
Physiological and Molecular Responses of Wheat to Low Light Intensity

 ,
,  ,
,  , ,
, ,  and
and
Abstract
1. Introduction
2. Materials and Methods
2.1. Experimental Design and Field Management
2.2. Determination of Chlorophyll Content and Dry Matter Accumulation
2.3. Determination of Chlorophyll Fluorescence Parameters
2.4. Label-Free Quantitative Proteomic Analysis
2.5. Statistical Analysis
3. Results
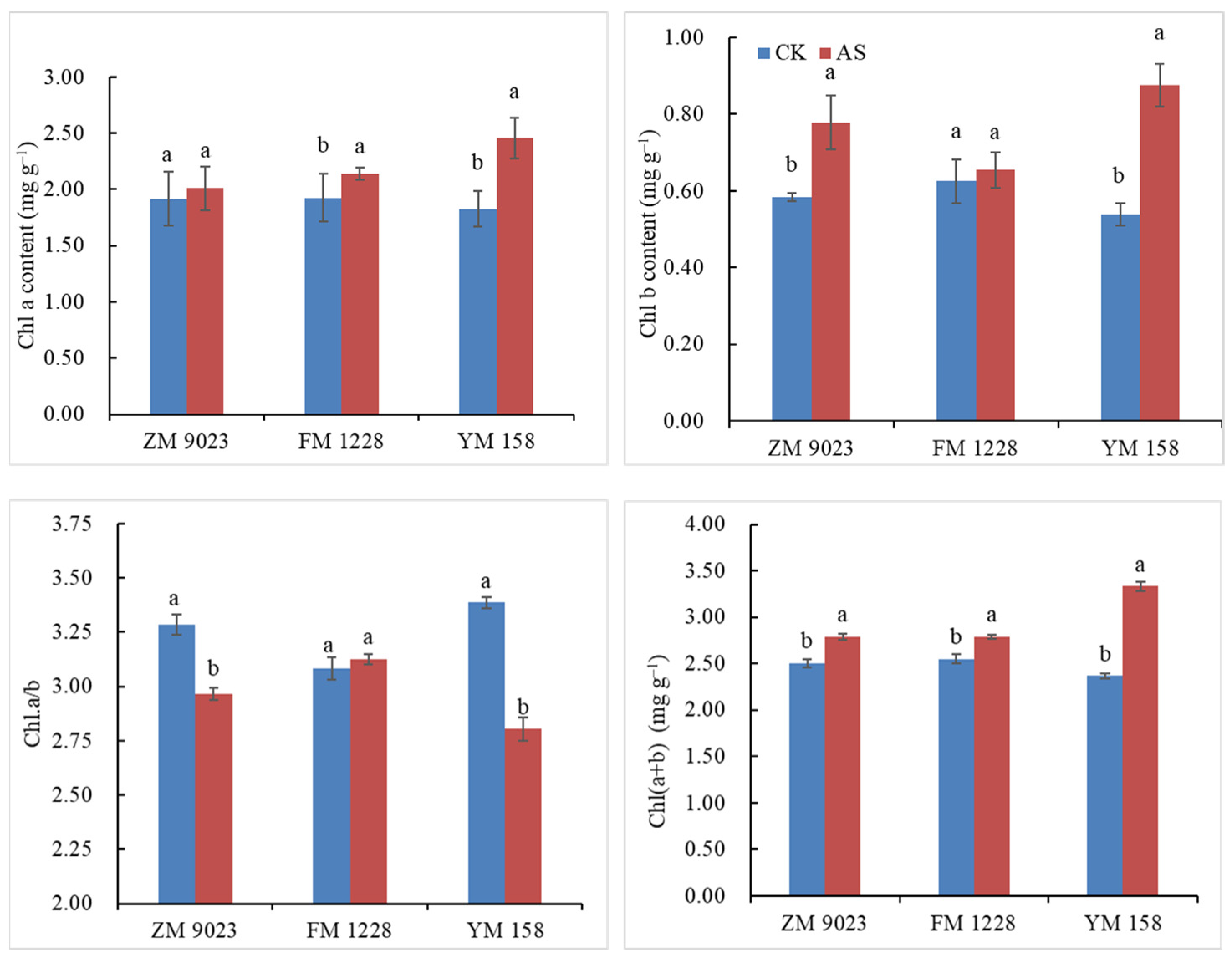
3.1. Chlorophyll Content
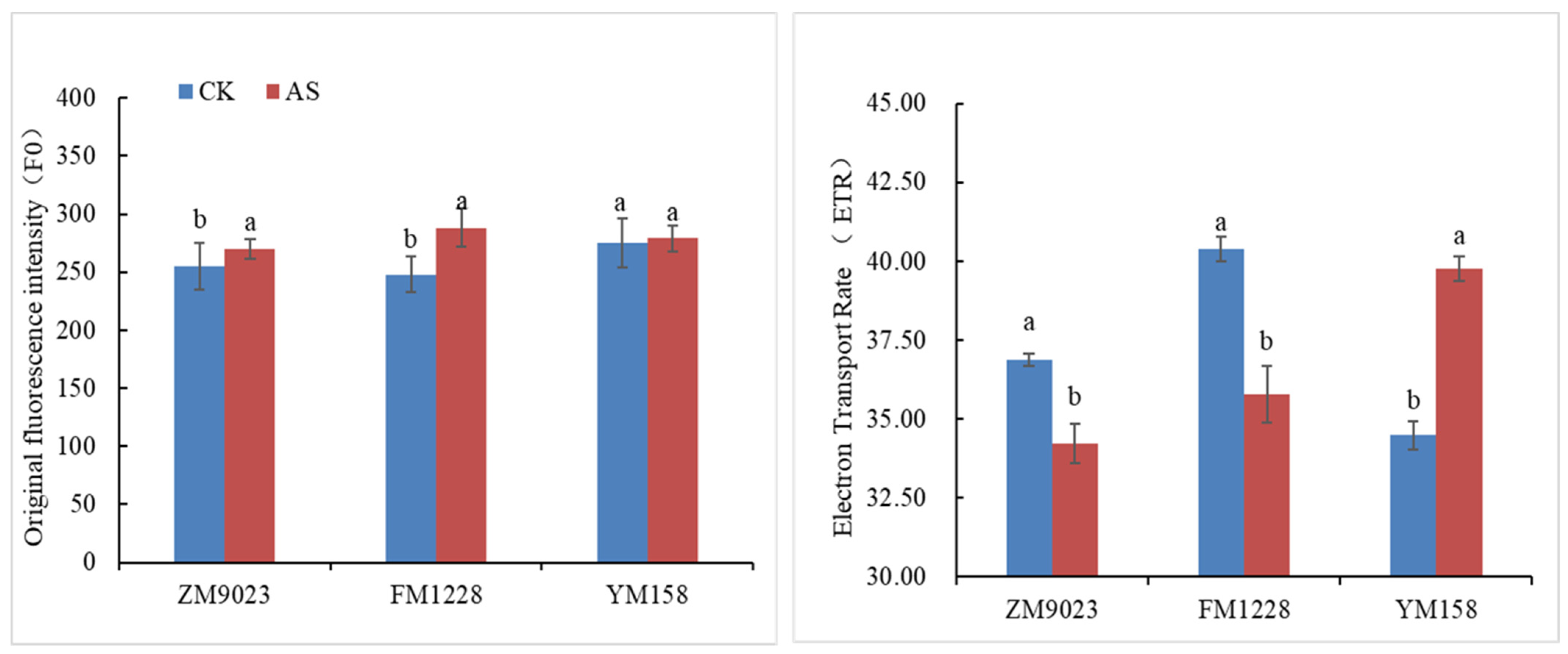
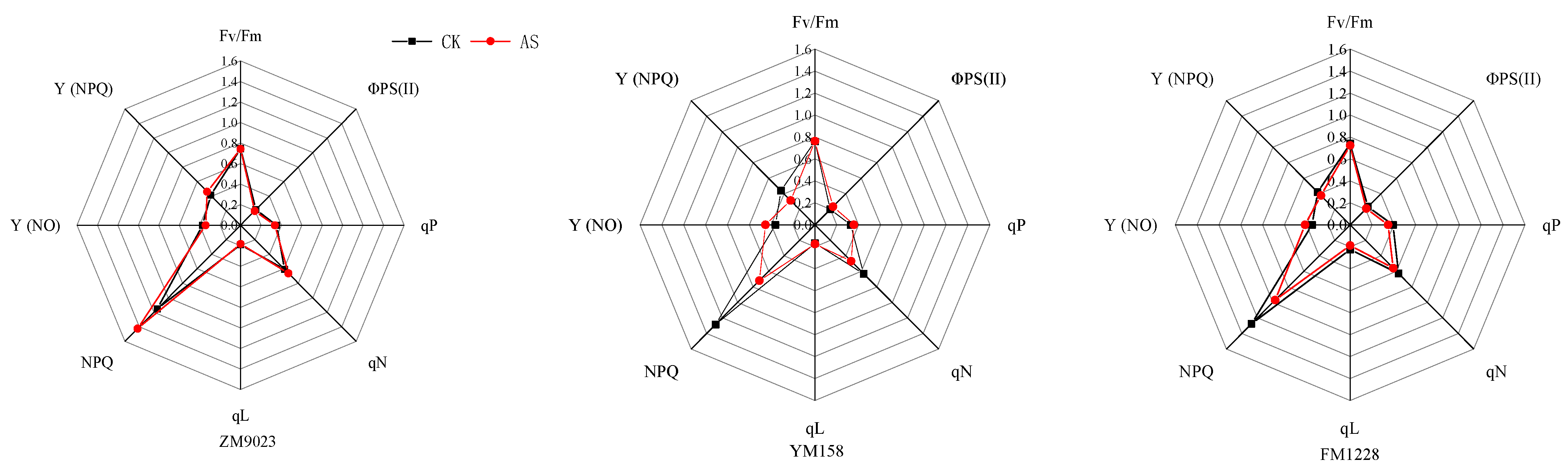
3.2. Chlorophyll Fluorescence Parameters
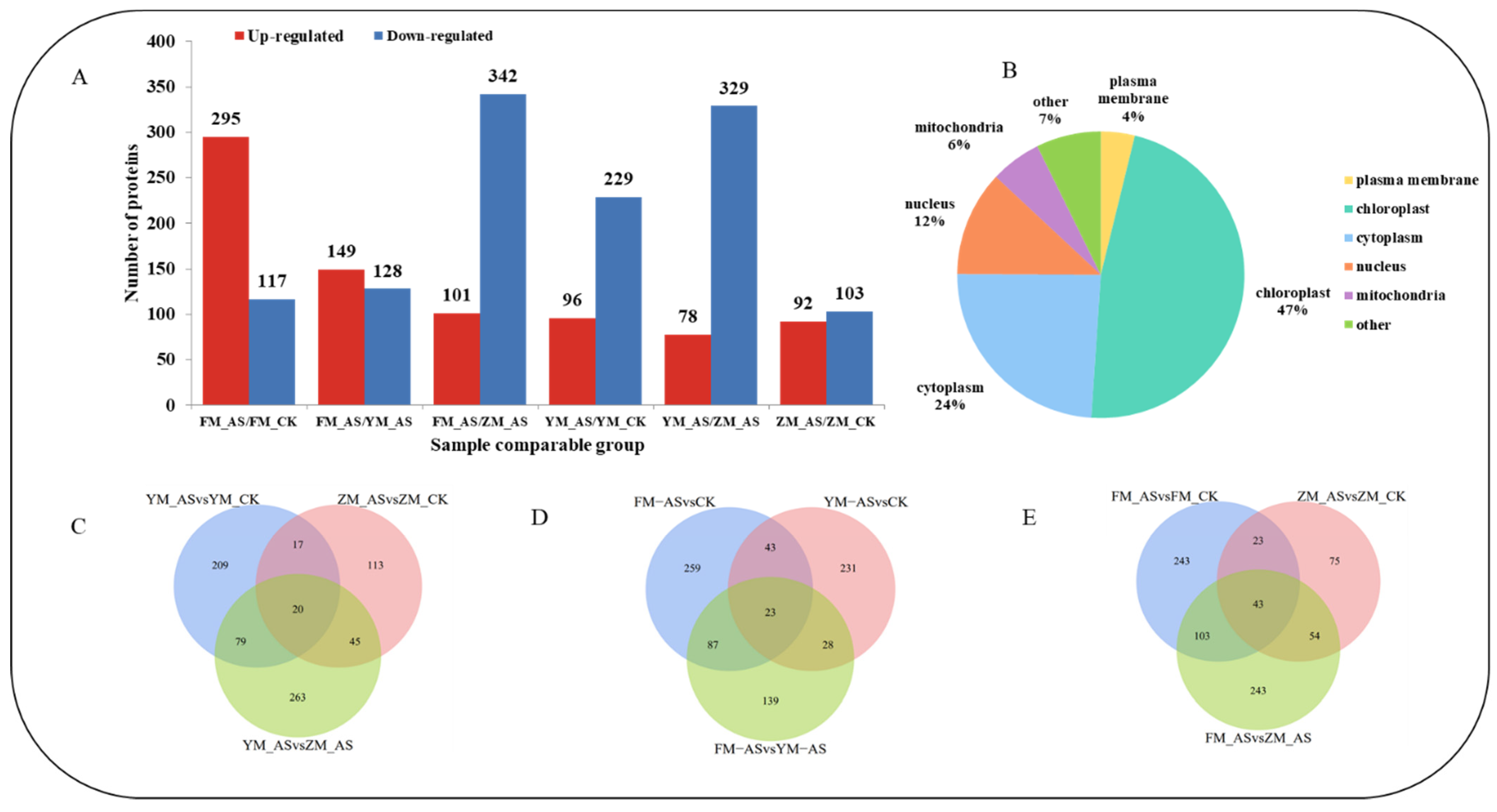
3.3. Label-Free Quantitative Proteomics Analysis in Wheat Leaves
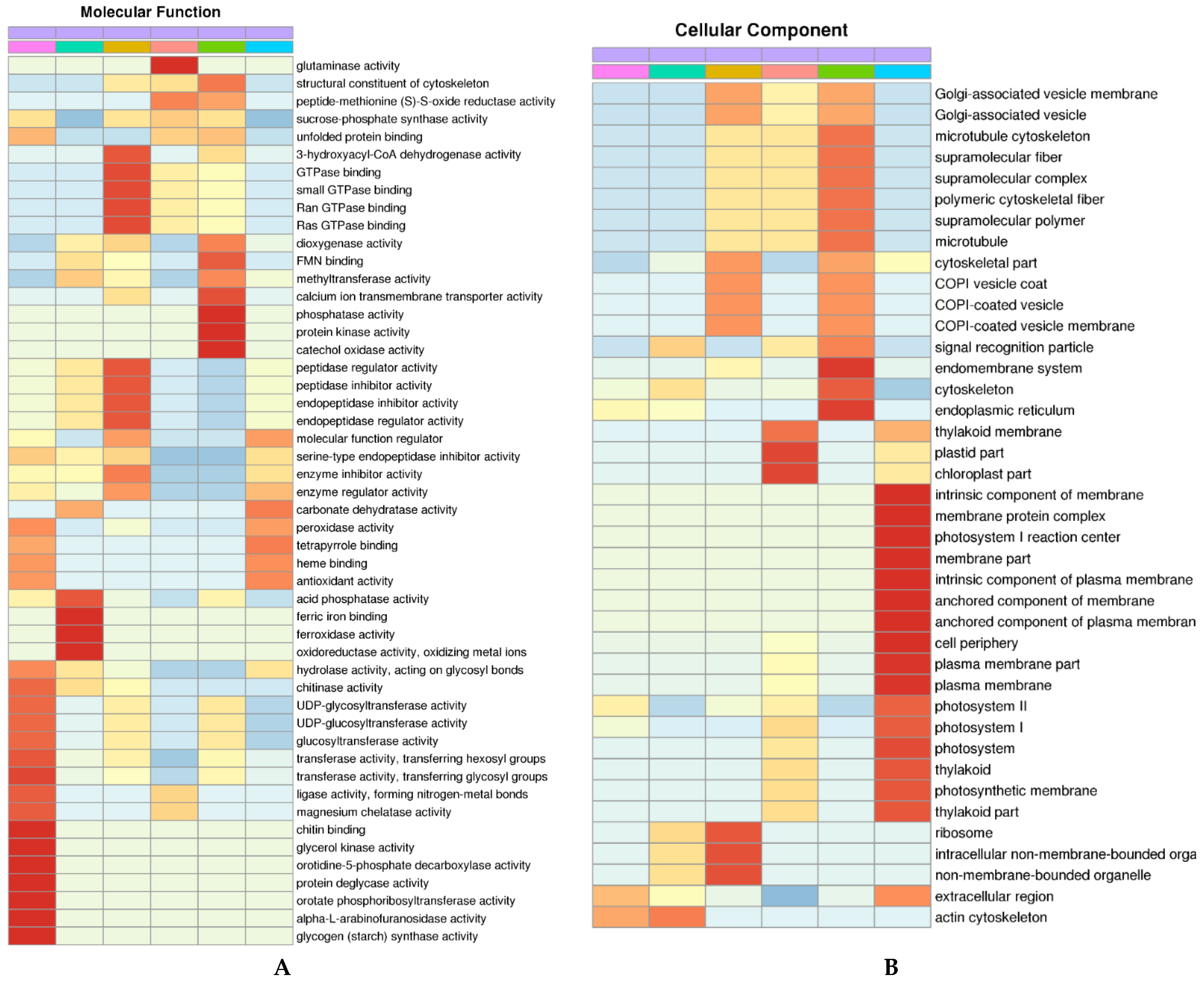
3.4. Gene Ontology Classification of DAPs
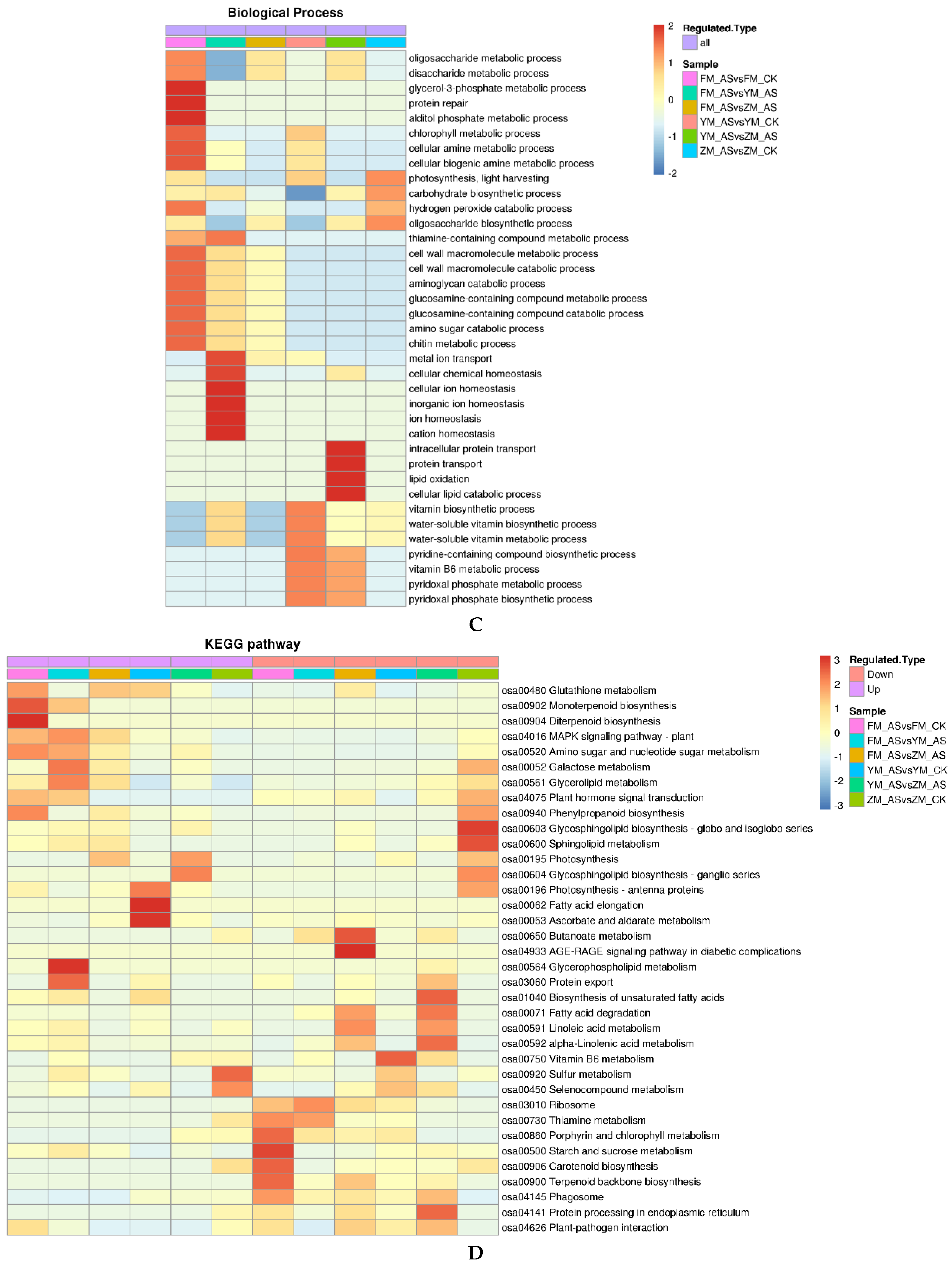
3.5. KEGG Pathway Analysis of DAPs
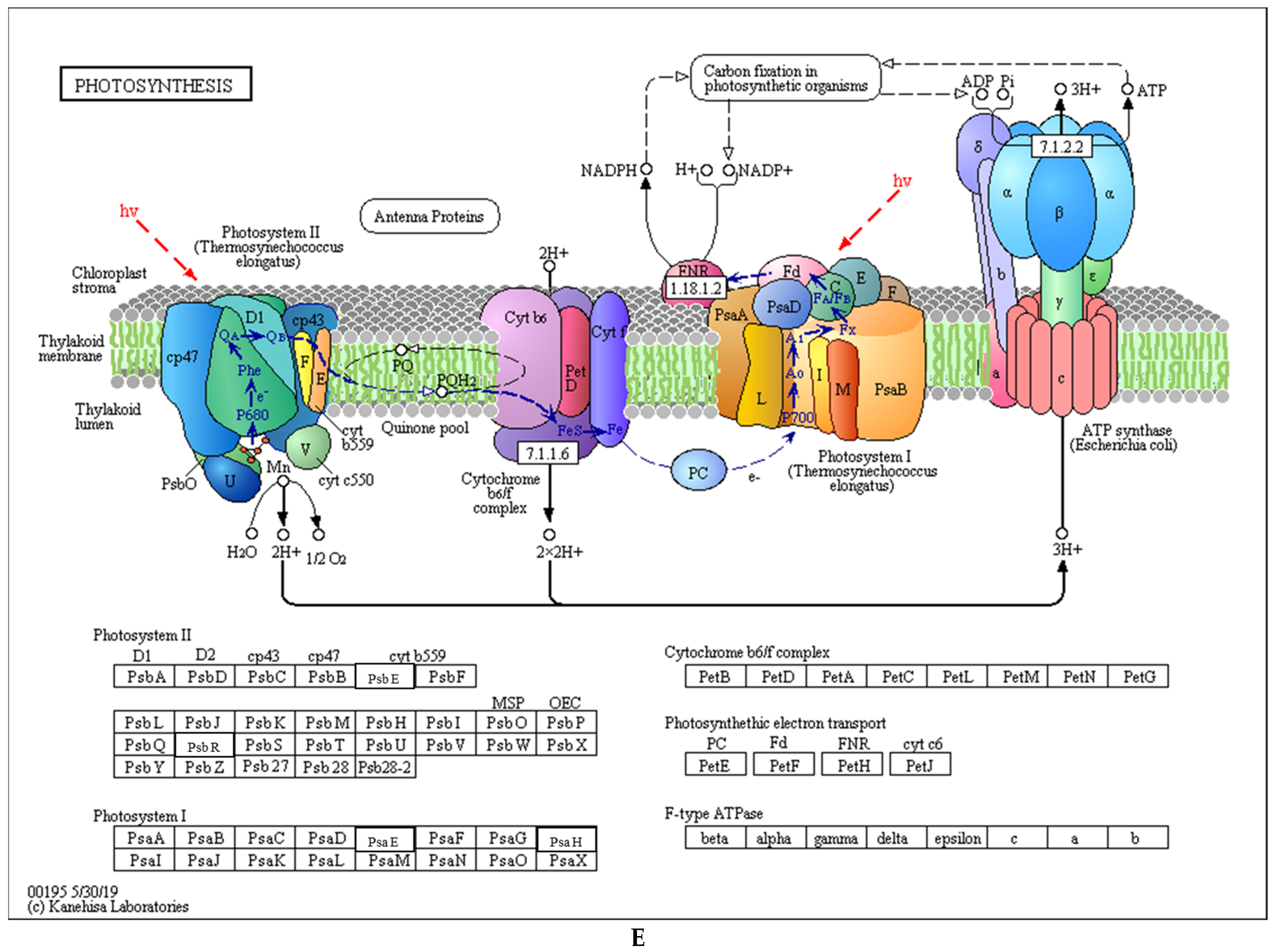
3.6. Differential Analysis of Wheat Photosynthetic Proteins under Low Light
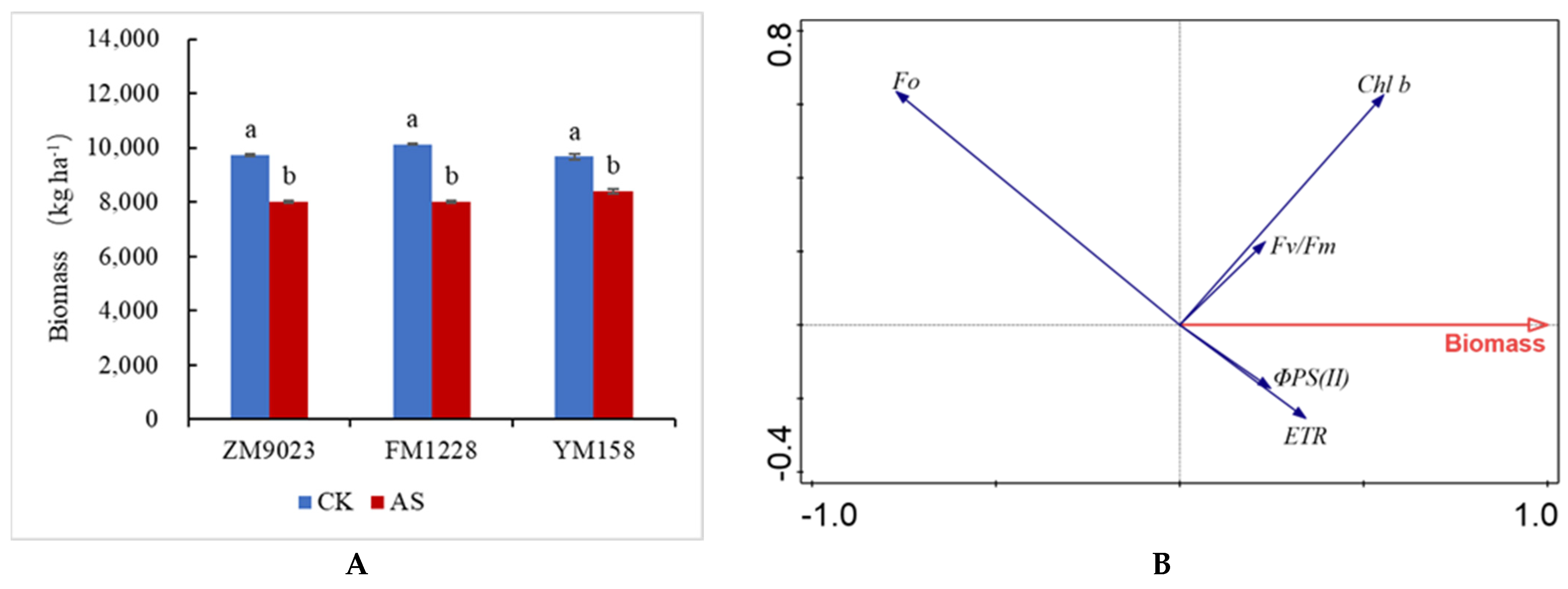
3.7. Effects of Shading on Dry Matter Accumulation
4. Discussion
4.1. Effects of Low Light on Dry Matter Accumulation and Leaf Physiology in Different Wheat Varieties
4.2. Responses of Chlorophyll Fluorescence Parameters to Low Light
4.3. Effects of Low Light on Proteomics across Genotypes
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Liu, K.; Harrison, M.T.; Wang, B.; Yang, R.; Yan, H.; Zou, J.; Liu, D.L.; Meinke, H.; Tian, X.; Ma, S.; et al. Designing high-yielding wheat crops under late sowing: A case study in southern China. Agron. Sustain. Dev. 2022, 42, 29. [Google Scholar] [CrossRef]
- Yan, H.; Harrison, M.T.; Liu, K.; Wang, B.; Feng, P.; Fahad, S.; Meinke, H.; Yang, R.; Liu, D.L.; Archontoulis, S.; et al. Crop traits enabling yield gains under more frequent extreme climatic events. Sci. Total Environ. 2022, 808, 152170. [Google Scholar] [CrossRef]
- Harrison, M.T. Climate change benefits negated by extreme heat. Nat. Food 2021, 2, 855–856. [Google Scholar] [CrossRef]
- Cai, J.; Jiang, D. The Effect of Climate Change on Winter Wheat Production in China. J. Agro-Environ. Sci. 2011, 30, 1726–1733. [Google Scholar]
- Liu, K.; Harrison, M.; Yan, H.; Liu, D.L.; Meinke, H.; Hoogenboom, G.; Wang, B.; Peng, B.; Guan, K.; Jaegermeyr, J.; et al. Silver lining to a climate crisis in multiple prospects for alleviating crop waterlogging under future climates. Res. Sq. 2022. [Google Scholar] [CrossRef]
- Harrison, M.T.; Evans, J.R.; Dove, H.; Moore, A.D. Recovery dynamics of rainfed winter wheat after livestock grazing 2. Light interception, radiation-use efficiency and dry-matter partitioning. Crop Pasture Sci. 2011, 62, 960–971. [Google Scholar] [CrossRef]
- Zhang, C.; Chu, H.; Chen, G.; Shi, D.; Zuo, M.; Wang, J.; Lu, C.; Wang, P.; Chen, L. Photosynthetic and biochemical activities in flag leaves of a newly developed superhigh-yield hybrid rice (Oryza sativa) and its parents during the reproductive stage. J. Plant Res. 2007, 120, 209–217. [Google Scholar] [CrossRef]
- Chen, Y.; Guerschman, J.; Shendryk, Y.; Henry, D.; Harrison, M.T. Estimating pasture biomass using sentinel-2 imagery and machine learning. Remote Sens. 2021, 13, 603. [Google Scholar] [CrossRef]
- Dong, B.; Yang, H.; Liu, H.; Qiao, Y.; Zhang, M.; Wang, Y.; Xie, Z.; Liu, M. Effects of Shading Stress on Grain Number, Yield, and Photosynthesis during Early Reproductive Growth in Wheat. Crop Sci. 2019, 59, 363–378. [Google Scholar] [CrossRef]
- Walker, B.J.; Kramer, D.M.; Fisher, N.; Fu, X. Flexibility in the Energy Balancing Network of Photosynthesis Enables Safe Operation under Changing Environmental Conditions. Plants 2020, 9, 301. [Google Scholar] [CrossRef]
- Ibrahim, A.; Harrison, M.T.; Meinke, H.; Zhou, M. Examining the yield potential of barley near-isogenic lines using a genotype by environment by management analysis. Eur. J. Agron. 2019, 105, 41–51. [Google Scholar] [CrossRef]
- Fan, Y.; Chen, J.; Wang, Z.; Tan, T.; Li, S.; Li, J.; Wang, B.; Zhang, J.; Cheng, Y.; Wu, X.; et al. Soybean (Glycine max L. Merr.) seedlings response to shading: Leaf structure, photosynthesis and proteomic analysis. BMC Plant Biol. 2019, 19, 34. [Google Scholar] [CrossRef] [PubMed]
- Leong, T.; Anderson, J. Adaptation of the thylakoid membranes of pea chloroplasts to light intensities. I. Study on the distribution of chlorophyll-protein complexes. Photosynth. Res. 1984, 5, 105–115. [Google Scholar] [CrossRef] [PubMed]
- Legris, M.; Ince, Y.; Fankhauser, C. Molecular mechanisms underlying phytochrome-controlled morphogenesis in plants. Nat. Commun. 2019, 10, 5219. [Google Scholar] [CrossRef] [PubMed]
- Poorter, H.; Niinemets, Ü.; Ntagkas, N.; Siebenkäs, A.; Mäenpää, M.; Matsubara, S.; Pons, T. A meta-analysis of plant responses to light intensity for 70 traits ranging from molecules to whole plant performance. New Phytol. 2019, 223, 1073–1105. [Google Scholar] [CrossRef] [PubMed]
- Yu, X.; Ming, X.; Xiong, M.; Zhang, C.; Yue, L.; Yang, L.; Fan, C. Partial shade improved the photosynthetic capacity and polysaccharide accumulation of the medicinal plant Bletilla ochracea Schltr. Photosynthetica 2022, 60, 12–22. [Google Scholar] [CrossRef]
- Kondo, T.; Gordon, J.B.; Pinnola, A.; Dall’Osto, L.; Bassi, R.; Schlau-Cohen, G.S. Microsecond and millisecond dynamics in the photosynthetic protein LHCSR1 observed by single-molecule correlation spectroscopy. Proc. Natl. Acad. Sci. USA 2019, 116, 11247–11252. [Google Scholar] [CrossRef] [PubMed]
- Guidi, L.; Lo Piccolo, E.; Landi, M. Chlorophyll fluorescence, photoinhibition and abiotic stress: Does it make any difference the fact to be a C3 or C4 species? Front. Plant Sci. 2019, 10, 174. [Google Scholar] [CrossRef]
- Rogowski, P.; Wasilewska-Dębowska, W.; Krupnik, T.; Drożak, A.; Zienkiewicz, M.; Krysiak, M.; Romanowska, E. Photosynthesis and organization of maize mesophyll and bundle sheath thylakoids of plants grown in various light intensities. Environ. Exp. Bot. 2019, 162, 72–86. [Google Scholar] [CrossRef]
- Arenas-Corraliza, M.; Rolo, V.; López-Díaz, M.; Moreno, G. Wheat and barley can increase grain yield in shade through acclimation of physiological and morphological traits in Mediterranean conditions. Sci. Rep. 2019, 9, 9547. [Google Scholar] [CrossRef]
- Arnon, D. Copper enzymes in isolated chloroplasts. Polyphenoloxidase in Beta vulgaris. Plant Physiol. 1949, 24, 1–15. [Google Scholar] [CrossRef] [PubMed]
- Gong, X.; Liu, C.; Ferdinand, U.; Dang, K.; Zhao, G.; Yang, P.; Feng, B. Effect of intercropping on leaf senescence related to physiological metabolism in proso millet (Panicum miliaceum L.). Photosynthetica 2019, 57, 993–1006. [Google Scholar] [CrossRef]
- Yun, F.; Liu, G.; Shi, H.; Song, J. Effects of Light and Nitrogen Interaction on Photosynthesis and Chlorophyll Fluorescence Characteristics in Flue-Cured Tobacco. Sci. Agric. Sin. 2010, 43, 932–941. [Google Scholar]
- Cheng, J.; Duan, W.; Tang, X.; Zhang, Y.; Li, B.; Wang, Y.; Yang, C.; Song, Z.; Wang, L.; Yang, J.; et al. Low sink demand caused net photosynthetic rate decrease is closely related to the irrecoverable damage of oxygen-releasing complex and electron receptor in peach trees. J. Plant Physiol. 2021, 266, 153510. [Google Scholar] [CrossRef]
- Meng, Z.; Lu, T.; Zhang, G.; Qi, M.; Tang, W.; Li, L.; Liu, Y.; Li, T. Photosystem inhibition and protection in tomato leaves under low light. Sci. Hortic. 2017, 217, 145–155. [Google Scholar] [CrossRef]
- Samaniego-Gámez, B.Y.; Garruña, R.; Tun-Suárez, J.; Kantun-Can, J.; Reyes-Ramírez, A.; Cervantes-Díaz, L. Bacillus spp. inoculation improves photosystem II efficiency and enhances photosynthesis in pepper plants. Chil. J. Agric. Res. 2016, 76, 409–416. [Google Scholar] [CrossRef]
- He, Z.; Tang, R.; Li, M.; Jin, M.; Xin, C.; Liu, J.; Hong, W. Response of photosynthesis and Chlorophyll fluorescence parameters of castanopsis kawakamii seedlings to forest gaps. Forests 2019, 11, 21. [Google Scholar] [CrossRef]
- Jia, M.; Li, D.; Colombo, R.; Wang, Y.; Wang, X.; Cheng, T.; Zhu, Y.; Yao, X.; Xu, C.; Ouer, G.; et al. Quantifying Chlorophyll Fluorescence Parameters from Hyperspectral Reflectance at the Leaf Scale under Various Nitrogen Treatment Regimes in Winter Wheat. Remote Sens. 2019, 11, 2838. [Google Scholar] [CrossRef]
- Elkins, C.; van Iersel, M.W. Longer photoperiods with the same daily light integral increase daily electron transport through photosystem II in lettuce. Plants 2020, 9, 1172. [Google Scholar] [CrossRef]
- Osmond, C.B.; Chow, W.S.; Robinson, S.A. Inhibition of non-photochemical quenching increases functional absorption cross-section of photosystem II as excitation from closed reaction centres is transferred to open centres, facilitating earlier light saturation of photosynthetic electron transport. Funct. Plant Biol. 2021, 49, 463–482. [Google Scholar] [CrossRef]
- Mu, H.; Jiang, D.; Wollenweber, B.; Dai, T.; Jing, Q.; Cao, W. Long-term low radiation decreases leaf photosynthesis, photochemical efficiency and grain yield in winter wheat. J. Agron. Crop Sci. 2010, 196, 38–47. [Google Scholar] [CrossRef]
- Zivcak, M.; Brestic, M.; Kalaji, H.M. Photosynthetic responses of sun-and shade-grown barley leaves to high light: Is the lower PSII connectivity in shade leaves associated with protection against excess of light? Photosynth. Res. 2014, 119, 339–354. [Google Scholar] [CrossRef] [PubMed]
- Weng, H.; Zeng, Y.; Cen, H.; He, M.; Meng, Y.; Liu, Y.; Wan, L.; Xu, H.; Li, H.; Fang, H.; et al. Characterization and detection of leaf photosynthetic response to citrus Huanglongbing from cool to hot seasons in two orchards. Trans. ASABE 2020, 63, 501–512. [Google Scholar] [CrossRef]
- Wang, Y.; Zhang, Z.; Liang, Y.; Han, Y.; Han, Y.; Tan, J. High potassium application rate increased grain yield of shading-stressed winter wheat by improving photosynthesis and photosynthate translocation. Front. Plant Sci. 2020, 11, 134. [Google Scholar] [CrossRef] [PubMed]
- Sadak, M.H. Physiological role of signal molecules in improving plant tolerance under abiotic stress. Int. J. Chem Tech. Res. 2016, 9, 46–60. [Google Scholar]
- Ragaey, M.M.; Sadak, M.S.; Dawood, M.F.A.; Mousa, N.H.S.; Hanafy, R.S.; Latef, A.A.H.A. Role of Signaling Molecules Sodium Nitroprusside and Arginine in Alleviating Salt-Induced Oxidative Stress in Wheat. Plants 2022, 11, 1786. [Google Scholar] [CrossRef]
- Liu, Q.; Wu, X.; Chen, B.; Ma, J.; Gao, J. Effects of low light on agronomic and physiological characteristics of rice including grain yield and quality. Rice Sci. 2014, 21, 243–251. [Google Scholar] [CrossRef]
- Xie, X.; Cheng, H.; Hou, C.; Ren, M. Integration of Light and Auxin Signaling in Shade Plants: From Mechanisms to Opportunities in Urban Agriculture. Int. J. Mol. Sci. 2022, 23, 3422. [Google Scholar] [CrossRef]
- Van, K.; Snel, J. The use of chlorophyll fluorescence nomenclature in plant stress physiology. Photosynth. Res. 1990, 25, 147–150. [Google Scholar] [CrossRef]
- Xu, Y.; Yang, M.; Cheng, F.; Liu, S.; Liang, Y. Effects of LED photoperiods and light qualities on in vitro growth and chlorophyll fluorescence of Cunninghamia lanceolata. BMC Plant Biol. 2020, 20, 269. [Google Scholar] [CrossRef]
- Schreiber, U. Pulse-amplitude-modulation (PAM) fluorometry and saturation pulse method: An overview. In Chlorophyll a Fluorescence; Springer: Dordrecht, The Netherlands, 2004; pp. 279–319. [Google Scholar] [CrossRef]
- Yamori, W.; Takahashi, S.; Makino, A.; Price, G.D.; Badger, M.R.; von Caemmerer, S. The roles of ATP synthase and the cytochrome b 6/f complexes in limiting chloroplast electron transport and determining photosynthetic capacity. Plant Physiol. 2011, 155, 956–962. [Google Scholar] [CrossRef] [PubMed]
- Klughammer, C.; Schreiber, U. Complementary PS II quantum yields calculated from simple fluorescence parameters measured by PAM fluorometry and the Saturation Pulse method. PAM Appl. Notes 2008, 1, 201–247. [Google Scholar]
- Li, X.; Wei, F.; Zeng, X. Advances in chlorophyll fluorescence analysis and its uses. Acta Bot. Boreali-Occident. Sin. 2006, 26, 2186–2196. [Google Scholar]
- Gao, J.; Liu, Z.; Zhao, B.; Liu, P.; Zhang, J. Physiological and comparative proteomic analysis provides new insights into the effects of shade stress in maize (Zea mays L.). BMC Plant Biol. 2020, 20, 60. [Google Scholar] [CrossRef] [PubMed]
- Geigenberger, P.; Fernie, A.R. Metabolic control of redox and redox control of metabolism in plants. Antioxid. Redox Signal. 2014, 21, 1389–1421. [Google Scholar] [CrossRef] [PubMed]
- Papenbrock, J.; Mock, H.P.; Tanaka, R.; Kruse, E.; Grimm, B. Role of magnesium chelatase activity in the early steps of the tetrapyrrole biosynthetic pathway. Plant Physiol. 2000, 122, 1161–1170. [Google Scholar] [CrossRef]
- Shen, Y.; Wang, X.; Wu, F.; Du, S.; Cao, Z.; Shang, Y.; Wang, X.; Peng, C.; Yu, X.; Zhu, S.; et al. The Mg-chelatase H subunit is an abscisic acid receptor. Nature 2006, 443, 823–826. [Google Scholar] [CrossRef]
- Kobayashi, K.; Mochizuki, N.; Yoshimura, N.; Motohashi, K.; Hisabori, T.; Masuda, T. Functional analysis of Arabidopsis thaliana isoforms of the Mg-chelatase CHLI subunit. Photochem. Photobiol. Sci. 2008, 7, 1188–1195. [Google Scholar] [CrossRef]
- Yuan, Z.; Ni, X.; Arif, M.; Dong, Z.; Zhang, L.; Tan, X.; Li, J.; Li, C. Transcriptomic Analysis of the Photosynthetic, Respiration, and Aerenchyma Adaptation Strategies in Bermudagrass (Cynodon dactylon) under Different Submergence Stress. Int. J. Mol. Sci. 2021, 22, 7905. [Google Scholar] [CrossRef]
- Ma, N.; Ma, X.; Li, A.; Cao, X.; Kong, L. Cloning and expression analysis of wheat pheophorbide a oxygenase gene TaPaO. Plant Mol. Biol. Report. 2012, 30, 1237–1245. [Google Scholar] [CrossRef]
- Aghdam, M.; Razavi, F. Octapeptide NOP-1 treatment delays yellowing in broccoli floret during low temperature storage. Postharvest Biol. Technol. 2021, 180, 111628. [Google Scholar] [CrossRef]
- Yang, F.; Feng, L.; Liu, Q.; Wu, X.; Fan, Y.; Raza, M.; Cheng, Y.; Chen, J.; Wang, X.; Yong, T.; et al. Effect of interactions between light intensity and red-to-far-red ratio on the photosynthesis of soybean leaves under shade condition. Environ. Exp. Bot. 2018, 150, 79–87. [Google Scholar] [CrossRef]
- Nugent, J. Oxygenic photosynthesis: Electron transfer in photosystem I and photosystem II. Eur. J. Biochem. 1996, 237, 519–531. [Google Scholar] [CrossRef] [PubMed]
- Fuciman, M.; Enriquez, M.; Polivka, T.; Dall’Osto, L.; Bassi, R.; Frank, H.A. Role of xanthophylls in light harvesting in green plants: A spectroscopic investigation of mutant LHCII and Lhcb pigment–protein complexes. J. Phys. Chem. B 2012, 116, 3834–3849. [Google Scholar] [CrossRef] [PubMed]
- Oh, S.; Montgomery, B. Mesophyll-specific phytochromes impact chlorophyll light-harvesting complexes (LHCs) and non-photochemical quenching. Plant Signal. Behav. 2019, 14, 1609857. [Google Scholar] [CrossRef]
- Lv, Y.; Li, Y.; Liu, X.; Xu, K. Photochemistry and proteomics of ginger (Zingiber officinale Roscoe) under drought and shading. Plant Physiol. Biochem. 2020, 151, 188–196. [Google Scholar] [CrossRef]
- Ganeteg, U.; Kulheim, C.; Andersson, J.; Jansson, S. Is each light-harvesting complex protein important for plant fitness? Plant Physiol. 2004, 134, 502–509. [Google Scholar] [CrossRef]
- Knoppová, J.; Sobotka, R.; Yu, J.; Bečková, M.; Pilný, J.; Trinugroho, J.P.; Csefalvay, L.; Bína, D.; Nixon, P.J.; Komenda, J. Assembly of D1/D2 complexes of photosystem II: Binding of pigments and a network of auxiliary proteins. Plant Physiol. 2022, 189, 790–804. [Google Scholar] [CrossRef]
- Terentyev, V.V.; Shukshina, A.K.; Ashikhmin, A.A.; Tikhonov, K.G.; Shitov, A.V. The main structural and functional characteristics of photosystem-II-enriched membranes isolated from wild type and cia3 mutant Chlamydomonas reinhardtii. Life 2020, 10, 63. [Google Scholar] [CrossRef]
- Haldrup, A.; Simpson, D.; Scheller, H. Down-regulation of the PSI-F subunit of photosystem I (PSI) in Arabidopsis thaliana: The PSI-F subunit is essential for photoautotrophic growth and contributes to antenna function. J. Biol. Chem. 2000, 275, 31211–31218. [Google Scholar] [CrossRef]
- Ihalainen, J.A.; Jensen, P.E.; Haldrup, A.; van Stokkum, I.H.; van Grondelle, R.; Scheller, H.V.; Dekker, J.P. Pigment organization and energy transfer dynamics in isolated photosystem I (PSI) complexes from Arabidopsis thaliana depleted of the PSI-G, PSI-K, PSI-L, or PSI-N subunit. Biophys. J. 2002, 83, 2190–2201. [Google Scholar] [CrossRef] [PubMed]
- Jensen, P.; Haldrup, A.; Zhang, S.; Scheller, H.V. The PSI-O subunit of plant photosystem I is involved in balancing the excitation pressure between the two photosystems. J. Biol. Chem. 2004, 279, 24212–24217. [Google Scholar] [CrossRef] [PubMed]
- Baniulis, D.; Yamashita, E.; Zhang, H.; Hasan, S.S.; Cramer, W.A. Structure–function of the cytochrome b6f complex. Photochem. Photobiol. 2008, 84, 1349–1358. [Google Scholar] [CrossRef]
- Suorsa, M.; Sirpio, S.; Allahverdiyeva, Y.; Paakkarinen, V.; Mamedov, F.; Styring, S.; Aro, E.-M. PsbR, a missing link in the assembly of the oxygen-evolving complex of plant photosystem II. J. Biol. Chem. 2006, 281, 145–150. [Google Scholar] [CrossRef] [PubMed]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Analysis | Software/Method | Version/Website | Version/Website |
|---|---|---|---|
| Mass spectrum data analysis | MaxQuant | v.1.6.6.0 http://www.maxquant.org/ | 12 August 2019 |
| Motif analysis | MoMo | v.5.0.2 http://meme-suite.org/tools/momo | 12 August 2019 |
| GO comment | InterProScan | v.5.14-53.0 http://www.ebi.ac.uk/interpro/ | 12 August 2019 |
| Domain annotation | InterProScan | v.5.14-53.0 http://www.ebi.ac.uk/interpro/ | 12 August 2019 |
| KEGG comment | KAAS | v.2.0 http://www.genome.jp/kaas-bin/kaas_main | 12 August 2019 |
| KEGG Mapper | v.2.5 http://www.kegg.jp/kegg/mapper.html | 12 August 2019 | |
| Subcellular localization | Wolfpsort | v.0.2 http://www.genscript.com/psort/wolf_psort.html | 12 August 2019 |
| CELLO | v.2.5 http://cello.life.nctu.edu.tw/ | 12 August 2019 | |
| Enrichment analysis | Perl module | v.1.31 https://metacpan.org/pod/Text::NSP::Measures::2D::Fisher | 12 August 2019 |
| Clustering heat map | R Package pheatmap | v.2.0.3 https://cran.r-project.org/web/packages/cluster/ | 12 August 2019 |
| Protein Accession | FM 1228_AS/ FM 1228_CK Ratio | FM 1228_AS/ FM 1228_CK p Value | YM 158_AS/ YM 158_CK Ratio | YM 158_AS/ YM 158_CK p Value | ZM 9023_AS/ ZM 9023_CK Ratio | ZM 9023_AS/ ZM 9023_CK p Value | Subcellular Localization | KEGG KO No. | KEGG Gene | KEGG pathway |
|---|---|---|---|---|---|---|---|---|---|---|
| A0A3B6MJI8 | 0.682 | 0.000319 | 0.872 | 0.008401 | 1.074 | 0.015861 | chloroplast | K00231 | PPOX,;protoporphyrinogen/coproporphyrinogen III oxidase | osa00860 Porphyrin and chlorophyll metabolism |
| A0A3B6D8J7 | 0.174 | 0.002397 | 0.563 | 0.020221 | 3.052 | 0.001818 | chloroplast | K03403 | chlH, bchH; magnesium chelatase subunit H | osa00860 Porphyrin and chlorophyll metabolism |
| A0A3B6RQT6 | 0.656 | 8.17E-05 | 0.897 | 0.008597 | 1.27 | 0.047158 | chloroplast | K03405 | chlI, bchI; magnesium chelatase subunit I, Mg-protoporphyrin IX chelatase | osa00860 Porphyrin and chlorophyll metabolism |
| A0A3B6I0Y7 | 0.823 | 0.005938 | 0.803 | 0.000179 | 0.786 | 0.018519 | chloroplast | K13071 | PAO, ACD1; pheophorbide a oxygenase | osa00860 Porphyrin and chlorophyll metabolism |
| A0A3B6C6E5 | 1.258 | 0.106482 | 1.23 | 0.021164 | 0.814 | 0.002942 | chloroplast | K08915 | Lhcb4; light-harvesting complex II chlorophyll a/b binding protein 4 | osa00196 Photosynthesis-antenna proteins |
| A0A3B6DA68 | 1.409 | 0.149784 | 5.011 | 0.006844 | 0.878 | 0.43954 | chloroplast | K08908 | Lhca2; light-harvesting complex I chlorophyll a/b binding protein 2 | osa00196 Photosynthesis-antenna proteins |
| A0A1D5RS51 | 1.723 | 0.000722 | 1.562 | 0.004579 | 0.6 | 0.002898 | chloroplast | K08912 | Lhcb1; light-harvesting complex II chlorophyll a/b binding protein 1 | osa00196 Photosynthesis-antenna proteins |
| W5GFA4 | 1.247 | 0.017805 | 1.808 | 0.001079 | 0.752 | 0.02286 | chloroplast | K08909 | Lhca3; light-harvesting complex I chlorophyll a/b binding protein 3 | osa00196 Photosynthesis-antenna proteins |
| A0A3B6MT65 | 1.263 | 0.132322 | 0.937 | 0.66118 | 0.594 | 0.000504 | nucleus | K02707 | psbE; photosystem II cytochrome b559 subunit alpha | osa00195 Photosynthesis |
| A0A3B6N0T3 | 0.966 | 0.015923 | 0.929 | 0.043041 | 0.868 | 0.11284 | chloroplast | K08901 | psbQ; photosystem II oxygen-evolving enhancer protein 3 | osa00195 Photosynthesis |
| A0A3B6NWL9 | 0.745 | 0.005485 | 1.028 | 0.63812 | 1.349 | 0.014479 | chloroplast | K03541 | psbR; photosystem II 10kDa protein | osa00195 Photosynthesis |
| A0A3B6GQA5 | 1.184 | 0.014319 | 1.779 | 0.011498 | 0.682 | 0.019324 | chloroplast | K02699 | psaL; photosystem I subunit XI | osa00195 Photosynthesis |
| A0A3B6MXE7 | 1.021 | 0.77492 | 1.207 | 0.007018 | 0.833 | 0.136616 | chloroplast | K02694 | psaF; photosystem I subunit III | osa00195 Photosynthesis |
| P60162 | 0.957 | 0.43904 | 1.293 | 0.033939 | 0.746 | 0.051481 | cytoplasm | K02635 | petB; cytochrome b6 | osa00195 Photosynthesis |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Li, X.; Yang, R.; Li, L.; Liu, K.; Harrison, M.T.; Fahad, S.; Wei, M.; Yin, L.; Zhou, M.; Wang, X. Physiological and Molecular Responses of Wheat to Low Light Intensity. Agronomy 2023, 13, 272. https://doi.org/10.3390/agronomy13010272
Li X, Yang R, Li L, Liu K, Harrison MT, Fahad S, Wei M, Yin L, Zhou M, Wang X. Physiological and Molecular Responses of Wheat to Low Light Intensity. Agronomy. 2023; 13(1):272. https://doi.org/10.3390/agronomy13010272
Chicago/Turabian StyleLi, Xiu, Rui Yang, Liulong Li, Ke Liu, Matthew Tom Harrison, Shah Fahad, Mingmei Wei, Lijun Yin, Meixue Zhou, and Xiaoyan Wang. 2023. "Physiological and Molecular Responses of Wheat to Low Light Intensity" Agronomy 13, no. 1: 272. https://doi.org/10.3390/agronomy13010272
APA StyleLi, X., Yang, R., Li, L., Liu, K., Harrison, M. T., Fahad, S., Wei, M., Yin, L., Zhou, M., & Wang, X. (2023). Physiological and Molecular Responses of Wheat to Low Light Intensity. Agronomy, 13(1), 272. https://doi.org/10.3390/agronomy13010272