
Ethyl Methanesulfonate Mutant Library Construction in Tartary Buckwheat with Agronomic Trait and Flavonoid Screening for Germplasm Innovation

Abstract
1. Introduction
2. Materials and Methods
2.1. EMS Mutation Library Construction
2.2. Planting and Phenotypic Characteristic Screening of Mutagenic Materials
2.3. Grain Morphology and Data Measurement
2.4. Flavonoid Content Determination via Near-Infrared Reflectance Spectroscopy (NIR)
NIR Model Establishment
3. Results
3.1. Construction of the Mutant Library
3.2. Screening of Mutant Phenotypes
3.2.1. Plant Type and Leaf Colour Variation
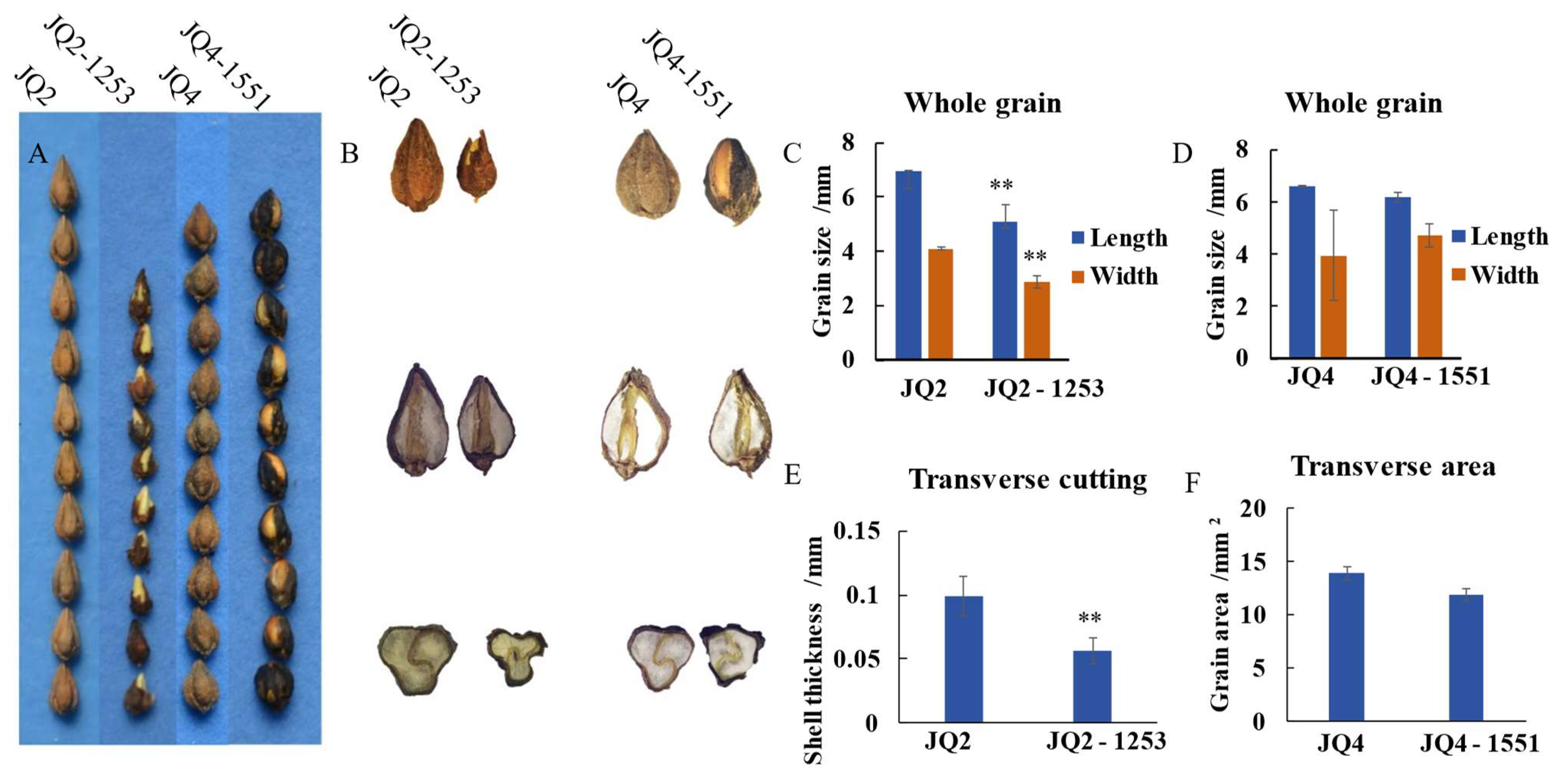
3.2.2. Grain Variation
3.2.3. Analysis of the 1000-Grain Weights (TGWs) of the Mutants
3.2.4. Mutant Analysis of Grain Weight per Plant (GWP)
3.2.5. Correlation Analysis of Mutant Grain Traits
3.3. Analysis of Differences in Flavonoid Content in the Mutants
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Joshi, D.C.; Chaudhari, G.V.; Sood, S.; Kant, L.; Pattanayak, A.; Zhang, K.; Fan, Y.; Janovská, D.; Meglič, V.; Zhou, M. Revisiting the versatile buckwheat: Reinvigorating genetic gains through integrated breeding and genomics approach. Planta 2019, 250, 783–801. [Google Scholar] [CrossRef]
- Joshi, D.C.; Zhang, K.; Wang, C.; Chandora, R.; Khurshid, M.; Li, J.; He, M.; Georgiev, M.I.; Zhou, M. Strategic enhancement of genetic gain for nutraceutical development in buckwheat: A genomics-driven perspective. Biotechnol. Adv. 2020, 39, 107479. [Google Scholar] [CrossRef]
- Shukla, A.; Srivastava, N.; Suneja, P.; Yadav, S.; Hussain, Z.; Rana, J.C.; Yadav, S. Genetic diversity analysis in buckwheat germplasm for nutritional traits. Indian J. Exp. Biol. 2018, 56, 827–837. [Google Scholar]
- Kwiecień, S.; Brzozowski, T.; Konturek, S.J. Effects of reactive oxygen species action on gastric mucosa in various models of mucosal injury. J. Physiol. Pharmacol. 2002, 53, 39–50. [Google Scholar]
- Synnerstad, I.; Johansson, M.; Nylander, O.; Holm, L. Intraluminal acid and gastric mucosal integrity: The importance of blood-borne bicarbonate. Am. J. Physiol. Gastrointest. Liver Physiol. 2001, 280, G121–G129. [Google Scholar] [CrossRef] [PubMed]
- Østergaard, L.; Yanofsky, M.F. Establishing gene function by mutagenesis in Arabidopsis thaliana. Plant J. 2004, 39, 682–696. [Google Scholar] [CrossRef]
- Wang, L.; Zheng, J.; Luo, Y.; Xu, T.; Zhang, Q.; Zhang, L.; Xu, M.; Wan, J.; Wang, M.B.; Zhang, C.; et al. Construction of a genomewide RNAi mutant library in rice. Plant Biotechnol. J. 2013, 11, 997–1005. [Google Scholar] [CrossRef] [PubMed]
- Nie, S.; Wang, B.; Ding, H.; Lin, H.; Zhang, L.; Li, Q.; Wang, Y.; Zhang, B.; Liang, A.; Zheng, Q.; et al. Genome assembly of the Chinese maize elite inbred line RP125 and its EMS mutant collection provide new resources for maize genetics research and crop improvement. Plant J. 2021, 108, 40–54. [Google Scholar] [CrossRef]
- Sun, J.; Luu, N.S.; Chen, Z.; Chen, B.; Cui, X.; Wu, J.; Zhang, Z.; Lu, T. Generation and Characterization of a Foxtail Millet (Setaria italica) Mutant Library. Front. Plant Sci. 2019, 10, 369. [Google Scholar] [CrossRef]
- Wang, D.; Li, Y.; Wang, H.; Xu, Y.; Yang, Y.; Zhou, Y.; Chen, Z.; Zhou, Y.; Gui, L.; Guo, Y.; et al. Boosting wheat functional genomics via an indexed EMS mutant library of KN9204. Plant Commun. 2023, 4, 100593. [Google Scholar] [CrossRef]
- Kodym, A.; Afza, R. Physical and chemical mutagenesis. Methods Mol. Biol. 2003, 236, 189–204. [Google Scholar]
- Koebner, R.; Hadfield, J. Large-scale mutagenesis directed at specific chromosomes in wheat. Genome 2001, 44, 45–49. [Google Scholar] [CrossRef]
- Kinane, J.T.; Jones, P.W. Isolation of wheat mutants with increased resistance to powdery mildew from small induced variant populations. Euphytica 2001, 117, 251–260. [Google Scholar] [CrossRef]
- An, C.; Beard, W.A.; Chen, D.; Wilson, S.H.; Makridakis, N.M. Understanding the loss-of-function in a triple missense mutant of DNA polymerase β found in prostate cancer. Int. J. Oncol. 2013, 43, 1131–1140. [Google Scholar] [CrossRef][Green Version]
- Wang, F.; Travins, J.; DeLaBarre, B.; Penard-Lacronique, V.; Schalm, S.; Hansen, E.; Straley, K.; Kernytsky, A.; Liu, W.; Gliser, C.; et al. Targeted inhibition of mutant IDH2 in leukemia cells induces cellular differentiation. Science 2013, 340, 622–626. [Google Scholar] [CrossRef]
- Qu, G.; Sun, Y.; Pang, H.; Wu, Q.; Wang, F.; Hu, S. EMS mutagenesis and ALS—Inhibitor herbicide—Resistant mutants of Brassica napus L. Chin. J. Oil Crop Sci. 2014, 36, 25–31. [Google Scholar]
- Gautam, V.; Swaminathan, M.; Akilan, M.; Gurusamy, A.; Suresh, M.; Kaithamalai, B.; Joel, A.J. Early flowering, good grain quality mutants through gamma rays and EMS for enhancing per day productivity in rice (Oryza sativa L.). Int. J. Radiat. Biol. 2021, 97, 1716–1730. [Google Scholar] [CrossRef] [PubMed]
- Jia, Z.; Cui, Y.; Li, Y.; Wang, X.; Du, Y.; Huang, S. Inducible positive mutant screening system to unveil the signaling pathway of late blight resistance. J. Integr. Plant Biol. 2010, 52, 476–484. [Google Scholar] [CrossRef] [PubMed]
- Ma, M.; Liu, L.; Zhang, L.; Zhou, J.; Cui, L. Research Progress of Buckwheat Breeding. J. Shanxi Agric. Sci. 2015, 43, 240–243. [Google Scholar]
- Li, G.; Shen, H.; Fan, L.; Cao, H. Heredity analysis of flavone and agronomic characters among radiated tartary buckwheat mutants. In Proceedings of the 10th International Symposium on Buckwheat, Xianyang, China, 14–18 August 2007; pp. 178–185. [Google Scholar]
- Zhao, G.; Tang, Y.; Wang, A. Selection of a New Tartary Buckwheat Variety ‘Xiqiao No.1’. Hortic. Seed 2002, 262–264. [Google Scholar]
- Zhang, C.; Li, X.; Zhang, Y. Selection of Jin Buckwheat (Sweet) 3 and Its Cultivation Techniques. J. Shanxi Agric. Sci. 2011, 39, 316–318. [Google Scholar]
- Ma, M.; Liu, L.; Zhang, L.; Cui, L.; Zhou, J. Morphological Identification and Analysis of EMS-Induced Mutants from Ciqiao. Crops 2019, 37–41+197–198. [Google Scholar] [CrossRef]
- Shimizu, A.; Yamaguchi, H.; Degi, K.; Morishita, T. Development of ‘Darumadattan’, a semidwarf lodging-resistant Tartary buckwheat cultivar, using gamma-ray irradiation. Breed. Sci. 2020, 70, 623–630. [Google Scholar] [CrossRef]
- Sun, Z.; Hou, S.; Hao, Y.; Wang, D.; Gao, H.; Han, Y.; Li, H.; Liu, L.; Zhou, M. EMS-assisted Mutagenesis of Tartary Buckwheat and Expression Analysis of Rutin Biosynthesis Genes in Selected Mutants. J. Plant Genet. Resour. 2020, 21, 402–408. [Google Scholar]
- Yao, Y.; Cheng, X.-Z.; Ren, G.-X. Application of Near-Infrared Reflectance Spectroscopy to the Evaluation of D-chiro-lnositol, Vitexin, and Isovitexin Contents in Mung Bean. Agric. Sci. China 2011, 10, 1986–1991. [Google Scholar] [CrossRef]
- Li, R.; Chen, Z.; Zheng, R.; Chen, Q.; Deng, J.; Li, H.; Huang, J.; Liang, C.; Shi, T. QTL mapping and candidate gene analysis for yield and grain weight/size in Tartary buckwheat. BMC Plant Biol. 2023, 23, 58. [Google Scholar] [CrossRef] [PubMed]
- Pan, F.; Shi, T.; Chen, Q.; Meng, Z.; Liang, C.; Chen, Q. Variation in Major Agronomic Traits and its Contribution to Grain Weight per Plant in Tartary Buckwheat Germplasm. Plant Sci. J. 2015, 33, 829–839. [Google Scholar]
- Zhou, Q.; He, P.; Tang, J.; Huang, K.; Huang, X. Increasing planting density can improve the yield of Tartary buckwheat. Front. Plant Sci. 2023, 14, 1313181. [Google Scholar] [CrossRef] [PubMed]
- Qin, X.; Feng, G.; Liu, D.; Liu, C.; Yang, X.; Yan, Z. Mutagenesis of Kidney Bean Cultivar A18-1 by EMS and Mutant Library Construction. Chin. Agric. Sci. Bull. 2020, 36, 14–21. [Google Scholar]
- Ahmad, J.; Jan, S.; Javid, W.; Bhat, S.A.; Tahir, I. Assessment on induced genetic variability and divergence in the mutagenised Tartary buckwheat populations developed using gamma rays and EMS mutagenesis. Ecol. Genet. Genom. 2023, 28, 100177. [Google Scholar] [CrossRef]
- Kalinova, J.; Vrchotova, N. Level of catechin, myricetin, quercetin and isoquercitrin in buckwheat (Fagopyrum esculentum Moench), changes of their levels during vegetation and their effect on the growth of selected weeds. J. Agric. Food Chem. 2009, 57, 2719–2725. [Google Scholar] [CrossRef]
- Chen, Q.F.; Huang, X.Y.; Li, H.Y.; Yang, L.J.; Cui, Y.S. Recent Progress in Perennial Buckwheat Development. Sustainability 2018, 10, 536. [Google Scholar] [CrossRef]
- Bai, C.Z.; Feng, M.L.; Hao, X.L.; Zhong, Q.M.; Tong, L.G.; Wang, Z.H. Rutin, quercetin, and free amino acid analysis in buckwheat (Fagopyrum) seeds from different locations. Genet. Mol. Res. 2015, 14, 19040–19048. [Google Scholar] [CrossRef]
- Wang, L.; Wei, M.; Shao, J.; Du, L.; Lin, X.; Wang, L. Content determination and analysis of flavonoids in buckwheat. J. Food Saf. Food Qual. 2018, 9, 5387–5392. [Google Scholar]
- Qin, P.; Wang, Q.; Shan, F.; Hou, Z.; Ren, G. Nutritional composition and flavonoids content of flour from different buckwheat cultivars. Int. J. Food Sci. Technol. 2010, 45, 951–958. [Google Scholar] [CrossRef]
- Chen, Y.; Qin, L.; Wen, A.; Mazhar, M.; Wang, H.; Zhu, Y. Three-solvent extracting method comprehensively evaluates phenolics profile and antioxidant activities of Tartary buckwheat. J. Food Process. Preserv. 2021, 45, e15020. [Google Scholar] [CrossRef]
- Gurdon, C.; Poulev, A.; Armas, I.; Satorov, S.; Tsai, M.; Raskin, I. Genetic and Phytochemical Characterization of Lettuce Flavonoid Biosynthesis Mutants. Sci. Rep. 2019, 9, 3305. [Google Scholar] [CrossRef]
- Attarzadeh, M.; Balouchi, H.; Rajaie, M.; Dehnavi, M.M.; Salehi, A. Improving growth and phenolic compounds of Echinacea purpurea root by integrating biological and chemical resources of phosphorus under water deficit stress. Ind. Crops Prod. 2020, 154, 112763. [Google Scholar] [CrossRef]
- Maddox, C.E.; Laur, L.M.; Tian, L. Antibacterial activity of phenolic compounds against the phytopathogen Xylella fastidiosa. Curr. Microbiol. 2010, 60, 53–58. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Year | Variety | Number of Etiolated Seedlings | Number of Non-Etiolated Seedlings | Number of Seedling Emergence | Seeding Number | Etiolation Rate (%) | Germination Rate (%) |
|---|---|---|---|---|---|---|---|
| 2020 | M0-JQ2 | 257 | 12,677 | 12,924 | 25,000 | 2.03 | 51.69 |
| M0-JQ4 | 200 | 14,078 | 14,278 | 25,000 | 1.42 | 57.11 | |
| 2021 | M1-JQ2 | 93 | 13,371 | 13,464 | 49,497 | 0.70 | 27.3 |
| M1-JQ4 | 11 | 10,777 | 10,788 | 33,845 | 0.10 | 31.9 | |
| 2022 | M2-JQ2 | 53 | 7613 | 7666 | 0.70 | ||
| M2-JQ4 | 47 | 3137 | 3184 | 1.50 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Guo, H.; Qin, Z.; Ren, W.; Feng, H.; Chen, W.; Liu, L.; Sun, Z. Ethyl Methanesulfonate Mutant Library Construction in Tartary Buckwheat with Agronomic Trait and Flavonoid Screening for Germplasm Innovation. Agronomy 2024, 14, 547. https://doi.org/10.3390/agronomy14030547
Guo H, Qin Z, Ren W, Feng H, Chen W, Liu L, Sun Z. Ethyl Methanesulfonate Mutant Library Construction in Tartary Buckwheat with Agronomic Trait and Flavonoid Screening for Germplasm Innovation. Agronomy. 2024; 14(3):547. https://doi.org/10.3390/agronomy14030547
Chicago/Turabian StyleGuo, Huajie, Zhiying Qin, Wei Ren, Hongmei Feng, Wenliang Chen, Longlong Liu, and Zhaoxia Sun. 2024. "Ethyl Methanesulfonate Mutant Library Construction in Tartary Buckwheat with Agronomic Trait and Flavonoid Screening for Germplasm Innovation" Agronomy 14, no. 3: 547. https://doi.org/10.3390/agronomy14030547
APA StyleGuo, H., Qin, Z., Ren, W., Feng, H., Chen, W., Liu, L., & Sun, Z. (2024). Ethyl Methanesulfonate Mutant Library Construction in Tartary Buckwheat with Agronomic Trait and Flavonoid Screening for Germplasm Innovation. Agronomy, 14(3), 547. https://doi.org/10.3390/agronomy14030547