The Influence of Tillage and Cover Cropping on Soil Microbial Parameters and Spring Wheat Physiology

 ,
, 

Abstract
1. Introduction
2. Materials and Methods
2.1. Microbiological Analyses
2.1.1. Counts of Microorganisms
- total bacterial count—on ready-made Merck standard agar after 5 days of incubation at 25 °C,
- fungi—on Martin agar [10] after 5 days of incubation at 24 °C,
- copiotrophs—on nutrient broth (NB) agar [11] after 5 days of incubation at 25 °C,
- oligotrophs—on diluted nutrient broth (DNB) agar [11] after 5 days of incubation at 25 °C and
- actinobacteria—on Pochon agar after 5 days of incubation at 25 °C [12].
2.1.2. Analyses of Enzymatic Activity
- dehydrogenases (EC 1.1.1.)—colorimetry with 1% TTC (triphenyltetrazolium chloride) as a substrate after 24-hour incubation at 30 °C at a wavelength of 485 nm, expressed as μmol triphenylformazan (TPF) kg−1/24 h−1 [13];
- acid and alkaline phosphatase (EC 3.1.3.2)—spectrophotometry with sodium p-nitrophenyl phosphate as a substrate after 1-hour incubation at 37 °C at a wavelength of 400 nm, expressed as μmol para-nitrophenol (PNP) g−1dm/h−1 (Novospac spectrophotometer) [14] and
- catalase (EC 1.11.1.6)—permanganometry with 0.3% hydrogen peroxide (H2O2) as a substrate after 20 minutes incubation at room temperature (about 20 °C), titrated with 0.02 M potassium permanganate (KMnO4) to light pink colour and expressed as mmol H2O2 g−1dm/min−1 [15].
2.1.3. Biological Index of Fertility
2.2. Genetic Analyses
- automatic demultiplexing of the samples,
- generating FASTQ files containing raw reads and
- classification of paired-end reads into individual taxonomic categories.
2.3. Physiological State of Plants
2.4. Statistical Analysis
- conventional tillage (CT) before sowing of spring wheat without a cover crop,
- CT skimming before sowing of the cover crop and sowing of spring wheat after CT,
- skimming before sowing of the cover crop and sowing of spring wheat directly (no-till (NT)) into mulch,
- sowing of the cover crop directly (NT) into the stubble and then sowing of spring wheat after CT and
- NT and sowing of spring wheat directly into mulch (NT).
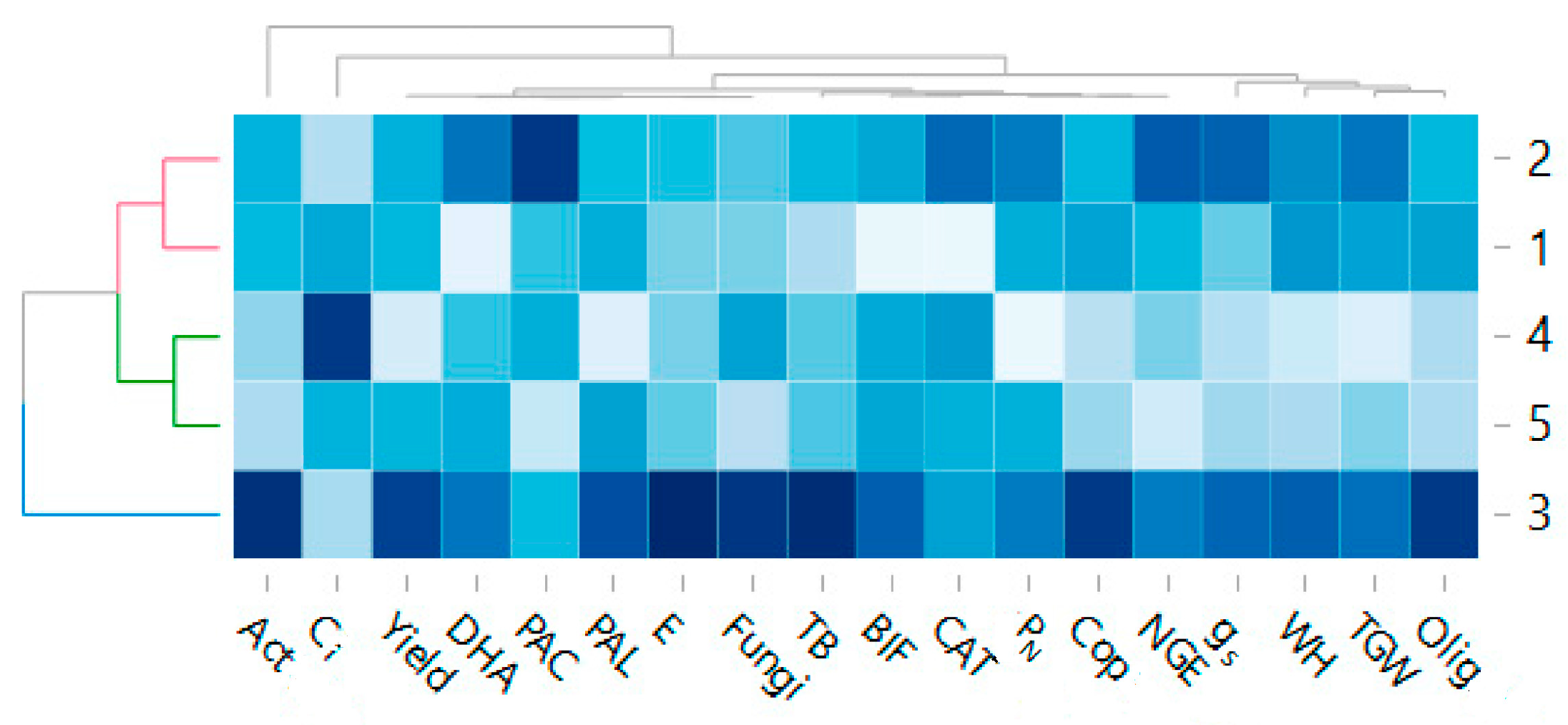
3. Results and Discussion
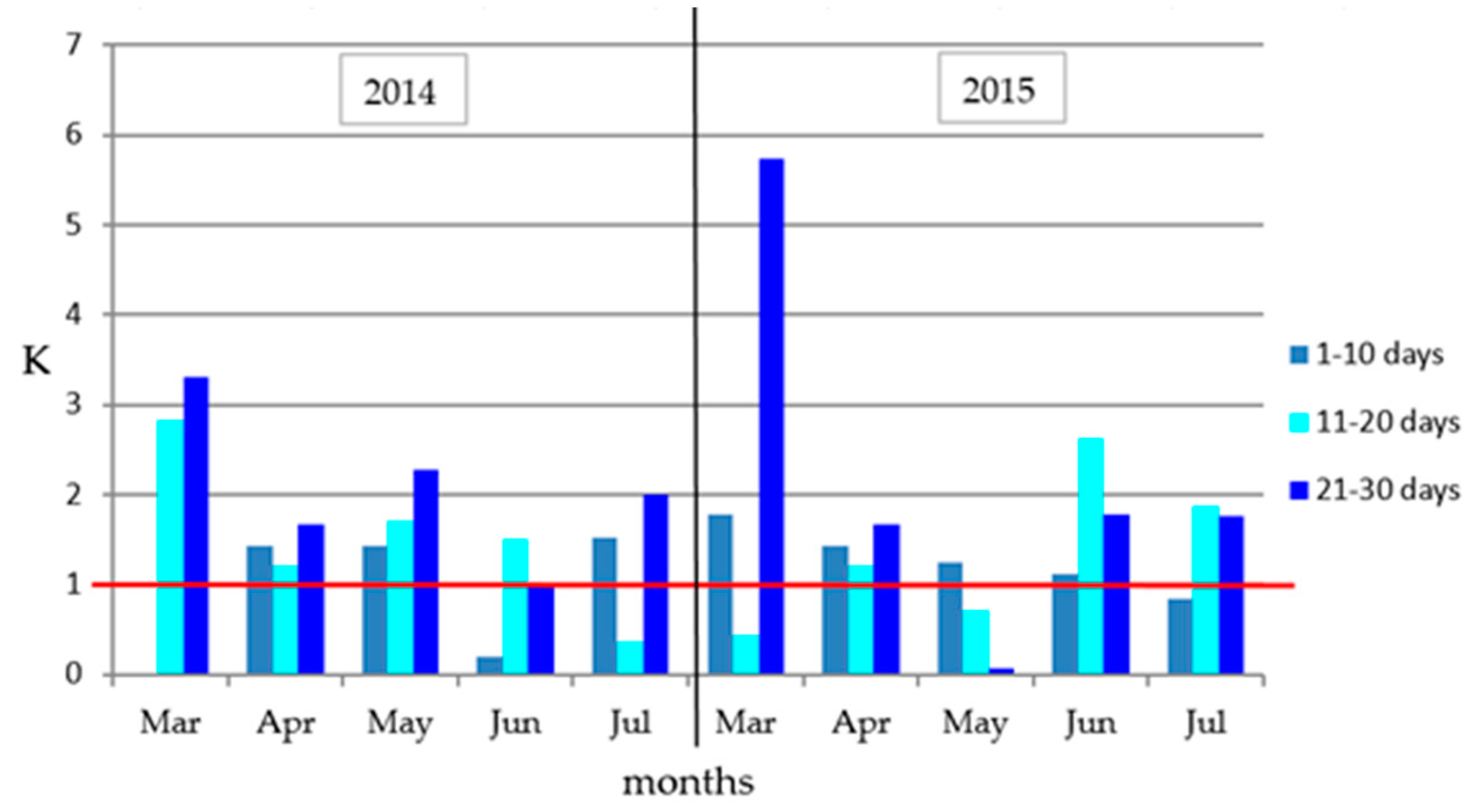
3.1. Weather Data
3.2. Soil Organic Matter Content
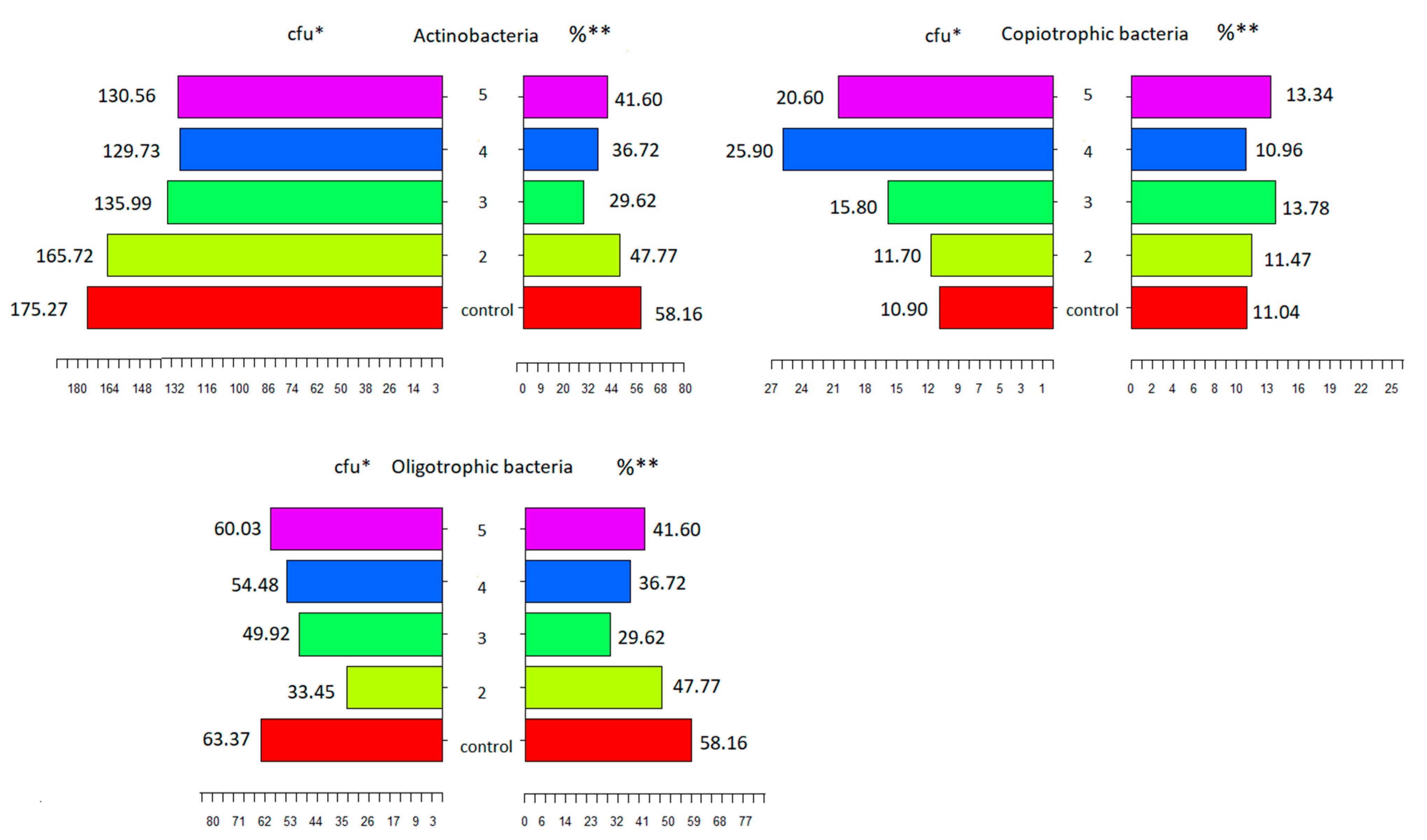
3.3. The Count of Microorganisms
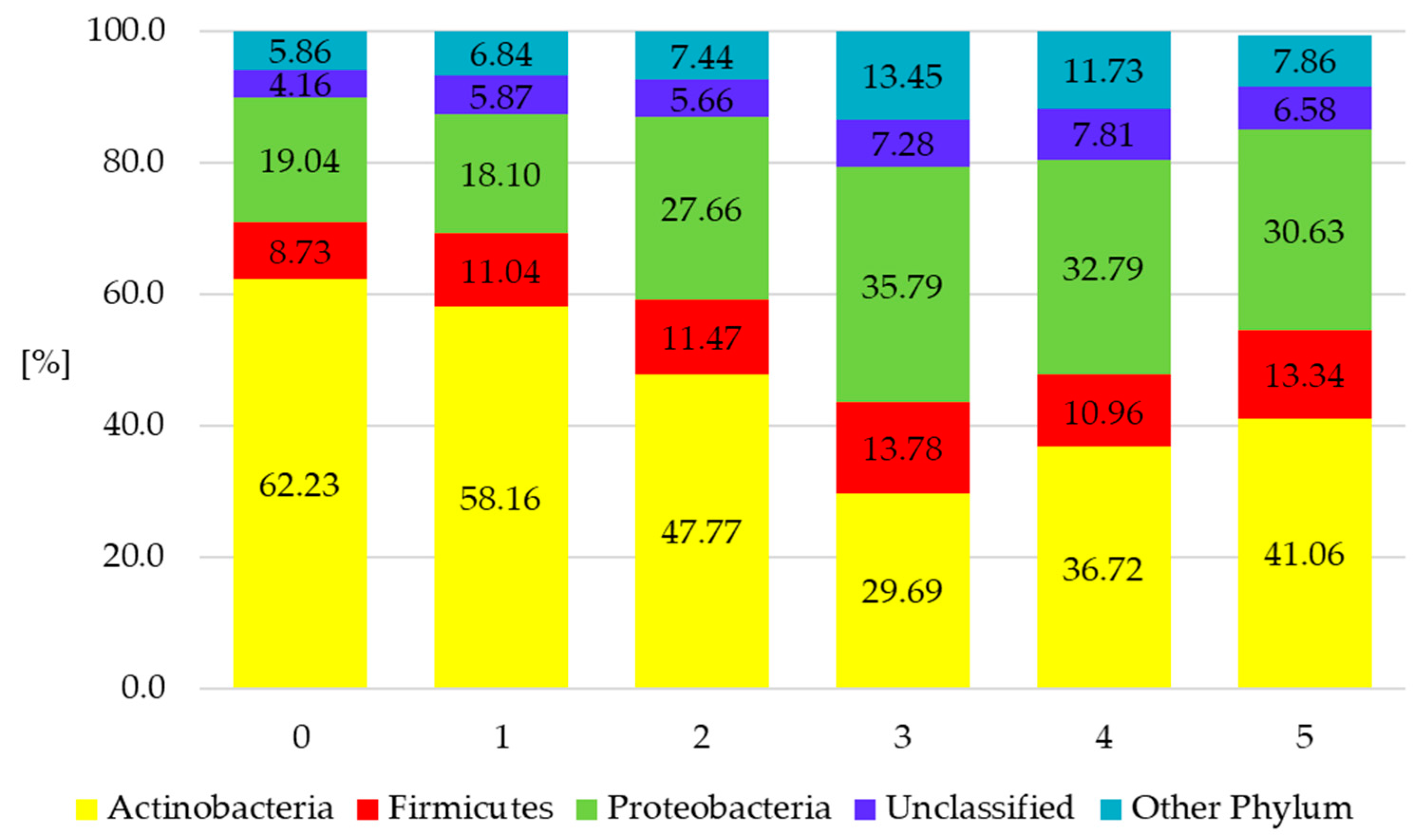
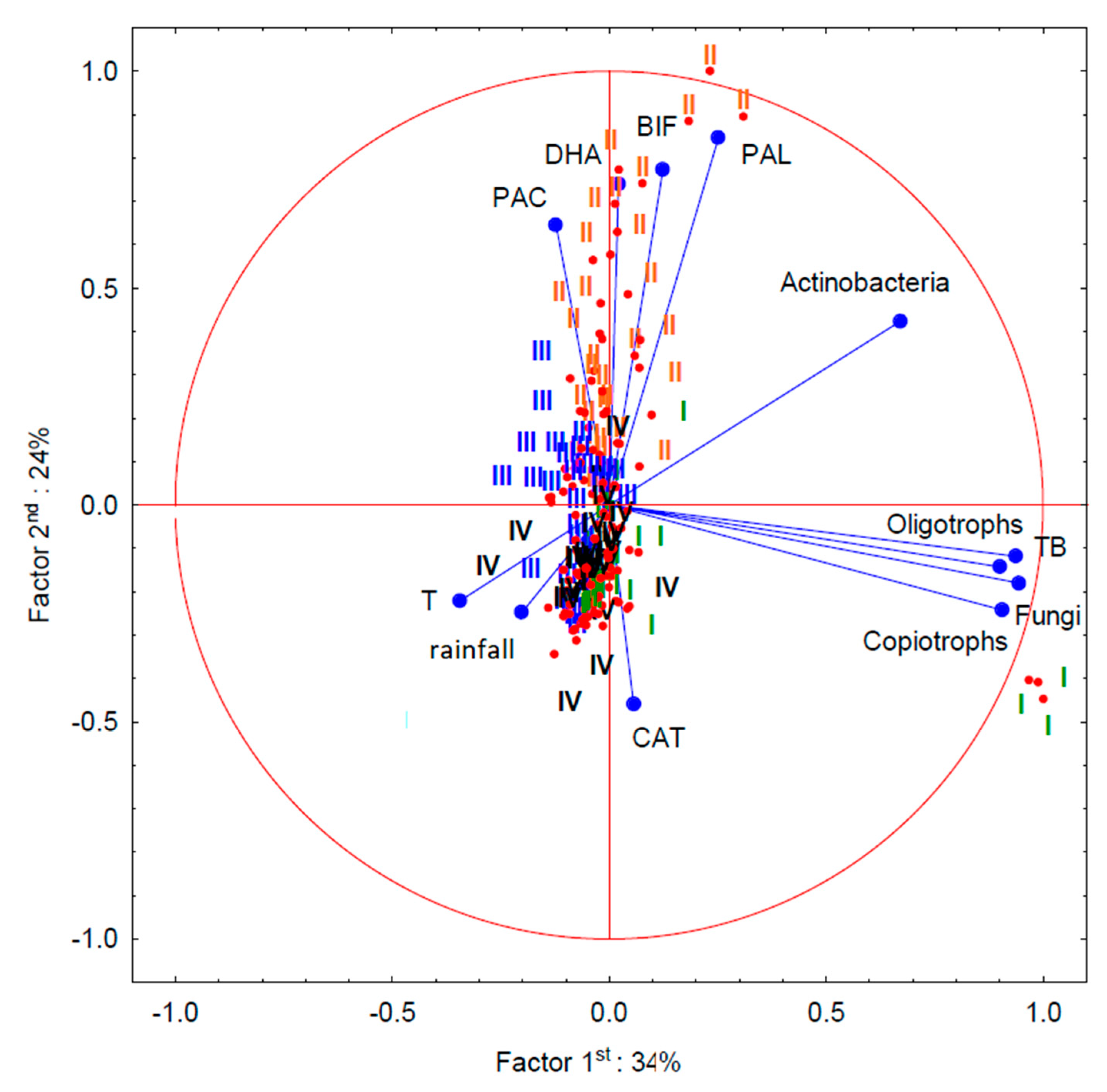
3.4. Influence of Various Cultivation Methods on Taxonomic Distribution of Soil Bacteria
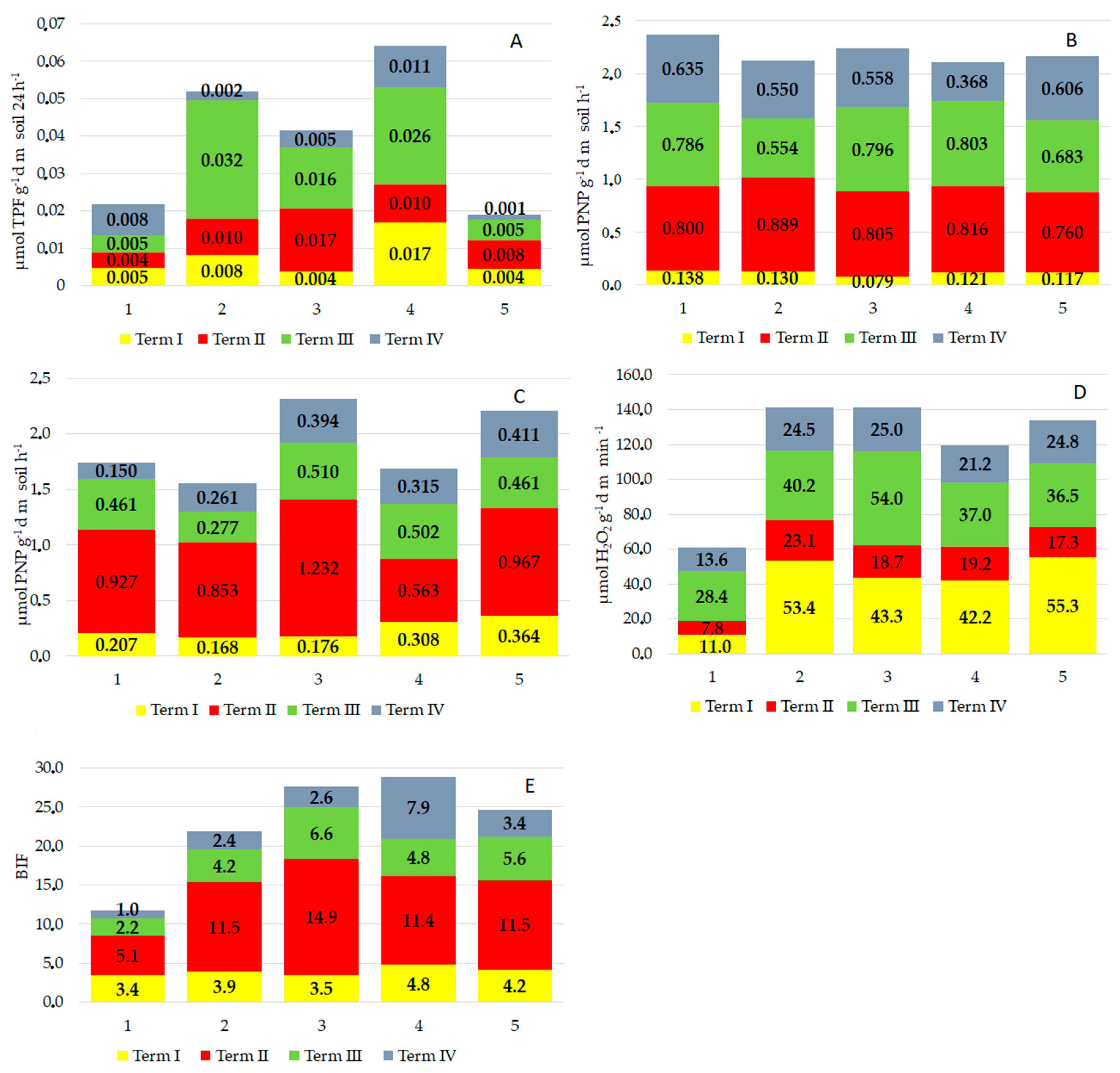
3.5. Biochemical Analyses of Soil
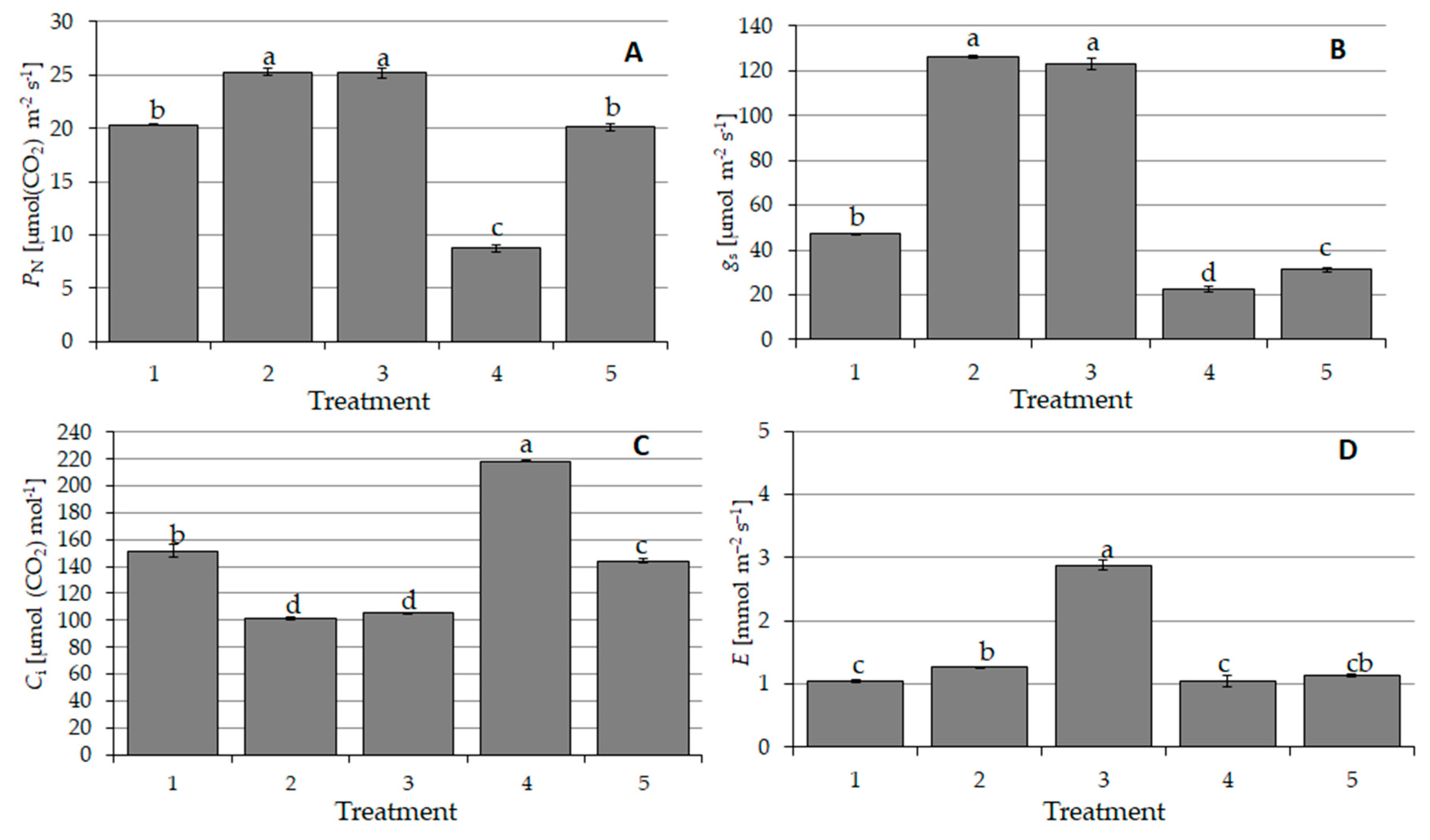
3.6. Physiological State of Wheat
4. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Hobbs, P.R.; Sayre, K.; Gupta, R. The role of conservation agriculture in sustainable agriculture. Philos. T. R. Soc. B. 2008, 363, 543–555. [Google Scholar] [CrossRef]
- Mbuthia, L.W.; Acosta-Martínez, V.; DeBruyn, J.; Schaeffer, S.; Tyler, D.; Odoi, E.; Mpheshea, M.; Walker, F.; Eash, N. Long term tillage, cover crop, and fertilization effects on microbial community structure, activity: Implications for soil quality. Soil Biol. Biochem. 2015, 89, 24–34. [Google Scholar] [CrossRef]
- Yao, R.J.; Yang, J.S.; Zhang, T.J.; Gao, P.; Yu, S.P.; Wang, X.P. Short-term effect of cultivation and crop rotation systems on soil quality indicators in a coastal newly reclaimed farming area. J. Soils Sed. 2013, 13, 1335–1350. [Google Scholar] [CrossRef]
- Bünemann, E.K.; Schwenke, G.D.; Van Zwieten, L. Impact of agricultural inputs on soil organisms—A review. Aust. J. Soil Res. 2006, 44, 379–406. [Google Scholar] [CrossRef]
- Malézieux, E.; Crozat, Y.; Dupraz, C.; Laurans, M.; Makowski, D.; Ozier-Lafontaine, H.; Rapidel, B.; de Tourdonnet, S.; Valantin-Morison, M. Mixing plant species in cropping systems: Concepts, tools and models: A review. In Sustainable agriculture; Springer: Berlin, Germany, 2009; pp. 329–353. [Google Scholar]
- Bocianowski, J.; Majchrzak, L. Analysis of effects of cover crop and tillage method combinations on the phenotypic traits of spring wheat (Triticum aestivum L.) using multivariate methods. Appl. Ecol. Environ. Res. 2019, 17, 15267–15276. [Google Scholar]
- Yang, Y.; Ding, J.; Zhang, Y.; Wu, J.; Zhang, J.; Pan, X.; Gao, C.; Wang, Y.; He, F. Effects of tillage and mulching measures on soil moisture and temperature, photosynthetic characteristics and yield of winter wheat. Agric. Water Manag. 2018, 201, 299–308. [Google Scholar] [CrossRef]
- Ahemad, M.; Khan, S.M. Functional aspects of plant growth promoting rhizobacteria: Recent Advancements. Insight Microbiol. 2011, 1, 39–54. [Google Scholar] [CrossRef]
- Bielińska, E.J.; Mocek-Płóciniak, A. Impact of the tillage system on the soil enzymatic activity. Archiv. Environ. Prot. 2012, 38, 75–82. [Google Scholar] [CrossRef]
- Martin, J.P. Use of acid, rose bengal and streptomycin in the plate method for estimating soil fungi. Soil Sci. 1950, 69, 215–232. [Google Scholar] [CrossRef]
- Ohta, H.; Hattori, T. Bacteria sensitive to nutrient broth medium in terrestrial environments. Soil Sci. Plant Nutr. 1980, 26, 99–107. [Google Scholar] [CrossRef]
- Grabińska-Łoniewska, A. Laboratory Classes in General Microbiology; Oficyna Wydawnicza Politechniki: Warsaw, Poland, 1999. [Google Scholar]
- Thalmann, A. Zur Methodik der Bestimmung der Dehydrogenase aktivität im Boden mittels triphenytetrazoliumchlorid (TTC). Landwirtsch Forsch. 1968, 21, 249–258. [Google Scholar]
- Tabatabai, M.A.; Bremner, J.M. Use of p-nitrophenyl phosphate for assay of soil phosphatase activity. Soil Biol. Biochem. 1969, 1, 301–307. [Google Scholar] [CrossRef]
- Johnson, J.L.; Temple, K.L. Some Variables Affecting the Measurement of “Catalase Activity” in Soil 1. Soil Sci. Soc. Am. 1964, 28, 207–209. [Google Scholar] [CrossRef]
- Saviozzi, A.; Levi-Minzi, R.; Cardelli, R.; Riffaldi, R. A comparison of soil quality in adjacent cultivated, forest and native grassland soils. Plant Soil 2001, 233, 251–259. [Google Scholar] [CrossRef]
- Stachowski, P. Evaluation of meteorological droughts in post-mining areas in Poland in Konin area—Central Pomeranian Scientific Society for Environmental Protection. Ann. Set Environ. Prot. 2010, 12, 587. [Google Scholar]
- Thomas, G.A.; Dalal, R.C.; Standley, J. No-till effects on organic matter, pH, cation exchange capacity and nutrient distribution in a Luvisol in the semi-arid subtropics. Soil Tillage Res. 2007, 94, 295–304. [Google Scholar] [CrossRef]
- López-Fando, C.; Pardo, M.T. Changes in soil chemical characteristics with different tillage practices in a semi-arid environment. Soil Tillage Res. 2009, 104, 278–284. [Google Scholar] [CrossRef]
- Plaza-Bonilla, D.; Nolot, J.M.; Passot, S.; Raffaillac, D.; Justes, E. Grain legume-based rotations managed under conventional tillage need cover crops to mitigate soil organic matter losses. Soil Tillage Res. 2016, 156, 33–43. [Google Scholar] [CrossRef]
- Kraska, P.; Mielniczuk, E. The occurrence of fungi on the stem base and roots of spring wheat (Triticum aestivum L.) grown in monoculture depending on tillage systems and catch crops. Acta Agrobot. 2012, 65, 79–90. [Google Scholar] [CrossRef]
- Majchrzak, L.; Sawinska, Z.; Natywa, M.; Skrzypczak, G.; Głowicka-Wołoszyn, R. Impact of different tillage systems on soil dehydrogenase activity and spring wheat infection. J. Agric. Sci. Technol. 2016, 18, 1871–1881. [Google Scholar]
- Arcand, M.M.; Helgason, B.L.; Lemke, R.L. Microbial crop residue decomposition dynamics in organic and conventionally managed soils. Appl. Soil Ecol. 2016, 107, 347–359. [Google Scholar] [CrossRef]
- Hartman, W.H.; Richardson, C.J. Differential nutrient limitation of soil microbial biomass and metabolic quotients (qCO2): Is there a biological stoichiometry of soil microbes? PLoS ONE 2013, 8, e57127. [Google Scholar] [CrossRef]
- Nacke, H.; Thürmer, A.; Wollherr, A.; Will, C.; Hodac, L.; Herold, N.; Schöning, I.; Schrumpf, M.; Danie, R. Pyrosequencing-based assessment of bacterial community structure along different management types in German forest and grassland soils. PLoS ONE 2011, 6, e17000. [Google Scholar] [CrossRef]
- Navarro-Noya, Y.E.; Gómez-Acata, S.; Montoya-Ciriaco, N.; Rojas-Valdez, A.; Suárez-Arriaga, M.C.; Valenzuela-Encinas, C.; Jiménez-Bueno, N.; Verhulst, N.; Govaerts, B.; Dendooven, L. Relative impacts of tillage, residue management and crop-rotation on soil bacterial communities in a semi-arid agroecosystem. Soil Biol. Biochem. 2013, 65, 86–95. [Google Scholar] [CrossRef]
- Wang, Z.; Liu, L.; Chen, Q.; Wen, X.; Liao, Y. Conservation tillage increases soil bacterial diversity in the dryland of northern China. Agron. Sustain. Dev. 2016, 36, 28. [Google Scholar] [CrossRef]
- Fierer, N.; Bradford, M.A.; Jackson, R.B. Toward an ecological classification of soil bacteria. Ecology 2007, 88, 1354–1364. [Google Scholar] [CrossRef]
- Rodrigues, J.L.; Pellizari, V.H.; Mueller, R.; Baek, K.; Jesus, E.D.C.; Paula, F.S.; Mirza, B.; Hamaoui, G.S., Jr.; Tsai, S.M.; Feigl, B.; et al. Conversion of the Amazon rainforest to agriculture results in biotic homogenization of soil bacterial communities. Proc. Natl. Acad. Sci. USA 2013, 110, 988–993. [Google Scholar] [CrossRef]
- Pascault, N.; Ranjard, L.; Kaisermann, A.; Bachar, D.; Christen, R.; Terrat, S.; Mathieu, O.; Lévêque, J.; Mougel, C.; Henault, C.; et al. Stimulation of different functional groups of bacteria by various plant residues as a driver of soil priming effect. Ecosystems 2013, 16, 810–822. [Google Scholar] [CrossRef]
- Swędrzyńska, D.; Małecka, I.; Blecharczyk, A.; Swędrzyński, A.; Starzyk, J. Effects of various long-term tillage systems on some chemical and biologicalpProperties of soil. Pol. J. Environ. Stud. 2013, 22, 1835–1844. [Google Scholar]
- Niewiadomska, A.; Sulewska, H.; Wolna-Maruwka, A.; Ratajczak, K.; Głuchowska, K.; Waraczewska, Z.; Budka, A. An assessment of the influence of Co-inoculation with endophytic bacteria and rhizobia, and the influence of PRP SOL and PRP EBV fertilisers on the microbial parameters of soil and nitrogenase activity in yellow lupine (Lupinus luteus L.) cultivation. Pol. J. Environ. Stud. 2018, 27, 2687–2702. [Google Scholar]
- Melero, S.; Panettieri, M.; Madejón, E.; Gómez Macpherson, H.; Moreno, F.; Murillo, J.M. Implementation of chiseling and mouldboard ploughing in soil after 8 years of no-till management in SW, Spain: Effect on soil quality. Soil Tillage Res. 2011, 112, 107–113. [Google Scholar] [CrossRef]
- Singh, G.; Bhattacharyya, R.; Das, T.K.; Sharma, A.R.; Ghosh, A.; Das, S.; Jha, P. Crop rotation and residue management effects on soil enzyme activities, glomalin and aggregate stability under zero tillage in the Indo-Gangetic Plains. Soil Tillage Res. 2018, 184, 291–300. [Google Scholar] [CrossRef]
- Morris, N.L.; Miller, P.C.H.; Orson, J.H.; Froud-Williams, R.J. The adoption of non-inversion tillage systems in the United Kingdom and the agronomic impact on soil, crops and the environment—A review. Soil Tillage Res. 2010, 108, 1–15. [Google Scholar] [CrossRef]
- Żebrowska, E.; Ciereszko, I. Acid phosphatases role in plant cells phosphate homeostasis. Adv. Cell Biol. 2009, 36, 583–599. [Google Scholar]
- Lemanowicz, J.; Koper, J. Activity of phosphatase and the content of phosphorus in soil under selected crops fertilised with slurry. Proc. ECOpole 2012, 6, 239–243. [Google Scholar]
- Niewiadomska, A.; Sulewska, H.; Wolna-Maruwka, A.; Ratajczak, K.; Waraczewska, Z.; Budka, A.; Głuchowska, K. The influence of biostimulants and foliar fertilisers on the process of biological nitrogen fixation and the level of soil biochemical activity in soybean (Glycine max L.) Cultivation. Appl. Ecol. Environ. Res. 2019, 17, 12649–12666. [Google Scholar] [CrossRef]
- Lemanowicz, J.; Koper, J. Changes in the content of available phosphorus and soil phosphatase activity as a result of fertilization. Soil Sci. Ann. 2010, 61, 140–145. [Google Scholar]
- Heflik, M.; Kandziora, M.; Nadgórska-Socha, A.; Ciepal, R. Acid phosphatase activity in plants grown in heavy metals contaminated sites. Environ. Prot. Nat. Res. 2007, 32, 151–154. [Google Scholar]
- Bonanomi, G.; Chiurazzi, M.; Caporaso, S.; Del Sorbo, G.; Moschetti, G.; Felice, S. Soil solarization with biodegradable materials and its impact on soil microbial communities. Soil Biol. Biochem. 2008, 40, 1989–1998. [Google Scholar] [CrossRef]
- James, D.W.; Suslow, T.V.; Steinback, K.E. Relationship between rapid, firm adhesion and long-term colonization of roots by bacteria. Appl. Environ. Microbiol. 1985, 50, 392–397. [Google Scholar] [CrossRef] [PubMed]
- Pięta, D.; Patkowska, E. Effect of root exudates of various plants on composition of bacteria and fungi communities with special regard to pathogenic soil-borne fungi. Acta Agrobot. 2001, 54, 95–104. [Google Scholar] [CrossRef]
- Siwik-Ziomek, A.; Szczepanek, M. Soil Extracellular Enzyme Activities and Uptake of N by Oilseed Rape Depending on Fertilization and Seaweed Biostimulant Application. Agron. J. 2019, 9, 480. [Google Scholar] [CrossRef]
- Pausch, J.; Kuzyakov, Y. Photoassimilate allocation and dynamics of hotspots in roots visualized by 14C phosphor imaging. J. Plant Nutr. Soil Sci. 2011, 174, 12–19. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Treatment | Soil Tillage System Under Spring Wheat | Dose of Glyphosate + Adjuvant AS 500 SL and Time of Application | Soil Tillage System Under Cover Crop (Yellow Lupine + Field Pea) |
|---|---|---|---|
| 1 | CT (spring ploughing) | - | - |
| 2 | CT (spring ploughing) | - | CT (skimming) |
| 3 | NT (no tillage) | 1.5 L/ha−1 + 1.5 L/ha−1 (before spring wheat sowing) | CT (skimming) |
| 4 | CT (spring ploughing) | 4.0 L/ha−1 + 1.5 L/ha−1 (before legume sowing) | NT (no tillage) |
| 5 | NT (no tillage) | 4.0 L/ha−1 + 1.5 L/ha−1 (before legume sowing) 1.5 L/ha−1 + 1.5 L/ha−1 (before spring wheat sowing) | NT (no tillage) |
| Month | Days | Term of Analyses | Average Temperature (°C) | Average Rainfall (mm) | ||
|---|---|---|---|---|---|---|
| 2014 | 2015 | 2014 | 2015 | |||
| March | 1–10 | 4.3 | 4.7 | 0.0 | 8.4 | |
| 11–20 | 7.7 | 5.0 | 21.8 | 2.2 | ||
| 21–31 | 7.9 | 5.8 | 26.0 | 33.3 | ||
| Average/Total | 6.6 | 5.2 | 47.8 | 43.9 | ||
| Long-term | 2.9 | 39.2 | ||||
| April | 1–10 | 8.8 | 4.8 | 12.5 | 25.0 | |
| 11–20 | 8.7 | 9.4 | 10.5 | 0.3 | ||
| 21–30 | 13.9 | 11.5 | 23.3 | 6.7 | ||
| Average/Total | 10.5 | 8.6 | 46.3 | 32.0 | ||
| Long-term | 8.1 | 37.4 | ||||
| May | 1–10 | 10.1 | 12.8 | 14.5 | 15.9 | |
| 11–20 | 1st term (BBCH 16–17) | 12.6 | 12.6 | 21.4 | 8.9 | |
| 21–31 | 16.5 | 13.6 | 37.6 | 0.8 | ||
| Average/Total | 13.1 | 13.0 | 73.5 | 25.6 | ||
| Long-term | 13.2 | 57.4 | ||||
| June | 1–10 | 17.5 | 16.3 | 3.3 | 18.3 | |
| 11–20 | 16.0 | 15.4 | 24.0 | 40.3 | ||
| 21–30 | 2nd term (BBCH 61–65) | 14.8 | 15.0 | 14.7 | 26.7 | |
| Average/Total | 16.1 | 15.5 | 42.0 | 85.3 | ||
| Long-term | 16.6 | 63.8 | ||||
| July | 1–10 | 20.5 | 20.3 | 31.0 | 17.0 | |
| 11–20 | 3rd term (BBCH 87–89) | 21.7 | 19.1 | 7.8 | 35.8 | |
| 21–31 | 4th term after the harvest | 22.2 | 18.2 | 44.3 | 32.1 | |
| Average/Total | 21.5 | 19.2 | 83.1 | 84.9 | ||
| Long-term | 18.3 | 81.3 | ||||
| Specification | Investigation Term | Treatments | ||||
|---|---|---|---|---|---|---|
| 1 * | 2 | 3 | 4 | 5 | ||
| C organic (g/kg−1 soil) | A ** | 10.8 | 12.9 | 10.1 | 10.9 | 10.4 |
| B ** | 10.7 | 11.8 | 11.0 | 12.4 | 11.9 | |
| N total (g/kg−1 soil) | A | 0.7 | 1.1 | 1.0 | 1.0 | 1.2 |
| B | 0.9 | 1.2 | 1.0 | 1.3 | 0.9 | |
| C:N | A | 12.6 | 14.4 | 11.9 | 11.8 | 10.2 |
| B | 10.5 | 13.2 | 12.3 | 10.0 | 14.3 | |
| Parameter | Term | Treatment | Interaction |
|---|---|---|---|
| Total count of bacteria | 87.78 * | 120.67 * | 99.67 * |
| Actinobacteria | 58.27 * | 21.60 * | 17.58 * |
| Fungi | 54.53 * | 74.44 * | 85.66 * |
| Oligotrophic bacteria | 45.96 * | 23.6 * | 35.58 * |
| Copiotrophic bacteria | 89.89 * | 101.92 * | 111.99 * |
| Experimental Treatment | Term of Analysis | |||
|---|---|---|---|---|
| 1st Term Emergence (BBCH 16–17) | 2nd Term Flowering (BBCH 61–65) | 3rd Term Technological Ripeness of Seeds (BBCH 87–89) | 4th Term After Harvest | |
| Total count of heterotrophic bacteria (cfu g−1/dm soil 105) | ||||
| 1 | 4.7 ± 3.8 | 6.3 ± 1.9 | 6.4 ± 2.4 | 14.5 ± 4.6 |
| 2 | 30.8 ± 12.1 | 20.3 ± 20.8 | 22.5 ± 3.8 | 45.5 ± 5.1 |
| 3 | 30.3 ± 7.1 | 55.4 ± 12.3 | 37.9 ± 7.6 | 26.5 ± 5.0 |
| 4 | 22.8 ± 8.4 | 15.0 ± 2.3 | 12.8 ± 4.3 | 40.4 ± 5.2 |
| 5 | 162.3 ± 52.8 | 15.9 ± 4.1 | 14.6 ± 4.5 | 37.9 ± 5.2 |
| Actinobacteria (cfu g−1/dm soil 104) | ||||
| 1 | 232.9 ± 58.0 | 243.1 ± 47.0 | 193.8 ± 30.6 | 175.3 ± 3.2 |
| 2 | 204.8 ± 79.8 | 231.4 ± 38.5 | 117.2 ± 27.5 | 165.7 ± 5.0 |
| 3 | 126.2 ± 21.8 | 247.9 ± 33.0 | 133.6 ± 27.8 | 136.0 ± 14.6 |
| 4 | 131.7 ± 45.9 | 149.2 ± 24.9 | 43.4 ± 13.7 | 129.7 ± 5.9 |
| 5 | 202.5 ± 31.0 | 125.2 ± 21.4 | 55.7 ± 18.1 | 130.6 ± 4.6 |
| Fungi (cfu g−1/dm soil 104) | ||||
| 1 | 0.3 ± 0.0 | 1.4 ± 0.6 | 1.4 ± 1.3 | 1.1 ± 0.7 |
| 2 | 1.5 ± 2.1 | 2.6 ± 0.9 | 2.3 ± 1.7 | 3.4 ± 1.0 |
| 3 | 0.6 ± 0.6 | 3.9 ± 0.1 | 4.1 ± 3.8 | 3.0 ± 1.2 |
| 4 | 3.2 ± 4.1 | 2.4 ± 0.2 | 3.8 ± 1.9 | 4.2 ± 1.2 |
| 5 | 33.2 ± 16.4 | 2.9 ± 0.3 | 2.0 ± 1.3 | 2.6 ± 2.1 |
| Oligotrophic bacteria (cfu g−1/dm soil 105) | ||||
| 1 | 35.8 ± 9.7 | 49.7 ± 9.2 | 33.0 ± 3.9 | 62.4 ± 2.5 |
| 2 | 37.1 ± 13.9 | 41.1 ± 5.5 | 22.3 ± 5.5 | 33.4 ± 3.1 |
| 3 | 38.6 ± 10.7 | 32.1 ± 3.6 | 33.3 ± 8.8 | 49.9 ± 5.0 |
| 4 | 56.9 ± 18.0 | 23.7 ± 5.3 | 31.6 ± 9.6 | 54.5 ± 5.6 |
| 5 | 150.3 ± 27.4 | 23.5 ± 9.1 | 32.6 ± 5.2 | 60.0 ± 2.5 |
| Copiotrophic bacteria (cfu g−1/dm soil 105) | ||||
| 1 | 14.1 ± 8.3 | 19.9 ± 4.6 | 12.2 ± 5.0 | 10.9 ± 5.2 |
| 2 | 14.6 ± 7.4 | 19.3 ± 5.4 | 19.1 ± 2.1 | 11.7 ± 5.3 |
| 3 | 20.2 ± 6.3 | 27.5 ± 7.2 | 21.9 ± 3.7 | 15.8 ± 5.5 |
| 4 | 10.1 ± 6.1 | 25.1 ± 6.5 | 13.3 ± 4.8 | 25.9 ± 5.9 |
| 5 | 74.6 ± 11.2 | 26.2 ± 5.2 | 15.2 ± 4.3 | 20.6 ± 5.5 |
| Parameter | Term | Treatment | Interaction |
|---|---|---|---|
| Dehydrogenase activity (DHA) | 45.3 * | 11.2 * | 4.9 * |
| Acid phosphatase activity (PAC) | 25.5 * | 1.4 ns | 3.9 * |
| Alkaline phosphatase activity (PAL) | 23.3 * | 1.4 * | 9.96 * |
| Catalase activity (CAT) | 41.04 * | 16.07 * | 3.24 * |
| Biological index of fertility (BIF) | 87.41 * | 16.66 * | 22.0 * |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Niewiadomska, A.; Majchrzak, L.; Borowiak, K.; Wolna-Maruwka, A.; Waraczewska, Z.; Budka, A.; Gaj, R. The Influence of Tillage and Cover Cropping on Soil Microbial Parameters and Spring Wheat Physiology. Agronomy 2020, 10, 200. https://doi.org/10.3390/agronomy10020200
Niewiadomska A, Majchrzak L, Borowiak K, Wolna-Maruwka A, Waraczewska Z, Budka A, Gaj R. The Influence of Tillage and Cover Cropping on Soil Microbial Parameters and Spring Wheat Physiology. Agronomy. 2020; 10(2):200. https://doi.org/10.3390/agronomy10020200
Chicago/Turabian StyleNiewiadomska, Alicja, Leszek Majchrzak, Klaudia Borowiak, Agnieszka Wolna-Maruwka, Zyta Waraczewska, Anna Budka, and Renata Gaj. 2020. "The Influence of Tillage and Cover Cropping on Soil Microbial Parameters and Spring Wheat Physiology" Agronomy 10, no. 2: 200. https://doi.org/10.3390/agronomy10020200
APA StyleNiewiadomska, A., Majchrzak, L., Borowiak, K., Wolna-Maruwka, A., Waraczewska, Z., Budka, A., & Gaj, R. (2020). The Influence of Tillage and Cover Cropping on Soil Microbial Parameters and Spring Wheat Physiology. Agronomy, 10(2), 200. https://doi.org/10.3390/agronomy10020200