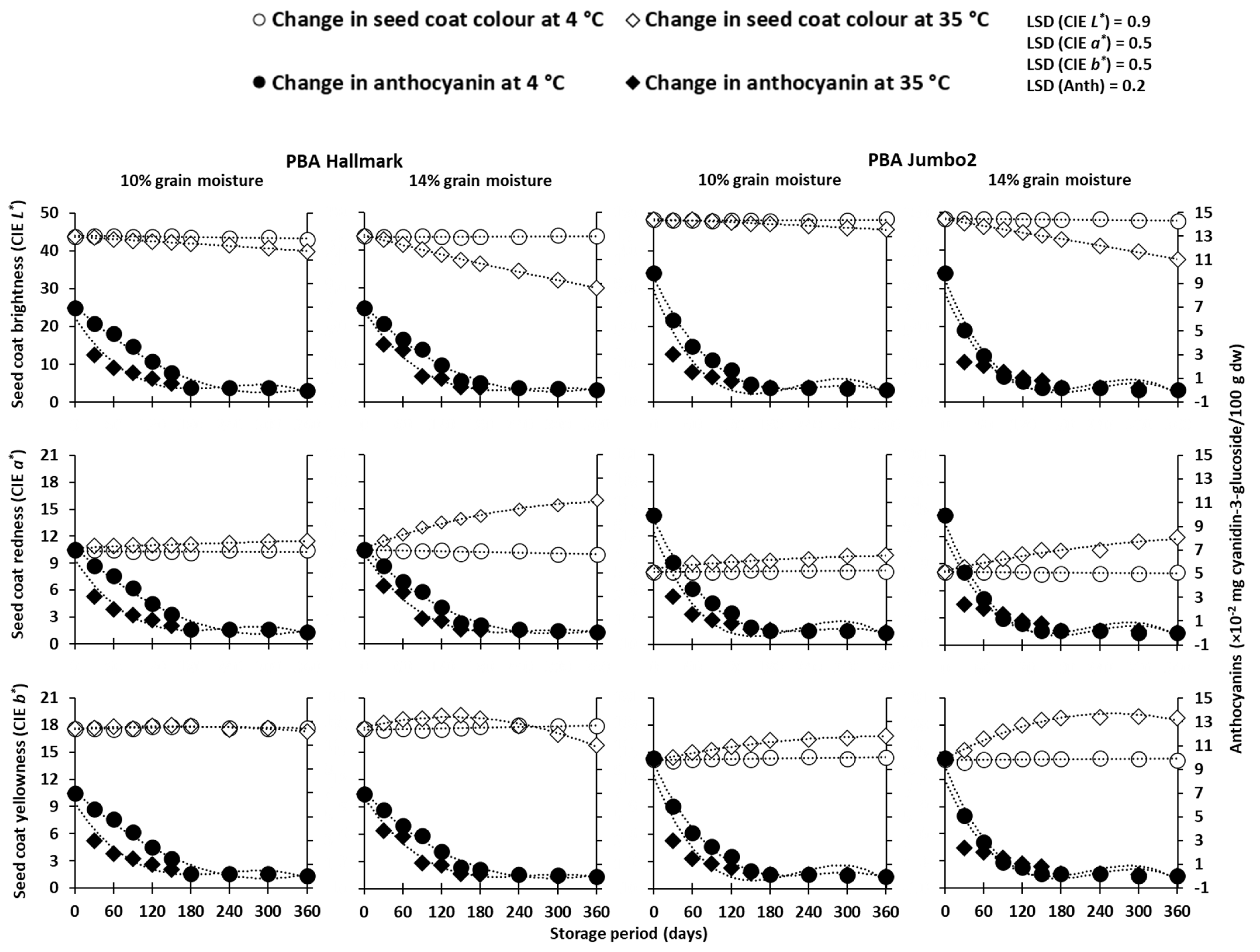
3. Results
4. Discussion
5. Conclusions
6. Future Research Direction
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Roy, S.; Islam, M.; Sarker, A.; Ismail, M.; Rafii, M.; Mondal, M.; Malek, M. Morphological characterization of lentil accessions: Qualitative characters. Bangladesh J. Bot. 2012, 41, 187–190. [Google Scholar] [CrossRef]
- Venora, G.; Grillo, O.; Shahin, M.A.; Symons, S.J. Identification of Sicilian landraces and Canadian cultivars of lentil using an image analysis system. Food Res. Int. 2007, 40, 161–166. [Google Scholar] [CrossRef]
- GTA. Australian Pulse Standards. Available online: https://www.graintrade.org.au/commodity_standards (accessed on 1 February 2024).
- CGC. Official Grain Grading Guide. Available online: https://www.grainscanada.gc.ca/en/grain-quality/official-grain-grading-guide/ (accessed on 1 February 2024).
- McDonald, L.; Panozzo, J. A review of the opportunities for spectral-based technologies in post-harvest testing of pulse grains. Legume Sci. 2022, 5, e175. [Google Scholar] [CrossRef]
- Mirali, M.; Purves, R.W.; Vandenberg, A. Phenolic profiling of green lentil (Lens culinaris Medic.) seeds subjected to long-term storage. Eur. Food Res. Technol. 2016, 242, 2161–2170. [Google Scholar] [CrossRef]
- Bhattarai, B.; Walker, C.K.; Wallace, A.J.; Nuttall, J.G.; Hepworth, G.; Panozzo, J.F.; Partington, D.L.; Fitzgerald, G.J. Storage Temperature and grain moisture effects on market and end use properties of red lentil. Agronomy 2023, 13, 2261. [Google Scholar] [CrossRef]
- Nasar-Abbas, S.M.; Siddique, K.H.M.; Plummer, J.A.; White, P.F.; Harris, D.; Dods, K.; D’Antuono, M. Faba bean (Vicia faba L.) seeds darken rapidly and phenolic content falls when stored at higher temperature, moisture and light intensity. LWT-Food Sci. Technol. 2009, 42, 1703–1711. [Google Scholar] [CrossRef]
- Aguilera, Y.; Dueñas, M.; Estrella, I.; Hernández, T.; Benitez, V.; Esteban, R.M.; Martín-Cabrejas, M.A. Evaluation of phenolic profile and antioxidant properties of pardina lentil as affected by industrial dehydration. J. Agric. Food Chem. 2010, 58, 10101–10108. [Google Scholar] [CrossRef] [PubMed]
- Bartolomé, B.; Estrella, I.; Hernández, T. Changes in phenolic compounds in lentils (Lens culinaris) during germination and fermentation. Z. Leb.-Forsch. A 1997, 205, 290–294. [Google Scholar] [CrossRef]
- Duenas, M.; Baoshan, S.; Hernandez, T.; Estrella, I.; Spranger, M.I. Proanthocyanidin composition in the seed coat of lentils (Lens culinaris L.). J. Agric. Food Chem. 2003, 51, 7999–8004. [Google Scholar] [CrossRef]
- Sant’Anna, V.; Gurak, P.D.; Ferreira Marczak, L.D.; Tessaro, I.C. Tracking bioactive compounds with colour changes in foods—A review. Dye. Pigment. 2013, 98, 601–608. [Google Scholar] [CrossRef]
- Lee, J. Degradation kinetics of grape skin and seed proanthocyanidins in a model wine system. Food Chem. 2010, 123, 51–56. [Google Scholar] [CrossRef]
- Walker, J.R.L.; Ferrar, P.H. Diphenol Oxidases, enzyme-catalysed browning and plant disease resistance. Biotechnol. Genet. Eng. Rev. 1998, 15, 457–498. [Google Scholar] [CrossRef] [PubMed]
- Amarowicz, R.; Carle, R.; Dongowski, G.; Durazzo, A.; Galensa, R.; Kammerer, D.; Maiani, G.; Piskula, M.K. Influence of postharvest processing and storage on the content of phenolic acids and flavonoids in foods. Mol. Nutr. Food Res. 2009, 53, S151–S183. [Google Scholar] [CrossRef]
- Ziegler, V.; Paraginski, R.T.; Ferreira, C.D. Grain storage systems and effects of moisture, temperature and time on grain quality—A review. J. Stored Prod. Res. 2021, 91, 101770. [Google Scholar] [CrossRef]
- Nozzolillo, C.; Bezada, M.D. Browning of lentil seeds, concomitant loss of viability, and the possible role of soluble tannins in both phenomena. Can. J. Plant Sci. 1984, 64, 815–824. [Google Scholar] [CrossRef]
- Caldas, G.V.; Blair, M.W. Inheritance of seed condensed tannins and their relationship with seed-coat color and pattern genes in common bean (Phaseolus vulgaris L.). Theor. Appl. Genet. 2009, 119, 131–142. [Google Scholar] [CrossRef]
- Lindemann, I.d.S.; Lang, G.H.; Ferreira, C.D.; Colussi, R.; Elias, M.C.; Vanier, N.L. Cowpea storage under nitrogen-modified atmosphere at different temperatures: Impact on grain structure, cooking quality, in vitro starch digestibility, and phenolic extractability. J. Food Process. Preserv. 2020, 44, e14368. [Google Scholar] [CrossRef]
- Beninger, C.W.; Gu, L.; Prior, R.L.; Junk, D.C.; Vandenberg, A.; Bett, K.E. Changes in polyphenols of the seed coat during the after-darkening process in pinto beans (Phaseolus vulgaris L.). J. Agric. Food Chem. 2005, 53, 7777–7782. [Google Scholar] [CrossRef] [PubMed]
- Ainsworth, E.A.; Gillespie, K.M. Estimation of total phenolic content and other oxidation substrates in plant tissues using Folin-Ciocalteu reagent. Nat. Protoc. 2007, 2, 875–877. [Google Scholar] [CrossRef]
- FAO/IAEA. Quantification of Tannins in Tree Foliage. A Laboratory Manual for the FAO/IAEA Co-Ordinated Research Project on Use of Nuclear and Related Techniques to Develop Simple Tannin Assays for Predicting and Improving the Safety and Efficiency of Feeding Ruminants on Tanniniferous Tree Foliage. 2000. FAO/IAEA Working Document IAEA, VIENNA. 2000. Available online: https://www.iaea.org/sites/default/files/21/06/nafa-aph-manual-pubd31022manual-tannin.pdf (accessed on 15 August 2023).
- Sintara, M.; Li, L.; Cunningham, D.G.; Prior, R.L.; Wu, X.; Chang, T. Single-laboratory validation for determination of total soluble proanthocyanidins in cranberry using 4-dimethylaminocinnamaldehyde. J. AOAC Int. 2019, 101, 805–809. [Google Scholar] [CrossRef]
- Zhao, X.; Corrales, M.; Zhang, C.; Hu, X.; Ma, Y.; Tauscher, B. Composition and thermal stability of anthocyanins from chinese purple corn (Zea mays L.). J. Agric. Food Chem. 2008, 56, 10761–10766. [Google Scholar] [CrossRef] [PubMed]
- Payne, R.; Murray, D.; Harding, S.; Baird, D.; Soutar, D. GenStat for Windows Introduction; VSN International: Hemel Hempstead, UK, 2009; Volume 204. [Google Scholar]
- Edmister, J.A.; Breene, W.M.; Serugendo, A. Influence of temperature, water activity and time on cookability and color of a stored Rwandan dry bean (Phaseolus vulgaris) mixture. J. Stored Prod. Res. 1990, 26, 121–126. [Google Scholar] [CrossRef]
- Taylor, A.; Min, T.; Paine, D. Maillard reactions cause browning in bean seed coats during ageing: Inhibition by aminoguanidine. In Seed Biology: Advances and Applications; Black, M., Bradford, K.J., Vazquez-Ramos, J., Eds.; CABI: Wallingford, UK, 2000; pp. 189–195. [Google Scholar]
- Duwadi, K.; Austin, R.S.; Mainali, H.R.; Bett, K.; Marsolais, F.; Dhaubhadel, S. Slow darkening of pinto bean seed coat is associated with significant metabolite and transcript differences related to proanthocyanidin biosynthesis. BMC Genom. 2018, 19, 260. [Google Scholar] [CrossRef] [PubMed]
- Freixas Coutin, J.A.; Munholland, S.; Silva, A.; Subedi, S.; Lukens, L.; Crosby, W.L.; Pauls, K.P.; Bozzo, G.G. Proanthocyanidin accumulation and transcriptional responses in the seed coat of cranberry beans (Phaseolus vulgaris L.) with different susceptibility to postharvest darkening. BMC Plant Biol. 2017, 17, 89. [Google Scholar] [CrossRef] [PubMed]
- Huang, P.-H.; Cheng, Y.-T.; Chan, Y.-J.; Lu, W.-C.; Li, P.-H. Effect of heat treatment on nutritional and chromatic properties of mung bean (Vigna radiata L.). Agronomy 2022, 12, 1365. [Google Scholar] [CrossRef]
- Prescott, C.E. Litter decomposition: What controls it and how can we alter it to sequester more carbon in forest soils? Biogeochemistry 2010, 101, 133–149. [Google Scholar] [CrossRef]
- Bento, J.A.C.; Ribeiro, P.R.V.; Bassinello, P.Z.; Brito, E.S.d.; Zocollo, G.J.; Caliari, M.; Soares Júnior, M.S. Phenolic and saponin profile in grains of carioca beans during storage. LWT 2021, 139, 110599. [Google Scholar] [CrossRef]



Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bhattarai, B.; Nuttall, J.G.; Li, M.; Suleria, H.A.R.; Wallace, A.J.; Fitzgerald, G.J.; Walker, C.K. Storage Temperature and Grain Moisture Effects on Phenolic Compounds as a Driver of Seed Coat Darkening in Red Lentil. Agronomy 2024, 14, 705. https://doi.org/10.3390/agronomy14040705
Bhattarai B, Nuttall JG, Li M, Suleria HAR, Wallace AJ, Fitzgerald GJ, Walker CK. Storage Temperature and Grain Moisture Effects on Phenolic Compounds as a Driver of Seed Coat Darkening in Red Lentil. Agronomy. 2024; 14(4):705. https://doi.org/10.3390/agronomy14040705
Chicago/Turabian StyleBhattarai, Bhawana, James G. Nuttall, Minhao Li, Hafiz A. R. Suleria, Ashley J. Wallace, Glenn J. Fitzgerald, and Cassandra K. Walker. 2024. "Storage Temperature and Grain Moisture Effects on Phenolic Compounds as a Driver of Seed Coat Darkening in Red Lentil" Agronomy 14, no. 4: 705. https://doi.org/10.3390/agronomy14040705
APA StyleBhattarai, B., Nuttall, J. G., Li, M., Suleria, H. A. R., Wallace, A. J., Fitzgerald, G. J., & Walker, C. K. (2024). Storage Temperature and Grain Moisture Effects on Phenolic Compounds as a Driver of Seed Coat Darkening in Red Lentil. Agronomy, 14(4), 705. https://doi.org/10.3390/agronomy14040705