

Comparison of the Efficacy of Trichoderma and Bacillus Strains and Commercial Biocontrol Products against Grapevine Botryosphaeria Dieback Pathogens

 , ,
, ,  ,
, 
Abstract
1. Introduction
2. Material and Methods
2.1. Plant Material
2.2. Fungal Isolates
2.3. Treatments
2.4. Production of T. harzianum and B. velezensis Treatments
2.5. Greenhouse Bioassays on Grafted Plants
2.6. Statistical Analyses
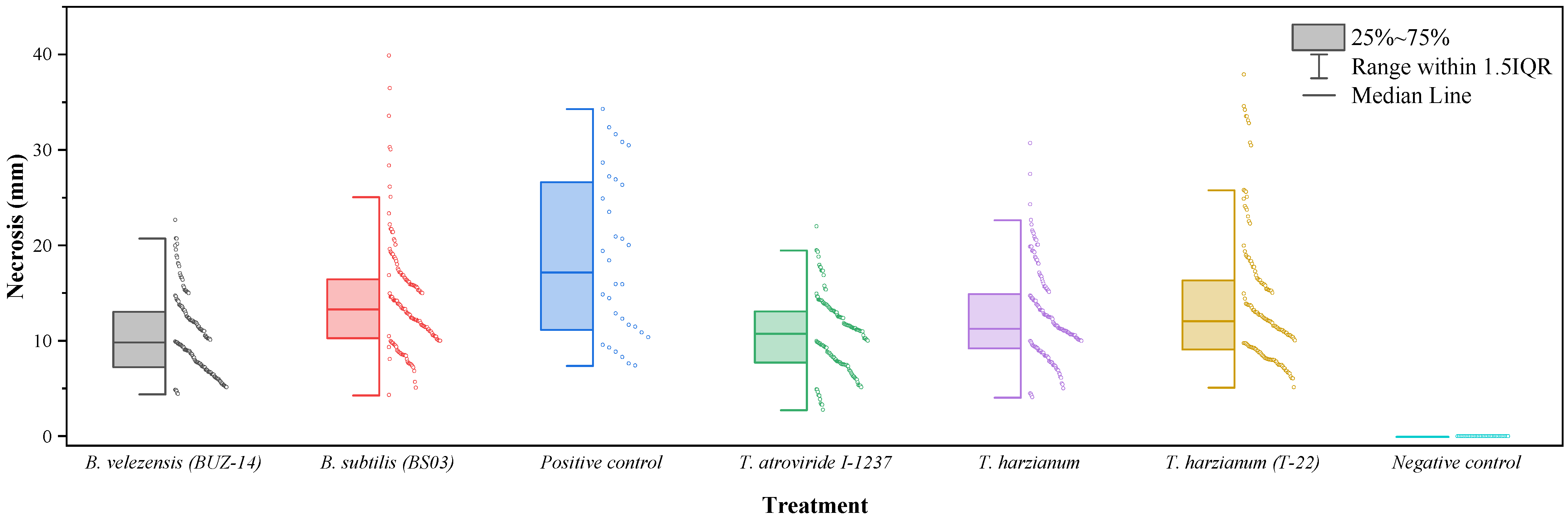
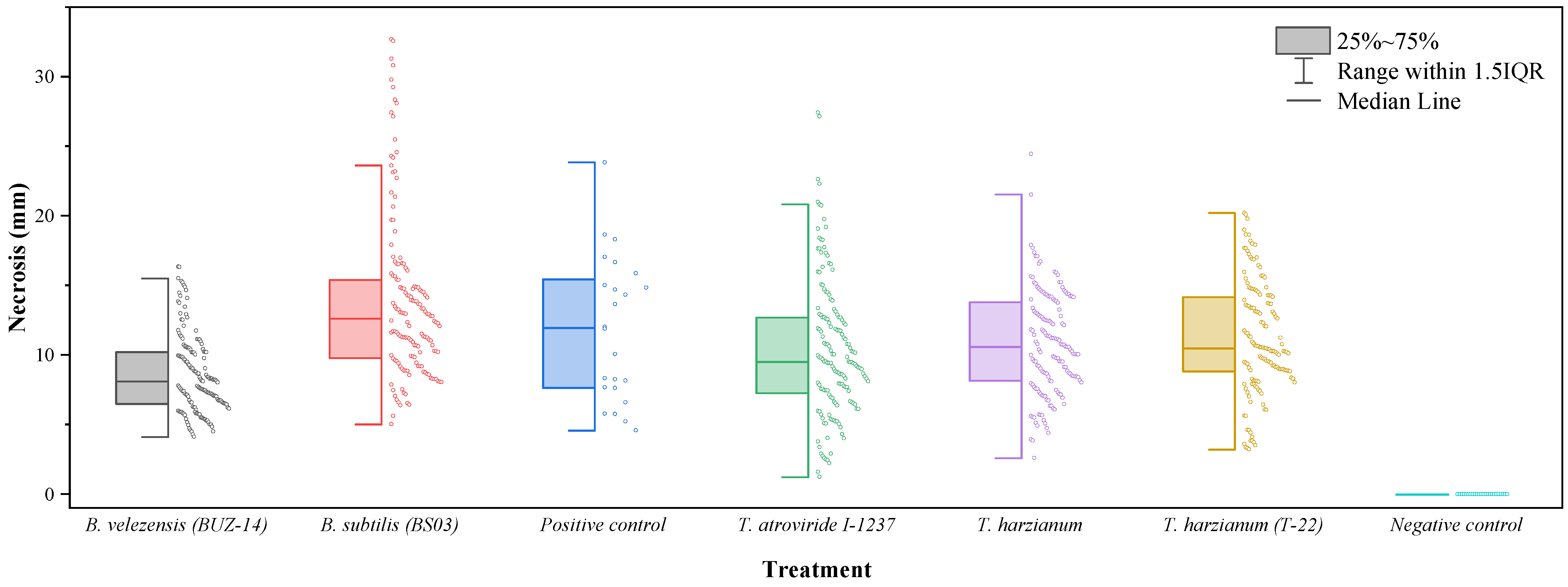
3. Results
3.1. Comparison of Efficacies against N. parvum
3.2. Comparison of Efficacies against D. seriata
4. Discussion
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
Appendix A

References
- Mondello, V.; Songy, A.; Battison, E.; Pinto, C.; Coppin, C.; Trozel-Aziz, P.; Clement, C.; Mugnai, L.; Fontaine, F. Grapevine trunk diseases: A review of fifteen years of trials for their control with chemicals and biocontrol agents. Plant Dis. 2018, 102, 1189–1217. [Google Scholar] [CrossRef]
- Gramaje, D. Enfermedades de la madera de la vid: Situación actual y estrategias de control. In Proceedings of the Jornadas Vitivinícolas; Instituto de Ciencias de la Vid y del Vino (ICVV): La Rioja, Spain, 2017. [Google Scholar]
- Gramaje, D.; Urbez-Torres, J.R.; Sosnowski, M.R. Managing grapevine trunk diseases with respect to etiology and epidemiology: Current strategies and future prospects. Plant Dis. 2018, 102, 12–39. [Google Scholar] [CrossRef] [PubMed]
- Leal, C.; Richet, N.; Guise, J.F.; Gramaje, D.; Armengol, J.; Fontaine, F.; Trotel-Aziz, P. Cultivar contributes to the beneficial effects of Bacillus subtilis PTA-271 and Trichoderma atroviride SC1 to protect grapevine against Neofusicoccum parvum. Front. Microbiol. 2021, 12, 726132. [Google Scholar] [CrossRef] [PubMed]
- Benítez, T.; Rincón, A.M.; Limón, M.C.; Codón, A.C. Biocontrol mechanisms of Trichoderma strains. Int. Microbiol. 2004, 7, 249–260. [Google Scholar] [PubMed]
- TariqJaveed, M.; Farooq, T.; Al-Hazmi, A.S.; Hussain, M.D.; Rehman, A.U. Role of Trichoderma as a biocontrol agent (BCA) of phytoparasitic nematodes and plant growth inducer. J. Invertebr. Pathol. 2021, 183, 107626. [Google Scholar] [CrossRef]
- Pertot, I.; Prodorutti, D.; Colombini, A.; Pasini, L. Trichoderma atroviride SC1 prevents Phaeomoniella chlamydospora and Phaeoacremonium aleophilum infection of grapevine plants during the grafting process in nurseries. BioControl 2016, 61, 257–267. [Google Scholar] [CrossRef]
- Pollard-Flamand, J.; Boule, J.; Hart, M.; Urbez-Torres, J.R. Biocontrol activity of Trichoderma species isolated from grapevines in British Columbia against Botryosphaeria dieback fungal pathogens. J. Fungi 2022, 8, 409. [Google Scholar] [CrossRef]
- Gramaje, D.; Armengol, J.; Barajas, E.; Berbegal, M.; Chacón, J.L.; Cibriaín Sabalza, J.F.; Díaz-Losada, E.; López-Manzanares, B.; Muñoz Gómez, R.; Martínez-Diz, M. Guía Sobre las Enfermedades Fúngicas de la Madera de la Vid; Ministerio de Agricultura, Pesca y Alimentación: Madrid, Spain, 2020. [Google Scholar]
- Kotze, C.; Van Niekerk, J.; Mostert, L.; Halleen, F.; Fourie, P.H. Evaluation of biocontrol agents for grapevine pruning wound protection against trunk pathogen infection. Phytopathol. Mediterr. 2011, 50, S247–S263. [Google Scholar]
- Urbez-Torres, J.R.; Tomaselli, E.; Pollard-Flamand, J.; Boule, J.; Gerin, D.; Pollastro, S. Characterization of Trichoderma isolates from southern Italy, and their potential biocontrol activity against grapevine trunk disease fungi. Phytopathol. Mediterr. 2020, 59, 425–439. [Google Scholar] [CrossRef]
- Blundell, R.; Eskalen, A. Biological and chemical pruning wound protectants reduce infection of grapevine trunk disease pathogens. Calif. Agric. 2021, 75, 128–134. [Google Scholar] [CrossRef]
- Martinez-Diz, M.D.; Diaz-Losada, E.; Diaz-Fernandez, A.; Bouzas-Cid, Y.; Gramaje, D. Protection of grapevine pruning wounds against Phaeomoniella chlamydospora and Diplodia seriata by commercial biological and chemical methods. Crop Prot. 2021, 143, 105465. [Google Scholar] [CrossRef]
- Rezgui, A.; Ben Ghnaya-Chakroun, A.; Vallance, J.; Bruez, E.; Hajlaoui, M.R.; Sadfi-Zouaoui, N.; Rey, P. Endophytic bacteria with antagonistic traits inhabit the wood tissues of grapevines from Tunisian vineyards. Biol. Control 2016, 99, 28–37. [Google Scholar] [CrossRef]
- Langa-Lomba, N.; Martín-Ramos, P.; Casanova-Gascón, J.; Julián-Lagunas, C.; González-García, V. Potential of native Trichoderma strains as antagonists for the control of fungal wood pathologies in young grapevine plants. Agronomy 2022, 12, 336. [Google Scholar] [CrossRef]
- van Lenteren, J.C.; Bolckmans, K.; Köhl, J.; Ravensberg, W.J.; Urbaneja, A. Biological control using invertebrates and microorganisms: Plenty of new opportunities. Biocontrol 2017, 63, 39–59. [Google Scholar] [CrossRef]
- Wallis, C.M. Nutritional niche overlap analysis as a method to identify potential biocontrol fungi against trunk pathogens. Biocontrol 2021, 66, 559–571. [Google Scholar] [CrossRef]
- Calvo, H.; Roudet, J.; Gracia, A.P.; Venturini, M.E.; Novello, V.; Fermaud, M. Comparison of efficacy and modes of action of two high-potential biocontrol Bacillus strains and commercial biocontrol products against Botrytis cinerea in table grapes. Oeno One 2021, 55, 228–243. [Google Scholar] [CrossRef]
- Calvo, H.; Mendiara, I.; Arias, E.; Blanco, D.; Venturini, M.E. The role of iturin A from B. amyloliquefaciens BUZ-14 in the inhibition of the most common postharvest fruit rots. Food Microbiol. 2019, 82, 62–69. [Google Scholar] [CrossRef]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2022. [Google Scholar]
- Alfonzo, A.; Lo Piccolo, S.; Conigliaro, G.; Ventorino, V.; Burruano, S.; Moschetti, G. Antifungal peptides produced by Bacillus amyloliquefaciens AG1 active against grapevine fungal pathogens. Ann. Microbiol. 2012, 62, 1593–1599. [Google Scholar] [CrossRef]
- Brown, A.A.; Travadon, R.; Lawrence, D.P.; Torres, G.; Zhuang, G.; Baumgartner, K. Pruning-wound protectants for trunk-disease management in California table grapes. Crop Prot. 2021, 141, 105490. [Google Scholar] [CrossRef]
- Halleen, F.; Fourie, P.H.; Lombard, P.J. Protection of grapevine pruning wounds against Eutypa lata by biological and chemical methods. Enol. Vitic. 2010, 31, 125–132. [Google Scholar] [CrossRef]
- Trotel-Aziz, P.; Abou-Mansour, E.; Courteaux, B.; Rabenoelina, F.; Clement, C.; Fontaine, F.; Aziz, A. Bacillus subtilis PTA-271 counteracts Botryosphaeria dieback in grapevine, triggering immune responses and detoxification of fungal phytotoxins. Front. Plant Sci. 2019, 10, 25. [Google Scholar] [CrossRef] [PubMed]
- Rusin, C.; Cavalcanti, F.R.; de Lima, P.C.G.; Faria, C.; Almanca, M.A.K.; Botelho, R.V. Control of the fungi Lasiodiplodia theobromae, the causal agent of dieback, in cv. Syrah grapevines. Acta Sci.-Agron. 2021, 43, e44785. [Google Scholar] [CrossRef]
- Alfonzo, A.; Conigliaro, G.; Torta, L.; Burruano, S.; Moschetti, G. Antagonism of Bacillus subtilis strain AG1 against vine wood fungal pathogens. Phytopathol. Mediterr. 2009, 48, 155–158. [Google Scholar]
- Sebestyen, D.; Perez-Gonzalez, G.; Goodell, B. Antioxidants and iron chelators inhibit oxygen radical generation in fungal cultures of plant pathogenic fungi. Fungal Biol. 2022, 126, 480–487. [Google Scholar] [CrossRef]
- John, S.; Scott, E.S.; Wicks, T.J.; Hunt, J.S. Interactions between Eutypa lata and Trichoderma harzianum. Phytopathol. Mediterr. 2004, 43, 95–104. [Google Scholar]
- Di Marco, S.; Osti, F.; Cesari, A. Experiments on the control of esca by Trichoderma. Phytopathol. Mediterr. 2004, 43, 108–115. [Google Scholar]
- John, S.; Wicks, T.J.; Hunt, J.S.; Lorimer, M.F.; Oakey, H.; Scott, E.S. Protection of grapevine pruning wounds from infection by Eutypa lata using Trichoderma harzianum and Fusarium lateritium. Australas. Plant Pathol. 2005, 34, 569–575. [Google Scholar] [CrossRef]
- Halleen, F.; Fourie, P.H. An integrated strategy for the proactive management of grapevine trunk disease pathogen infections in grapevine nurseries. S. Afr. J. Enol. Vitic. 2016, 37, 104–114. [Google Scholar] [CrossRef]
- van Jaarsveld, W.J.; Halleen, F.; Bester, M.C.; Pierron, R.J.; Stempien, E.; Mostert, L. Investigation of Trichoderma species colonization of nursery grapevines for improved management of black foot disease. Pest Manag. Sci. 2020, 77, 397–405. [Google Scholar] [CrossRef]
- Pintos, C.; Redondo, V.; Aguín, O.; Chaves, M.; Rial, C.; Mansilla, J. Evaluation of Trichoderma atroviride as biocontrol agent against five Botryosphaeriaceae grapevine trunk pathogens. Phytopathol. Mediterr. 2012, 51, 450. [Google Scholar]
- Reis, P.; Mondello, V.; Diniz, I.; Alves, A.; Rego, C.; Fontaine, F. Effect of the combined treatments with LC2017 and Trichoderma atroviride strain I-1237 on disease development and defense responses in vines infected by Lasiodiplodia theobromae. Agronomy 2022, 12, 996. [Google Scholar] [CrossRef]
- Berbegal, M.; Ramon-Albalat, A.; Leon, M.; Armengol, J. Evaluation of long-term protection from nursery to vineyard provided by Trichoderma atroviride SC1 against fungal grapevine trunk pathogens. Pest Manag. Sci. 2020, 76, 967–977. [Google Scholar] [CrossRef] [PubMed]
- Mutawila, C.; Halleen, F.; Mostert, L. Optimisation of time of application of Trichoderma biocontrol agents for protection of grapevine pruning wounds. Aust. J. Grape Wine Res. 2016, 22, 279–287. [Google Scholar] [CrossRef]
- Mutawila, C.; Fourie, P.H.; Halleen, F.; Mostert, L. Grapevine cultivar variation to pruning wound protection by Trichoderma species against trunk pathogens. Phytopathol. Mediterr. 2011, 50, S264–S276. [Google Scholar]
- Calvo, H.; Mendiara, I.; Arias, E.; Gracia, A.P.; Blanco, D.; Venturini, M.E. Antifungal activity of the volatile organic compounds produced by Bacillus velezensis strains against postharvest fungal pathogens. Postharvest Biol. Technol. 2020, 166, 111208. [Google Scholar] [CrossRef]
- Mutawila, C.; Vinale, F.; Halleen, F.; Lorito, M.; Mostert, L. Isolation, production and in vitro effects of the major secondary metabolite produced by Trichoderma species used for the control of grapevine trunk diseases. Plant Pathol. 2016, 65, 104–113. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Treatment | Concentration | Pathogen | Number of Replicates |
|---|---|---|---|
| Native Trichoderma harzianum | 1 × 107 conidia·mL−1 | D. seriata | 20 |
| N. parvum | 20 | ||
| Negative control | 4 | ||
| Trichoderma harzianum T-22 (Trianum-P®) | 1 × 107 conidia·mL−1 | D. seriata | 20 |
| N. parvum | 20 | ||
| Negative control | 4 | ||
| Trichoderma atroviride I-1237 (Esquive®) | 1 × 107 conidia·mL−1 | D. seriata | 20 |
| N. parvum | 20 | ||
| Negative control | 4 | ||
| Bacillus velezensis BUZ-14 | 1 × 107 CFU·mL−1 | D. seriata | 20 |
| N. parvum | 20 | ||
| Negative control | 4 | ||
| Bacillus subtilis BS03 (FUNGISEI®) | 1 × 107 CFU·mL−1 | D. seriata | 20 |
| N. parvum | 20 | ||
| Negative control | 4 | ||
| - | - | D. seriata positive control | 6 |
| N. parvum positive control | 6 |
| Treatment | Mean of Ranks | Groups | |||
|---|---|---|---|---|---|
| Negative control | 17.000 | A | |||
| B. velezensis (BUZ-14) | 360.994 | B | |||
| T. atroviride (I-1237) | 372.791 | B | |||
| T. harzianum (native) | 458.920 | C | |||
| T. harzianum (T-22) | 487.763 | C | |||
| B. subtilis (BS03) | 533.659 | C | D | ||
| N. parvum positive control | 623.578 | D | |||
| Treatment | Mean of Ranks | Groups | |||
|---|---|---|---|---|---|
| Negative control | 12.500 | A | |||
| B. velezensis (BUZ-14) | 297.784 | B | |||
| T. atroviride (I-1237) | 399.234 | C | |||
| T. harzianum (native) | 438.197 | C | |||
| T. harzianum (T-22) | 457.480 | C | |||
| D. seriata positive control | 470.813 | C | D | ||
| B. subtilis (BS03) | 548.569 | D | |||
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Langa-Lomba, N.; González-García, V.; Venturini-Crespo, M.E.; Casanova-Gascón, J.; Barriuso-Vargas, J.J.; Martín-Ramos, P. Comparison of the Efficacy of Trichoderma and Bacillus Strains and Commercial Biocontrol Products against Grapevine Botryosphaeria Dieback Pathogens. Agronomy 2023, 13, 533. https://doi.org/10.3390/agronomy13020533
Langa-Lomba N, González-García V, Venturini-Crespo ME, Casanova-Gascón J, Barriuso-Vargas JJ, Martín-Ramos P. Comparison of the Efficacy of Trichoderma and Bacillus Strains and Commercial Biocontrol Products against Grapevine Botryosphaeria Dieback Pathogens. Agronomy. 2023; 13(2):533. https://doi.org/10.3390/agronomy13020533
Chicago/Turabian StyleLanga-Lomba, Natalia, Vicente González-García, M. Eugenia Venturini-Crespo, José Casanova-Gascón, Juan J. Barriuso-Vargas, and Pablo Martín-Ramos. 2023. "Comparison of the Efficacy of Trichoderma and Bacillus Strains and Commercial Biocontrol Products against Grapevine Botryosphaeria Dieback Pathogens" Agronomy 13, no. 2: 533. https://doi.org/10.3390/agronomy13020533
APA StyleLanga-Lomba, N., González-García, V., Venturini-Crespo, M. E., Casanova-Gascón, J., Barriuso-Vargas, J. J., & Martín-Ramos, P. (2023). Comparison of the Efficacy of Trichoderma and Bacillus Strains and Commercial Biocontrol Products against Grapevine Botryosphaeria Dieback Pathogens. Agronomy, 13(2), 533. https://doi.org/10.3390/agronomy13020533