Abstract
Conservation management practices can improve soil health while minimizing deleterious effects of agriculture on the environment. However, adoption of these practices, particularly cover crops, is not widespread, as they often reduce crop yields compared to traditional management practices. The purpose of the current study was to determine if a two-species cover crop treatment of rye (Secale cereale L.) and crimson clover (Trifolium incarnatum L.) could increase soil health parameters and maximize soybean (Glycine max L.) yield greater than rye only in tilled and no-till Mississippi field soils. Enhanced microbial biomass and organic matter input from cover crops increased the activities of β-glucosidase, cellobiohydrolase, fluorescein diacetate hydrolysis, N-acetylglucosaminidase, and phosphatase in surface soils. Rye plus clover tended to elicit higher activities than rye only in no-till plots. Both cover crop treatments inhibited soybean yield in tilled plots by 11–25%. These results indicate that tillage exacerbates yield inhibition by cover crops in soybean and that double-species cover crop treatments were more consistent in increasing activities linked to nutrient cycling. Further study examining different combinations of cover crops in no-till systems is necessary to gain a better understanding of how they can be implemented to enhance soil health while maximizing crop yield.
1. Introduction
There has been increasing interest in the use of winter cover crops in row crop production to improve soil health, as well as the surrounding environment. As the name implies, these winter crops serve to cover the soil, decreasing the loss of soil water to evaporation [], while also shielding soil from rain impacts, thereby reducing erosion, increasing water infiltration, and ultimately improving the soil’s capacity to absorb water and maintain moisture []. Cover crops also function in nutrient preservation by absorbing nutrients from the soil, thus preventing their loss to leaching and erosion over the winter, and then releasing them back into the soil as their biomass degrades over the course of the summer growing season []. Some cover crop species also have inhibitory effects on weed populations []. Despite these benefits, there are several challenges that limit cover crop adoption in the southern US, including the increased costs they incur, such as the price of the seeds as well as the herbicides used for cover crop termination []. Cover crops often do not increase yield enough to offset these expenses, and in many cases, result in decreased cash crop yields [,], adding to the reluctance of their adoption.
Some of the issues resulting in cash crop yield inhibition by cover crops can be overcome by changes in management. Using chemical instead of mechanical termination and terminating at anthesis instead of earlier developmental stages have been found to decrease, but not eliminate, inhibitory effects of rye on the density, dry mass, and time to pod maturity in soybean []. Likewise, nitrogen (N) uptake and yield in corn can be increased by delaying hairy vetch cover crop termination in no-till systems []. Conversely, if not terminated early enough, cover crops can also serve as a reservoir for disease causing organisms in summer crops []. Differences in cover crop termination time can also alter weed density [], which in turn can influence yield if weed populations are large enough to outcompete the cash crop. The type of cover crops chosen can also impact their function in the field based on differences in chemical composition of plant tissues. Legumes generally have lower carbon (C):N and lignin:N ratios than cereal crops, resulting in a more rapid decomposition of legume biomass [,]. Such differences in decomposition may also influence cover crop efficacy if nutrients, such as N, are immobilized in cover crop biomass at early stages of cash crop development [,,]. Many cover crop species also contain allelopathic compounds that can elicit inhibitory effects in the main crop, with some exhibiting greater inhibition than others [,]. As such, careful selection of cover crop species may mitigate yield inhibition of the summer cash crop.
The soil microbial community plays a pivotal role in decomposition of cover crop biomass, and in turn, the rate of organic matter release back into the soil. Cover crops are known to cause shifts in microbial community composition, which can influence soil enzymes involved in nutrient cycling []. As such, assays of soil enzymes and microbial biomass can be an indicator for soil health and function in studies on the effects of cover crop treatment. Cover crop effects on soil enzymes can vary between species. For example, a previous study in the Mississippi Delta found balansa clover increased fluorescein diacetate (FDA) hydrolysis over twice as much as rye in a cotton cropping system, although this study did not report the effect of these treatments on yield []. Reddy [] assessed a wider range of cover crop species, including hairy vetch, crimson clover, rye, oat, wheat, subterranean clover, and Italian ryegrass, and found all species inhibited soybean yield relative to no cover controls, but did not include an assessment of soil health. Another more recent study conducted in the Mississippi Delta found that rye cover crop plots tended to display slightly higher microbial biomass and enzyme activities than plots with crimson clover as a winter cover, while having a greater inhibitory effect on soybean yield []. In contrast, the yield inhibition was less severe in tilled clover plots, and no inhibition was seen in response to clover in no-till plots []. The exact nature of this inhibition was unclear but appears to be due, in part, to allelopathic compounds in rye. As N fixation has been found to only supply 50–60% of soybean N demands [], slower degradation of rye biomass tying up nutrients and delaying the release of N back into the soil may have also contributed to the lower yields observed in rye plots. Given that rye grown in the presence of legumes has been found to have a lower C:N ratio compared to rye grown in a monoculture [], a rye-legume cover crop mix may provide more consistent soil health benefits than a rye only winter cover, since lower C:N ratios lead to faster decomposition. Much of the research on cover crop mixes has been conducted in geographic regions which have different agronomic conditions and needs compared to the Mississippi Delta, where the emphasis of past cover crop research has been on single-species treatments. As such, the purpose of the current study was to determine the effects of a double-species cover crop mix of rye and crimson clover compared to rye only on both soil health and soybean yield under till and no-till management in Mississippi Delta field soils. Given the status of soybean as a major crop in many countries across the world [], finding ways to reliably enhance soybean yield while preserving soil resources will also contribute toward minimizing environmental impacts of soybean production.
2. Materials and Methods
2.1. Experimental Design, Site History, and Management
The study site was located on the experimental research farm in Stoneville, MS, in the mid-south region of the USA (33.44330° latitude, −90.88303° longitude). The study field was arranged in a randomized block design with four blocks under till (T) and four blocks under no-till (NT) management. Each block was split into three plots (32 m × 8.4 m each) for winter cover crop treatments, resulting in a total of 24 plots (four replicate plots of each tillage–cover crop combination). Soil texture within the field site was predominantly Commerse series (fine-silty, mixed, superactive, nonacid, thermic Fluvaquentic Endoaquepts) with some Dowling series (very-fine, smectitic, nonacid, thermic Vertic Endoaquepts) in one end of the field. Replicate plots were arranged such that these soil textures were equally represented under each treatment. Field plots were initiated in the fall of 2000 for a study assessing the effects of Abruzzi rye (Secale cereale), Balansa clover (Trifolium michelianum), and no winter cover crop in a cotton (Gossypium hirsutum L.) production system []. No-till plots have not been plowed since initiation of the original study, while tilled plots have been plowed each fall. The field site remained under cotton production until the conclusion of the initial study in fall 2006, after which it was maintained under continuous soybean (Glycine max L.) until fall 2014, when a new study was established comparing effects of Elbon rye and crimson clover winter cover crop treatments in a soybean production system []. The current study was initiated in fall 2017, with the planting of new cover crop treatments. Treatment combinations for the current study were: till rye (TR), till rye+clover (TRC), till no cover (TNC), no-till rye (NTR), no-till rye+clover (NTRC), and no-till no cover (NTNC). Tilled plots were plowed following soybean harvest each fall. Cover crops were planted during October of each year, allowed to grow over the winter, and terminated with two applications of paraquat dichloride at a rate of 0.77 kg ha−1 (Gramoxone SL2.0, Syngenta, Greensboro, NC, USA), with each application being two weeks apart each April. Soybean (Asgrow AG4632) was planted in May and harvested in September of each year. Monthly precipitation and average daily air temperatures were obtained from the Mississippi State University Delta Agricultural Weather Center (http://deltaweather.extension.msstate.edu/coop-stoneville; accessed on 8 June 2021).
2.2. Sample Collection
Soil samples were collected each spring in April prior to cover crop termination, at mid-summer (late June 2018 and early July 2019), and in the fall during the window of time between soybean harvest and cover crop planting in October. For each timepoint, eight subsamples were collected from the top 0–5 cm soil at randomly selected locations within each plot using a 1.8 cm diameter soil probe. These subsamples were pooled to form one composite sample per plot. An additional 5–15 cm depth was collected at the spring timepoints in order to encompass the plow zone, which is the region of the soil profile disrupted by tilling. Soil samples were placed in plastic bags after collection, transported to the laboratory on ice, and stored at 5 °C until analysis. All soils were passed through a 2 mm sieve prior to analysis. Two plant biomass samples of winter growth were collected from each plot using a one square meter quadrat. Plant biomass samples were stored in paper bags and dried in a greenhouse for two months. Winter plant biomass was recorded as the average dry weight of the two quadrats collected from each plot.
2.3. Soil Sample Analyses
Soil moisture (SM) content was determined by weighing out 5 g fresh weight aliquots from each sample in triplicate, oven drying overnight at 105 °C, and calculated as the percentage moisture lost from drying. Soil organic matter (SOM) was determined by ashing oven dried soils (500 °C for two hours) and calculated as percentage of weight lost during ashing. Analysis of microbial biomass in soil samples was conducted using a chloroform-fumigation and extraction-based protocol. Briefly, two 12 g aliquots were weighed out from each soil sample. The first aliquot was extracted by shaking at 200 rpm for one hour in 50 mL of 0.5 M K2SO4 buffer. The second aliquot was chloroform fumigated for five days as described in Horwath and Paul [] prior to extraction. Extracts were centrifuged at 3000× g for 15 min and then gravity filtered through Whatman GF/F filter paper (GE Healthcare, Pittsburgh, PA, USA). Extracts were diluted 10-fold to decrease salt concentration for analysis on a Shimadzu total organic carbon (TOC)-L analyzer with total nitrogen module (TNM-L) to determine TOC and total nitrogen (TN) concentration. Concentrations of TOC and TN in fumigated and unfumigated soils were determined by multiplying diluted extract concentrations by ten and dividing by the dry weight equivalent of the soil from each extraction. Microbial biomass carbon (MBC) and microbial biomass nitrogen (MBN) were calculated using the equations:
where EC and EN are the difference in TOC and TN between fumigated and unfumigated soil aliquots, and kEC and kEN are constants of 0.35 and 0.68, respectively []. Activities of β-glucosidase, cellobiohydrolase, N-acetylglucosaminidase, and phosphatase in soil samples were determined using p-nitrophenol (pNP) linked assays in 96-well format as described in Jackson et al. []. Fluorescein diacetate (FDA) hydrolysis was assayed using a protocol from Schnürer and Rosswall [] modified for a 96-well plate format as described in Tyler [].
MBC = EC/kEC
MBN = EC/kEN
2.4. Statistics
All statistical analyses were conducted in JMP version 15.0.0. (SAS Institute Inc., Carey, NC, USA). Differences in winter plant biomass and soybean yield between tillage and cover crop treatments for each year were determined by two-way analysis of variance (ANOVA). Treatment effects on pH, SM, SOM, MBC, MBN, and enzyme activities over the entire study period were tested for by two-way repeated-measures ANOVA. Tillage treatment, cover crop treatment, and year were considered fixed, independent variables. Winter plant biomass, soybean yield, pH, SM, SOM, MBC, MBN, and soil enzyme activities were considered dependent variables. There were four replicates for each tillage–cover crop treatment combination. Differences between treatment combinations at specific timepoints were determined by individual two-way ANOVAs and Tukey’s Honestly Significant difference test using an alpha of 0.05. Pearson correlations were conducted to assess relationships between soil enzymes, microbial biomass, pH, SM, and SOM, and between cover crop biomass and soybean yield. Pearson correlations were also performed on annual cover crop biomass inputs for 2018 and 2019 with the soil enzyme activities for each timepoint in 2018 and 2019, respectively.
3. Results and Discussion
3.1. Cover Crop Biomass
Two-way repeated-measures ANOVA indicated no significant interactions between tillage and cover crop treatments. Rye+clover (RC) and rye (R) produced 300–600% more winter plant biomass than NC plots that were naturally colonized by native vegetation (Figure 1). Between the cover crop treatments, RC produced twice as much cover crop biomass compared to rye (R) in 2018 under both tilled (T) and no-till (NT) plots (p < 0.007; Figure 1). In 2019, RC biomass was also greater than R biomass, but to a lesser extent and only significant in T plots (p = 0.0388; Figure 1). There was a general increase in winter biomass from 2018 to 2019, with the greatest difference between years observed in R plots. Lower levels in 2018 could be due to colder temperatures observed in December 2017 and January 2018, compared to December 2018 and January 2019 (Figure 2), leading to slower cover crop growth. Differences in precipitation may also have contributed. November 2018 to May 2019 tended to have higher monthly precipitation than the previous winter season, with the exception of February and March of 2018, which received 25–50% higher precipitation than comparable months in 2019 (Figure 2). While R is generally considered to be a hardy cover crop, its growth may have benefitted from the warmer temperatures and higher rainfall in the winter leading up to the 2019 growing season, resulting in greater biomass production.
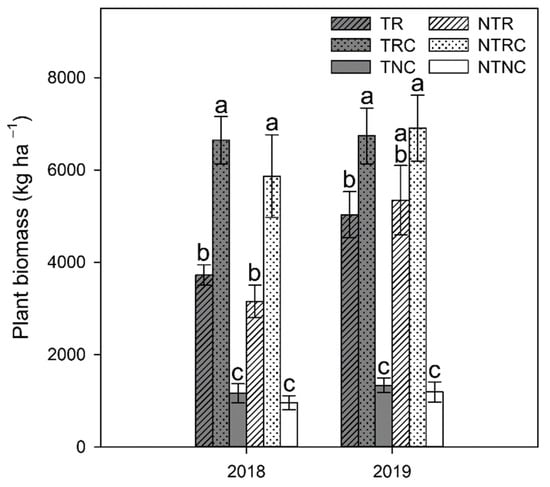
Figure 1.
Plant biomass from winter growth in tilled (T) and no-till (NT) plots planted with rye only (R), rye plus crimson clover (RC), or no winter cover crop (NC) in April 2018 and 2019. Values represent the mean ± standard error. Statistical significance between treatments for each timepoint are denoted by different letters.
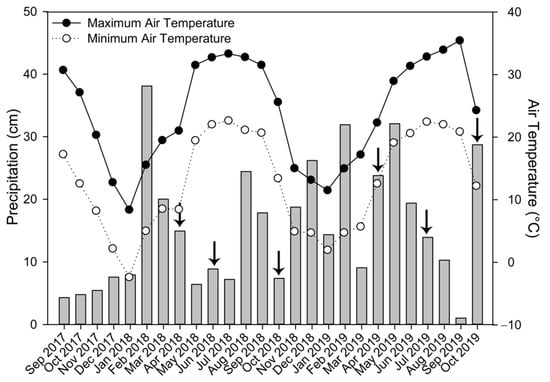
Figure 2.
Monthly precipitation (bars) and average minimum (open circle) and maximum (closed circle) air temperature during the study period. Arrows indicate soil sample collection timepoints.
In comparison, RC had more consistent biomass production between years, highlighting an advantage of double- over single-species cover crop mixes, as lower growth of one species may be compensated for by the second, resulting in greater reliability in cover crop biomass production across growing seasons than if a single species were planted. Synergistic interactions between cover crops, such as transfer of N from legume to grass species [] or differing structure of rye and clover resulting in an altered canopy that allowed for greater light interception [], may have also allowed both species to grow better under the colder, drier conditions in the 2018 winter season.
3.2. Soil Chemical Characteristics
Over the course of the two-year study period, SOM levels ranged from approximately 4.2% in TNC to 6.8% in NTRC treatments in the 0–5 cm depth (Figure 3C), and 4.7% in TNC to 5.2% in NTRC in 5–15 cm soil (Table 1). Two-way ANOVA of each timepoint indicated no significant differences between individual tillage and cover crop combinations (Figure 3C; Table 1). However, two-way repeated-measures ANOVA showed significant interactions between time and cover crop (p = 0.0372) and time and tillage (p = 0.019) for SOM in 0–5 cm soil. Average SOM for both cover crop treatments generally tended to be higher than NC, while NT tended to have higher SOM than T (Figure 3C). This is likely driven by cover crop biomass decomposition, which serves to return organic matter to the soil. Reddy et al. [] also found SOM to be higher in crimson clover and rye than no cover in soybean fields, and Moore et al. [] found rye cover crop increased SOM relative to no cover in a corn-soy rotation. Locke et al. [] found that cover crop and tillage effects on total C concentration were most pronounced in 0–2 cm soils, indicating that nutrient inputs from cover crop biomass degradation are concentrated in surface soils. This stratification of C and organic matter, with decreasing concentrations and treatment-based effects by depth, was also observed by Blanco-Canqui and Lal [], and is consistent with the results of the current study, with SOM not differing significantly between treatments in the 5–15 cm depth (Table 1).
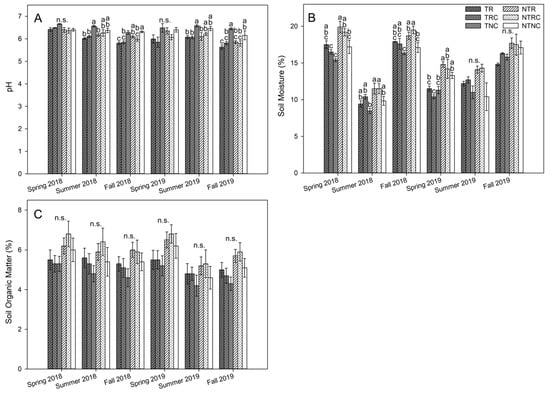
Figure 3.
Soil pH (A), soil moisture content (B), and soil organic matter (C) in 0–5 cm depth from soybean study plots during 2018 to 2019 growing seasons. Values represent the mean ± standard error (n = 4). Statistical significance between treatments for each timepoint are denoted by different letters (α = 0.05). Timepoints marked with n.s. indicate no significant differences between pairwise comparisons.

Table 1.
Soil chemical traits in 5–15 cm soil in spring 2018 and 2019 *.
Effects on soil moisture varied across time in 0–5 cm soil, with more differences in 2018 than 2019 (Figure 3B). Both NT (p ≤ 0.0127) and cover cropped plots (p ≤ 0.0415) had higher SM in all 2018 timepoints. In contrast, cover crops only increased soil moisture relative to no cover treatments in summer 2019 (p ≤ 0.041), and NT plots only had higher soil moisture than T plots in the spring (p < 0.0001) and fall (p = 0.00096) of that year. This variability could be due to differences in weather, as 2018 generally received less precipitation and the amount of rain received before sampling was lower in 2018 versus comparable 2019 timepoints (Figure 2). With the lower precipitation in 2018, retention of SM due to cover crop residues shielding the soil surface was more evident. These results are consistent with those of Zablotowicz et al. [], where soil moisture content in 0–5 cm soil was consistently higher in NT than T soils while the effects of rye and hairy vetch cover crops were less consistent across timepoints. Meanwhile, SM in deeper 5–15 cm soil did not differ significantly between treatments in the spring timepoints (Table 1).
Soil pH was slightly acidic, ranging from approximately 5.6 to 6.7, and varied by timepoint, tending to be higher in the spring and summer than in the fall (Figure 3A). Two-way repeated-measures ANOVA indicated an interaction between tillage and cover, with pH in T plots being more responsive to the effects of cover crop than NT in 0–5 cm soil (p = 0.048). Cover crops tended to decrease soil pH in this depth, with pH in tilled NC being higher than R (p ≤ 0.015) and RC (p ≤ 0.014) for all but the spring timepoints. Effects of cover crop on pH in no-till plots was variable but tended to be greatest in fall 2018 and summer 2019 (Figure 3A). No significant differences in pH were observed in 5–15 cm soils (Table 1).
3.3. Microbial Biomass
Two-way repeated-measures ANOVA indicated microbial biomass was enhanced by NT (MBC, p < 0.0001; MBN, p < 0.0001) and both cover crops treatments (MBC, p < 0.0001; MBN, p = 0.0057) in 0–5 cm soil throughout the study period (Figure 4), while no significant treatment effects were observed in the 5–15 cm depth (Table 2). This observation is consistent with prior observations of rye and crimson clover as individual treatments increasing MBC and MBN []. Similarly, conservation management incorporating rye cover crops also increased microbial biomass relative to conventional management practices in monoculture wheat and wheat-bean-potato rotations []. Interestingly, while cover crop-based differences in the current study were apparent in NT plots throughout the growing season, differences between cover crop and no cover in T plots were not significant in the spring timepoints prior to cover crop termination. They became more pronounced later in the growing season of both years (Figure 4), possibly the result of the organic matter input from degrading cover crop residues stimulating microbial growth in soils. Consistent with this, correlations between microbial biomass levels and yearly cover crop biomass inputs peaked in the summer timepoints (Table 3) while they were not significant in spring timepoints prior to cover crop termination before the start of cover crop decomposition, or the fall timepoint after the bulk of cover crop residues had been degraded. In addition, both MBC and MBN correlate significantly with SOM (Table 4).
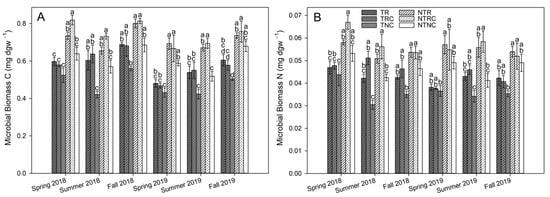
Figure 4.
Microbial biomass carbon (A) and microbial biomass nitrogen (B) in 0–5 cm soil from till (T) and no-till (NT) plots planted with winter cover crop treatments of rye only (R), rye+clover (RC), and no cover (NC) during the 2018 and 2019 soybean growing season. Values represent the mean ± standard error (n = 4). Statistical significance between treatments for each timepoint are denoted by different letters (α = 0.05).
Table 2.
Microbial biomass C and N in 5–15 cm soil in spring 2018 and 2019 *.
Table 3.
Correlations of with annual winter plant biomass input with enzyme activities and microbial biomass from each timepoint #.
Table 4.
Correlation matrix of chemical and biological traits in 0–5 cm soil *.
3.4. Soil Enzyme Activities
All enzymes were significantly impacted by cover crop treatments (p < 0.0001) and tillage (p ≤ 0.0358) according to two-way repeated-measures ANOVA. Generally, NT tended to have higher activities than T plots, while the response of soil enzymes to cover crop varied between tillage treatment (Figure 5). RC significantly enhanced enzyme activities compared to NC controls more often than R, and this trend was most notable in NT plots, where NTRC activities were significantly higher than NTNC for cellobiohydrolase at all timepoints (p ≤ 0.0242) as well as β-glucosidase (p ≤ 0.0366) and phosphatase (p ≤ 0.0410) in all but the fall 2019 timepoint (Figure 5A,B,E). For NAGase (Figure 5D), NTRC was only higher than NTNC at three timepoints (spring 2018 and spring and summer 2019; p ≤ 0.0359), and for FDA hydrolysis (Figure 5C), the relationship was only significant in summer 2018 (p ≤ 0.0057). Meanwhile, NTR plots only enhanced activities relative to NTNC for three enzymes across four timepoints: cellobiohydolase in fall 2018 (p = 0.0054), spring 2019 (p = 0.0426), and summer 2019 (p < 0.0001); phosphatase in fall 2018 (p = 0.025); and FDA hydrolysis in summer 2018 (p = 0.0098). In contrast to NT, T plots were more responsive to R cover crop treatment. Both TR and TRC enhanced multiple enzymes relative to TNC. However, TR enhanced β-glucosidase (p = 0.0419), FDA hydrolysis (p = 0.0104), and phosphatase (p = 0.0049) in fall 2018, when TRC did not (Figure 5A,C,E). In comparison, TRC only outperformed TR in enhancing β-glucosidase in summer 2018 (p = 0.0077) and phosphatase in spring and summer 2019 (p ≤ 0.0430). In a cotton system, Cordeiro et al. [] also observed differences in soil enzyme activities between mixed cover crop treatments, where three- and two-species cover crop mixes that included both grasses and legumes increased FDA hydrolysis and β-glucosidase relative to fallow and single-grass species cover treatments, with the greatest enhancement seen with the three-species mixture. These observations, coupled with the results from the current study, suggest that increasing the number and diversity of plant species in a cover crop mix will result in more consistent enhancement of soil activities. In contrast, Calderón et al. [] found that neither single-species nor a 10 species cover crop mix enhanced β-glucosidase or NAGase activities relative to fallow fields, with enzymes being more responsive to differences in irrigation. These results demonstrate how climatic differences can also impact soil activities, particularly in the semiarid region where the Calderón et al. [] study was conducted. However, the current study was conducted in the midsouth US and irrigated as needed, minimizing effects of drought on soil moisture and enzyme activities.
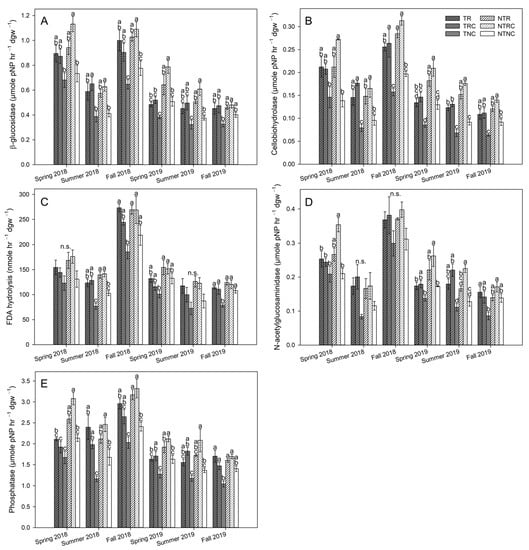
Figure 5.
Activities of β-glucosidase (A), cellobiohydrolase (B), FDA hydrolysis (C), N-acetylglucosaminidase (D), and phosphatase (E) in 0–5 cm soil from till (T) and no-till (NT) plots planted with winter cover crop treatments of rye only (R), rye+clover (RC), and no cover (NC) during the 2018 and 2019 soybean growing seasons. Values represent the mean ± standard error. Statistical significance between treatments for each timepoint are denoted by different letters (α = 0.05). Timepoints marked with n.s. indicated no significant differences between pairwise comparisons.
Effects of land management on soil enzyme activities in 5–15 cm soils were less noticeable (Figure 6). Two-way ANOVA indicated tillage enhanced the activities of β-glucosidase (p = 0.0002), cellobiohydrolase (p = 0.0178), and FDA hydrolysis (p = 0.0495) in 5–15 cm soil, but only in 2018 (Figure 6A–C). These results contrast with the effects observed in 0–5 cm soil, where NT plots had higher activities. Higher activities in tilled soil in the 5–15 cm depth are likely the result of above ground plant biomass being incorporated into the soil, thereby providing a substrate to soil microorganisms, stimulating their activity. No cover crop-based differences in β-glucosidase, NAGase, or FDA hydrolysis were observed in 5–15 cm soil. Cellobiohydrolase (p = 0.0081) and phosphatase (p = 0.0166) activities were higher in RC compared to NC plots at this depth in 2018 (Figure 6A,E), while the only significant effect observed in 2019 was that phosphatase activity was higher in TR than TNC (Figure 6E; p = 0.0456). As tillage was performed in the fall, cover crop biomass left on the soil surface had already been partially degraded over the course of the growing season. As such, it is not surprising that cover crop treatment effects in deeper soil were less pronounced. These results were consistent with Locke et al. [], where single-species cover crop treatments of Abruzzi rye and balansa clover enhanced FDA hydrolysis in surface soils but did not significantly impact activity in 5 15 cm in cotton fields. The enhancement of FDA activity observed by Locke et al. [] was attributed to higher biomass C providing binding sites and greater stability for soil enzymes as well as greater substrate availability for the soil microorganisms that produce these enzymes.
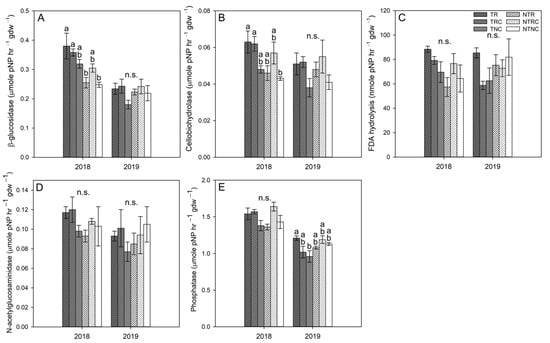
Figure 6.
Activities of β-glucosidase (A), cellobiohydrolase (B), FDA hydrolysis (C), N-acetylglucosaminidase (D), and phosphatase (E) in 5–15 cm soil from till (T) and no-till (NT) plots planted with winter cover crop treatments of rye only (R), rye+clover (RC), and no cover (NC) in spring 2018 and 2019. Values represent the mean ± standard error. Statistical significance between treatments for each timepoint are denoted by different letters (α = 0.05). Timepoints marked with n.s. indicated no significant differences between pairwise comparisons.
Organic matter input from crop residues is known to influence soil enzyme activity and function [,], and appears to be a driving factor for enhanced enzyme activities seen in the current study. Correlations were performed between annual biomass input and the soil enzyme activities for each timepoint to determine how enzymatic response may change over the course of the growing season as cover crop residues degrade (Table 3). These correlations peak in summer and decline slightly by the fall timepoint once a large portion of the above ground cover crop biomass has decomposed (Table 3). β-glucosidase and cellobiohydrolase activities from each timepoint significantly correlated with the cover crop biomass across all timepoints, with correlation coefficients ranging from 0.431 to 0.805, which is consistent with these two enzymes’ role in cellulose hydrolysis and organic matter degradation []. NAGase, involved in both C and N mineralization [], also correlated with cover crop biomass input in all but the fall 2018 timepoint, although to a lesser extent, with significant correlations ranging from 0.407 to 0.769. FDA hydrolysis, linked to general microbial hydrolytic activities [], also correlated with biomass inputs for all but the spring 2019 timepoint, with significant correlations ranging from 0.405 to 0.678. Meanwhile, phosphatase activities, linked to organic phosphate mineralization [], only correlated to yearly cover crop biomass inputs at four timepoints, mostly in 2019, ranging from 0.429 to 0.609.
All soil enzymes assayed also significantly correlated with soil moisture content, and phosphatase and FDA hydrolysis were weakly, negatively correlated with pH (Table 4). Consistent with the observed correlation to biomass inputs, enzymes involved in organic matter mineralization (β-glucosidase, r = 0.458; cellobiohydrolase, r = 0.368; NAGase, r = 0.296; phosphatase, r = 0.486) all correlated significantly with SOM (p < 0.0001). FDA hydrolysis also correlated with SOM, but to a lesser extent (r = 0.183; p = 0.028). Given that FDA hydrolysis is a measure of general microbial hydrolytic activity and includes the activities of multiple enzyme classes, including proteases, lipases, and esterases [,], the lower correlation to SOM is not surprising. Larger microbial communities in NT and cover cropped plots also appear to play a role in enzyme activity levels, as evidenced by high correlations of activities with MBC (r > 0.06; Table 4).
3.5. Effects on Soybean Yield
While conservation management is important for preserving soil health resources, ultimately, the primary goal for farmers is to optimize crop yields while minimizing management costs. As such, the impact of conservation management on yield must also be considered. Generally, soybean yields in cover crop treatments differed more in T than NT treatment plots (Figure 7). In 2018, yields in NT plots were approximately 10% higher than yields in T plots (p = 0.0002). Yields in cover cropped plots (CR and R) did not differ significantly within NT. However, both TR (p = 0.0207) and TRC (p = 0.0037) had 11–14% lower yields than TNC in 2018. Similarly, yields in TR and TRC were approximately 25% lower than TNC while there were no differences between cover cropped and NC treatments within NT plots in 2019 (Figure 7). This observation indicates that employing tillage when using cover crops exacerbates the inhibitory effect they can have on soybean yield. Water stress may be a contributing factor to this yield inhibition. Soybean grown in tilled soils has been found to be more susceptible to water stress, resulting in lower yields than soybean grown in no-till plots that had a greater soil moisture reserve to pull from []. While plots in the current study were irrigated, soybean plants could have experienced some water stress between irrigation and precipitation events. Such stress could have made the plants more susceptible to inhibition by allelopathic compounds released from decomposing cover crop residues, eliciting decreased yields in tilled cover crop soil. A prior study conducted at this site reported a higher incidence of bacteria associated with biocontrol of plant pathogens in tilled plots []. As R cover crops can act as a host to pathogens of summer crops [], lower yields in cover cropped plots in the current study may be the result of higher incidence of disease. Some evidence suggests that higher microbial activity in NT soils can result in disease suppressive soils that lessen effects of fungal pathogens [], offering another potential factor that may have contributed to the lack of significant yield inhibition seen in response to cover crops in NT plots.
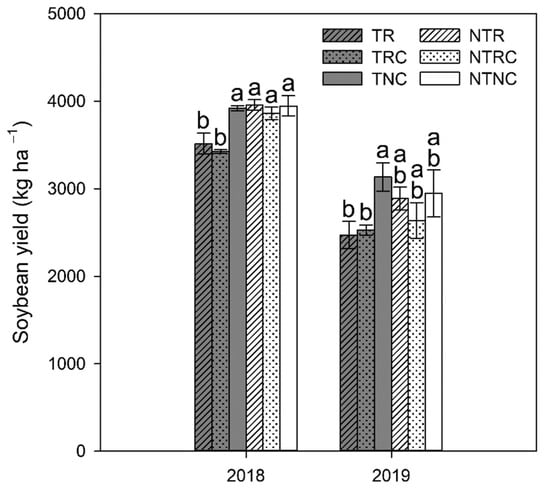
Figure 7.
Soybean yield in till (T) and no-till (NT) plots planted with winter cover crop treatments of rye only (R), rye+clover (RC), and no cover (NC) during the 2018 and 2019 soybean growing season. Values represent the mean ± standard error. Statistical significance between treatments for each timepoint is denoted by different letters (α = 0.05).
Soybean yield was negatively correlated with cover crop biomass (r = −0.44; p = 0.0018). When broken down by winter cover treatment, there was a significant negative correlation between rye biomass and soybean yield (r = −0.77; p = 0.0005), which is consistent with previous studies reporting inhibition of soybean yield by cereal rye as a winter cover [,]. Meanwhile, the correlations of biomass and yield within RC (r = −0.33; p = 0.21) were not statistically significant, suggesting that the presence of crimson clover may have counterbalanced some of rye’s inhibitory effects. Singh et al. [] also found that rye resulted in lower soybean yields, which was attributed to immobilizing soil N during early stages of soybean growth before root nodule establishment. Additionally, Reddy [] found that while crimson clover did inhibit soybean yield, it was to a lesser extent than the inhibition observed in cereal rye treatments. Zablotowicz et al. [] also found that rye inhibited yields while hairy vetch, a legume cover crop, did not. As yield inhibition was observed in response to both cover crop treatments in tilled soil to a similar extent, multiple factors could be contributing to yield loss.
In contrast to the above-mentioned trends of higher yield in clover versus rye cover crop treatments, Reddy et al. [] found that soybean yield was lower in crimson clover than in rye, which was attributed to the higher weed populations in crimson clover plots, highlighting the fact that differences on weed control between treatments can counter balance benefits to soil health. The current study employed weed control measures applied to all treatments, negating potential effects of weed populations on yield. Kessavalou and Walters [] found the inhibitory effects of rye cover on yield in a corn-soybean rotation system to be variable and attributed differences to timing of rye cover termination, suggesting that rye terminated at earlier developmental stages had greater allelopathic effects. In the current study, rye was terminated at the later developmental stage of anthesis in both years, negating developmental stage as a potential contributor to yield variability. However, co-cultivation with clover may have altered the chemical composition of rye biomass or root exudates, potentially impacting the concentration of allelopathic compounds or the speed of their release into the soil, as co-cultivation with a legume has been found to decrease its C:N ratio [], which in turn can increase the rate of decomposition.
4. Conclusions
Both single- and double-species cover crop treatments enhanced some aspects of soil health, with higher levels of microbial biomass and soil enzyme activities relative to no cover controls across most timepoints. These differences appear to be driven, in large part, by release of organic matter from degrading cover crop biomass residues over the course of the growing season. Rye with clover often had slightly higher activities than R only, particularly in NT plots. Despite these improvements, both single- and double-species cover crop treatments significantly inhibited soybean yield in T plots, indicating other soil traits respond differently to cover crop implementation. As such, further study is required to assess an expanded array of soil health parameters, including soil nutrient dynamics. Future research needs also include identification of ideal cover crop species, and possible optimization of cover crop planting and termination times to minimize allelopathy while timing degradation of biomass to coincide with cash crop nutrient needs. Yield inhibition was not observed in NT plots, suggesting this management approach will be ideal for future cover crop studies assessing yield. Meanwhile, farmers wishing to employ cover crops in Mississippi Delta field soils may get the best return of yield by using a NT management approach.
Funding
This research received no external funding.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
Not applicable.
Acknowledgments
The author would like to thank Paige Goodlett for assistance in collecting and processing soil samples. This research was supported by the U.S. Department of Agriculture, Agricultural Research Service. Mention of trade names or commercial products is solely for the purpose of providing specific information and does not imply recommendation or endorsement by the USDA. The findings and conclusions in this publication are those of the author and should not be construed to represent any official USDA or U.S. Government determination or policy. USDA is an equal opportunity provider and employer.
Conflicts of Interest
The author declares no conflict of interest.
References
- Locke, M.A.; Bryson, C.T. Herbicide-soil interactions in reduced tillage and plant residue management systems. Weed Sci. 1997, 45, 307–320. [Google Scholar] [CrossRef]
- Blanco-Canqui, H.; Shaver, T.M.; Lindquist, J.L.; Shapiro, C.A.; Elmore, R.W.; Francis, C.A.; Hergert, G.W. Cover Crops and Ecosystem Services: Insights from Studies in Temperate Soils. Agron. J. 2015, 107, 2449–2474. [Google Scholar] [CrossRef] [Green Version]
- Teasdale, J.R.; Brandsæter, L.O.; Calegari, A.; Neto, F.S. Cover crops and weed management. In Non-Chemical Weed Management: Principles, Concepts and Technology; CABI: Egham, UK, 2007; pp. 49–64. [Google Scholar]
- Bergtold, J.S.; Ramsey, S.; Maddy, L.; Williams, J.R. A review of economic considerations for cover crops as a conservation practice. Renew. Agric. Food Syst. 2019, 34, 62–76. [Google Scholar] [CrossRef]
- Mischler, R.A.; Curran, W.S.; Duiker, S.W.; Hyde, J.A. Use of a Rolled-rye Cover Crop for Weed Suppression in No-Till Soybeans. Weed Technol. 2010, 24, 253–261. [Google Scholar] [CrossRef]
- Reddy, K.N. Effects of Cereal and Legume Cover Crop Residues on Weeds, Yield, and Net Return in Soybean (Glycine max). Weed Technol. 2001, 15, 660–668. [Google Scholar] [CrossRef]
- Westgate, L.R.; Singer, J.W.; Kohler, K.A. Method and Timing of Rye Control Affects Soybean Development and Resource Utilization. Agron. J. 2005, 97, 806–816. [Google Scholar] [CrossRef]
- Sainju, U.M.; Singh, B.P. Tillage, Cover Crop, and Kill-Planting Date Effects on Corn Yield and Soil Nitrogen. Agron. J. 2001, 93, 878–886. [Google Scholar] [CrossRef]
- Acharya, J.; Bakker, M.G.; Moorman, T.B.; Kaspar, T.C.; Lenssen, A.W.; Robertson, A.E. Time Interval Between Cover Crop Termination and Planting Influences Corn Seedling Disease, Plant Growth, and Yield. Plant Dis. 2017, 101, 591–600. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mirsky, S.B.; Curran, W.S.; Mortenseny, D.M.; Ryany, M.R.; Shumway, D.L. Timing of Cover-Crop Management Effects on Weed Suppression in No-Till Planted Soybean using a Roller-Crimper. Weed Sci. 2011, 59, 380–389. [Google Scholar] [CrossRef]
- Cookson, W.; Beare, M.; Wilson, P. Effects of prior crop residue management on microbial properties and crop residue decomposition. Appl. Soil Ecol. 1998, 7, 179–188. [Google Scholar] [CrossRef]
- Jahanzad, E.; Barker, A.V.; Hashemi, M.; Eaton, T.; Sadeghpour, A.; Weis, S.A. Nitrogen Release Dynamics and Decomposition of Buried and Surface Cover Crop Residues. Agron. J. 2016, 108, 1735–1741. [Google Scholar] [CrossRef]
- Kessavalou, A.; Walters, D.T. Winter Rye as a Cover Crop Following Soybean under Conservation Tillage. Agron. J. 1997, 89, 68–74. [Google Scholar] [CrossRef]
- Kessavalou, A.; Walters, D.T. Winter Rye Cover Crop Following Soybean Under Conservation Tillage: Residual Soil Nitrate. Agron. J. 1999, 91, 643–649. [Google Scholar] [CrossRef]
- Shekoofa, A.; Safikhan, S.; Raper, T.B.; Butler, S.A. Allelopathic Impacts of Cover Crop Species and Termination Timing on Cotton Germination and Seedling Growth. Agronomy 2020, 10, 638. [Google Scholar] [CrossRef]
- Barnes, J.P.; Putnam, A.R. Rye residues contribute weed suppression in no-tillage cropping systems. J. Chem. Ecol. 1983, 9, 1045–1057. [Google Scholar] [CrossRef] [PubMed]
- Fernandez, A.L.; Sheaffer, C.C.; Wyse, D.L.; Staley, C.; Gould, T.J.; Sadowsky, M.J. Associations between soil bacterial community structure and nutrient cycling functions in long-term organic farm soils following cover crop and organic fertilizer amendment. Sci. Total Environ. 2016, 566–567, 949–959. [Google Scholar] [CrossRef] [Green Version]
- Locke, M.A.; Zablotowicz, R.M.; Steinriede, R.W.; Testa, S.; Reddy, K.N. Conservation Management in Cotton Production: Long-Term Soil Biological, Chemical, and Physical Changes. Soil Sci. Soc. Am. J. 2013, 77, 974–984. [Google Scholar] [CrossRef]
- Tyler, H.L. Winter cover crops and no till management enhance enzyme activities in soybean field soils. Pedobiologia 2020, 81-82, 150666. [Google Scholar] [CrossRef]
- Salvagiotti, F.; Cassman, K.; Specht, J.; Walters, D.; Weiss, A.; Dobermann, A. Nitrogen uptake, fixation and response to fertilizer N in soybeans: A review. Field Crop. Res. 2008, 108, 1–13. [Google Scholar] [CrossRef] [Green Version]
- Sullivan, P.G.; Parrish, D.J.; Luna, J.M. Cover crop contributions to N supply and water conservation in corn production. Am. J. Altern. Agric. 1991, 6, 106–113. [Google Scholar] [CrossRef]
- Pagano, M.C.; Miransari, M. The importance of soybean production worldwide. In Abiotic and Biotic Stresses in Soybean Production; Miransari, M., Ed.; Academic Press: San Diego, CA, USA, 2016; pp. 1–26. [Google Scholar]
- Horwath, W.R.; Paul, E.A. Microbial Biomass. In Methods of Soil Analysis: Part 2—Microbiological and Biochemical Properties; Bottomley, P.S., Angle, J.S., Weaver, R.W., Eds.; Soil Science Society of America Book Series: Madison, WI, USA, 1994; pp. 753–773. [Google Scholar]
- Jackson, C.R.; Tyler, H.L.; Millar, J. Determination of Microbial Extracellular Enzyme Activity in Waters, Soils, and Sediments using High Throughput Microplate Assays. J. Vis. Exp. 2013, 80, e50399. [Google Scholar] [CrossRef] [Green Version]
- Schnürer, J.; Rosswall, T. Fluorescein Diacetate Hydrolysis as a Measure of Total Microbial Activity in Soil and Litter. Appl. Environ. Microbiol. 1982, 43, 1256–1261. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jensen, E.S. Barley uptake of N deposited in the rhizosphere of associated field pea. Soil Biol. Biochem. 1996, 28, 159–168. [Google Scholar] [CrossRef]
- Keating, B.A.; Carberry, P.S. Resource capture and use in intercropping: Solar radiation. Field Crop. Res. 1993, 34, 273–301. [Google Scholar] [CrossRef]
- Reddy, K.N.; Zablotowicz, R.M.; Locke, M.A.; Koger, C.H. Cover crop, tillage, and herbicide effects on weeds, soil properties, microbial populations, and soybean yield. Weed Sci. 2003, 51, 987–994. [Google Scholar] [CrossRef]
- Moore, E.B.; Wiedenhoeft, M.H.; Kaspar, T.C.; Cambardella, C.A. Rye Cover Crop Effects on Soil Quality in No-Till Corn Silage-Soybean Cropping Systems. Soil Sci. Soc. Am. J. 2014, 78, 968–976. [Google Scholar] [CrossRef]
- Blanco-Canqui, H.; Lal, R. No-Tillage and Soil-Profile Carbon Sequestration: An On-Farm Assessment. Soil Sci. Soc. Am. J. 2008, 72, 693–701. [Google Scholar] [CrossRef] [Green Version]
- Zablotowicz, R.M.; Reddy, K.N.; Weaver, M.A.; Mengistu, A.; Krutz, L.J.; Gordon, R.E.; Bellaloui, N. Cover crops, tillage, and glyphosate effects on chemical and biological properties of a lower Mississippi Delta soil and soybean yield. Environ. Res. J. 2010, 4, 227–251. [Google Scholar]
- Lupwayi, N.Z.; Larney, F.J.; Blackshaw, R.E.; Kanashiro, D.A.; Pearson, D.C. Phospholipid fatty acid biomarkers show positive soil microbial community responses to conservation soil management of irrigated crop rotations. Soil Tillage Res. 2017, 168, 1–10. [Google Scholar] [CrossRef]
- Cordeiro, C.F.D.S.; Rodrigues, D.R.; Rocha, C.H.; Araujo, F.F.; Echer, F.R. Glomalin and microbial activity affected by cover crops and nitrogen management in sandy soil with cotton cultivation. Appl. Soil Ecol. 2021, 167, 104026. [Google Scholar] [CrossRef]
- Calderón, F.J.; Nielsen, D.; Acosta-Martínez, V.; Vigil, M.F.; Lyon, D. Cover Crop and Irrigation Effects on Soil Microbial Communities and Enzymes in Semiarid Agroecosystems of the Central Great Plains of North America. Pedosphere 2016, 26, 192–205. [Google Scholar] [CrossRef]
- Bending, G.D.; Turner, M.K.; Jones, J.E. Interactions between crop residue and soil organic matter quality and the functional diversity of soil microbial communities. Soil Biol. Biochem. 2002, 34, 1073–1082. [Google Scholar] [CrossRef]
- Woodward, J. Synergism in cellulase systems. Bioresour. Technol. 1991, 36, 67–75. [Google Scholar] [CrossRef]
- Ekenler, M.; Tabatabai, M.A. β-Glucosaminidase Activity as an Index of Nitrogen Mineralization in Soils. Commun. Soil Sci. Plant Anal. 2004, 35, 1081–1094. [Google Scholar] [CrossRef]
- Turner, B.L.; McKelvie, I.D.; Haygarth, P.M. Characterisation of water-extractable soil organic phosphorus by phosphatase hydrolysis. Soil Biol. Biochem. 2002, 34, 27–35. [Google Scholar] [CrossRef]
- Guilbault, G.G.; Kramer, D.N. Fluorometric Determination of Lipase, Acylase, Alpha-, and Gamma-Chymotrypsin and Inhibitors of These Enzymes. Anal. Chem. 1964, 36, 409–412. [Google Scholar] [CrossRef]
- Rotman, B.; Papermaster, B.W. Membrane properties of living mammalian cells as studied by enzymatic hydrolysis of fluorogenic esters. Proc. Natl. Acad. Sci. USA 1966, 55, 134–141. [Google Scholar] [CrossRef] [Green Version]
- Webber, C.L.; Gebhardt, M.R.; Kerr, H.D. Effect of Tillage on Soybean Growth and Seed Production. Agron. J. 1987, 79, 952–956. [Google Scholar] [CrossRef]
- Tyler, H.L. Shifts in bacterial community in response to conservation management practices within a soybean production system. Biol. Fertil. Soils 2021, 57, 1–12. [Google Scholar] [CrossRef]
- Bakker, M.G.; Acharya, J.; Moorman, T.B.; Robertson, A.E.; Kaspar, T.C. The Potential for Cereal Rye Cover Crops to Host Corn Seedling Pathogens. Phytopathology 2016, 106, 591–601. [Google Scholar] [CrossRef] [Green Version]
- Sturz, A.V.; Carter, M.R.; Johnston, H.W. A review of plant disease, pathogen interactions and microbial antagonism under conservation tillage in temperate humid agriculture. Soil Tillage Res. 1997, 41, 169–189. [Google Scholar] [CrossRef]
- Singh, G.; Thilakarathne, A.D.G.M.; Williard, K.W.J.; Schoonover, J.E.; Cook, R.L.; Gage, K.L.; McElroy, R. Tillage and legume non-legume cover cropping effects on corn–soybean production. Agron. J. 2020, 112, 2636–2648. [Google Scholar] [CrossRef]
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the author. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).