
Characterizing Genotype-Specific Rice Architectural Traits Using Smart Mobile App and Data Modeling

 ,
, 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Experiment Design and Data Collection
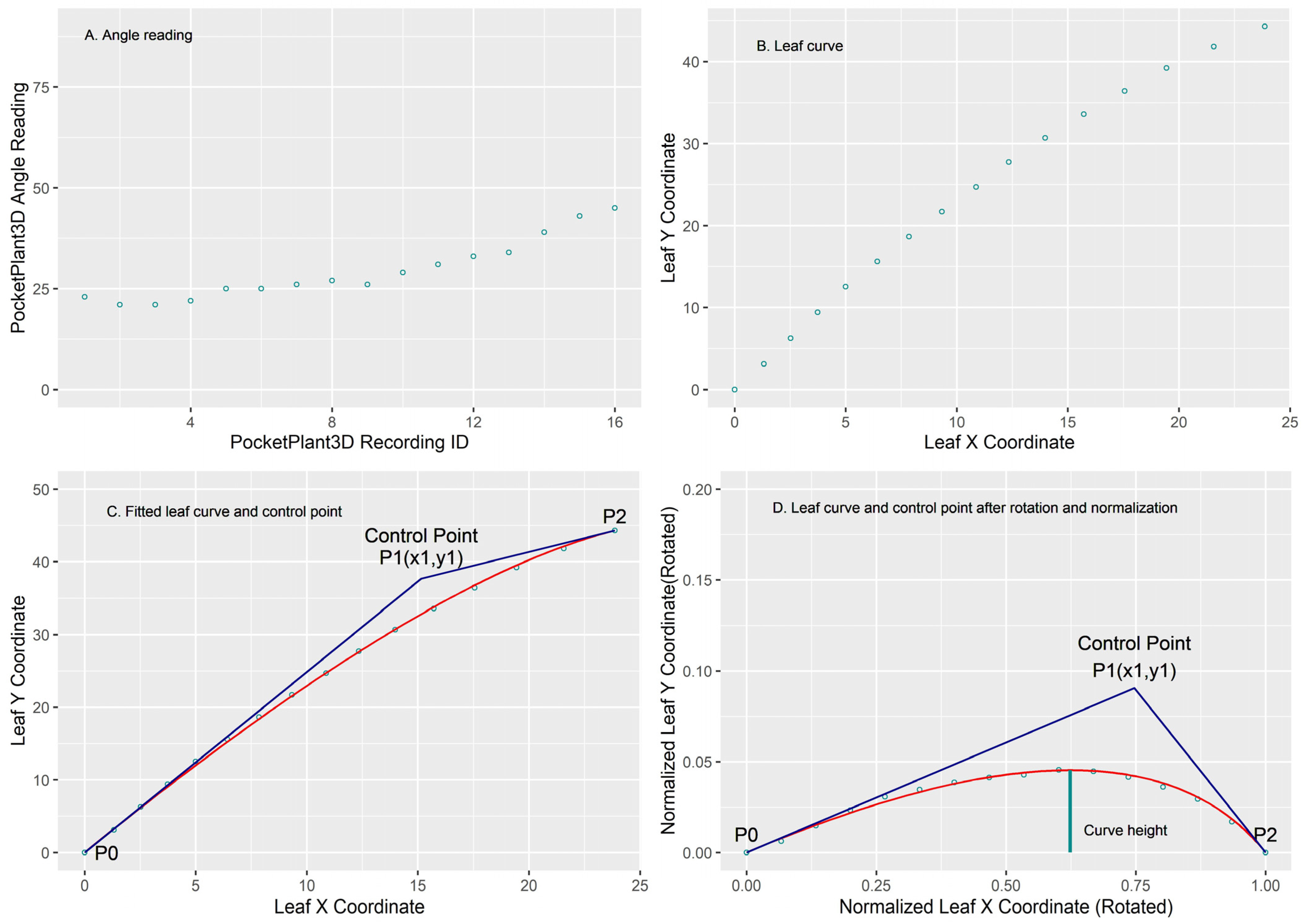
2.2. Calculation of Leaf Insertion Angle, Elevation Angle, and Curve Height
2.3. Leaf Azimuth Distribution and Leaf Phyllotaxis
2.4. Canopy Leaf Angle Distribution and Light Extinction Coefficient
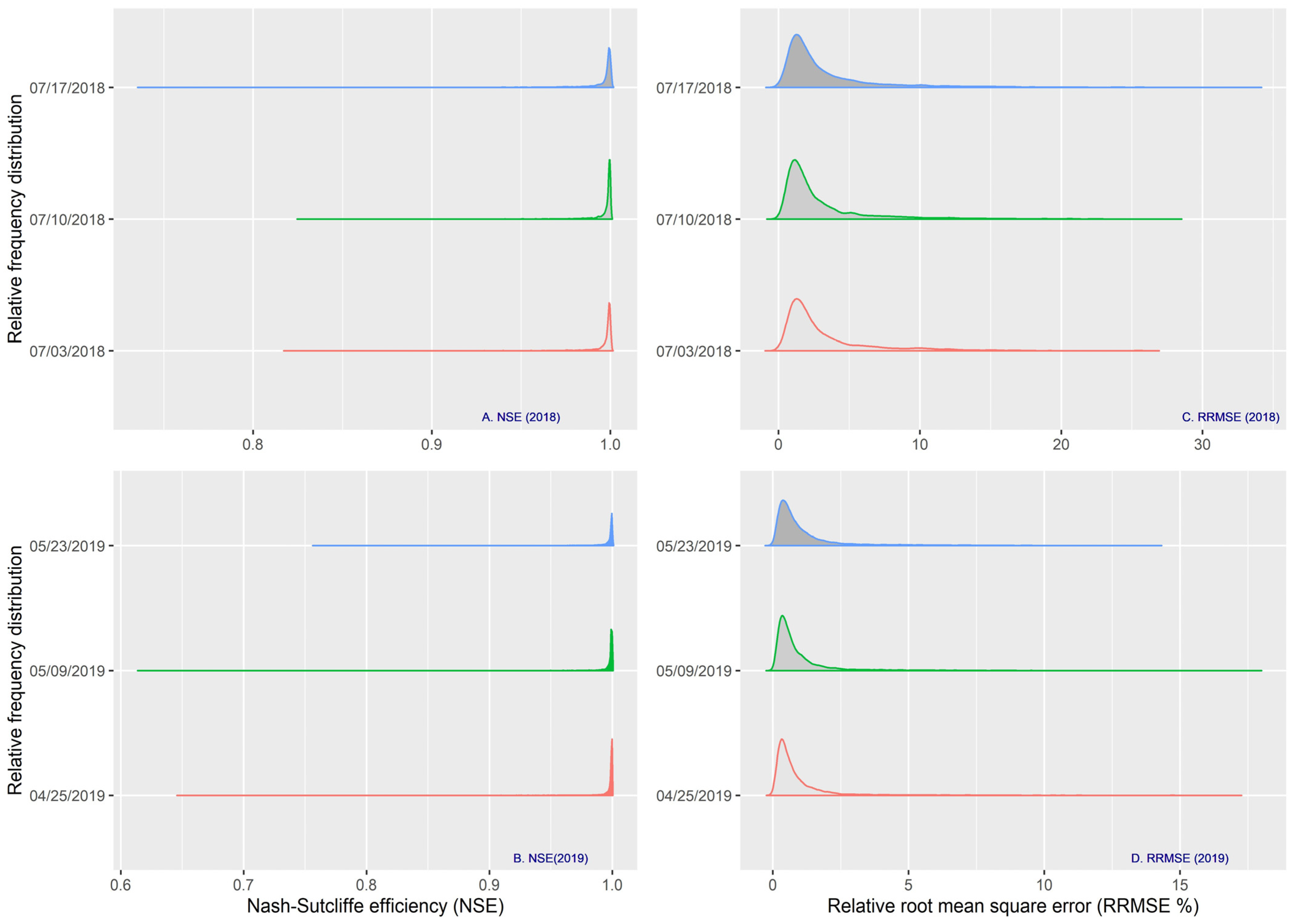
2.5. Curve Fitting and Statistical Analysis
3. Results
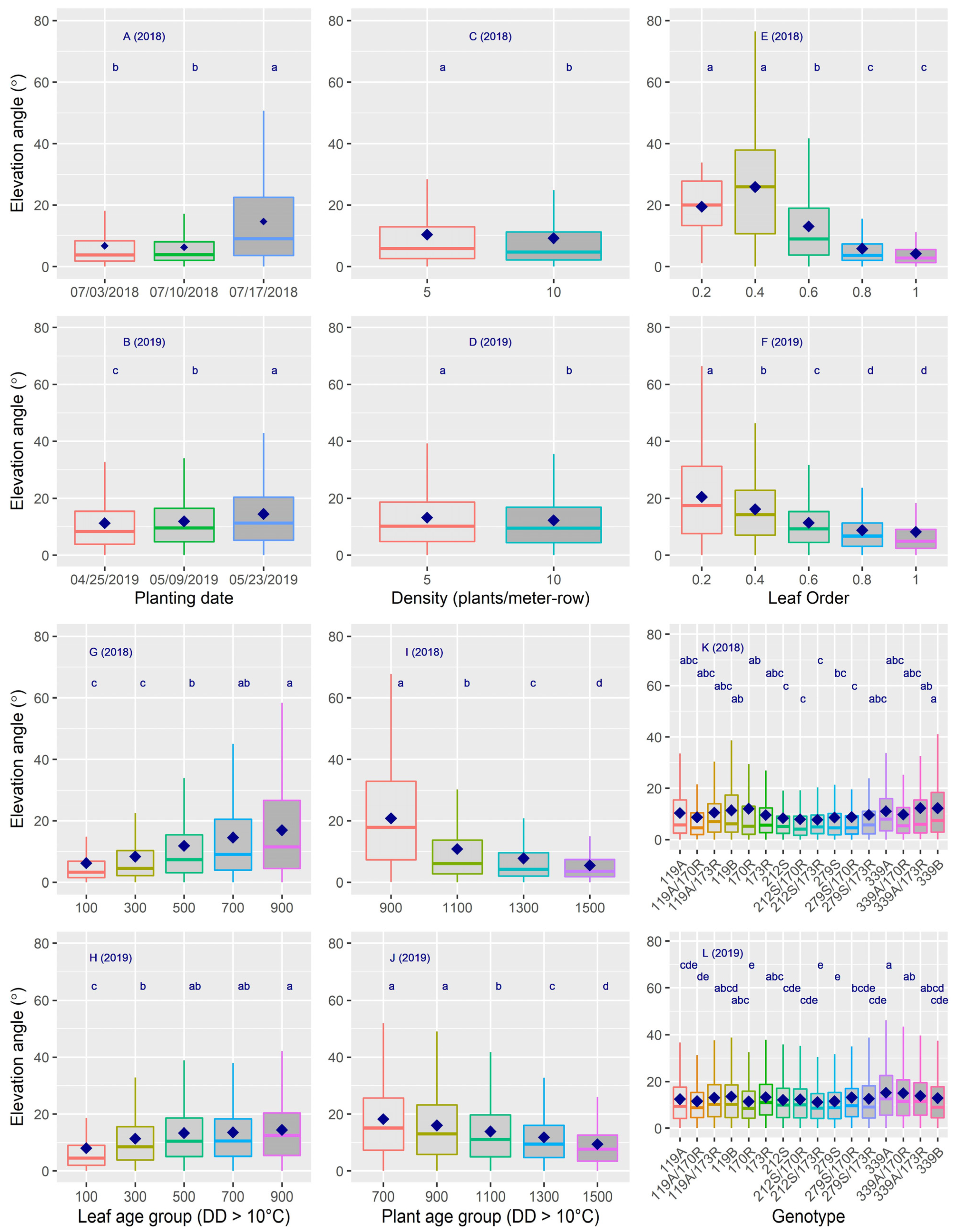
3.1. Analysis of Variance on Leaf Insertion Angle and Leaf Curvature (Elevation Angle and Curve Height)
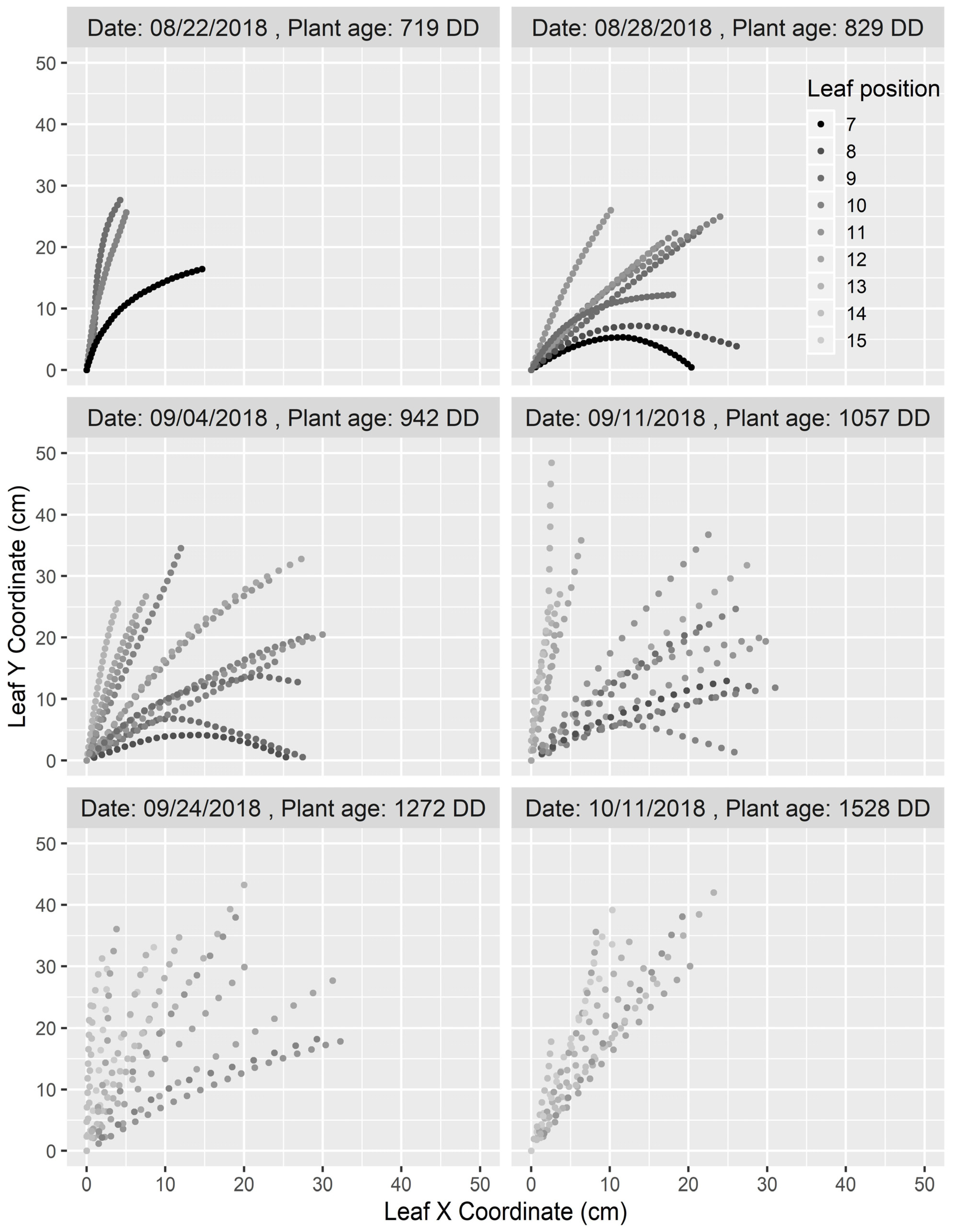
3.2. Dynamics of Leaf Insertion Angle
3.3. Dynamics of Leaf Elevation Angle
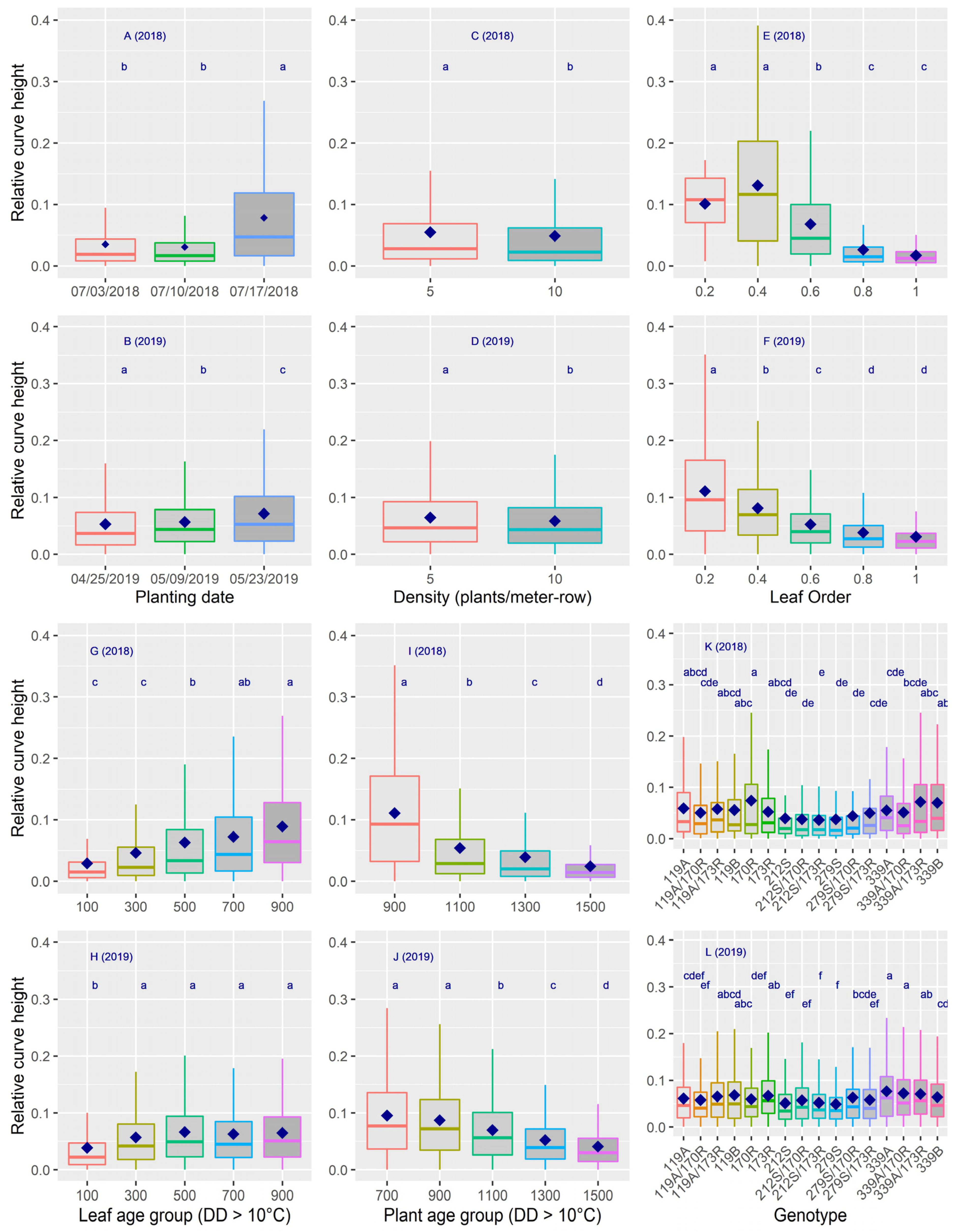
3.4. Dynamics of Curve Height
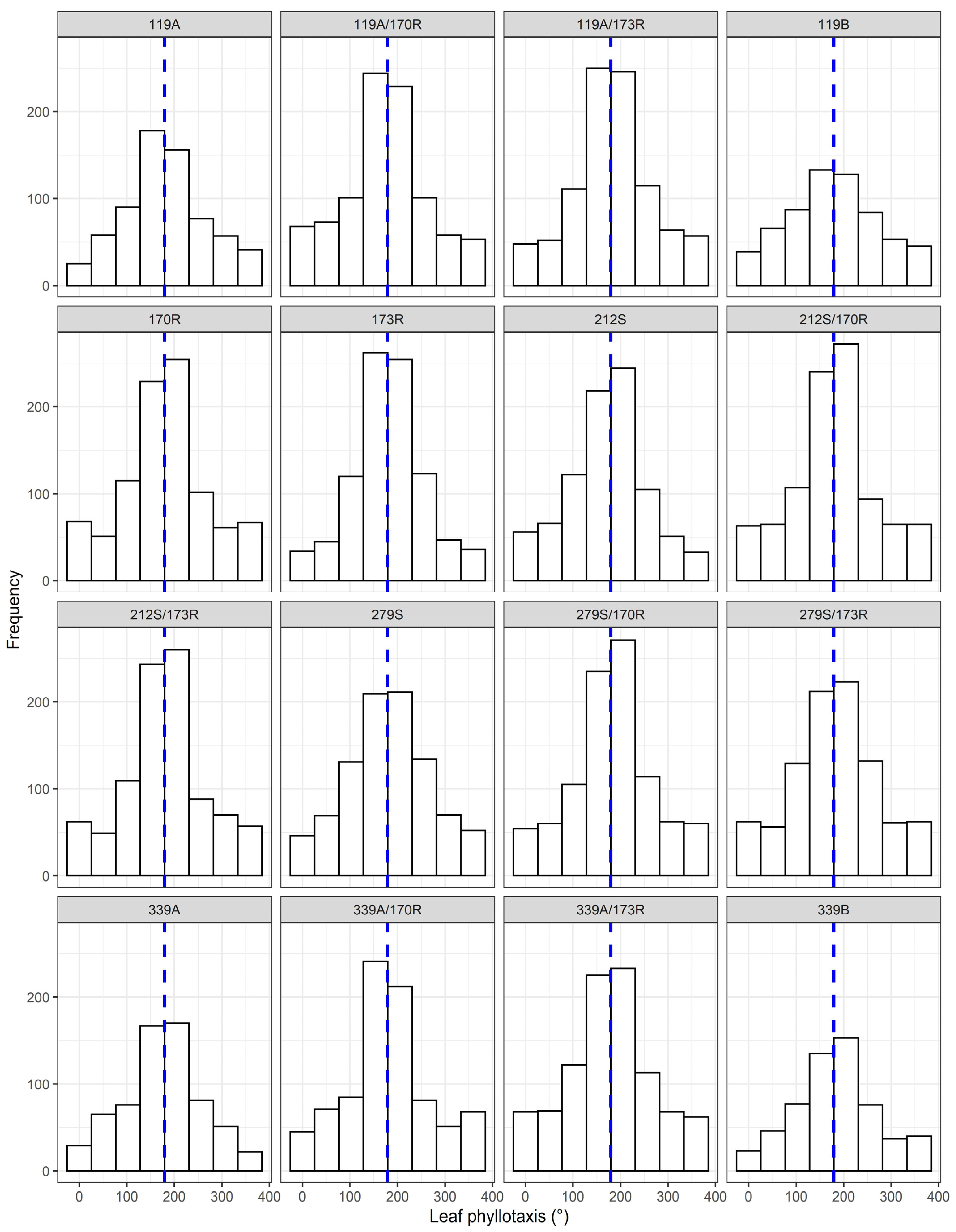
3.5. Leaf Azimuth Distribution and Leaf Phyllotaxis
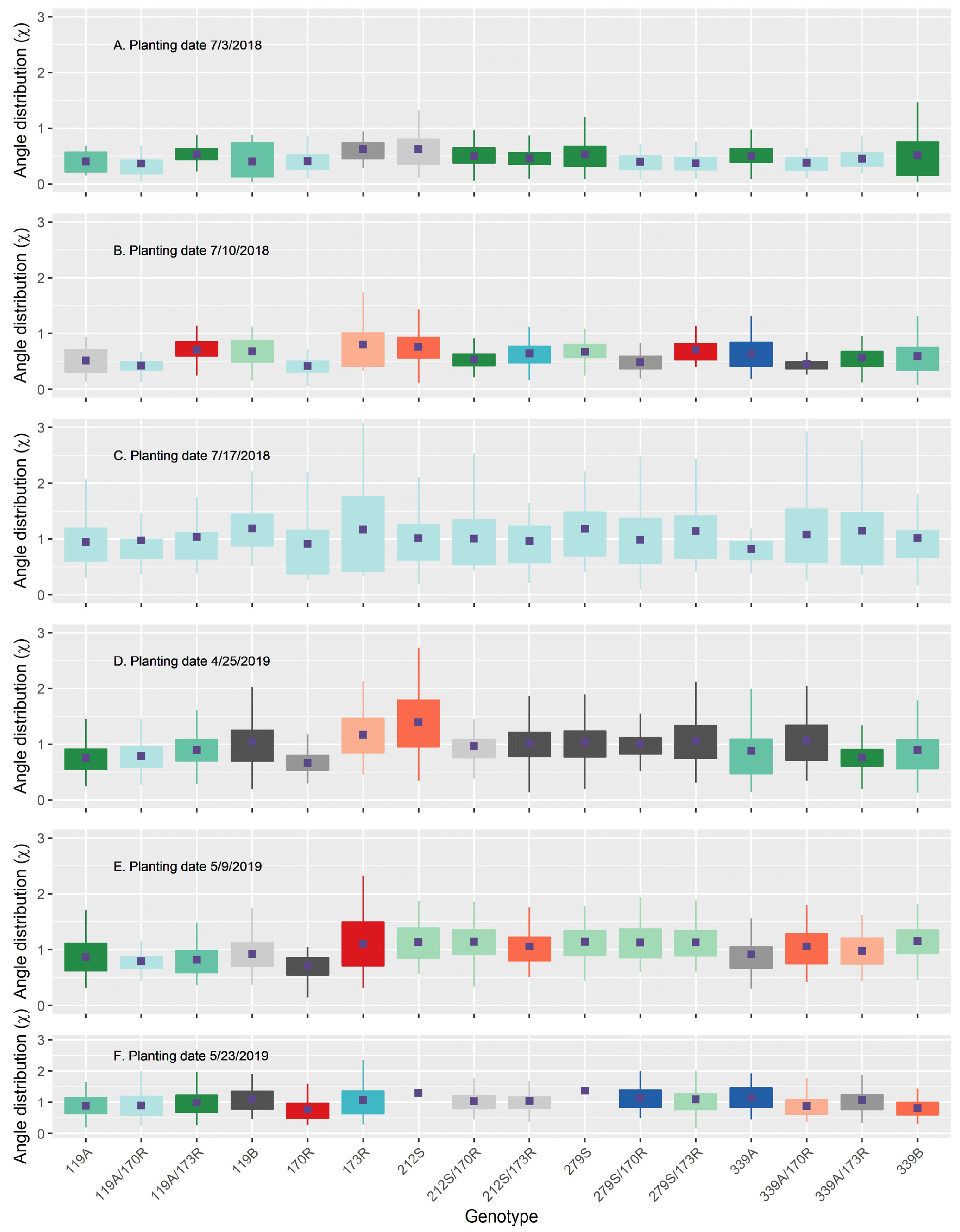
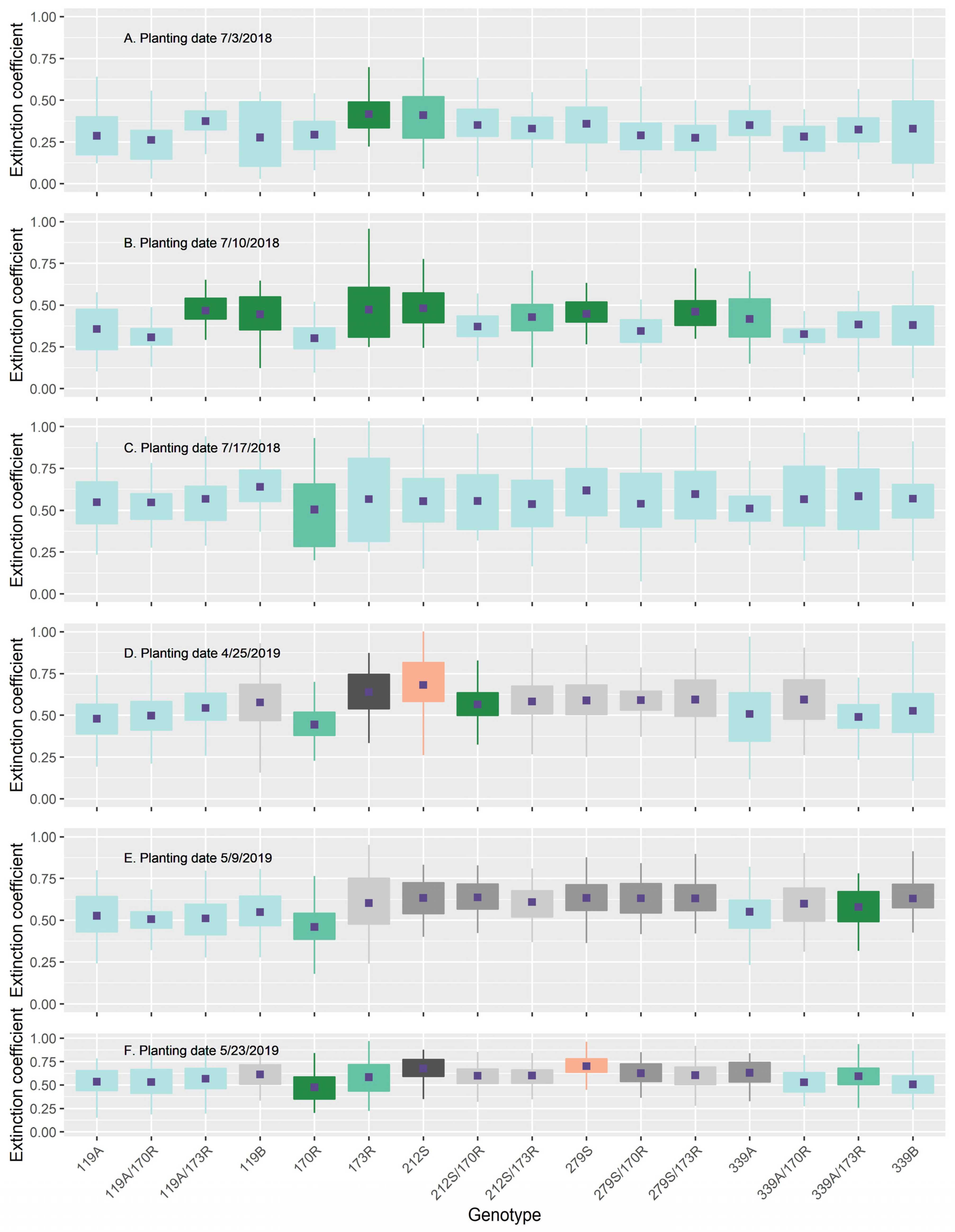
3.6. Canopy Leaf Angle Distribution and Light Extinction Coefficient
4. Discussion
4.1. Rice Leaf Curvature Modeling
4.2. Leaf Angle Dynamics and Distribution Pattern
4.3. Canopy Leaf Angle Distribution and Light Capture
4.4. Leaf Area Index Estimation through Leaf Curvature Modeling and Leaf Area Integration
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Evers, J.B.; Marcelis, L.F.M. Functional—Structural plant modeling of plants and crops. In Improving Organic Animal Farming; Burleigh Dodds Science Publishing: Cambridge, MA, USA, 2019; Volume 75, pp. 45–68. [Google Scholar]
- Küpers, J.; van Gelderen, K.; Pierik, R. Location Matters: Canopy Light Responses over Spatial Scales. Trends Plant Sci. 2018, 23, 865–873. [Google Scholar] [CrossRef]
- Huber, M.; Nieuwendijk, N.M.; Pantazopoulou, C.K.; Pierik, R. Light signalling shapes plant–plant interactions in dense canopies. Plant Cell Environ. 2021, 44, 1014–1029. [Google Scholar] [CrossRef]
- Casal, J.J. Photoreceptor signaling networks in plant responses to shade. In Annual Review of Plant Biology; Merchant, S.S., Ed.; Annual Reviews: Palo Alto, CA, USA, 2013; Volume 64, pp. 403–427. [Google Scholar]
- Demotes-Mainard, S.; Péron, T.; Corot, A.; Bertheloot, J.; Le Gourrierec, J.; Pelleschi-Travier, S.; Crespel, L.; Morel, P.; Huché-Thélier, L.; Boumaza, R.; et al. Plant responses to red and far-red lights, applications in horticulture. Environ. Exp. Bot. 2016, 121, 4–21. [Google Scholar] [CrossRef]
- De Kroon, H.; Huber, H.; Stuefer, J.F.; Van Groenendael, J.M. A modular concept of phenotypic plasticity in plants. New Phytol. 2005, 166, 73–82. [Google Scholar] [CrossRef]
- Chelle, M.; Evers, J.B.; Combes, D.; Varlet-Grancher, C.; Vos, J.; Andrieu, B. Simulation of the three-dimensional distribution of the red:far-red ratio within crop canopies. New Phytol. 2007, 176, 223–234. [Google Scholar] [CrossRef] [PubMed]
- Hoogenboom, G.; Porter, C.H.; Boote, K.J.; Shelia, V.; Wilkens, P.W.; Singh, U.; White, J.W.; Asseng, S.; Lizaso, J.I.; Moreno, L.P.; et al. The DSSAT crop modeling ecosystem. In Advances in Crop Modeling for a Sustainable Agriculture; Boote, K.J., Ed.; Burleigh Dodds Science Publishing: Cambridge, UK, 2019; pp. 173–216. [Google Scholar] [CrossRef]
- Jones, J.W.; Hoogenboom, G.; Porter, C.H.; Boote, K.J.; Batchelor, W.D.; Hunt, L.A.; Wilkens, P.W.; Singh, U.; Gijsman, A.J.; Ritchie, J.T. The DSSAT cropping system model. Eur. J. Agron. 2003, 18, 235–265. [Google Scholar] [CrossRef]
- Holzworth, D.P.; Huth, N.I.; Devoil, P.G.; Zurcher, E.; Herrmann, N.; McLean, G.; Chenu, K.; van Oosterom, E.J.; Snow, V.; Murphy, C.; et al. APSIM–Evolution towards a new generation of agricultural systems simulation. Environ. Model. Softw. 2014, 62, 327–350. [Google Scholar] [CrossRef]
- Evers, J.B. Simulating crop growth and development using functional-structural plant modeling. In Canopy Photosynthesis: From Basics to Applications; Hikosaka, K., Niinemets, U., Anten, N.P.R., Eds.; Springer: Dordrecht, The Netherlands, 2016; Volume 42, pp. 219–236. [Google Scholar]
- Auzmendi, I.; Hanan, J.S. Investigating tree and fruit growth through functional–structural modelling: Implications of carbon autonomy at different scales. Ann. Bot. 2020, 126, 775–788. [Google Scholar] [CrossRef]
- Louarn, G.; Song, Y. Two decades of functional-structural plant modelling: Now addressing fundamental questions in systems biology and predictive ecology. Ann. Bot. 2020, 126, 501–509. [Google Scholar] [CrossRef]
- Prusinkiewicz, P.; Lindenmayer, A. The Algorithmic Beauty of Plants; Springer: New York, NY, USA, 1991; p. 228. [Google Scholar]
- Fournier, C.; Andrieu, B. A 3d architectural and process-based model of maize development. Ann. Bot. 1998, 81, 233–250. [Google Scholar] [CrossRef] [Green Version]
- Room, P.; Hanan, J.; Prusinkiewicz, P. Virtual plants: New perspectives for ecologists, pathologists and agricultural scientists. Trends Plant Sci. 1996, 1, 33–38. [Google Scholar] [CrossRef]
- Reyes, F.; Pallas, B.; Pradal, C.; Vaggi, F.; Zanotelli, D.; Tagliavini, M.; Gianelle, D.; Costes, E. MuSCA: A multi-scale source–sink carbon allocation model to explore carbon allocation in plants. An application to static apple tree structures. Ann. Bot. 2020, 126, 571–585. [Google Scholar] [CrossRef] [PubMed]
- Zhang, B.; DeAngelis, D.L. An overview of agent-based models in plant biology and ecology. Ann. Bot. 2020, 126, 539–557. [Google Scholar] [CrossRef] [PubMed]
- DeJong, T.M.; Da Silva, D.; Vos, J.; Gutierrez, A.E. Using functional–structural plant models to study, understand and integrate plant development and ecophysiology. Ann. Bot. 2011, 108, 987–989. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sievänen, R.; Godin, C.; DeJong, T.M.; Nikinmaa, E. Functional–structural plant models: A growing paradigm for plant studies. Ann. Bot. 2014, 114, 599–603. [Google Scholar] [CrossRef]
- Peng, B.; Guan, K.; Tang, J.; Ainsworth, E.A.; Asseng, S.; Bernacchi, C.J.; Cooper, M.; Delucia, E.H.; Elliott, J.W.; Ewert, F.; et al. Towards a multiscale crop modelling framework for climate change adaptation assessment. Nat. Plants 2020, 6, 338–348. [Google Scholar] [CrossRef]
- Yin, X.; van der Linden, C.G.; Struik, P.C. Bringing genetics and biochemistry to crop modelling, and vice versa. Eur. J. Agron. 2018, 100, 132–140. [Google Scholar] [CrossRef] [Green Version]
- Jang, S.; An, G.; Li, H.-Y. Rice Leaf Angle and Grain Size Are Affected by the OsBUL1 Transcriptional Activator Complex. Plant Physiol. 2017, 173, 688–702. [Google Scholar] [CrossRef]
- Wang, R.; Liu, C.; Li, Q.; Chen, Z.; Sun, S.; Wang, X. Spatiotemporal Resolved Leaf Angle Establishment Improves Rice Grain Yield via Controlling Population Density. iScience 2020, 23, 101489. [Google Scholar] [CrossRef]
- Zhao, S.-Q.; Hu, J.; Guo, L.-B.; Qian, Q.; Xue, H.-W. Rice leaf inclination2, a VIN3-like protein, regulates leaf angle through modulating cell division of the collar. Cell Res. 2010, 20, 935–947. [Google Scholar] [CrossRef] [Green Version]
- Zhu, Y.; Chang, L.; Tang, L.; Jiang, H.; Zhang, W.; Cao, W. Modelling leaf shape dynamics in rice. NJAS-Wagening. J. Life Sci. 2009, 57, 73–81. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Y.; Tang, L.; Liu, X.; Liu, L.; Cao, W.; Zhu, Y. Modeling the leaf angle dynamics in rice plant. PLoS ONE 2017, 12, e0171890. [Google Scholar]
- Zhang, Y.-H.; Tang, L.; Liu, X.-J.; Liu, L.-L.; Cao, W.-X.; Zhu, Y. Modeling curve dynamics and spatial geometry characteristics of rice leaves. J. Integr. Agric. 2017, 16, 2177–2190. [Google Scholar] [CrossRef] [Green Version]
- Duncan, W.G. Leaf Angles, Leaf Area, and Canopy Photosynthesis 1. Crop. Sci. 1971, 11, 482–485. [Google Scholar] [CrossRef]
- Weiss, M.; Baret, F.; Smith, G.J.; Jonckheere, I.; Coppin, P. Review of methods for in situ leaf area index (lai) determination part ii. Estimation of lai, errors and sampling. Agric. For. Meteorol. 2004, 121, 37–53. [Google Scholar] [CrossRef]
- Sarlikioti, V.; De Visser, P.H.B.; Buck-Sorlin, G.H.; Marcelis, L.F.M. How plant architecture affects light absorption and photosynthesis in tomato: Towards an ideotype for plant architecture using a functional–structural plant model. Ann. Bot. 2011, 108, 1065–1073. [Google Scholar] [CrossRef] [Green Version]
- Zou, X.; Mõttus, M.; Tammeorg, P.; Torres, C.L.; Takala, T.; Pisek, J.; Mäkelä, P.; Stoddard, F.; Pellikka, P. Photographic measurement of leaf angles in field crops. Agric. For. Meteorol. 2014, 184, 137–146. [Google Scholar] [CrossRef] [Green Version]
- Mantilla-Perez, M.B.; Fernandez, M.G.S. Differential manipulation of leaf angle throughout the canopy: Current status and prospects. J. Exp. Bot. 2017, 68, 5699–5717. [Google Scholar] [CrossRef] [Green Version]
- Lang, A. Leaf-Area and Average Leaf Angle from Transmission of Direct Sunlight. Aust. J. Bot. 1986, 34, 349–355. [Google Scholar] [CrossRef]
- Sinoquet, H.; Thanisawanyangkura, S.; Mabrouk, H.; Kasemsap, P. Characterization of the Light Environment in Canopies Using 3D Digitising and Image Processing. Ann. Bot. 1998, 82, 203–212. [Google Scholar] [CrossRef] [Green Version]
- Watanabe, T.; Hanan, J.S.; Room, P.M.; Hasegawa, T.; Nakagawa, H.; Takahashi, W. Rice Morphogenesis and Plant Architecture: Measurement, Specification and the Reconstruction of Structural Development by 3D Architectural Modelling. Ann. Bot. 2005, 95, 1131–1143. [Google Scholar] [CrossRef] [Green Version]
- Sasidharan, R.; Chinnappa, C.C.; Staal, M.; Elzenga, J.T.M.; Yokoyama, R.; Nishitani, K.; Voesenek, L.; Pierik, R. Light quality-mediated petiole elongation in arabidopsis during shade avoidance involves cell wall modification by xyloglucan endotransglucosylase/hydrolases. Plant Physiol. 2010, 154, 978–990. [Google Scholar] [CrossRef] [Green Version]
- Wang, H.; Zhang, W.; Zhou, G.; Yan, G.; Clinton, N. Image-based 3D corn reconstruction for retrieval of geometrical structural parameters. Int. J. Remote Sens. 2009, 30, 5505–5513. [Google Scholar] [CrossRef]
- Pisek, J.; Ryu, Y.; Alikas, K. Estimating leaf inclination and G-function from leveled digital camera photography in broadleaf canopies. Trees 2011, 25, 919–924. [Google Scholar] [CrossRef]
- Raabe, K.; Pisek, J.; Sonnentag, O.; Annuk, K. Variations of leaf inclination angle distribution with height over the growing season and light exposure for eight broadleaf tree species. Agric. For. Meteorol. 2015, 214–215, 2–11. [Google Scholar] [CrossRef]
- Zhang, X.-C.; Lu, C.-G.; Hu, N.; Yao, K.-M.; Zhang, Q.-J.; Dai, Q.-G. Simulation of Canopy Leaf Inclination Angle in Rice. Rice Sci. 2013, 20, 434–441. [Google Scholar] [CrossRef]
- Itakura, K.; Hosoi, F. Automatic Leaf Segmentation for Estimating Leaf Area and Leaf Inclination Angle in 3D Plant Images. Sensors 2018, 18, 3576. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Qi, J.; Xie, D.; Li, L.; Zhang, W.; Mu, X.; Yan, G. Estimating Leaf Angle Distribution from Smartphone Photographs. IEEE Geosci. Remote Sens. Lett. 2019, 16, 1190–1194. [Google Scholar] [CrossRef]
- Raju, S.K.K.; Adkins, M.; Enersen, A.; De Carvalho, D.S.; Studer, A.J.; Ganapathysubramanian, B.; Schnable, P.S.; Schnable, J.C. Leaf Angle eXtractor: A high-throughput image processing framework for leaf angle measurements in maize and sorghum. Appl. Plant Sci. 2020, 8, e11385. [Google Scholar]
- Zhang, Y.; Teng, P.; Aono, M.; Shimizu, Y.; Hosoi, F.; Omasa, K. 3D monitoring for plant growth parameters in field with a single camera by multi-view approach. J. Agric. Meteorol. 2018, 74, 129–139. [Google Scholar] [CrossRef] [Green Version]
- Shaaf, S.; Bretani, G.; Biswas, A.; Fontana, I.M.; Rossini, L. Genetics of barley tiller and leaf development. J. Integr. Plant Biol. 2018, 61, 226–256. [Google Scholar] [CrossRef] [Green Version]
- Yang, W.; Feng, H.; Zhang, X.; Zhang, J.; Doonan, J.H.; Batchelor, W.D.; Xiong, L.; Yan, J. Crop Phenomics and High-Throughput Phenotyping: Past Decades, Current Challenges, and Future Perspectives. Mol. Plant 2020, 13, 187–214. [Google Scholar] [CrossRef] [Green Version]
- Gibbs, J.A.; Pound, M.P.; French, A.; Wells, D.; Murchie, E.; Pridmore, T.P. Approaches to three-dimensional reconstruction of plant shoot topology and geometry. Funct. Plant Biol. 2017, 44, 62–75. [Google Scholar] [CrossRef] [Green Version]
- Paulus, S.; Schumann, H.; Kuhlmann, H.; Léon, J. High-precision laser scanning system for capturing 3D plant architecture and analysing growth of cereal plants. Biosyst. Eng. 2014, 121, 1–11. [Google Scholar] [CrossRef]
- Bailey, B.N.; Mahaffee, W.F. Rapid measurement of the three-dimensional distribution of leaf orientation and the leaf angle probability density function using terrestrial LiDAR scanning. Remote Sens. Environ. 2017, 194, 63–76. [Google Scholar] [CrossRef] [Green Version]
- Itakura, K.; Hosoi, F. Estimation of Leaf Inclination Angle in Three-Dimensional Plant Images Obtained from Lidar. Remote Sens. 2019, 11, 344. [Google Scholar] [CrossRef] [Green Version]
- Hosoi, F.; Nakai, Y.; Omasa, K. Estimating the leaf inclination angle distribution of the wheat canopy using a portable scanning lidar. J. Agric. Meteorol. 2009, 65, 297–302. [Google Scholar] [CrossRef] [Green Version]
- Hosoi, F.; Omasa, K. Estimation of vertical plant area density profiles in a rice canopy at different growth stages by high-resolution portable scanning lidar with a lightweight mirror. ISPRS J. Photogramm. Remote Sens. 2012, 74, 11–19. [Google Scholar] [CrossRef]
- Stovall, A.E.L.; Masters, B.; Fatoyinbo, L.; Yang, X. Tlsleaf: Automatic leaf angle estimates from single-scan terrestrial laser scanning. New Phytol. 2021, 232, 1876–1892. [Google Scholar] [CrossRef]
- Vicari, M.B.; Pisek, J.; Disney, M. New estimates of leaf angle distribution from terrestrial LiDAR: Comparison with measured and modelled estimates from nine broadleaf tree species. Agric. For. Meteorol. 2019, 264, 322–333. [Google Scholar] [CrossRef]
- McNeil, B.E.; Pisek, J.; Lepisk, H.; Flamenco, E.A. Measuring leaf angle distribution in broadleaf canopies using UAVs. Agric. For. Meteorol. 2016, 218, 204–208. [Google Scholar] [CrossRef]
- Nevalainen, O.; Honkavaara, E.; Tuominen, S.; Viljanen, N.; Hakala, T.; Yu, X.; Hyyppä, J.; Saari, H.; Pölönen, I.; Imai, N.N.; et al. Individual Tree Detection and Classification with UAV-Based Photogrammetric Point Clouds and Hyperspectral Imaging. Remote Sens. 2017, 9, 185. [Google Scholar] [CrossRef] [Green Version]
- Xu, S.; Zaidan, M.A.; Honkavaara, E.; Hakala, T.; Viljanen, N.; Porcar-Castell, A.; Liu, Z.G.; Atherton, J. On the Estimation of the Leaf Angle Distribution from Drone Based Photogrammetry. In Proceedings of the IEEE International Geoscience and Remote Sensing Symposium (IGARSS), Waikoloa, HI, USA, 26 September–2 October 2020; pp. 4379–4382. [Google Scholar]
- Huang, W.; Niu, Z.; Wang, J.; Liu, L.; Zhao, C.; Liu, Q. Identifying Crop Leaf Angle Distribution Based on Two-Temporal and Bidirectional Canopy Reflectance. IEEE Trans. Geosci. Remote Sens. 2006, 44, 3601–3609. [Google Scholar] [CrossRef]
- Zhao, K.; Garcia, M.; Liu, S.; Guo, Q.; Chen, G.; Zhang, X.; Zhou, Y.; Meng, X. Terrestrial lidar remote sensing of forests: Maximum likelihood estimates of canopy profile, leaf area index, and leaf angle distribution. Agric. For. Meteorol. 2015, 209–210, 100–113. [Google Scholar] [CrossRef]
- Dassot, M.; Constant, T.; Fournier, M. The use of terrestrial LiDAR technology in forest science: Application fields, benefits and challenges. Ann. For. Sci. 2011, 68, 959–974. [Google Scholar] [CrossRef] [Green Version]
- Liu, J.; Skidmore, A.K.; Wang, T.; Zhu, X.; Premier, J.; Heurich, M.; Beudert, B.; Jones, S. Variation of leaf angle distribution quantified by terrestrial LiDAR in natural European beech forest. ISPRS J. Photogramm. Remote Sens. 2019, 148, 208–220. [Google Scholar] [CrossRef]
- Confalonieri, R.; Paleari, L.; Foi, M.; Movedi, E.; Vesely, F.M.; Thoelke, W.; Agape, C.; Borlini, G.; Ferri, I.; Massara, F.; et al. PocketPlant3D: Analysing canopy structure using a smartphone. Biosyst. Eng. 2017, 164, 1–12. [Google Scholar] [CrossRef]
- Paleari, L.; Movedi, E.; Vesely, F.M.; Confalonieri, R. Tailoring parameter distributions to specific germplasm: Impact on crop model-based ideotyping. Sci. Rep. 2019, 9, 1–9. [Google Scholar]
- Paleari, L.; Vesely, F.; Ravasi, R.; Movedi, E.; Tartarini, S.; Invernizzi, M.; Confalonieri, R. Analysis of the Similarity between in Silico Ideotypes and Phenotypic Profiles to Support Cultivar Recommendation—A Case Study on Phaseolus vulgaris L. Agronomy 2020, 10, 1733. [Google Scholar] [CrossRef]
- Tang, L.; Shi, C.-L.; Zhu, Y.; Jing, Q.; Cao, W.-X. A Quantitative Analysis on Leaf Curvature Characteristics in Rice. Crop. Modeling Decis. Support 2009, 32, 71–76. (In Chinese) [Google Scholar]
- Welham, S.J.; Gezan, S.A.; Clark, S.J.; Mead, A. Statistical Methods in Biology: Design and Analysis of Experiments and Regression; CRC Press Taylor & Francis Group: Boca Raton, FL, USA, 2015. [Google Scholar]
- Beuzelin, J.M.; Wilson, L.T.; Showler, A.T.; Meszaros, A.; Wilson, B.E.; Way, M.O.; Reagan, T.E. Oviposition and larval development of a stem borer, eoreuma loftini, on rice and non-crop grass hosts. Entomol. Exp. Appl. 2013, 146, 332–346. [Google Scholar] [CrossRef]
- Yang, H.; Luo, W.; He, H.; Xie, X. Rice leaf blade 3d morphology modeling and computer simulation. J. Agric. Mech. Res. 2008, 12, 33–36. (In Chinese) [Google Scholar]
- Farin, G. Curves and Surfaces for Computer Aided Geometric Design—A Practical Guide, 2nd ed.; Academic Press Inc.: London, UK, 1990; p. 444. [Google Scholar]
- Foley, J.D.; Feiner, S.K.; Hughes, J.F.; Dam, A.V. Computer Graphics: Principles and Practice, 2nd ed.; Addison-Wesley: Boston, MA, USA, 1990. [Google Scholar]
- Campbell, G.S. Derivation of an angle density function for canopies with ellipsoidal leaf angle distributions. Agric. For. Meteorol. 1990, 49, 173–176. [Google Scholar] [CrossRef]
- Campbell, G.S. Extinction coefficients for radiation in plant canopies calculated using an ellipsoidal inclination angle distribution. Agric. For. Meteorol. 1986, 36, 317–321. [Google Scholar] [CrossRef]
- Moriasi, D.N.; Arnold, J.G.; Van Liew, M.W.; Bingner, R.L.; Harmel, R.D.; Veith, T.L. Model evaluation guidelines for systematic quantification of accuracy in watershed simulations. Trans. ASABE 2007, 50, 885–900. [Google Scholar] [CrossRef]
- SAS. The Glm Procedure 9.3 User’s Guide; SAS Institute Inc.: Cary, NC, USA, 2015. [Google Scholar]
- Ma, P.L.; Ding, W.L.; Gu, H. The visual modeling of rice leaf based on opengl and bezier curved surface. J. Zhejiang Univ. Technol. 2010, 38, 36–40. (In Chinese) [Google Scholar]
- Liu, H.W.; Wu, B.; Zhang, H.Y.; Li, F.; Shao, Y.H. Research on rice leaf geometric model and its visualization. Comput. Eng. 2009, 35, 263–265. (In Chinese) [Google Scholar]
- Liu, H.W.; Liang, Y.Y.; Zhang, H. Research on the rice leaf morphological formation and its visualization. In Advanced Manufacturing Systems; Yang, D.G., Gu, T.L., Zhou, H.Y., Zeng, J.M., Jiang, Z.Y., Eds.; Trans Tech Publications Ltd.: Bäch SZ, Switzerland, 2011; Volume 201–203, pp. 2504–2508. [Google Scholar]
- Dornbusch, T.; Wernecke, P.; Diepenbrock, W. Description and visualization of graminaceous plants with an organ-based 3D architectural model, exemplified for spring barley (Hordeum vulgare L.). Vis. Comput. 2007, 23, 569–581. [Google Scholar] [CrossRef]
- Pieruschka, R.; Poorter, H. Phenotyping plants: Genes, phenes and machines introduction. Funct. Plant Biol. 2012, 39, 813–820. [Google Scholar] [CrossRef]
- Hoshikawa, K. The Growing Rice Plant: An Anatomical Monograph, 1st ed.; Nobunkyo Press: Tokyo, Japan, 1989; p. 310. [Google Scholar]
- Zhou, H.; Xia, D.; Zeng, J.; Jiang, G.; He, Y. Dissecting combining ability effect in a rice NCII-III population provides insights into heterosis in indica-japonica cross. Rice 2017, 10, 39. [Google Scholar] [CrossRef] [Green Version]
- Perez, R.P.; Fournier, C.; Cabrera-Bosquet, L.; Artzet, S.; Pradal, C.; Brichet, N.; Chen, T.; Chapuis, R.; Welcker, C.; Tardieu, F. Changes in the vertical distribution of leaf area enhanced light interception efficiency in maize over generations of selection. Plant Cell Environ. 2019, 42, 2105–2119. [Google Scholar] [CrossRef]
- Dingkuhn, M.; Penning De Vries, F.; Datta, S.K.D.; Van Laar, H. Concepts for a new plant type for direct seeded flooded tropical rice. In Direct Seeded Flooded Rice in the Tropics; IRRI: Manila, Philippines, 1991; pp. 17–38. [Google Scholar]
- Sinclair, T.R.; Sheehy, J.E. Erect Leaves and Photosynthesis in Rice. Science 1999, 283, 1455. [Google Scholar] [CrossRef]
- Murchie, E.; Chen, Y.-Z.; Hubbart, S.; Peng, S.; Horton, P. Interactions between Senescence and Leaf Orientation Determine in Situ Patterns of Photosynthesis and Photoinhibition in Field-Grown Rice. Plant Physiol. 1999, 119, 553–564. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zheng, B.; Shi, L.; Ma, Y.; Deng, Q.; Li, B.; Guo, Y. Comparison of architecture among different cultivars of hybrid rice using a spatial light model based on 3-D digitising. Funct. Plant Biol. 2008, 35, 900–910. [Google Scholar] [CrossRef]
- Hopkins, R.; Schmitt, J.; Stinchcombe, J.R. A latitudinal cline and response to vernalization in leaf angle and morphology inArabidopsis thaliana(Brassicaceae). New Phytol. 2008, 179, 155–164. [Google Scholar] [CrossRef] [PubMed]
- Uto, K.; Dalla Mura, M.; Sasaki, Y.; Shinoda, K. Estimation of Leaf Angle Distribution Based on Statistical Properties of Leaf Shading Distribution. In Proceedings of the IEEE International Geoscience and Remote Sensing Symposium (IGARSS), Waikoloa, HI, USA, 26 September–2 October 2020; pp. 5195–5198. [Google Scholar]
- Zhu, B.; Liu, F.; Xie, Z.; Guo, Y.; Li, B.; Ma, Y. Quantification of light interception within image-based 3-D reconstruction of sole and intercropped canopies over the entire growth season. Ann. Bot. 2020, 126, 701–712. [Google Scholar] [CrossRef] [Green Version]
- Burgess, A.; Retkute, R.; Herman, T.; Murchie, E.H. Exploring Relationships between Canopy Architecture, Light Distribution, and Photosynthesis in Contrasting Rice Genotypes Using 3D Canopy Reconstruction. Front. Plant Sci. 2017, 8, 734. [Google Scholar] [CrossRef] [PubMed]
- Sakamoto, T.; Morinaka, Y.; Ohnishi, T.; Sunohara, H.; Fujioka, S.; Ueguchi-Tanaka, M.; Mizutani, M.; Sakata, K.; Takatsuto, S.; Yoshida, S.; et al. Erect leaves caused by brassinosteroid deficiency increase biomass production and grain yield in rice. Nat. Biotechnol. 2005, 24, 105–109. [Google Scholar] [CrossRef]
- Mueller-Linow, M.; Pinto-Espinosa, F.; Scharr, H.; Rascher, U. The leaf angle distribution of natural plant populations: Assessing the canopy with a novel software tool. Plant Methods 2015, 11, 1–6. [Google Scholar] [CrossRef] [Green Version]
- Kuo, K.T.; Itakura, K.; Hosoi, F. Leaf segmentation based on k-means algorithm to obtain leaf angle distribution using terrestrial lidar. Remote Sens. 2019, 11, 2536. [Google Scholar] [CrossRef] [Green Version]
- Watson, D.J. Comparative Physiological Studies on the Growth of Field Crops: I. Variation in Net Assimilation Rate and Leaf Area between Species and Varieties, and within and between Years. Ann. Bot. 1947, 11, 41–76. [Google Scholar] [CrossRef]
- Lang, A.; Yueqin, X.; Norman, J. Crop structure and the penetration of direct sunlight. Agric. For. Meteorol. 1985, 35, 83–101. [Google Scholar] [CrossRef]
- Zou, X.; Mõttus, M. Retrieving crop leaf tilt angle from imaging spectroscopy data. Agric. For. Meteorol. 2015, 205, 73–82. [Google Scholar] [CrossRef]
- Jonckheere, I.; Fleck, S.; Nackaerts, K.; Muys, B.; Coppin, P.; Weiss, M.; Baret, F. Review of methods for in situ leaf area index determination: Part I. Theories, sensors and hemispherical photography. Agric. For. Meteorol. 2004, 121, 19–35. [Google Scholar] [CrossRef]
- Yan, G.; Hu, R.; Luo, J.; Weiss, M.; Jiang, H.; Mu, X.; Xie, D.; Zhang, W. Review of indirect optical measurements of leaf area index: Recent advances, challenges, and perspectives. Agric. For. Meteorol. 2019, 265, 390–411. [Google Scholar] [CrossRef]
- Confalonieri, R.; Foi, M.; Casa, R.; Aquaro, S.; Tona, E.; Peterle, M.; Boldini, A.; De Carli, G.; Ferrari, A.; Finotto, G.; et al. Development of an app for estimating leaf area index using a smartphone. Trueness and precision determination and comparison with other indirect methods. Comput. Electron. Agric. 2013, 96, 67–74. [Google Scholar] [CrossRef]
- Hirooka, Y.; Homma, K.; Shiraiwa, T. Parameterization of the vertical distribution of leaf area index (LAI) in rice (Oryza sativa L.) using a plant canopy analyzer. Sci. Rep. 2018, 8, 1–9. [Google Scholar] [CrossRef] [Green Version]
- Antunes, M.A.; Walter-Shea, E.A.; Mesarch, M.A. Test of an extended mathematical approach to calculate maize leaf area index and leaf angle distribution. Agric. For. Meteorol. 2001, 108, 45–53. [Google Scholar] [CrossRef]
- Fukuda, S.; Koba, K.; Okamura, M.; Watanabe, Y.; Hosoi, J.; Nakagomi, K.; Maeda, H.; Kondo, M.; Sugiura, D. Novel technique for non-destructive LAI estimation by continuous measurement of NIR and PAR in rice canopy. Field Crop. Res. 2021, 263, 108070. [Google Scholar] [CrossRef]











{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Thermogenic Male Sterile | Cytoplasmic Male Sterile | |||
|---|---|---|---|---|
| Restorer | 212S | 279S | 119A | 339A |
| 170R | 212S × 170R | 279S × 170R | 119A × 170R | 339A × 170R |
| 173R | 212S × 173R | 279S × 173R | 119A × 173R | 339A × 173R |
| Maintainers | - | - | 119B | 339B |
| Year | Plant 3D Traits | Variance Explained (%) | |||||
|---|---|---|---|---|---|---|---|
| Planting Date | Genotype (PD) 1 | Density (PD) 1 | Leaf Order (PD G D) 1 | Leaf Age (PD G D LO) 1 | Total | ||
| 2018 | Insertion angle | 8.1 ** | 7.1 ** | 0.2 ** | 25.9 ** | 22.4 ** | 63.7 |
| Elevation angle | 13.5 | 3.8 ** | 0.1 * | 28.6 ** | 8.9 ** | 54.9 | |
| Curve height | 11.6 ** | 5.6 ** | 0.3 ** | 31.4 | 8.0 * | 56.8 | |
| 2019 | Insertion angle | 0.1 ** | 5.2 ** | 0.1 | 8.8 ** | 18.4 ** | 32.6 |
| Elevation angle | 1.7 ** | 1.9 ** | 0.2 ** | 12.9 ** | 7.2 ** | 23.8 | |
| Curve height | 2.0 ** | 2.6 ** | 0.4 ** | 15.5 ** | 6.3 ** | 26.9 | |
| Plant 3D Traits | Variance Explained (%) | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|
| Planting Date | Genotype (PD) 1 | Density (PD) 1 | Leaf Order (PD G D) 1 | Leaf Age 2 | Leaf Length 2 | Leaf Width 2 | Leaf Area 2 | Specific Leaf Weight 2 | Total | |
| Insertion angle | 0.1 | 9.9 ** | 0.2 | 25.3 ** | 36.9 ** | 11.1 ** | 5.3 | 4.8 | 2.6 | 96.2 |
| Elevation angle | 2.2 ** | 4.8 ** | 0.0 | 25.8 ** | 22.7 * | 14.0 ** | 11.1 ** | 9.4 ** | 5.4 ** | 95.4 |
| Curve height | 1.6 ** | 7.4 ** | 0.0 | 29.9 ** | 16.8 | 13.6 * | 12.7 ** | 6.6 | 6.1 * | 94.7 |
| Plant 3D Traits | Year | Variance Explained (%) | |||||
|---|---|---|---|---|---|---|---|
| Planting Date | Genotype (PD) 1 | Density (PD) 1 | Octant (PD G D) 1 | Leaf Order (PD G D LO) 1 | Total | ||
| Azimuth angle | 2018 | 0.2 | 3.9 ** | 0.1 | 41.6 ** | - | 45.8 |
| 2019 | 0.4 | 2.48 ** | 0.1 | 30.7 ** | - | 33.7 | |
| Phyllotaxis | 2018 | 0.1 | 1.9 | 0.1 | - | 4.2 | 6.2 |
| 2019 | 0.0 | 0.3 | 0.0 | - | 1.0 | 1.3 | |
| Plant 3D Traits | Year | Variance Explained (%) | |||
|---|---|---|---|---|---|
| Planting Date | Genotype (PD) 1 | Density (PD) 1 | Total | ||
| Leaf angle distribution | 2018 | 24.0 ** | 4.4 ** | 0.3 * | 28.7 |
| 2019 | 0.5 ** | 14.4 ** | 0.9 ** | 15.7 | |
| Light extinction coefficient | 2018 | 27.5 ** | 6.7 ** | 0.5 ** | 34.6 |
| 2019 | 0.6 ** | 15.3 ** | 0.8 ** | 16.7 | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yang, Y.; Paleari, L.; Wilson, L.T.; Confalonieri, R.; Astaldi, A.Z.; Buratti, M.; Yan, Z.; Christensen, E.; Wang, J.; Samonte, S.O.P.B. Characterizing Genotype-Specific Rice Architectural Traits Using Smart Mobile App and Data Modeling. Agronomy 2021, 11, 2428. https://doi.org/10.3390/agronomy11122428
Yang Y, Paleari L, Wilson LT, Confalonieri R, Astaldi AZ, Buratti M, Yan Z, Christensen E, Wang J, Samonte SOPB. Characterizing Genotype-Specific Rice Architectural Traits Using Smart Mobile App and Data Modeling. Agronomy. 2021; 11(12):2428. https://doi.org/10.3390/agronomy11122428
Chicago/Turabian StyleYang, Yubin, Livia Paleari, Lloyd T. Wilson, Roberto Confalonieri, Adriano Z. Astaldi, Mirko Buratti, Zongbu Yan, Eric Christensen, Jing Wang, and Stanley Omar P. B. Samonte. 2021. "Characterizing Genotype-Specific Rice Architectural Traits Using Smart Mobile App and Data Modeling" Agronomy 11, no. 12: 2428. https://doi.org/10.3390/agronomy11122428
APA StyleYang, Y., Paleari, L., Wilson, L. T., Confalonieri, R., Astaldi, A. Z., Buratti, M., Yan, Z., Christensen, E., Wang, J., & Samonte, S. O. P. B. (2021). Characterizing Genotype-Specific Rice Architectural Traits Using Smart Mobile App and Data Modeling. Agronomy, 11(12), 2428. https://doi.org/10.3390/agronomy11122428