Abstract
Nitric oxide (NO) plays an important role in plant stress responses. However, the mechanisms underlying NO-induced stress resistance to cadmium (Cd) stress in rice remain elusive. In this study, rat neuron NO synthase (nNOS)-overexpressing rice plants with higher endogenous NO level showed higher cadmium stress tolerance than the wild-type plants. The results showed that nNOS-overexpressing rice plants accumulated less cadmium in the roots and shoots by downregulating the expression of Cd uptake and transport related genes including OsCAL1, OsIRT2, OsNramp5, and OsCd1. Moreover, nNOS-overexpressing rice plants accumulated less hydrogen peroxide (H2O2), accompanying with higher expression of antioxidant enzyme genes (OsCATA, OsCATB, and OsPOX1) and corresponding higher enzyme activities under cadmium stress. Furthermore, the transcription of melatonin biosynthetic genes, including OsASMT1, OsTDC1, OsTDC3, and OsSNAT2, was also upregulated in nNOS-overexpressing plants, resulting in increased content of melatonin under cadmium treatment compared with the wild-type controls. Taken together, this study indicates that nNOS overexpression improves Cd tolerance of rice seedlings through decreasing cadmium accumulation and enhancing the antioxidant capacity and melatonin biosynthesis of the plants.
1. Introduction
Rice is one of the most important crops in Asia. However, rice production safety is threatened by the toxic heavy metal cadmium (Cd), due to the increasing problem of Cd pollution. When rice plants are grown in Cd-polluted soil, they can absorb Cd through their roots, which is then transported to the shoots and grains. The excessive accumulation of Cd in soil not only inhibits rice growth, it also endangers human health through the food chain [1]. In order to resist Cd toxicity, rice has evolved many resistance strategies. On the one hand, rice reduces Cd accumulation by affecting Cd uptake, transport and chelation. In recent years, many Cd uptake- and transport-related genes, including OsCAL1 (cadmium accumulation in leaf 1), OsIRT2 (iron-regulated metal transporter2), OsNramp5 (natural resistance-associated macrophage protein 5), and OsCd1 (cadmium transporter gene 1), have been identified [2]. On the other hand, rice can reduce cell damage caused by Cd-induced overexpressed reactive oxygen species (ROS) by activating antioxidant enzymes such as superoxide dismutase (SOD), catalase (CAT) and peroxidase (POX) [1]. Thus, improving the cadmium tolerance and decreasing the cadmium accumulation in rice through biotechnology has become an urgent task given the increasing problems related to cadmium pollution.
It has been reported that many signaling molecules in plants are involved in the Cd stress response. Nitric oxide is a signaling molecule that is involved in plant growth and development processes, as well as in responses to environmental stresses [3,4,5]. Recently, researchers have increasingly been reporting that NO modulates the resistance of plants to Cd stress by affecting physiological metabolic processes such as reactive oxygen species (ROS), photosynthesis, chlorophyll synthesis, and cadmium uptake [6,7,8], but the role of NO in response to Cd stress is still disputed. The Cd-stress-induced change in NO levels in many plants is influenced by Cd concentration, treatment time, and plant species [9,10,11]. For example, the NO level was substantially increased in the roots of barley and rice under Cd treatment [12,13], while Cd stress inhibited endogenous NO generation in peas and rice [14,15]. Additionally, NO may play different roles in the same biological process in different plants. For instance, inhibition of NO accumulation by means of NO scavenger 2-[4-carboxyphenyl]-4,4,5,5-tetramethylimidazoline-1-oxy-3-oxide (c-PTIO) or NOS inhibitor Nω-nitro-L-arginine-methylester(L-NAME) was shown to result in the prevention of Cd-stress-induced oxidant damage in Arabidopsis and yellow lupine [16,17], but application of the NO donor sodium nitroprusside (SNP) decreased ROS accumulation in Cd-stressed Brassica juncca and rice seedlings [18,19]. In addition to the above-mentioned NO-mediated Cd tolerance in plants, NO also serves as a gas messenger, and is involved in signaling transduction by regulating relevant gene expression and S-nitrosylation modifications of target proteins [20,21].
Moreover, NO can also regulate hormone homeostasis such as indole-3-acetic acid (IAA) or melatonin (N-acetyl-5-methoxytryptamine), so as to alleviate Cd toxicity in plants [22,23]. In recent decades, an increasing number of studies have reported that Cd stress can induce melatonin accumulation, and exogenous melatonin can improve Cd tolerance in different plants [24]. Melatonin is synthesized via four continual enzymatic reactions from tryptophan, requiring at least six enzymes: tryptophan decarboxylase (TDC), tryptophan hydroxylase (TPH), tryptamine 5-hydroxylase (T5H), N-acetylserotonin methyltransferase (ASMT), and serotonin N-acetyltransferase (SNAT) [24]. Many results have been reported showing that exogenous melatonin can alleviate Cd-induced oxidative damage by activating antioxidant systems [25,26], and can decrease Cd accumulation by regulating the transcription of iron-transport genes [25,27]. Although melatonin and NO play similar roles in response to Cd stress in plants, the relationship between them is still unclear.
Knowledge about the roles of NO in plants has largely been obtained through the exogenous application of NO donors such as SNP, the NO scavenger c-PTIO, and the NOS inhibitor L-NAME, which might be affected by differences in the concentration and time point of application of chemical treatments [3]. In addition, whether the application of NO donors or scavengers reflects the physiological status of NO is still unclear [28,29]. Therefore, plant materials with different endogenous NO contents should be used to assess the function of NO in response to Cd stress. Although many reports indicate that higher plants possess arginine-dependent NO synthase (NOS) activity, no NOS coding gene has yet been found in higher plants. Lin et al. (2012) reported a rice mutant Osnoe1 that showed increased NO levels. OsNOE1 encodes a rice catalase OsCATC, but whether its role arises from H2O2 or NO accumulation—or their crosstalk—remains elusive [30]. Therefore, it may be more accurate to investigating in the vivo roles of NO in plants through specific modulation of endogenous NO levels with no significant effect on plant development.
Overexpression of rat nNOS increases both NOS activity and NO content in transgenic Arabidopsis, tobacco, and rice plants, thus increasing tolerance to stresses from drought, salt and pathogens [31,32,33]. To assess the role of endogenous NO in response to Cd stress, nNOS-overexpressing rice plants are used in the current study. Our results indicate that the nNOS overexpression in rice plants improves Cd tolerance and decreases cadmium accumulation. Furthermore, the transgenic rice plants show enhanced antioxidant capacity, higher melatonin content, and changed expression of related genes under Cd stress.
2. Materials and Methods
2.1. Plant Materials and Growth Conditions
Rice (Oryza sativa L. cv. Zhonghua11) was used both as the wild type and for the generation of nNOS transgenic plants. Rice seeds were sterilized with 70% (v/v) ethanol for 5 min and subsequently with 5% (w/v) NaClO for 30 min, washed at least three times with sterile water, and then plated on agar medium containing 1/2 MS medium in plant growth chambers (50% humidity, 200 μmol m–2s–1, 14 h light/10 h dark cycle, and 28–30 °C).
2.2. Stress Treatments and Plant Sampling
To determine a suitable Cd treatment concentration, we transferred 2-day-old wild-type (WT) rice seedlings germinated on half-strength Murashige and Skoog (1/2 MS) plates to new plates containing 0, 50, 100, or 200 μM CdCl2, and the seedlings were photographed and root and shoot lengths were measured using Image J software (Version 8.0) at 1, 2, 3, 4, and 5 days after transfer.
To test the effects of exogenous SNP treatment on the root and shoot growth under normal and cadmium-stress conditions, we transferred germinated rice seedlings to 1/2 MS medium containing 0, 20, 50, or 100 μM SNP with or without 200 μM CdCl2, and the seedlings were photographed and root and shoot lengths measured at 1, 2, 3, 4, and 5 days after transfer.
To evaluate the plants’ tolerance to cadmium stress, rice seeds were plated on agar medium containing 1/2 MS medium for 2 days in a plant growth chamber. Uniformly germinated rice seedlings were then transferred to 1/2 MS medium supplemented with 200 μM CdCl2. After 1, 2, 3, 4, and 5 d of growth, the seedlings were photographed and root and shoot lengths were measured. At least 24 seedlings were analyzed per treatment.
To measure the transcript levels of selected genes and physiological parameters including H2O2 content, CAT and POX activity, chlorophyll content, cadmium content, and melatonin content under Cd stress, the roots, shoots, or whole seedlings of the tested plants under the different treatment conditions were sampled at the designated time for further analysis.
2.3. Measurement of NO Content
One-week-old wild-type rice seedlings were treated in 200 μM CdCl2, and the NO content in the roots was assayed using the specific fluorescent probe DAF-FM DA at 24 h post treatment [33]. For DAF-FM DA imaging, the primary roots of the seedlings were incubated in 2 mL EP tubes with 10 μM DAF-FM DA in 20 mM HEPES-NaOH, pH 7.5, for 1 h, and rinsed three times with sterile water. Then, the samples were examined under an Olympus BX60 (Olympus, Tokyo, Japan) differential interference contrast microscope equipped with a CCD Olympus dp72 camera (Olympus, Tokyo, Japan) with an excitation of 488 nm and an emission of 515 nm. At least 24 seedlings per treatment were analyzed.
2.4. Measurement of NOS Activity
NOS activity was measured as previously described [33]. Briefly, approximately 0.5 g of rice seedlings was frozen and ground with liquid nitrogen, and then extracted with 2 mL buffer (50 mM Tris-HCl, pH 7.4, 1 mM EDTA, 1 mM dithiothreitol, 1 mM leupeptin, 1 mM pepstatin, and 1 mM phenylmethylsulfonyl fluoride). After centrifuging at 12,000× g for 15 min at 4 °C, the supernatant was used as the enzyme extract. NOS activity was assayed using the NOS assay kit (Beyotime, Shanghai, China), following the manufacturer’s instructions.
2.5. Measurement of H2O2 Content, CAT Activity and POX Activity
The rice seedlings treated with or without 200 μM CdCl2 for 5 days were used for the measurement of H2O2 content, CAT activity and POX activity. H2O2 content was measured using the Hydrogen Peroxide Assay Kit (Beyotime, Shanghai, China), following the manufacturer’s instructions.
To measure the activities of CAT and POX, the total protein from seedlings was extracted using 0.05 M potassium phosphate buffer (pH 7.0), and the extract was centrifuged at 12,000× g for 15 min at 4 °C. Subsequently, the supernatant was used as the enzyme extract. CAT activity was detected using a Catalase Assay Kit (Beyotime), in accordance with the manufacturer’s instructions. POX activity was measured as described previously [33]. The reaction mixture contained 0.1 mL of enzyme extract, 0.029 M potassium phosphate buffer (pH 5.5), 0.1% (v/v) H2O2 and 0.01 M guaiacol as substrates. The oxidation of guaiacol was monitored by the absorbance measured at 470 nm every 10 s.
2.6. Quantitative Real-Time PCR
Rice seedlings treated with or without 200 μM CdCl2 for 5 days were sampled for qRT-PCR as previously described [33]. The total RNA was extracted from rice leaves using TRIzol reagent (Invitrogen, CA, USA). We used 1 μg of total RNA treated with RQ1 RNase-free DNase (Promega, Beijing, China) for cDNA synthesis with an RT kit (Toyobo, Shanghai, China) in accordance with the manufacturer’s instructions. Quantitative real-time PCR assays were performed on a Bio-Rad CFX96 apparatus with the dye SYBR Green I (Invitrogen). The rice gene eEF1α was chosen as the internal control for the following analysis. The gene-specific primers are listed in the Supplemental Data, Table S1.
2.7. Measurement of Cd Content
The Cd content was analyzed according to a method described previously [34]. Briefly, rice seedlings treated with or without 200 μM CdCl2 for 5 days and roots and shoots were sampled; then, the samples were washed with sterile water and then dried at 80 °C for 1 d. The dried samples were digested with HNO3/HClO4 (4:1, v/v) at 180 °C and then the digested solution was diluted with sterile water for determination of Cd content using an atomic absorption spectrometer.
2.8. Measurement of Chlorophyll Content
The chlorophyll content was determined according to a previously described method [35]. Briefly, 0.5 g fresh leaves of plants from different lines which treated with or without 200 μM CdCl2 for 5 days were collected and incubated in 20 mL of 80% (v/v) acetone and kept in darkness for 24 h. After centrifugation, the extracted solutions were used for the total chlorophyll content determination.
2.9. Measurement of Melatonin Content
The melatonin content in rice tissues was determined as described previously [36]. Briefly, after treatment with or without 200 μM CdCl2 for 5 days, root and shoot samples were extracted with acetone: methanol: water (v:v:v = 89:10:1) and centrifuged. Subsequently, the supernatant was used for the melatonin content determination using a melatonin enzyme-linked immunosorbent assay (ELISA) kit.
2.10. Statistical Analysis
All experiments were performed in at least three independent biological replicates and three technical repetitions. The significance of differences was determined with ANOVA or Student’s t-test, as indicated in the figure legends.
3. Results
3.1. Exogenous NO Donor Alleviated Cd Toxicity in Rice Seedlings
To elucidate how NO mediates cadmium stress in rice, we examined the root and shoot growth in different Cd concentrations. As shown in Supplementary Figure S1A,B, the root and shoot length were markedly inhibited by 200 μM CdCl2 compared with the untreated control. Therefore, 200 μM CdCl2 was used in the subsequent experiments based on the above results.
Nitric oxide may regulate plant growth in a dose-dependent manner. We tested this hypothesis through experiments employing the exogenous application of different concentrations of NO donor SNP. As shown in Figure 1A,B, the root and shoot lengths were markedly reduced in the 50 and 100 μM SNP treatment groups compared with the untreated controls, whereas the plants treated with 20 μM SNP showed no significant changes compared with those in the control group. Moreover, the application of Cd in the 50 and 100 μM SNP treatment groups abrogated this effect, whereas Cd and 20 μM SNP alleviated the inhibitory effects on the growth of the roots and shoots compared with Cd treatment alone (Figure 1C,D). Therefore, 20 μM SNP was used in subsequent experiments.
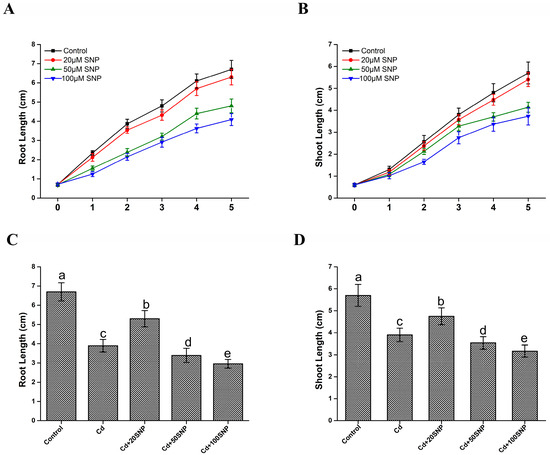
Figure 1.
Effects of exogenous SNP treatment on root and shoot growth under normal and cadmium-stress conditions. Two-day-old seedlings germinated on 1/2MS plates were transferred to new plates new plates containing 0, 20, 50, or 100 μM SNP, and then root (A) and shoot (B) lengths were measured at 1, 2, 3, 4, and 5 days after transfer. (C,D) The root (C) and shoot lengths (D) of rice seedlings were measured after 5 days of growth on the medium (1/2 MS containing 0, 20, 50, or 100 μM SNP with 200 μM CdCl2).The results shown are the means ± SD. Values were derived from three independent biological experiments. Different letters indicate significantly different values (p < 0.05 by Tukey’s test). Cd, 200 μM CdCl2; 20 SNP, 20 μM NP; 50 SNP, 50 μM SNP; 100 SNP, 100 μM SNP.
To verify whether Cd changed the NO level in rice seedlings, 1-week-old wild-type rice seedlings were treated in 200 μM CdCl2, and the NO level in the roots was assayed at 24 h post treatment. The results of fluorescence analysis showed increased NO levels induced by cadmium, which were strongly reduced by an NOS inhibitor (L-NAME) (Figure 2A), suggesting that Cd stress can modulate NO content, possibly through NOS activity.
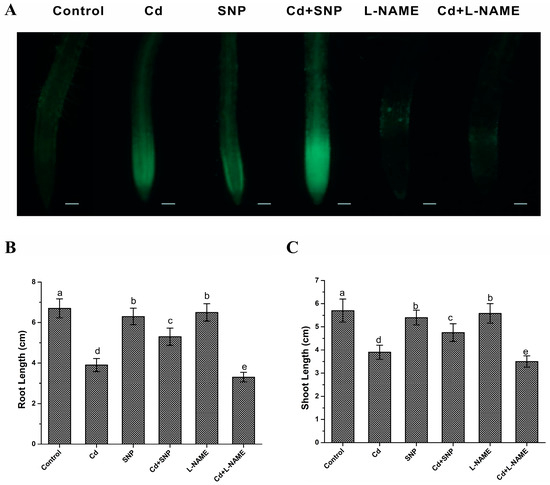
Figure 2.
Endogenous NO level in response to Cd stress and the effect of exogenous NO donor on Cd toxicity in rice seedlings. (A) Effect of cadmium stress on the endogenous NO level in the roots of wild-type rice plants. One-week-old wild-type plants were treated with 200 μM CdCl2 combined with or without 1 mM L-NAME for 24 h, and then NO production was measured. NO fluorescence in roots of the nNOS-overexpressing lines and wild-type plants examined using DAF-FM DA. (B,C) Effects of SNP or L-NAME on the root (B) and shoot (C) growth under cadmium stress. The results shown are the mean ± SD. Values were derived from three independent biological experiments. Different letters indicate significant differences using Tukey’s multiple comparison test at p < 0.05. Bars = 50 μm; Cd, 200 μM CdCl2; SNP, 20 μM SNP; L-NAME, 1 mM L-NAME.
Then, the effects of exogenous NO treatment on Cd resistance in rice seedlings were examined. Germinated wild-type rice seedlings were transferred to 1/2 MS medium containing 200 μM CdCl2 with or without 1 mM L-NAME or 20 μM SNP, and then root and shoot lengths were measured 5 d after transfer. As shown in Figure 2B,C, the combined application of CdCl2 and SNP reduced the inhibitory effect of cadmium treatment on the root and shoot lengths, whereas CdCl2 and L-NAME treatment abrogated the inhibitory effect on the growth of roots and shoots compared with cadmium treatment alone. Taken together, NO is induced by Cd stress and exogenous NO donor can alleviate Cd toxicity in rice seedlings. These findings suggest that NO plays a positive role in the cadmium-mediated inhibition of the growth of rice seedlings.
3.2. nNOS-Overexpressing Rice Plants Demonstrated Improved Tolerance to Cadmium Stress
Due to the lack of materials with altered NO levels, the role of endogenous NO in rice responses to Cd stress is still limited. As such, in this study, the tolerance of nNOS-overexpressing rice plants 35S:nNOS (#2, #8 and #20), which had higher NOS activity and NO content, to Cd stress was investigated (Figure 3A,B). For this purpose, 2-day-old seedlings of both wild-type and transgenic lines were transferred onto 1/2 MS medium with or without supplementation with 200 μM CdCl2, and both shoot and root lengths were assayed at 5 days after transfer. Although all the tested transgenic lines exhibited shoot and root lengths similar to those of the wild-type lines when grown in normal 1/2 MS medium, the transgenic lines were less sensitive to cadmium stress in terms of changes in shoot and root lengths (Figure 3C–F). Under Cd stress, the chlorophyll content of the wild-type plants decreased remarkably, whereas that of the nNOS-overexpressing rice plants remained relatively higher (Figure 3G). These results indicate that nNOS overexpression in rice can improve cadmium tolerance at the seedling stage.
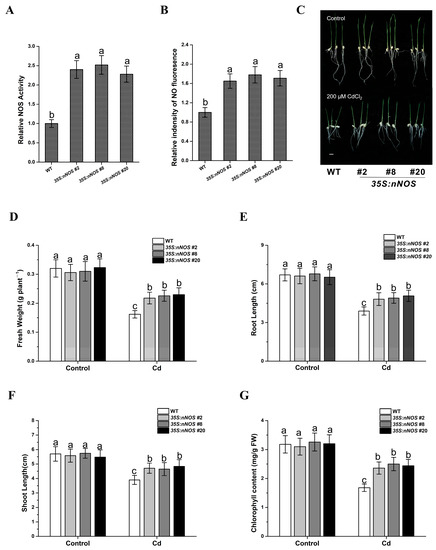
Figure 3.
The nNOS-overexpressing lines showed enhanced cadmium tolerance. (A) NOS activities of the nNOS-overexpressing lines and wild-type plants were determined using a NOS assay kit. The relative NOS activity is expressed using the NOS activity of the wild type as the standard (1). (B) Relative NO content of the nNOS-overexpressing lines and wild-type plants examined using DAF-FM DA and expressed using the fluorescence of the wild-type plants as the standard (1). (C) Germinated wild-type and nNOS-overexpressing plants at 5 d after treatment with or without 200 μM CdCl2. (D–F) The fresh weight (D), root length (E), shoot length (F), and total chlorophyll content (G) were assayed with both wild-type plants and three nNOS-overexpressing lines after 5 days of growth on the medium (1/2 MS with or without 200 μM CdCl2). The results shown are the means ± SD. Values were derived from three independent biological experiments, and the different letters indicate significant differences between the annotated columns (p < 0.05 by Tukey’s test). Bars = 1 cm; FW, fresh weight; Cd, 200 μM CdCl2.
3.3. nNOS-Overexpressing Rice Plants Accumulate Less Cadmium under Cadmium Stress
To determine whether the improved Cd tolerance of nNOS-overexpressing plants was associated with Cd accumulation in plant tissues, we transferred 2-day-old seedlings of both the wild-type and transgenic lines onto 1/2 MS medium with or without supplementation with 200 μM CdCl2, and then the Cd contents of the shoot and roots were assayed 5 days after transfer. Under Cd stress, the Cd contents significantly decreased in both the roots and shoots of nNOS transgenic rice plants compared with wild-type plants (Figure 4A,B). Then, we used reverse-transcription quantitative PCR (RT-qPCR) to examine the expression levels of genes involved in Cd transport. Compared with the WT control, the expressions of the cadmium transporter genes OsCAL1, OsIRT2, OsNramp5, and OsCd1 were significantly downregulated in the nNOS-overexpressing lines under cadmium treatment, whereas cadmium induced their expression (Figure 4C–F), suggesting that nitric oxide may reduce the accumulation of cadmium ions in rice seedlings by reducing the expressions of the genes encoding cadmium transporters.
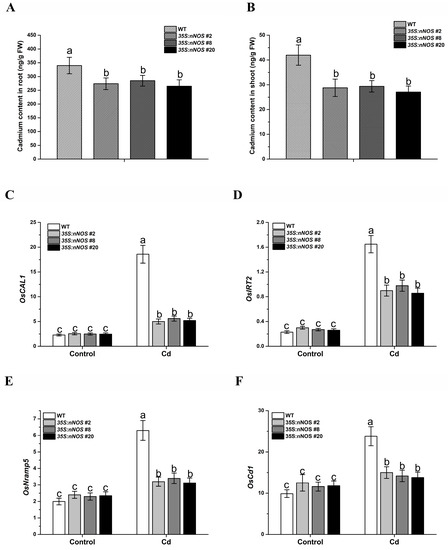
Figure 4.
nNOS-overexpressing rice plants accumulated less cadmium under cadmium stress. Germinated wild-type and transgenic plants were treated with or without 200 μM CdCl2 for 5 days, and then root (A) and shoot (B) cadmium contents, and the expression of OsCAL1 (C), OsIRT2 (D), OsNramp5 (E), and OsCd1 (F) were assayed. The results shown are the means ± SD. Values were derived from three independent biological experiments, and the different letters indicate significant differences between the annotated columns (p < 0.05 by Tukey’s test). FW, fresh weight; Cd, 200 μM CdCl2.
3.4. ROS-Scavenging Capacity of nNOS-Overexpressing Rice Plants Was Enhanced
Much of the injury caused by cadmium at the cellular level is associated with oxidative damage due to ROS [1]. Additionally, treatment with the NO donor SNP can decrease ROS accumulation, thus alleviating oxidative damage in cadmium-stressed plants [7]. Therefore, we expected that our transgenic plants with higher NO contents would also repress cadmium-induced ROS accumulation, thereby achieving higher tolerance to Cd stress. For this purpose, we assayed the H2O2 contents of both wild-type and transgenic plants. The results showed that the transgenic plants repressed the H2O2 accumulation induced by cadmium stress in the wild-type plants; however, both wild-type and transgenic plants accumulated similar contents of H2O2 under normal conditions (Figure 5A). The reduced Cd-induced H2O2 content in transgenic plants may result from changes in the activities of antioxidant enzymes that can scavenge H2O2, such as CAT and POX. As expected, the transgenic plants showed much higher CAT and POX activities under Cd-stress conditions compared with the wild-type plants (Figure 5B,C). Furthermore, qRT-PCR analysis showed that the expression levels of antioxidant enzymes genes such as OsCATA, OsCATB, and OsPOX1 in the transgenic lines were significantly higher than those in wild-type plants when subjected to Cd stress (Figure 5D–F). These results suggest that endogenous NO improves Cd tolerance by increasing ROS-scavenging capacity under Cd stress.
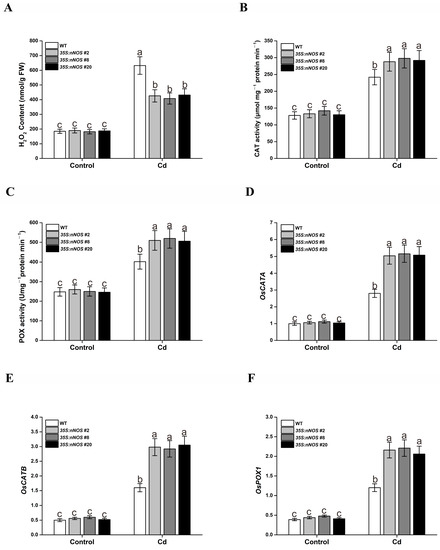
Figure 5.
The nNOS-overexpressing lines exhibited improved ROS-scavenging capacity under cadmium stress. Germinated wild-type and nNOS-overexpressing plants were treated with or without 200 μM CdCl2 for 5 days, and then H2O2 contents (A), enzymatic activities of CAT (B) and POX (C), and the expression of OsCATA (D), OsCATB (E) and OsPOX1 (F) were assayed. The results shown are the means ± SD. Values were derived from three independent biological experiments, and the different letters indicate significant differences between the annotated columns (p < 0.05 by Tukey’s test). FW, fresh weight; Cd, 200 μM CdCl2.
3.5. nNOS Transgenic Plants Showed Increased Melatonin Levels under Cadmium Stress
Many studies have reported that melatonin plays a similar role to NO in the tolerance of plants to Cd stress [24]. However, the relationship between NO and melatonin in response to Cd stress in plants is still unclear [37]. In this study, the melatonin content in nNOS-overexpressing transgenic lines was measured under cadmium stress. For this purpose, 2-day-old seedlings of both the wild-type and transgenic lines were transferred onto 1/2 MS medium with or without supplementation with 200 μM CdCl2, and the melatonin contents were measured 5 d after transfer. As shown in Figure 6A,B, the melatonin levels significantly increased in the cadmium-treated plants compared with the untreated plants, and the melatonin contents in the nNOS-overexpressing transgenic lines were markedly higher than those in the wild-type plants. The relative expression levels of the genes of the melatonin metabolism process, including OsASMT1, OsTDC3, OsTDC1, and OsSNAT2, increased under cadmium treatment in the transgenic lines compared with the WT lines (Figure 6C–F). All these results indicate that NO may reduce cadmium accumulation and improve cadmium tolerance in rice seedlings through modulation of melatonin biosynthesis.
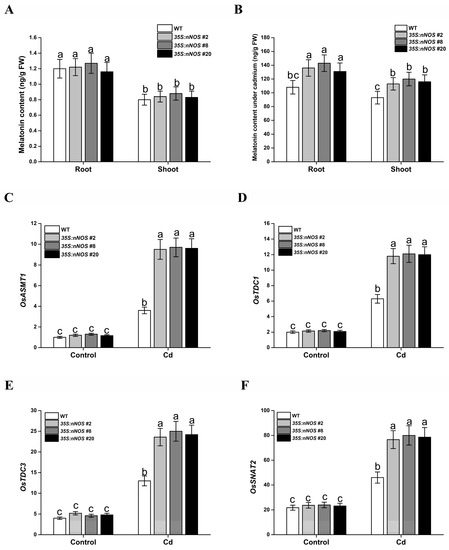
Figure 6.
nNOS-overexpressing rice plants accumulate less cadmium under cadmium stress. Germinated wild-type and nNOS-overexpressing plants were treated with or without 200 μM CdCl2 for 5 days, and then root (A) and shoot (B) melatonin contents, and the expression of OsASMT1 (C), OsTDC1 (D), OsTDC3 (E), and OsSNAT2 (F) were assayed. The results shown are the means ± SD. Values were derived from three independent biological experiments, and the different letters indicate significant differences between the annotated columns (p < 0.05 by Tukey’s test). FW, fresh weight; Cd, 200 μM CdCl2.
4. Discussion
Recently, many studies have reported the function of NO in response to Cd stress in plants. However, the role of NO in plant responses to Cd stress remains controversial [7]. This controversy may be due to the complex properties of NO, the functions of which depend on its location and concentration, as well as the plant species and plant development stage.
We present herein our results, showing that 200 μM CdCl2 treatment induces NO accumulation in the roots of rice seedlings. This result is consistent with those of studies in which NO accumulation was enhanced after treatment with different concentrations of cadmium [11,13], although several studies have reported a decrease in NO content in rice upon cadmium treatment [10,15,38]. This difference may be due to the use of different Cd concentrations, treatment times, plant sizes, or genotypes. In addition, most results regarding NO function were achieved with the application of NO donors or scavengers, which may not adequately replicate the function of endogenous NO in plants. Therefore, an exploration of the roles of endogenous NO in response to Cd in rice also appears to be of great interest and importance. In this study, we also examined the effects of exogenous NO treatment on Cd resistance in rice seedlings, and further investigated the tolerance of nNOS-overexpressing rice plants with higher NOS activity and NO content to Cd stress. Consistent with the results of studies in which the application of an NO donor enhanced cadmium stress, the transgenic rice plants showed higher Cd tolerance and lower Cd accumulation.
When plants experience cadmium stress, the antioxidant system is activated to reduce the overproduction of ROS. Exogenous NO can alleviate Cd-induced oxidative damage by activating the antioxidant system and strengthening its ability to scavenge ROS [19,39]. NO can also scavenge excess ROS by directly binding to O2− to form peroxynitrite (ONOO−) [40]. Our results indicates that transgenic nNOS rice plants with higher NO levels accumulated less H2O2 under Cd stress conditions, possibly due to the upregulation of the expression of antioxidant enzyme genes (OsCATA, OsCATB, and OsPOX1). Whether the transgenic plants affect other ROS and how these antioxidant enzyme genes are regulated under Cd stress remains to be explored.
Exogenous application of an NO donor also affects the fixation of Cd and Cd uptake, thereby regulating the Cd tolerance and Cd accumulation of plants. For example, 0.1 mM SNP treatment increases the accumulation of Cd in the cell wall of roots but decreases Cd accumulation in the soluble fraction of leaves and roots in rice under Cd stress [15]. Exogenous NO can also reduce Cd uptake and transport in rice [19,39]. However, these studies mainly focus on the morphological and physiological indexes. In this study, nNOS-overexpressing rice plants accumulated less cadmium under Cd stress conditions, possibly due to the downregulation of the transcription of Cd uptake- and transport-related genes such as OsCAL1, OsIRT2, OsNramp5, and OsCd1.
Similar to the physiological functions of NO, melatonin alleviates the Cd-induced inhibition of seedling growth [24]. However, the relationship between NO and melatonin is still elusive. For example, exogenous melatonin improved Cd tolerance by reducing NO accumulation, resulting in lower Cd accumulation in Brassica pekinensis (Lour.) Rupr. [41], but triggered endogenous NO and alleviated Cd toxicity by increasing the activities of antioxidant enzymes in wheat seedlings [26]. These results suggest that NO may act as a downstream signal in response to Cd stress. However, Lee et al., in 2017, reported that cadmium-induced melatonin synthesis in rice requires NO [23]. In this paper, we present data showing that NO upgregulates the expression of melatonin-biosynthesis-related genes, thus enhancing melatonin accumulation under cadmium stress. These findings suggest that NO may improve Cd tolerance by promoting melatonin biosynthesis. The complicated relationship between NO and melatonin requires further investigation.
Overall, this study showed the effects of exogenous and endogenous NO on the resistance of rice seedlings to Cd stress. NO was induced by Cd stress and exogenous NO donors were able to alleviate Cd toxicity in a dose-dependent manner. Rice plants overexpressing rat nNOS showed increases in both NOS activity and NO content, resulting in improved Cd stress tolerance. Further analyses indicated that improving endogenous NO enhances cadmium tolerance in rice through modulation of cadmium accumulation, antioxidant capacity and melatonin biosynthesis (Figure 7). However, the manner in which NO modulates the expression of genes involved in these pathways and whether NO affects other pathways in order to alleviate Cd stress need to be further investigated.
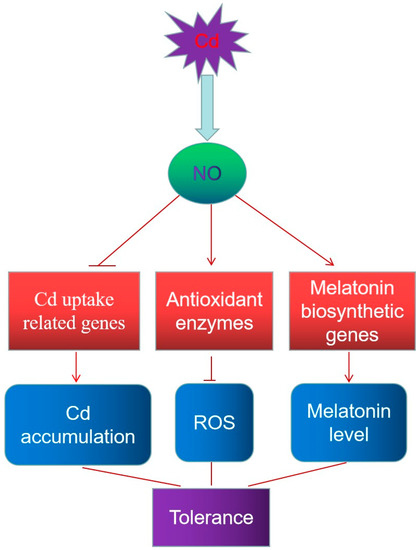
Figure 7.
Model of the role of endogenous NO in Cd stress. Cd stress induces NO accumulation, and the generated NO enhances Cd tolerance through modulation of the contents of Cd, ROS, and melatonin, possibly by regulating the transcription level of genes involved in Cd uptake and antioxidant and melatonin biosynthesis.
5. Conclusions
When rice plants were stressed with Cd, the growth and development were significantly inhibited and excessive Cd was accumulated. The application of an exogenous NO donor alleviated Cd toxicity, and nNOS-overexpressing rice plants with higher NO levels showed improved Cd stress tolerance. Moreover, the transgenic rice plants accumulated less Cd, and had higher melatonin levels and stronger ROS-scavenging capacity, as well as regulating the transcription of related genes under Cd conditions to achieve higher tolerance to Cd stress. In summary, improving endogenous NO levels through exogenous application of SNP or transgenic technology can improve Cd tolerance and reduce Cd accumulation in rice seedlings.
Supplementary Materials
The following supporting information can be downloaded at: https://www.mdpi.com/article/10.3390/agronomy13081978/s1, Table S1. Primers used for qRT-PCR analysis. F: forward; R: reverse; Figure S1. Cadmium stress repress root and shoot growth of rice seedlings.
Author Contributions
Data curation, G.Z. and T.Y.; Formal analysis, W.C.; Investigation, H.D. and P.W.; Methodology, W.C., W.W., H.D., B.C., G.Z. and P.W.; Project administration, W.C. and Y.Z.; Resources, B.C.; Supervision, Y.Z.; Writing—original draft, W.C. and T.Y.; Writing—review and editing, W.W. and T.Y. All authors have read and agreed to the published version of the manuscript.
Funding
This study was mainly supported by the Hubei Provincial Natural Science Foundation (2019CFB399) and Knowledge Innovation Program of Wuhan-Basi Research (2020020601012298).
Data Availability Statement
The datasets generated and/or analyzed during the study are available from the senior author upon reasonable request.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Genchi, G.; Sinicropi, M.S.; Lauria, G.; Carocci, A.; Catalano, A. The effects of cadmium toxicity. Int. J. Environ. Res. Public Health 2020, 17, 3782. [Google Scholar] [CrossRef]
- Zhang, J.; Zhu, Y.; Yu, L.; Yang, M.; Zou, X.; Yin, C.; Lin, Y. Research advances in cadmium uptake, transport and resistance in rice (Oryza sativa L.). Cells 2022, 11, 569. [Google Scholar] [CrossRef]
- Kumar, D.; Ohri, P. Say “no” to plant stresses: Unravelling the role of nitric oxide under abiotic and biotic stress. Nitric Oxide 2022, 130, 36–57. [Google Scholar] [CrossRef] [PubMed]
- Khan, M.; Ali, S.; Azzawi, T.N.I.A.; Yun, B. Nitric oxide acts as a key signaling molecule in plant development under stressful conditions. Int. J. Mol. Sci. 2023, 24, 4782. [Google Scholar] [CrossRef] [PubMed]
- Gupta, K.J.; Kaladhar, V.C.; Fitzpatrick, T.B.; Fernie, A.R.; Møller, I.M.; Loake, G.J. Nitric oxide regulation of plant metabolism. Mol. Plant 2021, 15, 228–242. [Google Scholar] [CrossRef]
- Gill, S.S.; Hasanuzzaman, M.; Nahar, K.; Macovei, A.; Tuteja, N. Importance of nitric oxide in cadmium stress tolerance in crop plants. Plant Physiol. Biochem. 2012, 63, 254–261. [Google Scholar] [CrossRef] [PubMed]
- Meng, Y.; Jing, H.; Huang, J.; Shen, R.; Zhu, X. The role of nitric oxide signaling in plant responses to cadmium stress. Int. J. Mol. Sci. 2022, 23, 6901. [Google Scholar] [CrossRef] [PubMed]
- Singh, S.; Shrivastava, A.K.; Singh, V.K. Arsenic and cadmium are inhibitors of cyanobacterial dinitrogenase reductase (nifh1) gene. Funct. Integr. Genom. 2014, 14, 571–580. [Google Scholar] [CrossRef]
- Xiong, J.; Lu, H.; Lu, K.; Duan, Y.; An, L.; Zhu, C. Cadmium decreases crown root number by decreasing endogenous nitric oxide, which is indispensable for crown root primordia initiation in rice seedlings. Planta 2009, 230, 599–610. [Google Scholar] [CrossRef]
- Piacentini, D.; Ronzan, M.; Fattorini, L.; Rovere, F.D.; Massimi, L.; Altamura, M.M.; Falasca, G. Nitric oxide alleviates cadmium- but not arsenic-induced damages in rice roots. Plant Physiol. Biochem. 2020, 151, 729–742. [Google Scholar] [CrossRef]
- Wang, T.; Shi, Z.Q.; Hu, L.; Xu, X.; Han, F.X.; Zhou, L.; Chen, J. Thymol ameliorates cadmium-induced phytotoxicity in the root of rice (Oryza sativa) seedling by decreasing endogenous nitric oxide generation. J. Agric. Food Chem. 2017, 65, 7396–7405. [Google Scholar] [CrossRef] [PubMed]
- Valentovicová, K.; Halusková, L.; Huttová, J.; Mistrík, I.; Tamás, L. Effect of cadmium on diaphorase activity and nitric oxide production in barley root tips. J. Plant Physiol. 2009, 167, 10–14. [Google Scholar] [CrossRef]
- Yang, L.; Ji, J.; Harris, S.; Karen, R.; Wang, H.; Wang, H.; Abd, A.; Elsayed, F.; Luo, Y.; Hu, X. The dynamic changes of the plasma membrane proteins and the protective roles of nitric oxide in rice subjected to heavy metal cadmium stress. Front. Plant Sci. 2016, 7, 190. [Google Scholar] [CrossRef] [PubMed]
- Rodríguez-serrano, M.; Romero-puertas, M.C.; Pazmiño, D.M.; Testillano, P.S.; Risueño, M.C.; Río, L.A.D.; Sandalio, L.M. Cellular response of pea plants to cadmium toxicity: Cross talk between reactive oxygen species, nitric oxide, and calcium. Plant Physiol. 2009, 150, 229–243. [Google Scholar] [CrossRef] [PubMed]
- Xiong, J.; An, L.; Lu, H.; Zhu, C. Exogenous nitric oxide enhances cadmium tolerance of rice by increasing pectin and hemicellulose contents in root cell wall. Planta 2009, 230, 755–765. [Google Scholar] [CrossRef]
- Arasimowicz-jelonek, M.; Floryszak-wieczorek, J.; Deckert, J.; Rucińska-sobkowiak, R.; Gzyl, J.; Pawlak-sprada, S.; Abramowski, D.; Jelonek, T.; Gwóźdź, E.A. Nitric oxide implication in cadmium-induced programmed cell death in roots and signaling response of yellow lupine plants. Plant Physiol. Biochem. 2012, 58, 124–134. [Google Scholar] [CrossRef] [PubMed]
- Michele, R.D.; Vurro, E.; Rigo, C.; Costa, A.; Elviri, L.; Valentin, M.D.; Careri, M.; Zottini, M.; Toppi, L.S.D.; Schiavo, F.L. Nitric oxide is involved in cadmium-induced programmed cell death in arabidopsis suspension cultures. Plant Physiol. 2009, 150, 217–228. [Google Scholar] [CrossRef] [PubMed]
- Verma, K.; Mehta, S.K.; Shekhawat, G.S. Nitric oxide (NO) counteracts cadmium induced cytotoxic processes mediated by reactive oxygen species (ROS) in brassica juncea: Cross-talk between ROS, NO and antioxidant responses. Biometals 2013, 26, 255–269. [Google Scholar] [CrossRef]
- Singh, P.; Shah, K. Evidences for reduced metal-uptake and membrane injury upon application of nitric oxide donor in cadmium stressed rice seedlings. Plant Physiol. Biochem. 2014, 83, 180–184. [Google Scholar] [CrossRef]
- Leitner, M.; Vandelle, E.; Gaupels, F.; Bellin, D.; Delledonne, M. NO signals in the haze: Nitric oxide signalling in plant defence. Curr. Opin. Plant Biol. 2009, 12, 451–458. [Google Scholar] [CrossRef]
- Grün, S.; Lindermayr, C.; Sell, S.; Durner, J. Nitric oxide and gene regulation in plants. J. Exp. Bot. 2006, 57, 507–516. [Google Scholar] [CrossRef] [PubMed]
- Xu, J.; Wang, W.; Yin, H.; Liu, X.; Sun, H.; Mi, Q. Exogenous nitric oxide improves antioxidative capacity and reduces auxin degradation in roots of Medicago truncatula seedlings under cadmium stress. Plant Soil 2009, 326, 321. [Google Scholar] [CrossRef]
- Lee, K.; Choi, G.H.; Back, K. Cadmium-induced melatonin synthesis in rice requires light, hydrogen peroxide, and nitric oxide: Key regulatory roles for tryptophan decarboxylase and caffeic acid O-methyltransferase. J. Pineal Res. 2017, 63, e12441. [Google Scholar] [CrossRef]
- Gu, Q.; Wang, C.; Xiao, Q.; Chen, Z.; Han, Y. Melatonin confers plant cadmium tolerance: An update. Int. J. Mol. Sci. 2021, 22, 11704. [Google Scholar] [CrossRef] [PubMed]
- Gu, Q.; Chen, Z.; Yu, X.; Cui, W.; Pan, J.; Zhao, G.; Xu, S.; Wang, R.; Shen, W. Melatonin confers plant tolerance against cadmium stress via the decrease of cadmium accumulation and reestablishment of microrna-mediated redox homeostasis. Plant Sci. 2017, 261, 28–37. [Google Scholar] [CrossRef] [PubMed]
- Kaya, C.; Okant, M.; Ugurlar, F.; Alyemeni, M.N.; Ashraf, M.; Ahmad, P. Melatonin-mediated nitric oxide improves tolerance to cadmium toxicity by reducing oxidative stress in wheat plants. Chemosphere 2019, 225, 627–638. [Google Scholar] [CrossRef] [PubMed]
- He, J.; Zhuang, X.; Zhou, J.; Sun, L.; Wan, H.; Li, H.; Lyu, D. Exogenous melatonin alleviates cadmium uptake and toxicity in apple rootstocks. Tree Physiol. 2020, 40, 746–761. [Google Scholar] [CrossRef] [PubMed]
- Terrón-Camero, L.C.; Peláez-Vico, M.Á.; Del-val, C.; Sandalio, L.M.; Romero-Puertas, M.C. Role of nitric oxide in plant responses to heavy metal stress: Exogenous application versus endogenous production. J. Exp. Bot. 2019, 70, 4477–4488. [Google Scholar] [CrossRef] [PubMed]
- Gupta, K.J.; Fernie, A.R.; Kaiser, W.M.; Dongen, J.T.V. On the origins of nitric oxide. Trends Plant Sci. 2010, 16, 160–168. [Google Scholar] [CrossRef]
- Lin, A.; Wang, Y.; Tang, J.; Xue, P.; Li, C.; Liu, L.; Hu, B.; Yang, F.; Loake, G.J.; Chu, C. Nitric oxide and protein S-nitrosylation are integral to hydrogen peroxide-induced leaf cell death in rice. Plant Physiol. 2012, 158, 451–464. [Google Scholar] [CrossRef]
- Chun, H.J.; Park, H.C.; Koo, S.C.; Lee, J.H.; Park, C.Y.; Choi, M.S.; Kang, C.H.; Baek, D.; Cheong, Y.H.; Yun, D.; et al. Constitutive expression of mammalian nitric oxide synthase in tobacco plants triggers disease resistance to pathogens. Mol. Cells 2012, 34, 463–471. [Google Scholar] [CrossRef] [PubMed]
- Shi, H.; Li, R.; Cai, W.; Liu, W.; Wang, C.; Lu, Y. Increasing nitric oxide content in arabidopsis thaliana by expressing rat neuronal nitric oxide synthase resulted in enhanced stress tolerance. Plant Cell Physiol. 2011, 53, 344–357. [Google Scholar] [CrossRef] [PubMed]
- Cai, W.; Liu, W.; Wang, W.; Fu, Z.; Han, T.; Lu, Y. Overexpression of rat neurons nitric oxide synthase in rice enhances drought and salt tolerance. PLoS ONE 2015, 10, e0131599. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Q.; Cai, W.; Ji, T.; Ye, L.; Lu, Y.; Yuan, T. WRKY13 enhances cadmium tolerance by promoting D-cysteine desulfhydrase and hydrogen sulfide production. Plant Physiol. 2020, 183, 345–357. [Google Scholar] [CrossRef] [PubMed]
- Qian, L.; Song, F.; Xia, J.; Wang, R. A Glucuronic acid-producing endophyte pseudomonas sp. MCS15 reduces cadmium uptake in rice by inhibition of ethylene biosynthesis. Front. Plant Sci. 2022, 13, 876545. [Google Scholar] [CrossRef]
- Wei, Y.; Liu, G.; Chang, Y.; Lin, D.; Reiter, R.J.; He, C.; Shi, H. Melatonin biosynthesis enzymes recruit wrky transcription factors to regulate melatonin accumulation and transcriptional activity on w-box in cassava. J. Pineal Res. 2018, 65, e12487. [Google Scholar] [CrossRef]
- He, H.; He, L. Crosstalk between melatonin and nitric oxide in plant development and stress responses. Physiol. Plantarum 2020, 170, 218–226. [Google Scholar] [CrossRef]
- Zhang, L.; Chen, Z.; Zhu, C. Endogenous nitric oxide mediates alleviation of cadmium toxicity induced by calcium in rice seedlings. J. Environ. Sci. 2012, 24, 940–948. [Google Scholar] [CrossRef]
- Mostofa, M.G.; Rahman, M.M.; Ansary, M.M.U.; Fujita, M.; Tran, L.P. Interactive effects of salicylic acid and nitric oxide in enhancing rice tolerance to cadmium stress. Int. J. Mol. Sci. 2019, 20, 5798. [Google Scholar] [CrossRef]
- Delledonne, M.; Zeier, J.; Marocco, A.; Lamb, C. Signal interactions between nitric oxide and reactive oxygen intermediates in the plant hypersensitive disease resistance response. Proc. Natl. Acad. Sci. USA 2001, 98, 13454–13459. [Google Scholar] [CrossRef]
- Wang, T.; Song, J.; Liu, Z.; Liu, Z.; Cui, J. Melatonin alleviates cadmium toxicity by reducing nitric oxide accumulation and IRT1 expression in chinese cabbage seedlings. Environ. Sci. Pollut. Res. 2020, 28, 15394–15405. [Google Scholar] [CrossRef] [PubMed]







Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).