
Exploiting Agronomic and Biochemical Traits to Develop Heat Resilient Cotton Cultivars under Climate Change Scenarios

 , , , ,
, , , ,
Abstract
:1. Introduction
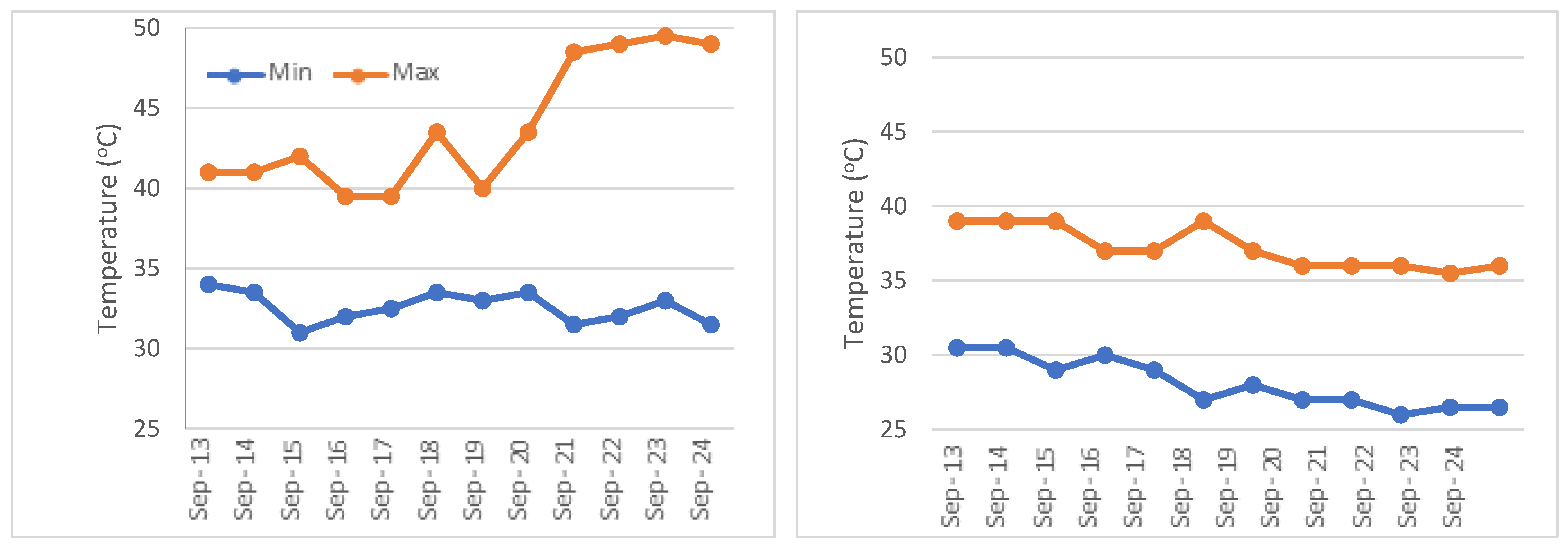
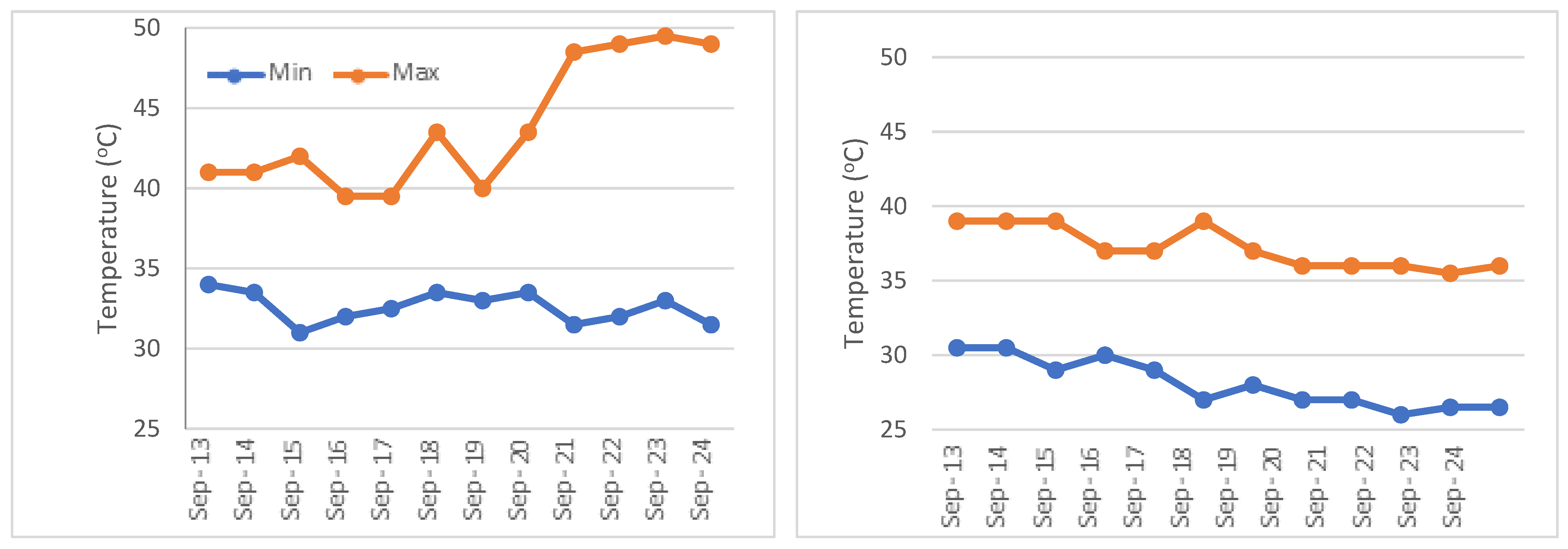
2. Materials and Methods
2.1. Biochemical and Yield-Related Parameters
2.2. Statistical Analysis
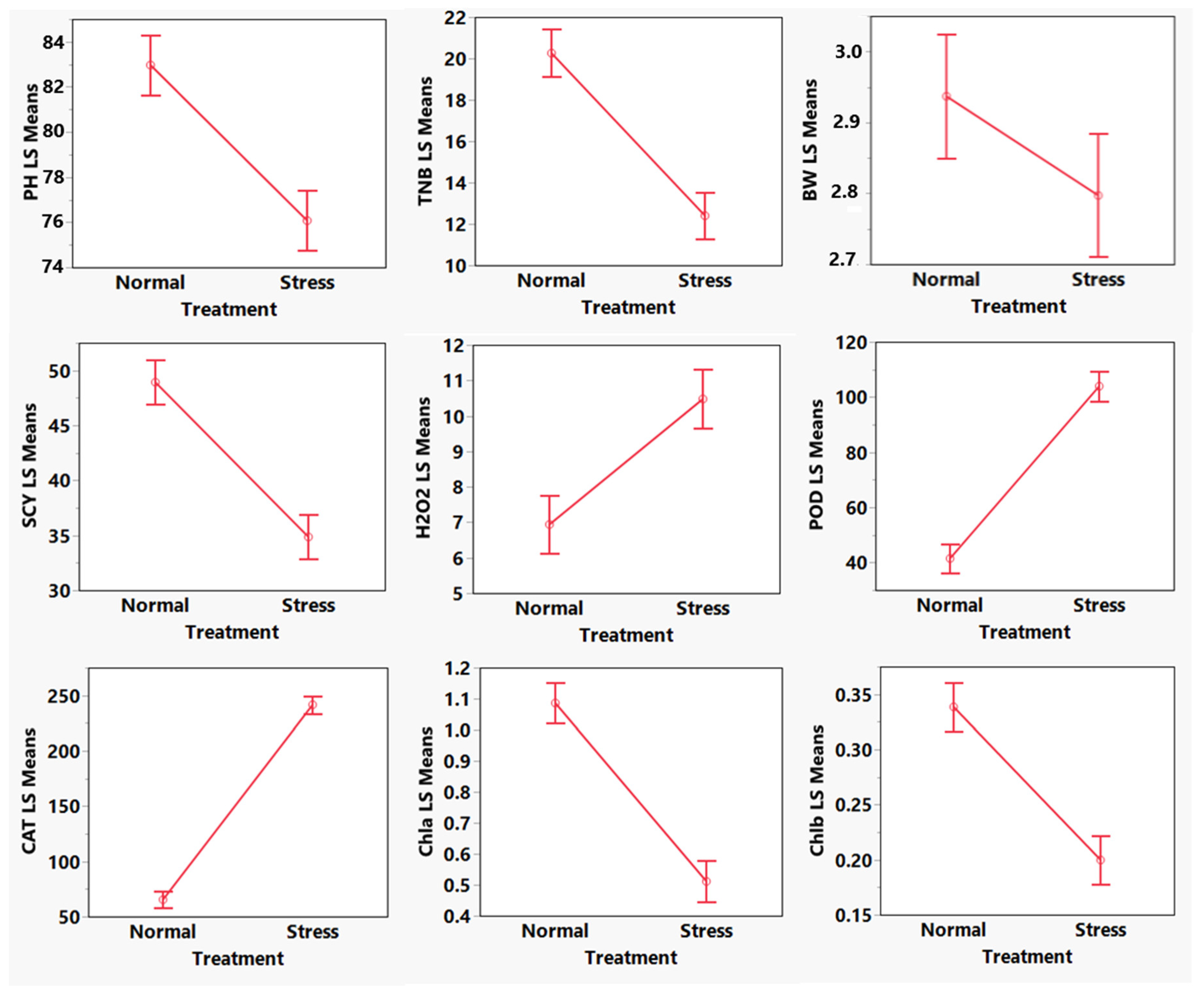
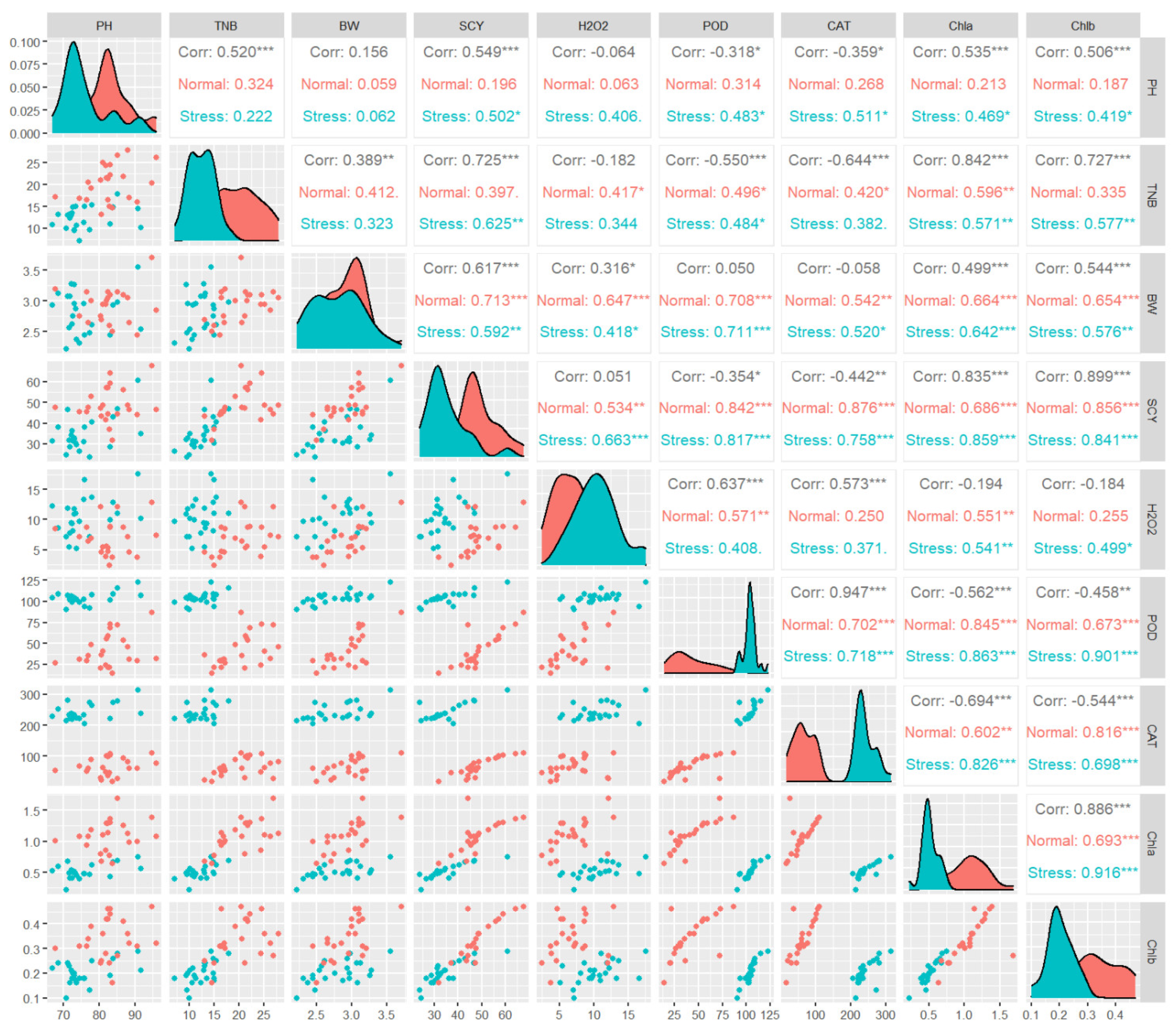
3. Results
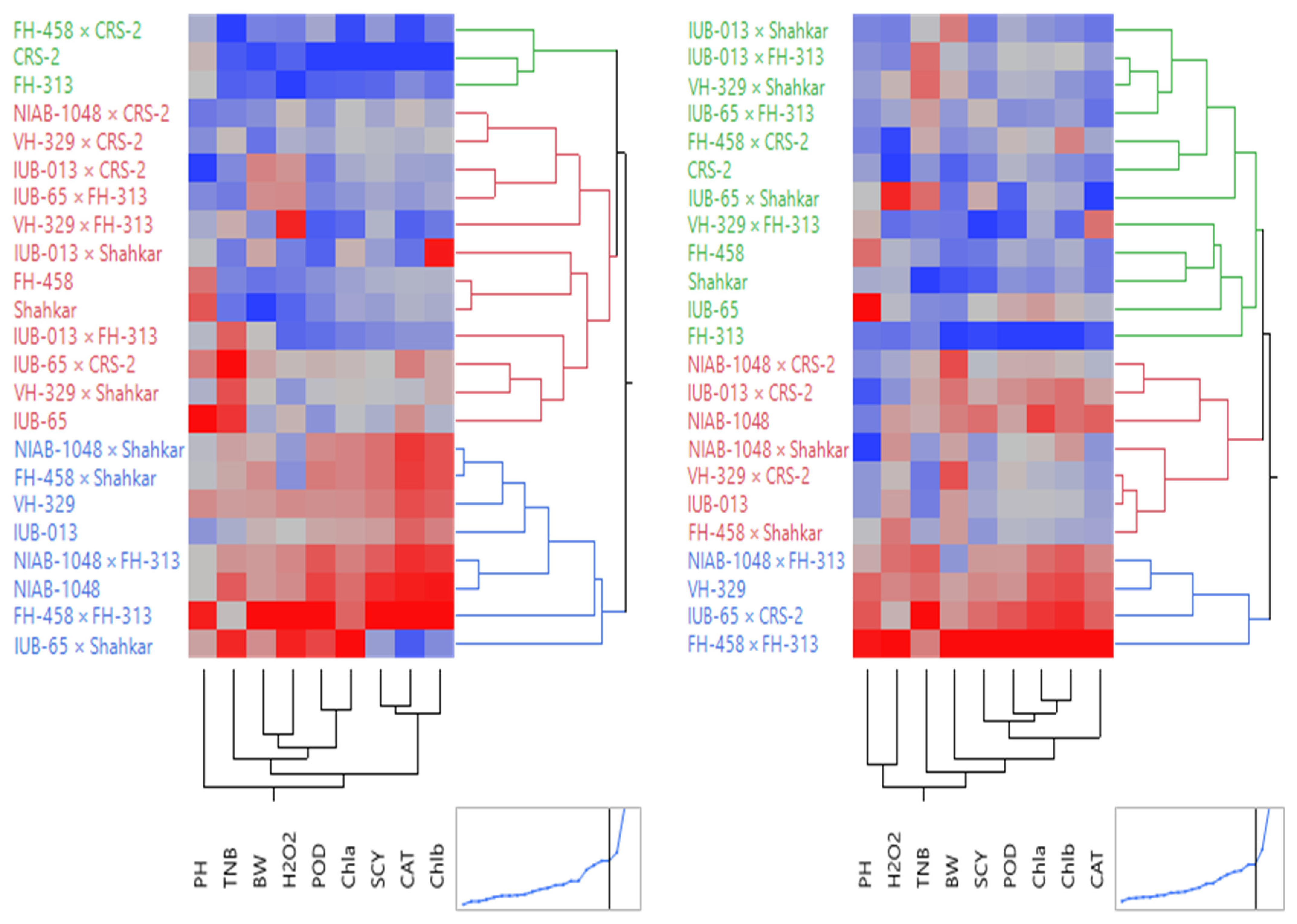
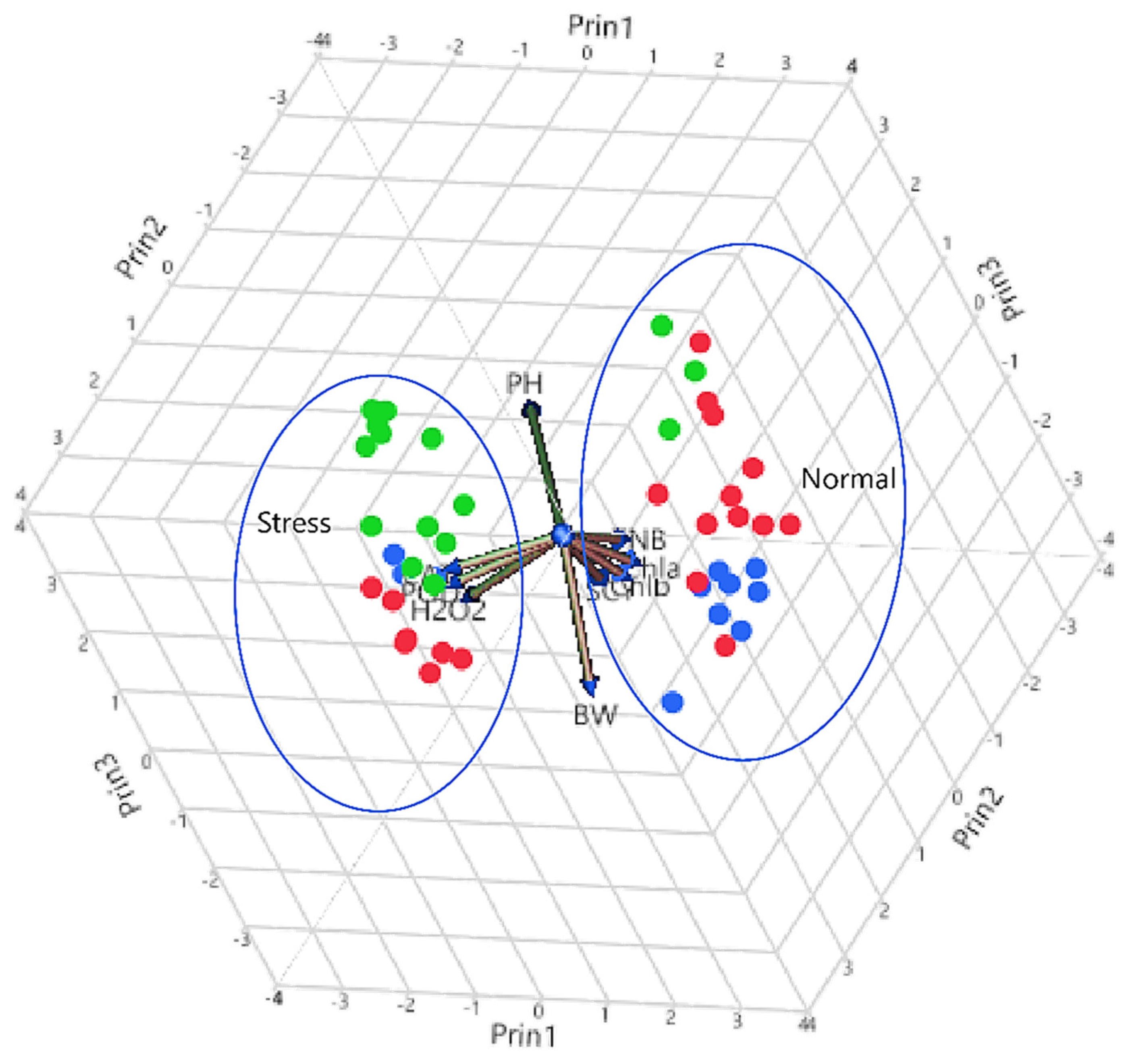
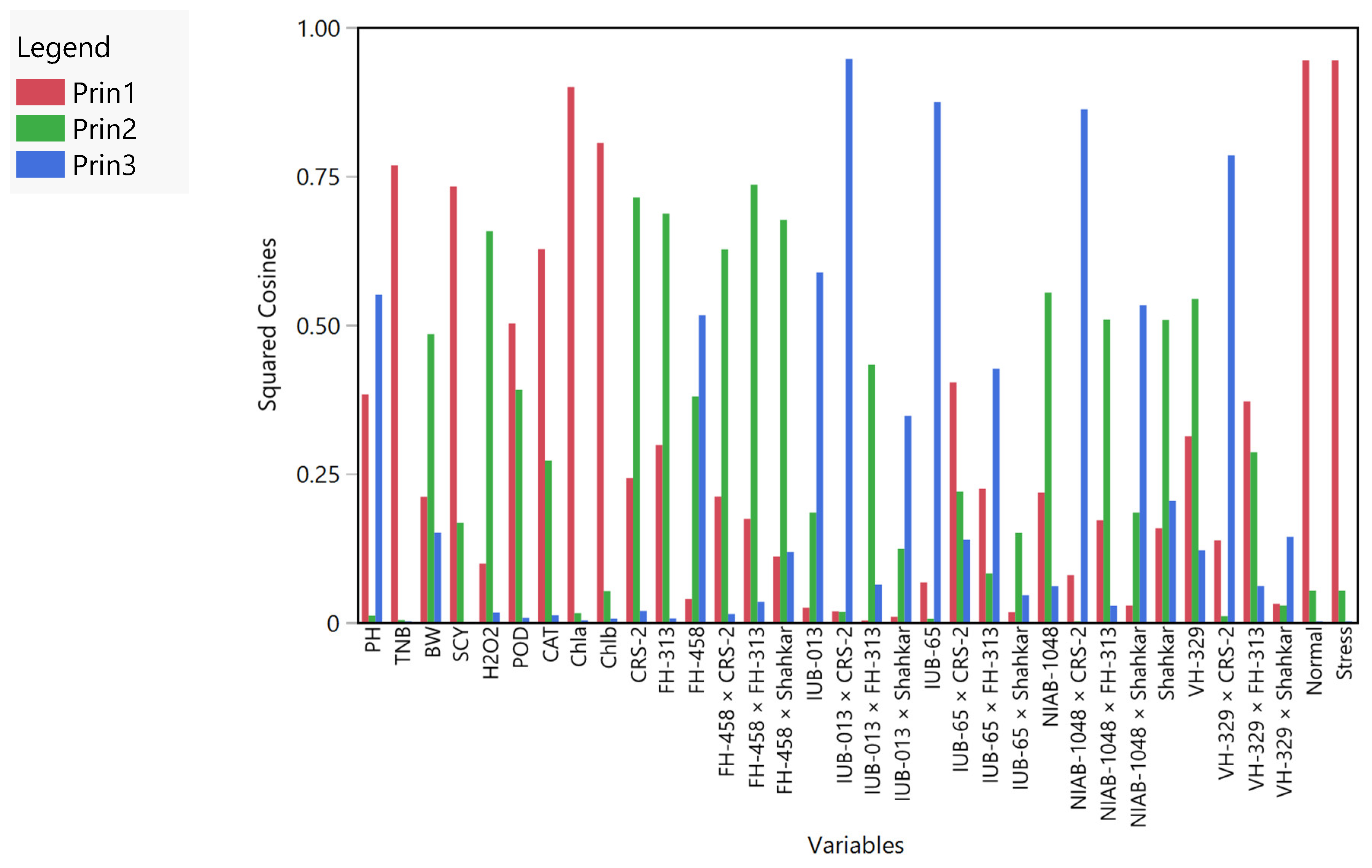
4. Cluster Analysis
Principal Component Analysis (PCA)
5. Discussion
6. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Shahrajabian, M.H.; Sun, W.; Cheng, Q. Considering white gold, cotton for its fiber, seed oil, traditional and modern health benefits. J. Biol. Environ. Sci. 2020, 14, 25–39. [Google Scholar]
- Lee, J.; Nadolnyak, D.A.; Hartarska, V.M. Impact of climate change on agricultural production in Asian countries: Evidence from panel study. In Proceedings of the Southern Agricultural Economics Association Annual Meeting, Birmingham, AL, USA, 4–7 February 2012. [Google Scholar]
- Wahid, A.; Gelani, S.; Ashraf, M.; Foolad, M.R. Heat tolerance in plants: An overview. Environ. Exp. Bot. 2007, 61, 199–223. [Google Scholar] [CrossRef]
- Echer, F.R.; Oosterhuis, D.M.; Loka, D.A.; Rosolem, C.A. High Night Temperatures During the Floral Bud Stage Increase the Abscission of Reproductive Structures in Cotton. J. Agron. Crop. Sci. 2014, 200, 191–198. [Google Scholar] [CrossRef]
- Rafiq, A.; Iqbal, M.S.; Ibrar, D.; Mahmood, T.; Naveed, M.S.; Naeem, M.K. A review on heat stress response in different genotypes of tomato crop (Solanum lycopersicon L.). Int. J. Mod. Agric. 2013, 2, 64–71. [Google Scholar] [CrossRef]
- El Sabagh, A.; Hossain, A.; Islam, M.S.; Barutcular, C.; Ratnasekera, D.; Gormus, O.; Amanet, K.; Mubeen, M.; Nasim, W.; Fahad, S.; et al. Drought and Heat Stress in Cotton (Gossypium hirsutum L.): Consequences and Their Possible Mitigation Strategies. In Agronomic Crops: Volume 3: Stress Responses and Tolerance; Hasanuzzaman, M., Ed.; Springer: Singapore, 2020; pp. 613–634. [Google Scholar]
- Rizhsky, L.; Liang, H.; Shuman, J.; Shulaev, V.; Davletova, S.; Mittler, R. When Defense Pathways Collide. The Response of Arabidopsis to a Combination of Drought and Heat Stress. Plant Physiol. 2004, 134, 1683–1696. [Google Scholar] [CrossRef] [Green Version]
- Saleem, M.A.; Malik, W.; Qayyum, A.; Ul-Allah, S.; Ahmad, M.Q.; Afzal, H.; Amjid, M.W.; Ateeq, M.F.; Zia, Z.U. Impact of heat stress responsive factors on growth and physiology of cotton (Gossypium hirsutum L.). Mol. Biol. Rep. 2021, 48, 1069–1079. [Google Scholar] [CrossRef] [PubMed]
- Oosterhuis, D.M.; Snider, J.L. High temperature stress on floral development and yield of cotton. In Stress Physiology in Cotton; Oosterhuis, D.M., Ed.; The Cotton Foundation: Cordova, TN, USA, 2011; pp. 1–24. [Google Scholar]
- Singh, R.P.; Prasad, P.V.; Sunita, K.; Giri, S.; Reddy, K.R. Influence of high temperature and breeding for heat tolerance in cotton: A review. Adv. Agron. 2007, 93, 313–385. [Google Scholar]
- Xu, W.; Zhou, Z.; Zhan, D.; Zhao, W.; Meng, Y.; Chen, B.; Liu, W.; Wang, Y. The difference in the formation of thermotolerance of two cotton cultivars with different heat tolerance. Arch. Agron. Soil Sci. 2020, 66, 58–69. [Google Scholar] [CrossRef]
- Dabbert, T.; Gore, M.A. Challenges and perspectives on improving heat and drought stress resilience in cotton. J. Cotton Sci. 2014, 18, 393–409. [Google Scholar]
- Ozturk, M.; Hakeem, K.R.; Ashraf, M.; Ahmad, M.S.A. Crop Production Technologies for Sustainable Use and Conservation: Physiological and Molecular Advances; CRC Press: Boca Raton, FL, USA, 2019. [Google Scholar]
- Iqbal, M.; Ul-Allah, S.; Naeem, M.; Ijaz, M.; Sattar, A.; Sher, A. Response of cotton genotypes to water and heat stress: From field to genes. Euphytica 2017, 213, 1–11. [Google Scholar] [CrossRef]
- Farooq, A.; Shakeel, A.; Chattha, W.S.; Khan, T.M.; Azhar, M.T.; Saeed, A. Genetic variability in cotton germplasm: Predicting the agro physiological markers for high-temperature tolerance. J. Agric. Sci. 2021, 1–12. [Google Scholar] [CrossRef]
- Sarwar, M.; Saleem, M.; Najeeb, U.; Shakeel, A.; Ali, S.; Bilal, M. Hydrogen peroxide reduces heat-induced yield losses in cotton (Gossypium hirsutum L.) by protecting cellular membrane damage. J. Agron. Crop. Sci. 2017, 203, 429–441. [Google Scholar] [CrossRef]
- Liu, Z.; Yuan, Y.L.; Liu, S.Q.; Yu, X.N.; Rao, L.Q. Screening for high-temperature tolerant cotton cultivars by testing in vitro pollen germination, pollen tube growth and boll retention. J. Integr. Plant Biol. 2006, 48, 706–714. [Google Scholar] [CrossRef]
- Conaty, W.; Burke, J.; Mahan, J.; Neilsen, J.; Sutton, B. Determining the optimum plant temperature of cotton physiology and yield to improve plant-based irrigation scheduling. Crop Sci. 2012, 52, 1828–1836. [Google Scholar] [CrossRef]
- Lokhande, S.; Reddy, K.R. Quantifying temperature effects on cotton reproductive efficiency and fiber quality. Agron. J. 2014, 106, 1275–1282. [Google Scholar] [CrossRef]
- Reddy, K.; Hodges, H.; Reddy, V. Temperature effects on cotton fruit retention. Agron. J. 1992, 84, 26–30. [Google Scholar] [CrossRef]
- Wullschleger, S.D.; Oosterhuis, D.M. A rapid leaf-disc sampler for psychrometric water potential measurements. Plant Physiol. 1986, 81, 684–685. [Google Scholar] [CrossRef] [Green Version]
- Cottee, N.; Tan, D.; Bange, M.; Cothren, J.; Campbell, L. Multi-level determination of heat tolerance in cotton (Gossypium hirsutum L.) under field conditions. Crop Sci. 2010, 50, 2553–2564. [Google Scholar] [CrossRef]
- Zhang, J.; Zou, D.; Li, Y.; Sun, X.; Wang, N.-N.; Gong, S.-Y.; Zheng, Y.; Li, X.-B. GhMPK17, a cotton mitogen-activated protein kinase, is involved in plant response to high salinity and osmotic stresses and ABA signaling. PLoS ONE 2014, 9, e95642. [Google Scholar]
- Kamal, M.; Saleem, M.; Shahid, M.; Awais, M.; Khan, H.; Ahmed, K. Ascorbic acid triggered physiochemical transformations at different phenological stages of heat-stressed Bt cotton. J. Agron. Crop. Sci. 2017, 203, 323–331. [Google Scholar] [CrossRef]
- Kocsy, G.; Szalai, G.; Sutka, J.; Páldi, E.; Galiba, G. Heat tolerance together with heat stress-induced changes in glutathione and hydroxymethylglutathione levels is affected by chromosome 5A of wheat. Plant Sci. 2004, 166, 451–458. [Google Scholar] [CrossRef]
- Roychoudhury, A.; Basu, S.; Sengupta, D.N. Antioxidants and stress-related metabolites in the seedlings of two indica rice varieties exposed to cadmium chloride toxicity. Acta Physiol. Plant. 2012, 34, 835–847. [Google Scholar] [CrossRef]
- Suzuki, N.; Koussevitzky, S.; Mittler, R.; Miller, G. ROS and redox signalling in the response of plants to abiotic stress. Plant Cell Environ. 2012, 35, 259–270. [Google Scholar] [CrossRef]
- Ali, B.; Iqbal, M.S.; Shah, M.K.N.; Shabbir, G.; Cheema, N.M. Genetic analysis for various traits in Gossypium hirsutum L. Pak. J. Agric. Res. 2011, 24, 8–13. [Google Scholar]
- Singh, S.; Vikram, P.; Sehgal, D.; Burgueño, J.; Sharma, A.; Singh, S.K.; Sansaloni, C.P.; Joynson, R.; Brabbs, T.; Ortiz, C.; et al. Harnessing genetic potential of wheat germplasm banks through impact-oriented-prebreeding for future food and nutritional security. Sci. Rep. 2018, 8, 12527. [Google Scholar] [CrossRef] [Green Version]
- Azhar, F.; Ali, Z.; Akhtar, M.; Khan, A.; Trethowan, R. Genetic variability of heat tolerance, and its effect on yield and fibre quality traits in upland cotton (Gossypium hirsutum L.). Plant Breed. 2009, 128, 356–362. [Google Scholar] [CrossRef]
- Gupta, P.; Langridge, P.; Mir, R. Marker-assisted wheat breeding: Present status and future possibilities. Mol. Breed. 2010, 26, 145–161. [Google Scholar] [CrossRef]
- Mangi, N.; Nazir, M.F.; Wang, X.; Iqbal, M.S.; Sarfraz, Z.; Jatoi, G.H.; Mahmood, T.; Ma, Q.; Shuli, F. Dissecting Source-Sink Relationship of Subtending Leaf for Yield and Fiber Quality Attributes in Upland Cotton (Gossypium hirsutum L.). Plants 2021, 10, 1147. [Google Scholar] [CrossRef]
- Jehanzeb Farooq, M.R.; Muhammad, A.; Muhammad, R.; Khalid, M.; Petrescu-Mag, V. Multivariate analysis for CLCuD and various morphological traits in some advanced lines of cotton Gossypium hirsutum L. Adv. Agric. Bot. Int. J. Bioflux Soc. 2015, 7, 241–247. [Google Scholar]
- Singh, R.; Gupta, M. Multivariate analysis of divergence in upland cotton. Indian J. Genet. Plant Breed. 1968, 47, 11–14. [Google Scholar]
- Rathinavel, K. Principal Component Analysis with Quantitative Traits in Extant Cotton Varieties (Gossypium Hirsutum L.) and Parental Lines for Diversity. Curr. Agric. Res. J. 2018, 6, 54–64. [Google Scholar] [CrossRef]
- Bange, M.P.; Baker, J.T.; Bauer, P.J.; Broughton, K.J.; Constable, G.A.; Luo, Q.; Oosterhuis, D.M.; Osanai, Y.; Payton, P.; Tissue, D.T. Climate Change and Cotton Production in Modern Farming Systems; CABI: Oxfordshire, UK, 2016. [Google Scholar]
- Jans, Y.; von Bloh, W.; Schaphoff, S.; Müller, C. Global cotton production under climate change—Implications for yield and water consumption. Hydrol. Earth Syst. Sci. 2020, 25, 2027–2044. [Google Scholar] [CrossRef]
- Soviadan, M.K.; Koffi-Tessio, E.M.; Enete, A.A.; Nweze, N.J. Impact of Climate Change on Cotton Production: Case of Savannah Region, Northern Togo. Agric. Sci. 2019, 10, 927–947. [Google Scholar] [CrossRef] [Green Version]
- Sousa, C.C.; Damasceno-Silva, K.J.; Bastos, E.A.; Rocha, M.M. Selection of cowpea progenies with enhanced drought-tolerance traits using principal component analysis. Genet. Mol. Res. 2015, 14, 15981–15987. [Google Scholar] [CrossRef]
- Mohammadi, S.A.; Prasanna, B. Analysis of genetic diversity in crop plants—Salient statistical tools and considerations. Crop Sci. 2003, 43, 1235–1248. [Google Scholar] [CrossRef] [Green Version]
- Muhammad, A.; Amir, S.; Khan, T.M.; Irfan, A. Genetic basis of variation for high temperature tolerance in upland cotton. Int. J. Agric. Biol. 2018, 20, 2637–2646. [Google Scholar]
- Both, A.-J.; Benjamin, L.; Franklin, J.; Holroyd, G.; Incoll, L.D.; Lefsrud, M.G.; Pitkin, G. Guidelines for measuring and reporting environmental parameters for experiments in greenhouses. Plant Methods 2015, 11, 1–18. [Google Scholar] [CrossRef]
- Velikova, V.; Yordanov, I.; Edreva, A. Oxidative stress and some antioxidant systems in acid rain-treated bean plants: Protective role of exogenous polyamines. Plant Sci. 2000, 151, 59–66. [Google Scholar] [CrossRef]
- Liu, D.; Zou, J.; Meng, Q.; Zou, J.; Jiang, W. Uptake and accumulation and oxidative stress in garlic (Allium sativum L.) under lead phytotoxicity. Ecotoxicology 2009, 18, 134–143. [Google Scholar] [CrossRef] [PubMed]
- Arnon, D.I. Copper enzymes in isolated chloroplasts. Polyphenoloxidase in Beta vulgaris. Plant Physiol. 1949, 24, 1. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gür, A.; Demirel, U.; Özden, M.; Kahraman, A.; Çopur, O. Diurnal gradual heat stress affects antioxidant enzymes, proline accumulation and some physiological components in cotton (Gossypium hirsutum L.). Afr. J. Biotechnol. 2010, 9, 1008–1015. [Google Scholar]
- Majeed, S.; Malik, T.A.; Rana, I.A.; Azhar, M.T. Antioxidant and physiological responses of upland cotton accessions grown under high-temperature regimes. Iran. J. Sci. Technol. Trans. A Sci. 2019, 43, 2759–2768. [Google Scholar] [CrossRef]
- Rana, R.; Khan, S.; Ali, Z.; Khan, A.; Khan, I. Elucidation of thermotolerance diversity in cotton (Gossypium hirsutum L.) using physio-molecular approaches. Genet. Mol. Res. 2011, 10, 1156–1167. [Google Scholar] [CrossRef] [PubMed]
- Ali, S.; Khan, T.M.; Shakeel, A.; Saleem, M.F. Genetic basis of variation for physiological and yield contributing traits under normal and high temperature stress in Gossypium hirsutum L. Pak. J. Agric. Sci. 2020, 57, 1491–1501. [Google Scholar]
- Joseph, B.; Jini, D.; Sujatha, S. Development of salt stress-tolerant plants by gene manipulation of antioxidant enzymes. Asian J. Agric. Res. 2011, 5, 17–27. [Google Scholar] [CrossRef] [Green Version]
- Jiang, Y.; Huang, B. Drought and heat stress injury to two cool-season turfgrasses in relation to antioxidant metabolism and lipid peroxidation. Crop. Sci. 2001, 41, 436–442. [Google Scholar] [CrossRef]
- Phung, T.-H.; Jung, S. Alterations in the porphyrin biosynthesis and antioxidant responses to chilling and heat stresses in Oryza sativa. Biol. Plant. 2015, 59, 341–349. [Google Scholar] [CrossRef]
- Shu, S.; Yuan, L.-Y.; Guo, S.-R.; Sun, J.; Yuan, Y.-H. Effects of exogenous spermine on chlorophyll fluorescence, antioxidant system and ultrastructure of chloroplasts in Cucumis sativus L. under salt stress. Plant Physiol. Biochem. 2013, 63, 209–216. [Google Scholar] [CrossRef]
- Chalanika De Silva, H.C.; Asaeda, T. Effects of heat stress on growth, photosynthetic pigments, oxidative damage and competitive capacity of three submerged macrophytes. J. Plant Interact. 2017, 12, 228–236. [Google Scholar] [CrossRef] [Green Version]
- Chapepa, B.; Mubvekeri, W.; Mare, M.; Kutywayo, D. Correlation and path coefficient analysis of polygenic traits of upland cotton genotypes grown in Zimbabwe. Cogent Food Agric. 2020, 6, 1823594. [Google Scholar] [CrossRef]
- Khalid, H.; Khan, I.A.; Sadaqat, H.A.; Muhammad, A. Genotypic and phenotypic correlation analysis of yield and fiber quality determining traits in upland cotton (Gossypim hirsutum). Int. J. Agric. Biol. 2010, 12, 348–352. [Google Scholar]
- Malik, T.A. Correlation for economic traits in upland cotton. ACTA Sci. Agric. 2018, 2, 59–62. [Google Scholar]
- Chunthaburee, S.; Dongsansuk, A.; Sanitchon, J.; Pattanagul, W.; Theerakulpisut, P. Physiological and biochemical parameters for evaluation and clustering of rice cultivars differing in salt tolerance at seedling stage. Saudi J. Biol. Sci. 2016, 23, 467–477. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jamil, A.; Khan, S.J.; Ullah, K. Genetic diversity for cell membrane thermostability, yield and quality attributes in cotton (Gossypium hirsutum L.). Genet. Resour. Crop. Evol. 2020, 67, 1405–1414. [Google Scholar] [CrossRef]
- Saeed, F.; Shabbir, R.H.; Farooq, J.; Riaz, M.; Mahmood, K. Genetic Diversity Analysis for Earliness, Fiber Quality and Cotton Leaf Curl Virus in Gossypium hirsutum L. Accessions. Cotton Genom. Genet. 2015, 6, 1–7. [Google Scholar]
- Shabbir, R.H.; Bashir, Q.A.; Shakeel, A.; Khan, M.M.; Farooq, J.; Fiaz, S.; Ijaz, B.; Noor, M.A. Genetic divergence assessment in upland cotton (Gossypium hirsutum L.) using various statistical tools. J. Glob. Innov. Agric. Soc. Sci. 2016, 4, 62–69. [Google Scholar] [CrossRef]
- Amna, N.; Jehanzeb, F.; Abid, M.; Muhammad, S.; Muhammad, R. Estimation of genetic diversity for CLCuV, earliness and fiber quality traits using various statistical procedures in different crosses of Gossypium hirsutum L. Вестник Аграрнoй Науки 2013, 43, 2–9. [Google Scholar]
- Isong, A.; Balu, P.A.; Ramakrishnan, P. Association and principal component analysis of yield and its components in cultivated cotton. Electron. J. Plant Breed. 2017, 8, 857–864. [Google Scholar] [CrossRef]
- Javed, M.; Hussain, S.; Baber, M. Assessment of genetic diversity of cotton genotypes for various economic traits against cotton leaf curl disease (CLCuD). Genet. Mol. Res. 2017, 16, 1–12. [Google Scholar] [CrossRef]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Traits Source | Genotype | Treatment | Genotype * Treatment | |
|---|---|---|---|---|
| Nparm | 22 | 1 | 22 | |
| DFNum | 22 | 1 | 22 | |
| PH | F Ratio Prob > F | 14.2846 <0.0001 * | 106.3898 <0.0001 * | 1.8302 0.0429 * |
| TNB | F Ratio Prob > F | 8.8080 <0.0001 * | 357.3079 <0.0001 * | 3.5284 0.0002 * |
| BW | F Ratio Prob > F | 8.9352 <0.0001 * | 12.5570 0.0009 * | 2.2602 0.0102 * |
| SCY | F Ratio Prob > F | 21.8997 <0.0001 * | 426.5928 <0.0001 * | 4.1679 <0.0001 * |
| H2O2 | F Ratio Prob > F | 9.6527 <0.0001 * | 98.0262 <0.0001 * | 2.4577 0.0053 * |
| POD | F Ratio Prob > F | 34.9447 <0.0001 * | 5496.441 <0.0001 * | 18.7102 <0.0001 * |
| CAT | F Ratio Prob > F | 54.9865 <0.0001 * | 16325.80 <0.0001 * | 15.0780 <0.0001 * |
| Chla | F Ratio Prob > F | 20.3758 <0.0001 * | 1484.163 <0.0001 * | 8.9729 <0.0001 * |
| Chlb | F Ratio Prob > F | 16.6234 <0.0001 * | 559.5363 <0.0001 * | 6.4504 <0.0001 * |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zafar, M.M.; Manan, A.; Razzaq, A.; Zulfqar, M.; Saeed, A.; Kashif, M.; Khan, A.I.; Sarfraz, Z.; Mo, H.; Iqbal, M.S.; et al. Exploiting Agronomic and Biochemical Traits to Develop Heat Resilient Cotton Cultivars under Climate Change Scenarios. Agronomy 2021, 11, 1885. https://doi.org/10.3390/agronomy11091885
Zafar MM, Manan A, Razzaq A, Zulfqar M, Saeed A, Kashif M, Khan AI, Sarfraz Z, Mo H, Iqbal MS, et al. Exploiting Agronomic and Biochemical Traits to Develop Heat Resilient Cotton Cultivars under Climate Change Scenarios. Agronomy. 2021; 11(9):1885. https://doi.org/10.3390/agronomy11091885
Chicago/Turabian StyleZafar, Muhammad Mubashar, Abdul Manan, Abdul Razzaq, Misbah Zulfqar, Asif Saeed, Muhammad Kashif, Azeem Iqbal Khan, Zareen Sarfraz, Huijuan Mo, Muhammad Shahid Iqbal, and et al. 2021. "Exploiting Agronomic and Biochemical Traits to Develop Heat Resilient Cotton Cultivars under Climate Change Scenarios" Agronomy 11, no. 9: 1885. https://doi.org/10.3390/agronomy11091885
APA StyleZafar, M. M., Manan, A., Razzaq, A., Zulfqar, M., Saeed, A., Kashif, M., Khan, A. I., Sarfraz, Z., Mo, H., Iqbal, M. S., Shakeel, A., & Ren, M. (2021). Exploiting Agronomic and Biochemical Traits to Develop Heat Resilient Cotton Cultivars under Climate Change Scenarios. Agronomy, 11(9), 1885. https://doi.org/10.3390/agronomy11091885