
Responses of Cynanchum taiwanianum and Its Bioactive Compound Biosynthesis to Levels of Nitrogen and Potassium Fertilization

Abstract
:1. Introduction
2. Materials and Methods
2.1. Chemicals
2.2. Plant Materials and Experimental Conditions
2.3. Experimental Designs
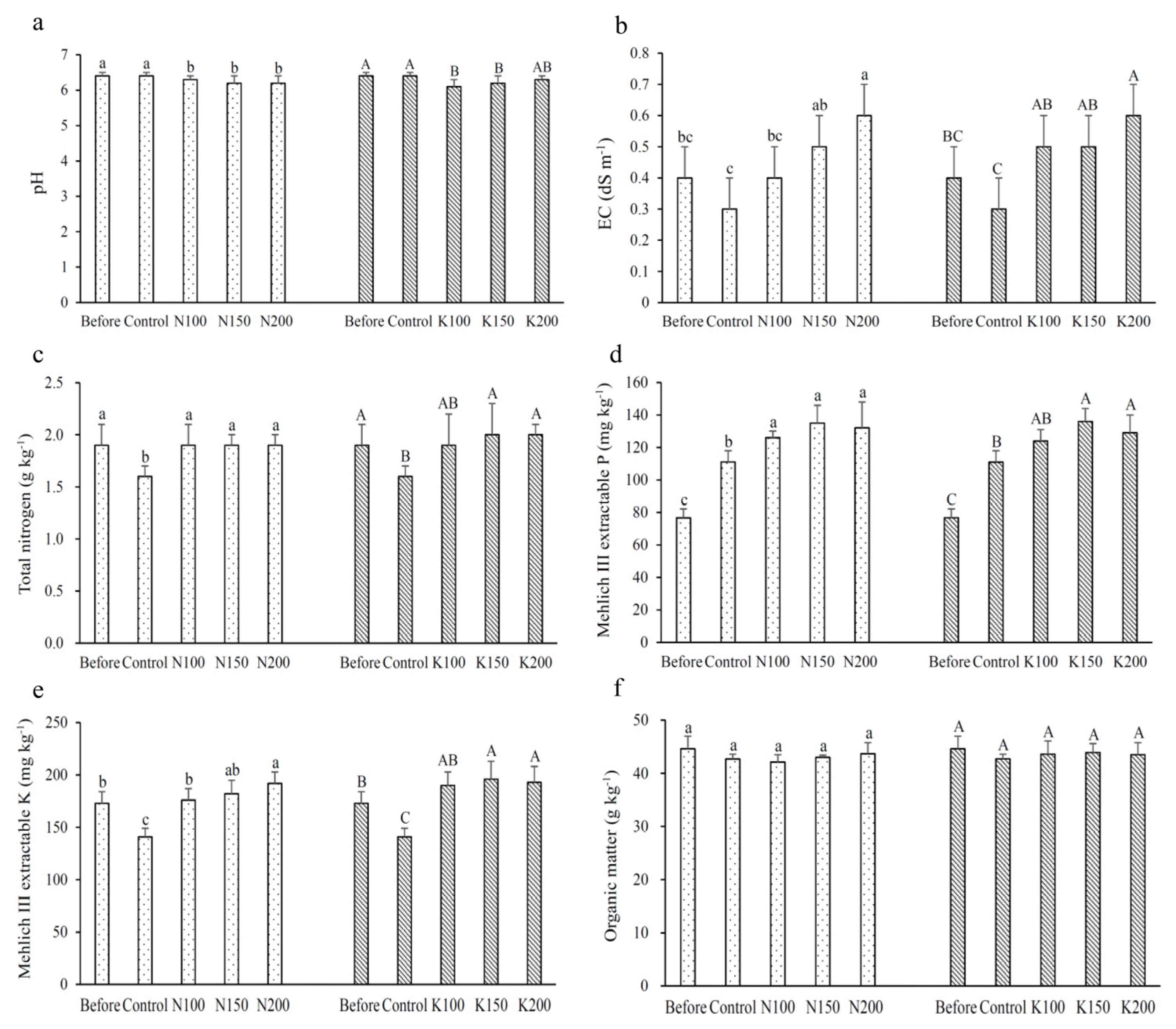
2.4. Soil Chemical Analysis
2.5. Plant Sample Processing
2.6. Analysis of Total Flavonoid and Total Phenolic Contents
2.7. Chromatographic Analysis of Polyphenolic Constituents
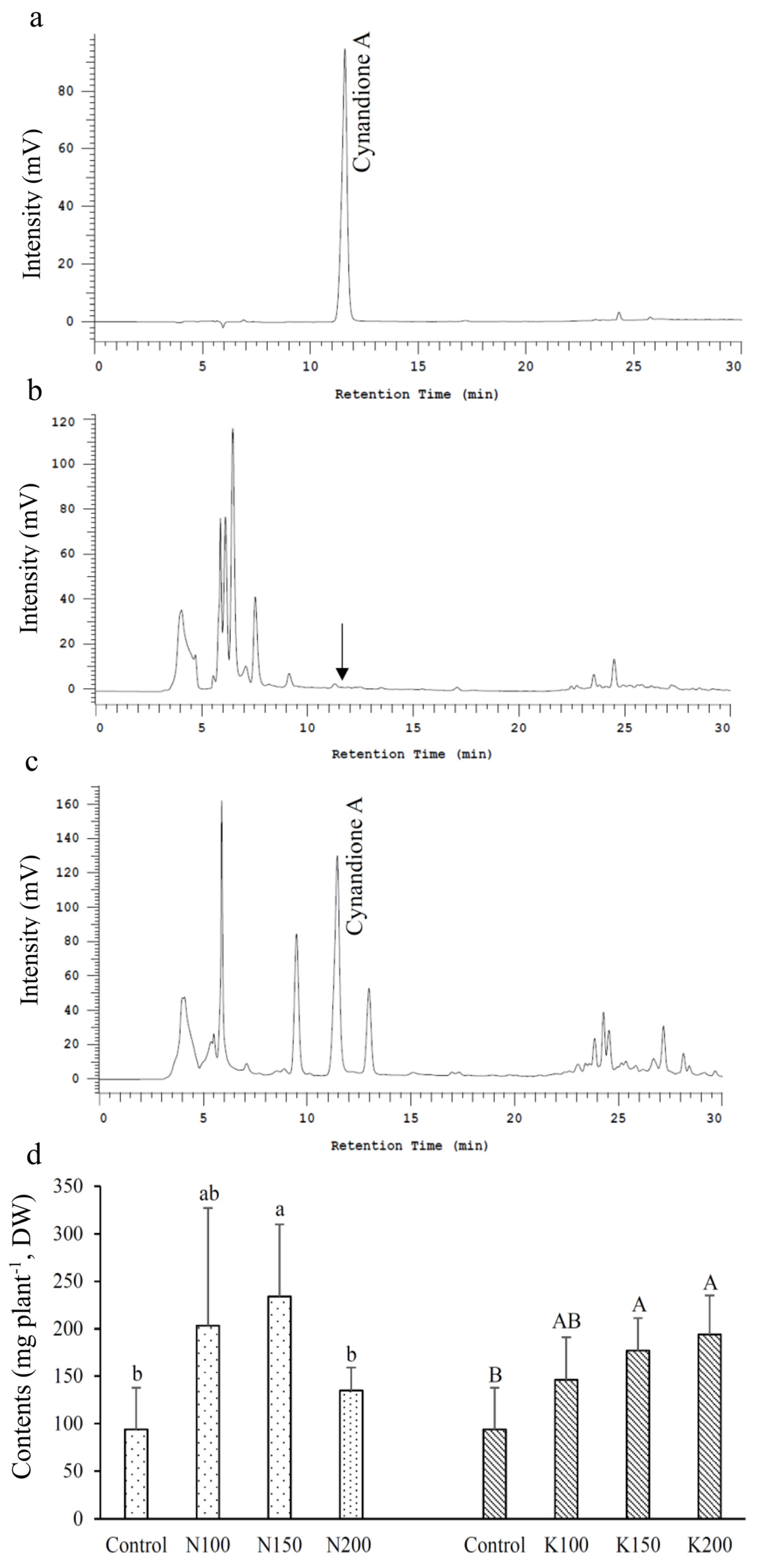
2.8. Analysis of Cynandione A Content
2.9. Statistical Analysis
3. Results
3.1. Chemical Properties of Soils
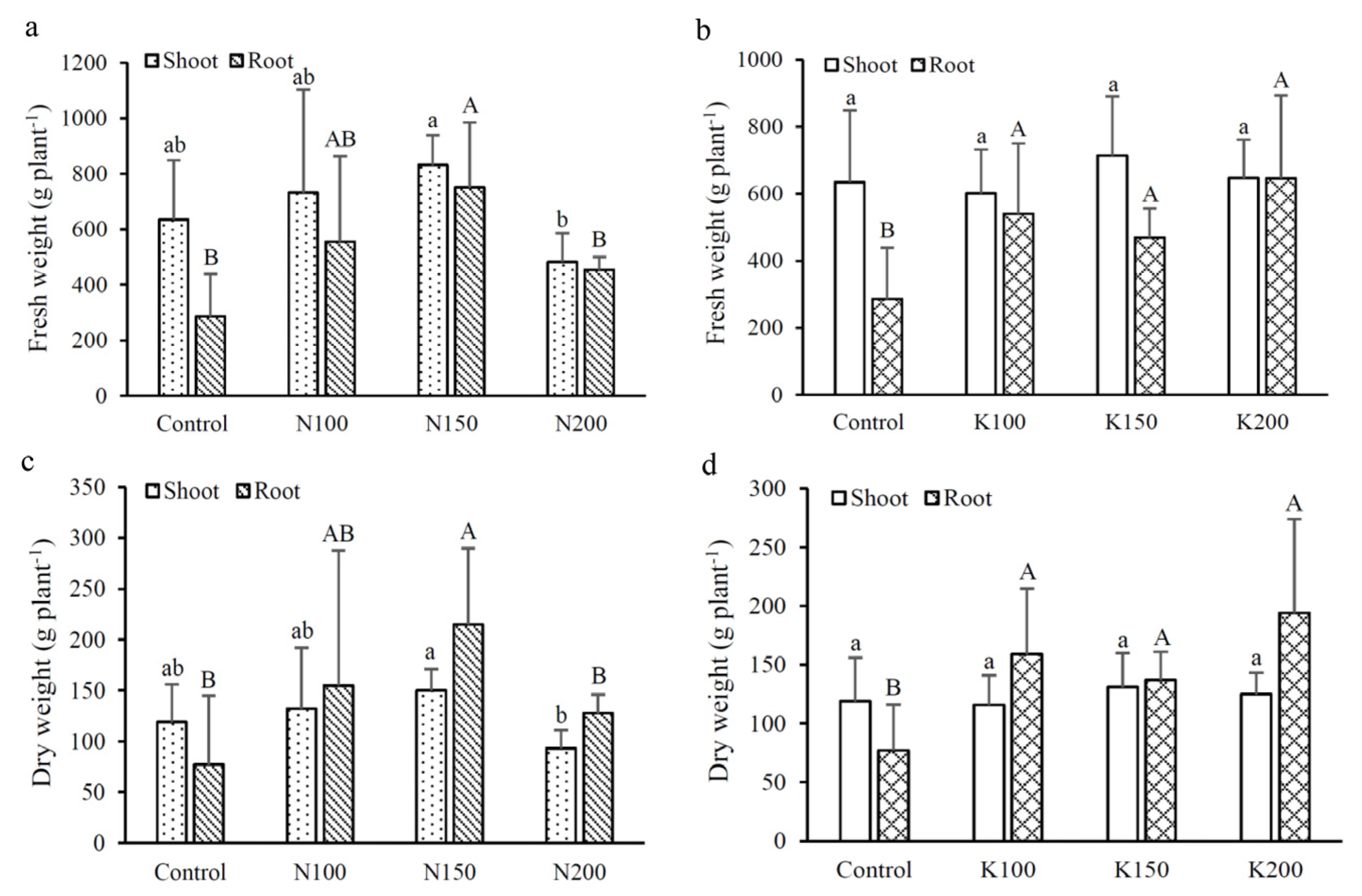
3.2. Biomass Production
3.3. Nutrient Uptake in Plant Parts
3.4. Contents of Total Phenols, Total Flavanoids and Polyphenolic Compounds
3.5. Cynandione A Content
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Lin, C.C.; Yen, M.H.; Wu, Y.W.; Xu, G.J. Histological study on crude drugs Pai-wei, Pai-chein and indigenous species of Cynanchum in Taiwan. Am. J. Chin. Med. 1995, 23, 305–312. [Google Scholar] [CrossRef] [PubMed]
- Lee, H.I. Studies of Antioxidant, Antimicrobial and Anti-Inflammatory Activities of Cynanchum taiwanianum Yamazaki. Ph.D. Thesis, Department of Food Science, National Pingtung University of Science and Technology, Pingtung, Taiwan, 2012; 173p. [Google Scholar]
- Huang, P.L.; Lu, C.M.; Yen, M.H.; Wu, R.R.; Lin, C.N. Acetophenones from Cynanchum taiwanianum. Phytochemistry 1996, 41, 293–295. [Google Scholar] [CrossRef]
- Huang, P.L.; Won, S.J.; Day, S.H.; Lin, C.N. A cytotoxic acetophenone with a novel skeleton, isolated from Cynanchum taiwanianum. Helv. Chim. Acta 1999, 82, 1716–1720. [Google Scholar] [CrossRef]
- Lin, C.N.; Huang, P.L.; Wang, J.J.; Day, S.H.; Lin, H.C.; Wang, J.P.; Ko, Y.L.; Teng, C.M. Stereochemistry and biological activities of constituents from Cynanchum taiwanianum. Biochim. Biophys. Acta 1998, 1380, 115–122. [Google Scholar] [CrossRef]
- Lin, C.N.; Huang, P.L.; Lu, C.M.; Yen, M.H.; Wu, R.R. Revised structure for five acetophenones from Cynanchum taiwanianum. Phytochemistry 1997, 44, 1359–1363. [Google Scholar]
- Lin, Y.L.; Lin, T.C.; Kuo, Y.H. Two acetophenone glucosides, cynanonesides A and B, from Cynanchum taiwanianum and revision of the structure for cynandione A. J. Nat. Prod. 1997, 60, 368–370. [Google Scholar] [CrossRef]
- Lin, Y.L.; Wu, Y.M.; Kuo, Y.H. Revised structures for four acetophenones from Cynanchum taiwanianum. Phytochemistry 1997, 45, 1057–1061. [Google Scholar] [CrossRef]
- Lin, Y.L.; Lin, T.C.; Kuo, Y.H. Five new pregnane glycosides from Cynanchum taiwanianum. J. Nat. Prod. 1995, 58, 1167–1173. [Google Scholar] [CrossRef]
- Lee, H.I.; Guo, J.H.; Wu, C.C.; Ti, M.C.; Lan, K.P.; Hu, C.Y.; Lo, C.Y.; Yang, C.C.; Lin, Y.F.; Wang, T.C. Anti-inflammatory effects of Cynanchum taiwanianum rhizome aqueous extract in IL-1β-induced NRK-52E cells. Pharm. Biol. 2011, 49, 437–444. [Google Scholar] [CrossRef]
- Yue, R.; Yuan, X.; Liu, X.; Zhang, J.; Jiang, P.; He, C.; Shan, L.; Yu, Y.; Zhang, W. Cynandione A mitigates ischemic injuries in rats with cerebral ischemia. J. Neurochem. 2012, 121, 451–464. [Google Scholar] [CrossRef]
- Yang, S.B.; Lee, S.M.; Park, J.H.; Lee, T.H.; Baek, N.I.; Park, H.J.; Lee, H.; Kim, J. Cynandione A from Cynanchum wilfordii attenuates the production of inflammatory mediators in LPS-induced BV-2 microglial cells via NF-κB inactivation. Biol. Pharm. Bull. 2014, 37, 1390–1396. [Google Scholar] [CrossRef] [Green Version]
- Lee, M.K.; Yeo, H.; Kim, J.; Markelonis, G.J.; Oh, T.H.; Kim, Y.C. Cynandione A from Cynanchum wilfordii protects cultured cortical neurons from toxicity induced by H2O2, L-glutamate, and kainate. J. Neurosci. Res. 2000, 59, 259–264. [Google Scholar] [CrossRef]
- Jiang, H.W.; Gu, S.S.; Cao, L.; Jiang, S.S.; Lin, J.; Chen, Y.; Wang, L.; Jiao, H.; Li, J.Y.; Zhao, W.M.; et al. Potential hypoglycemic effect of acetophenones from the root bark of Cynanchum wilfordii. Nat. Prod. Res. 2019, 33, 2314–2321. [Google Scholar] [CrossRef] [PubMed]
- Han, Q.Q.; Yin, M.; Wang, Z.Y.; Liu, H.; Ao, J.P.; Wang, Y.X. Cynandione A alleviates neuropathic pain through α7-nAChR-dependent IL-10/β-endorphin signaling complexes. Front. Pharmacol. 2021, 11, 614450. [Google Scholar] [CrossRef] [PubMed]
- Chung, R.S.; Chen, C.C.; Ng, L.T. Nitrogen fertilization affects the growth performance, betaine and polysaccharide concentrations of Lycium barbarum. Ind. Crops Prod. 2010, 32, 650–655. [Google Scholar] [CrossRef]
- Sugiyama, M.; Takahashi, M.; Katsube, T.; Koyama, A.; Itamura, H. Effects of applied nitrogen amounts on the functional components of Mulberry (Morus alba L.) leaves. J. Agric. Food Chem. 2016, 64, 6923–6929. [Google Scholar] [CrossRef] [PubMed]
- Westermann, D.T.; James, D.W.; Tindall, T.A.; Hurst, R.L. Nitrogen and potassium fertilization of potatoes: Sugars and starch. Am. Potato J. 1994, 71, 433–453. [Google Scholar] [CrossRef]
- Koch, M.; Naumann, M.; Pawelzik, E.; Gransee, A.; Thiel, H. The importance of nutrient management for potato production part I: Plant nutrition and yield. Potato Res. 2020, 63, 97–119. [Google Scholar] [CrossRef] [Green Version]
- Epstein, E.; Bloom, A.J. Mineral Nutrition of Plants: Principles and Perspectives, 2nd ed.; Sinauer Associates Inc.: Sunderland, MA, USA, 2005; p. 400. [Google Scholar]
- Kováčik, J.; Klejdus, B.; Bačkor, M.; Repčák, M. Phenylalanine ammonia-lyase activity and phenolic compounds accumulation in nitrogen-deficient Matricaria chamomilla leaf rosettes. Plant Sci. 2007, 172, 393–399. [Google Scholar] [CrossRef]
- Taiz, L.; Zeiger, E.; Møller, I.M.; Murphy, A. Plant Physiology and Development; Sinauer Associates, Inc.: Sunderland, MA, USA, 2015; 700p. [Google Scholar]
- Nguyen, P.M.; Kwee, E.M.; Niemeyer, E.D. Potassium rate alters the antioxidant capacity and phenolic concentration of basil (Ocimum basilicum L.) leaves. Food Chem. 2010, 123, 1235–1241. [Google Scholar] [CrossRef]
- Redovniković, I.R.; Bogović, M.; Belko, D.; Delonga, K.; Fabek, S.; Novak, B.; Toth, N. Influence of potassium fertilization on the levels of phenolic compounds in sweet potato (Ipomoea batatas L.) leaves. J. Hortic. Sci. Biotechnol. 2012, 87, 47–51. [Google Scholar] [CrossRef]
- Gremigni, P.; Wong, M.T.F.; Edwards, N.K.; Harris, D.; Hamblin, J. Potassium nutrition effects on seed alkaloid concentrations, yield and mineral content of lupins (Lupinus angustifolius). Plant Soil 2001, 234, 131–142. [Google Scholar] [CrossRef]
- Ehsanipour, A.; Razmjoo, J.; Zeinali, H. Effect of nitrogen rates on yield and quality of fennel (Foeniculum vulgare Mill.) accessions. Ind. Crops Prod. 2012, 35, 121–125. [Google Scholar] [CrossRef]
- Heimler, D.; Romani, A.; Ieri, F. Plant polyphenol content, soil fertilization and agricultural management: A review. Eur. Food Res. Technol. 2017, 243, 1107–1115. [Google Scholar] [CrossRef]
- Stumpf, B.; Yan, F.; Honermeier, B. Nitrogen fertilization and maturity influence the phenolic concentration of wheat grain (Triticum aestivum). J. Plant Nutr. Soil Sci. 2015, 178, 118–125. [Google Scholar] [CrossRef]
- McLean, E.O. Soil pH and lime requirement. In Methods of Soil Analysis. Part 2. Chemical and Microbiological Properties, 2nd ed.; Page, A.L., Ed.; American Society of Agronomy Inc.: Madison, WI, USA, 1982; pp. 199–224. [Google Scholar]
- Rhoades, J.D. Soluble salts. In Methods of Soil Analysis. Part 2. Chemical and Microbiological Properties, 2nd ed.; Page, A.L., Ed.; American Society of Agronomy Inc.: Madison, WI, USA, 1982; pp. 167–178. [Google Scholar]
- Murphy, J.; Riley, J.P. A modified single solution method for the determination of phosphate in natural waters. Anal. Chim. Acta 1962, 27, 31–36. [Google Scholar] [CrossRef]
- Mehlich, A. Mehlich 3 soil test extractant: A modification of Mehlich 2 extractant. Commun. Soil Sci. Plant Anal. 1984, 15, 1409–1416. [Google Scholar] [CrossRef]
- Bremner, J.M.; Mulvaney, C.S. Nitrogen-total methods of soil analysis. In Methods of Soil Analysis. Part 2. Chemical and Microbiological Properties, 2nd ed.; Page, A.L., Ed.; American Society of Agronomy Inc.: Madison, WI, USA, 1982; pp. 595–624. [Google Scholar]
- Soon, Y.K.; Abboud, S. A comparison of some methods for soil organic carbon determination. Commun. Soil Sci. Plant Anal. 1991, 22, 943–954. [Google Scholar] [CrossRef]
- Wu, S.J.; Ng, L.T. Antioxidant and free radical scavenging activities of wild bitter melon (Momordica charantia Linn. var. abbreviata Ser.) in Taiwan. LWT-Food Sci. Technol. 2008, 41, 323–330. [Google Scholar] [CrossRef]
- Huang, S.H.; Ng, L.T. Quantification of polyphenolic content and bioactive constituents of some commercial rice varieties in Taiwan. J. Food Compos. Anal. 2012, 26, 122–127. [Google Scholar] [CrossRef]
- Cai, Z.; Wang, B.; Xu, M.; Zhang, H.; Zhang, L.; Gao, S. Nitrification and acidification from urea application in red soil (Ferralic Cambisol) after different long-term fertilization treatments. J. Soils Sediments 2014, 14, 1526–1536. [Google Scholar] [CrossRef]
- Citak, S.; Sonmez, S. Effects of chemical fertilizer and different organic manures application on soil pH, EC and organic matter content. J. Food Agric. Environ. 2011, 9, 739–741. [Google Scholar]
- Machado, R.M.A.; Serralheiro, R.P. Soil salinity: Effect on vegetable crop growth. Management practices to prevent and mitigate soil salinization. Horticulturae 2017, 3, 30. [Google Scholar] [CrossRef]
- Lin, Y.R.; Cheng, C.H.; Tseng, T.Y.; Wang, S.L.; Guo, H.Y. Soil properties and organic carbon sequestration in the long-term afforested stands of plain areas. Taiwan J. Agric. Chem. Food Sci. 2011, 49, 260–274. [Google Scholar]
- Allison, M.F.; Flower, J.H.; Allen, E.J. Responses of potato (Solanum tuberosum) to potassium fertilizers. J. Agric. Sci. 2001, 136, 407–426. [Google Scholar] [CrossRef] [Green Version]
- Pen, W.N.; Hsin, T.W.; Lin, W.H. The effect of plant densities and potassium fertilizations on root tuber size and yield of sweet potato. Bull. Taoyuan Dist. Agric. Res. Ext. Stn. 1994, 17, 1–8. [Google Scholar]
- Kung, T.L.; Jiang, J.L.; Hsin, T.W. The effect of nitrogen and potassium fertilizers on agronomic characteristics and tuber yield of Chinese yam (Dioscorea batatas). Bull. Taoyuan Dist. Agric. Res. Ext. Stn. 1999, 36, 1–6. [Google Scholar]
- Zhang, F.; Wang, J.H.; Yu, S.L.; Chen, Y.H.; Dong, Q.Y. Accumulation and distribution of nitrogen, phosphorus, and potassium in Cynanchum bungei Decne. and their relationships to dry matter production. Soil Sci. Plant Nutr. 2006, 12, 369–373. [Google Scholar]
- Haeder, H.E.; Mengel, K.; Forster, H. The effect of potassium on translocation of photosynthates and yield pattern of potato plants. J. Agric. Food Chem. 1973, 24, 1479–1487. [Google Scholar] [CrossRef]
- Porter, G.A.; Sisson, J.A. Response of Russet Burbank and Shepody potatoes to nitrogen fertilizer in two cropping systems. Am. Potato J. 1991, 68, 425–443. [Google Scholar] [CrossRef]
- Chen, X.G.; Li, H.M.; Zhang, A.J.; Shi, X.M.; Tang, Z.H.; Wei, M.; Shi, C.Y. Effect of paclobutrazol under different N application rates on photosynthesis and starch accumulation in edible sweet potato. Acta Agron. Sin. 2012, 38, 1728–1733. [Google Scholar]
- Li, Z.G.; Zhang, R.H.; Xia, S.J.; Wang, L.; Liu, C.; Zhang, R.Q.; Fan, Z.H.; Chen, F.; Liu, Y. Interactions between N, P and K fertilizers affect the environment and the yield and quality of satsumas. Glob. Ecol. Conserv. 2019, 19, e00663. [Google Scholar] [CrossRef]
- Yan, Z.; Kim, N.; Han, W.; Guo, Y.; Han, T.; Du, E.; Fang, J. Effects of nitrogen and phosphorus supply on growth rate, leaf stoichiometry, and nutrient resorption of Arabidopsis thaliana. Plant Soil 2015, 388, 147–155. [Google Scholar] [CrossRef]
- Thomson, C.; Marschner, H.; Römheld, V. Effect of nitrogen fertilizer form on pH of the bulk soil and rhizosphere, and on the growth, phosphorus, and micronutrient uptake of bean. J. Plant Nutr. 1993, 16, 493–506. [Google Scholar] [CrossRef]
- Ruan, J.; Zhang, F.; Wong, M.H. Effect of nitrogen form and phosphorus source on the growth, nutrient uptake and rhizosphere soil property of Camellia sinensis L. Plant Soil 2000, 223, 65–73. [Google Scholar] [CrossRef]
- Tsai, J.H.; Wu, T.Y. Soil information and fertilization recommendation of taro in paddy field at Miaoli district. Bull. Taoyuan Dist. Agric. Res. Ext. Stn. 2017, 6, 1–14. [Google Scholar]
- Wills, R.B.; Bone, K.; Morgan, M. Herbal products: Active constituents, modes of action and quality control. Nutr. Res. Rev. 2000, 13, 47–77. [Google Scholar] [CrossRef]
- Amarowicz, R.; Cwalina-Ambroziak, B.; Janiak, M.A.; Bogucka, B. Effect of N fertilization on the content of phenolic compounds in Jerusalem artichoke (Helianthus tuberosus L.) tubers and their antioxidant capacity. Agronomy 2020, 10, 1215. [Google Scholar] [CrossRef]
- Jones, C.G.; Hartley, S.E. A protein competition model of phenolic allocation. Oikos 1999, 86, 27–44. [Google Scholar] [CrossRef]
- Mittelstraß, K.; Treutter, D.; Pleßl, M.; Heller, W.; Elstner, E.F.; Heiser, I. Modification of primary and secondary metabolism of potato plants by nitrogen application differentially affects resistance to Phytophthora infestans and Alternaria solani. Plant Biol. 2006, 8, 653–661. [Google Scholar] [CrossRef] [PubMed]
- Rasouli, H.; Hosein Farzaei, M.; Khodarahmi, R. Polyphenols and their benefits: A review. Int. J. Food Prop. 2017, 20, 1700–1741. [Google Scholar] [CrossRef] [Green Version]
- Chen, Z.S.; Lai, J.S.; Kao, Y.H. The constituents of Cynanchum taiwanianum. J. Chin. Chem. Soc. 1991, 38, 393–396. [Google Scholar] [CrossRef]
- Li, Y.; Piao, D.; Zhang, H.; Woo, M.H.; Lee, J.H.; Moon, D.C.; Lee, S.H.; Chang, H.W.; Son, J.K. Quality assessment and discrimination of the roots of Cynanchum auriculatum and Cynanchum wilfordii by HPLC-UV analysis. Arch. Pharm. Res. 2013, 36, 335–344. [Google Scholar] [CrossRef] [PubMed]
- Wink, M. Physiology of accumulation of secondary metabolites with special reference to alkaloids. In Cell Cultures and Somatic Cell Genetics of Plants; Constabel, F., Vasil, I.K., Eds.; Academic Press: San Diego, CA, USA, 1987; Volume 4, pp. 17–42. [Google Scholar]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Shoots | ||||||
| Concentrations (mg kg−1 DW) | ||||||
| Gallic acid | Catechin | Chlorogenic acid | Epicatechin | Ferulic acid | Cinnamic acid | |
| Nitrogen fertilization | ||||||
| Control | 518 ± 63 a | 551 ± 233 a | 335 ± 73 a | 495 ± 100 a | 383 ± 271 a | 974 ± 513 a |
| N100 | 523 ± 92 a | 464 ± 209 a | 357 ± 77 a | 538 ± 170 a | 252 ± 123 a | 980 ± 478 a |
| N150 | 438 ± 185 a | 343 ± 74 a | 285 ± 117 a | 467 ± 257 a | 299 ± 125 a | 991 ± 203 a |
| N200 | 424 ± 228 a | 340 ± 191 a | 294 ± 117 a | 531 ± 148 a | 272 ± 153 a | 1157 ± 139 a |
| Potassium fertilization | ||||||
| Control | 518 ± 63 a | 551 ± 233 a | 335 ± 73 a | 495 ± 100 a | 383 ± 271 a | 974 ± 513 bc |
| K100 | 416 ± 93 a | 363 ± 82 a | 319 ± 87 a | 538 ± 185 a | 414 ± 161 a | 1427 ± 167 a |
| K150 | 461 ± 91 a | 456 ± 128 a | 281 ± 44 a | 425 ± 125 a | 186 ± 51 a | 597 ± 133 c |
| K200 | 523 ± 207 a | 493 ± 120 a | 347 ± 36 a | 504 ± 92 a | 382 ± 149 a | 1091 ± 115 ab |
| Roots | ||||||
| Concentrations (mg kg−1 DW) | ||||||
| Gallic acid | Catechin | Chlorogenic acid | Vanillic acid | Epicatechin | Ferulic acid | |
| Nitrogen fertilization | ||||||
| Control | 100 ± 43 a | 652 ± 293 a | 74 ± 4 a | 86 ± 44 a | 2550 ± 741 a | 608 ± 117 a |
| N100 | 106 ± 16 a | 712 ± 268 a | 71 ± 19 a | 116 ± 32 a | 2488 ± 454 a | 842 ± 388 a |
| N150 | 68 ± 34 a | 682 ± 230 a | 80 ± 28 a | 133 ± 62 a | 2521 ± 738 a | 719 ± 365 a |
| N200 | 110 ± 9 a | 631 ± 244 a | 71 ± 13 a | 135 ± 45 a | 1844 ± 328 a | 634 ± 292 a |
| Potassium fertilization | ||||||
| Control | 100 ± 43 a | 652 ± 293 a | 74 ± 4 a | 86 ± 44 a | 2550 ± 741 a | 608 ± 117 a |
| K100 | 95 ± 39 a | 467 ± 110 a | 61 ± 4 a | 75 ± 10 a | 1802 ± 113 a | 494 ± 82 a |
| K150 | 125 ± 13 a | 826 ± 291 a | 63 ± 6 a | 101 ± 10 a | 2501 ± 404 a | 667 ± 194 a |
| K200 | 87 ± 55 a | 491 ± 267 a | 61 ± 7 a | 79 ± 18 a | 1766 ± 521 a | 749 ± 413 a |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tseng, H.-M.; Lu, T.-M.; Ng, L.-T. Responses of Cynanchum taiwanianum and Its Bioactive Compound Biosynthesis to Levels of Nitrogen and Potassium Fertilization. Agronomy 2022, 12, 180. https://doi.org/10.3390/agronomy12010180
Tseng H-M, Lu T-M, Ng L-T. Responses of Cynanchum taiwanianum and Its Bioactive Compound Biosynthesis to Levels of Nitrogen and Potassium Fertilization. Agronomy. 2022; 12(1):180. https://doi.org/10.3390/agronomy12010180
Chicago/Turabian StyleTseng, Han-Mei, Tzy-Ming Lu, and Lean-Teik Ng. 2022. "Responses of Cynanchum taiwanianum and Its Bioactive Compound Biosynthesis to Levels of Nitrogen and Potassium Fertilization" Agronomy 12, no. 1: 180. https://doi.org/10.3390/agronomy12010180
APA StyleTseng, H.-M., Lu, T.-M., & Ng, L.-T. (2022). Responses of Cynanchum taiwanianum and Its Bioactive Compound Biosynthesis to Levels of Nitrogen and Potassium Fertilization. Agronomy, 12(1), 180. https://doi.org/10.3390/agronomy12010180