
FAM107A Inactivation Associated with Promoter Methylation Affects Prostate Cancer Progression through the FAK/PI3K/AKT Pathway


Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Clinical Specimens and Cell Culture
2.2. Western Blotting
2.3. Immunohistochemistry
2.4. Quantitative Real-Time PCR
2.5. Methylation-Specific PCR (MSP)
2.6. Cell Transfection
2.7. Cell Proliferation Assay Cells
2.8. Colony Assay
2.9. Transwell Invasion Analysis
2.10. Wound Healing Assay
2.11. Immunofluorescence
2.12. Flow Cytometry
2.13. Xenograft Assay
2.14. Statistical Analysis
3. Result
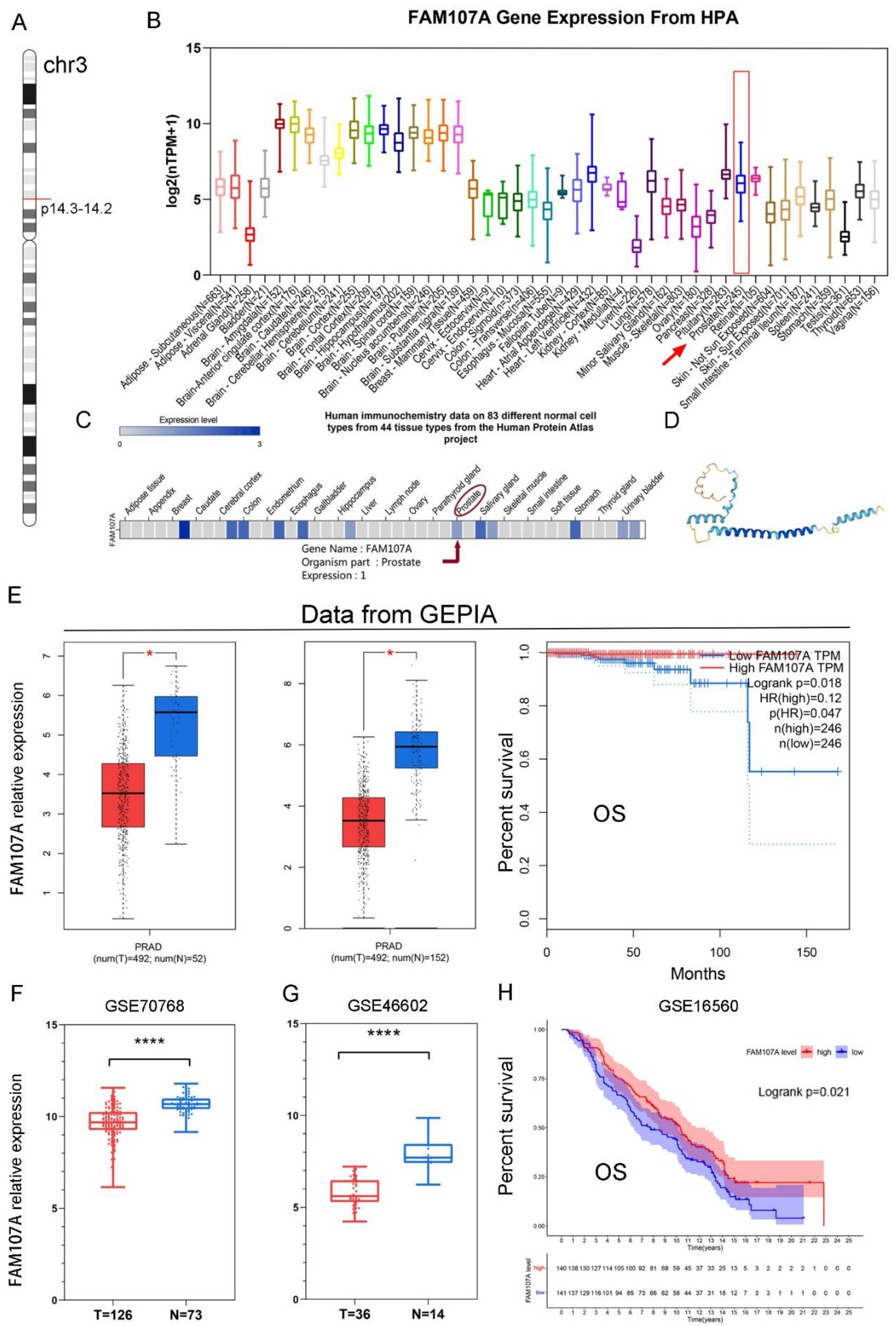
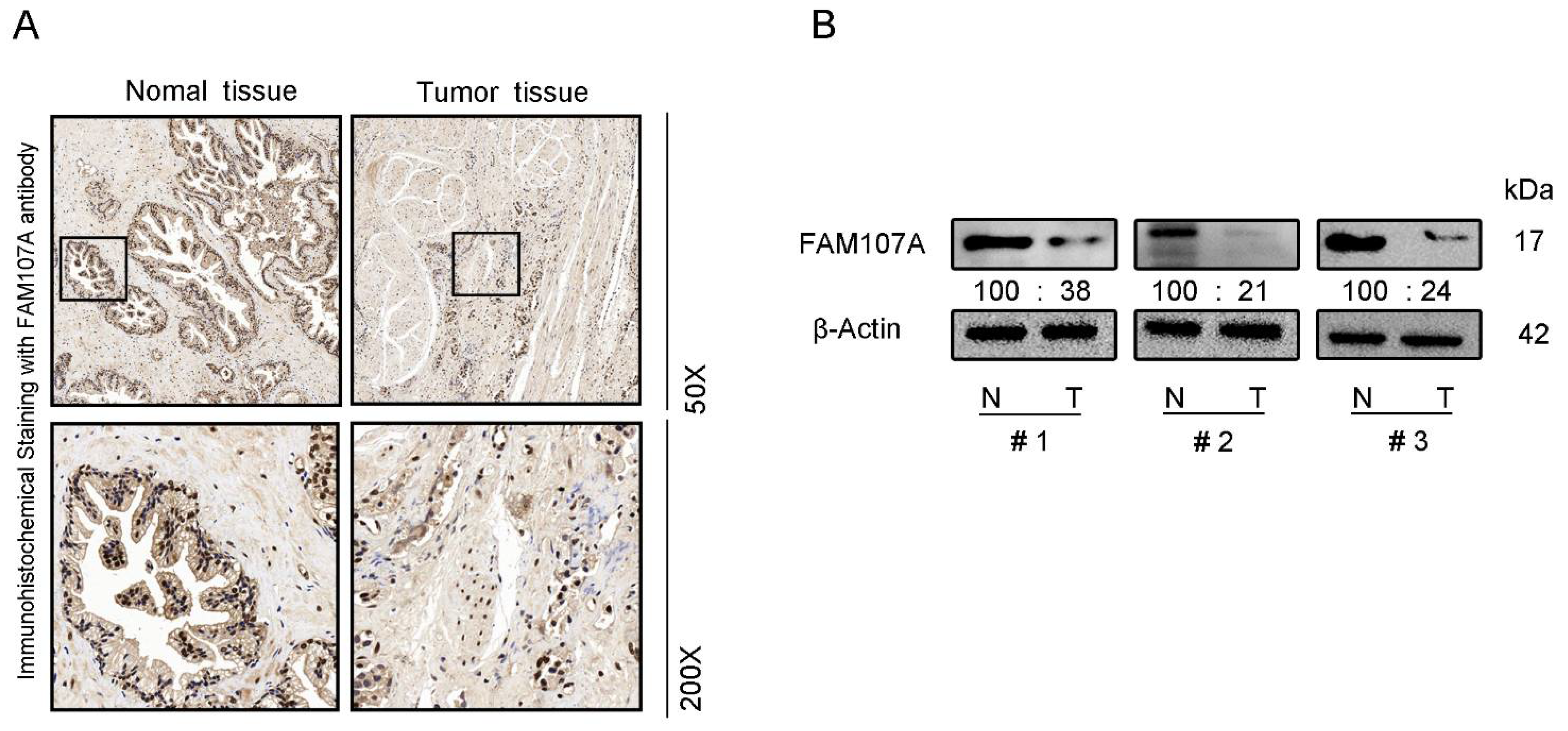
3.1. FAM107A Expression and Prognostic Analysis in Prostate Tissue
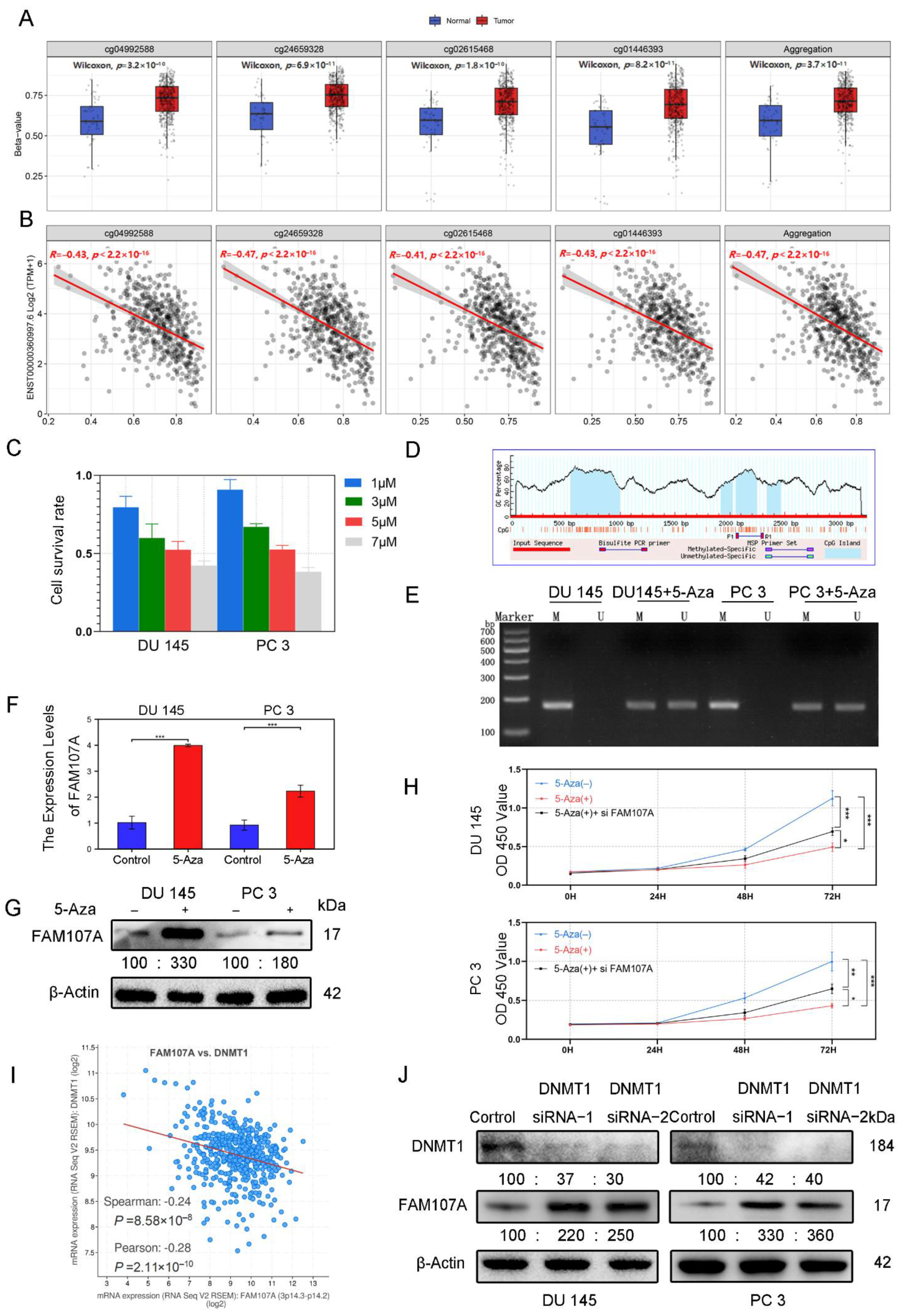
3.2. DNA Promoter Methylation Was Responsible for the Downregulation of FAM107A Expression
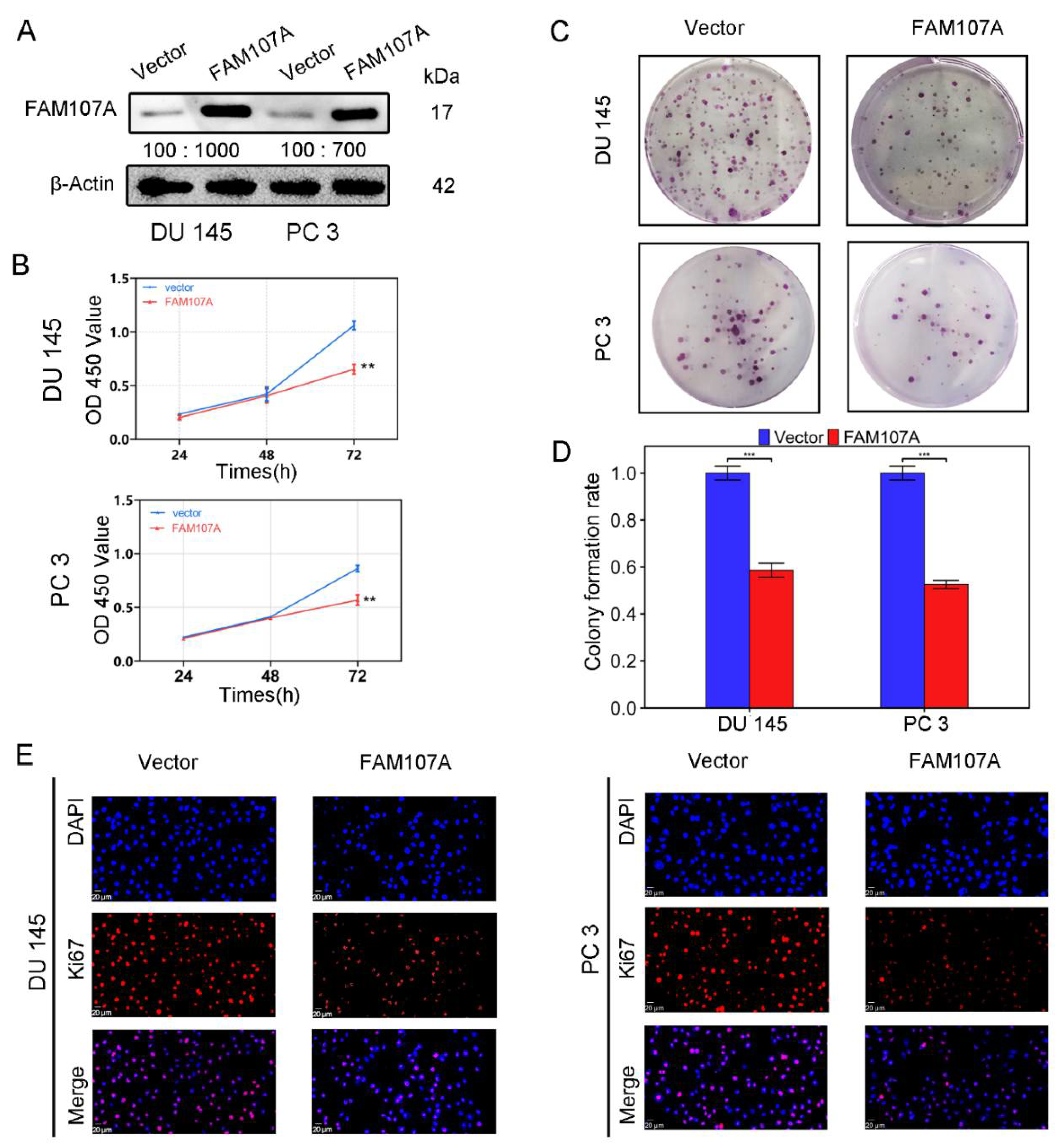
3.3. Overexpression of FAM107A Inhibited PCa Cell Growth and Invasion In Vitro
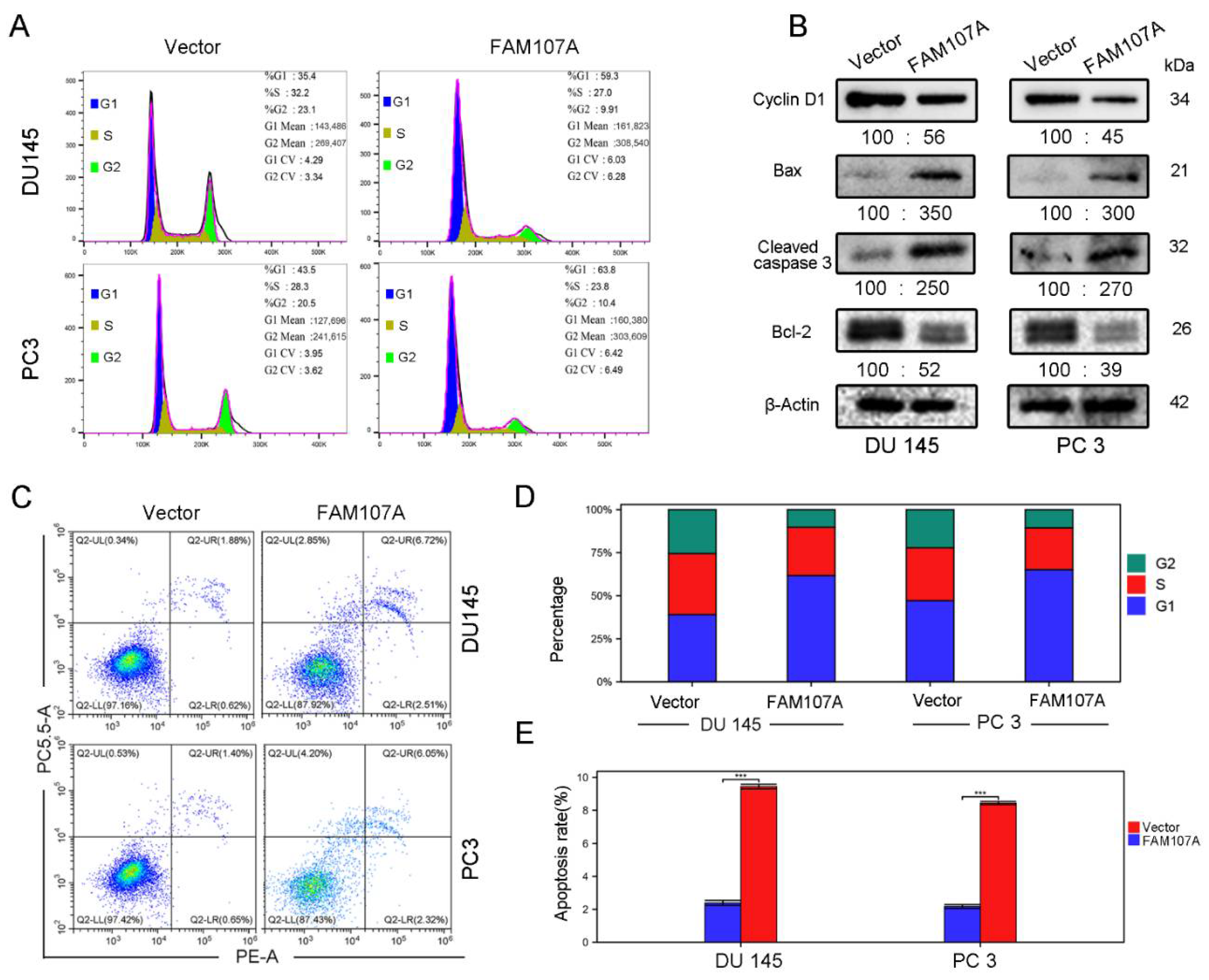
3.4. Regulation of the Cell Cycle and Apoptosis by FAM107A
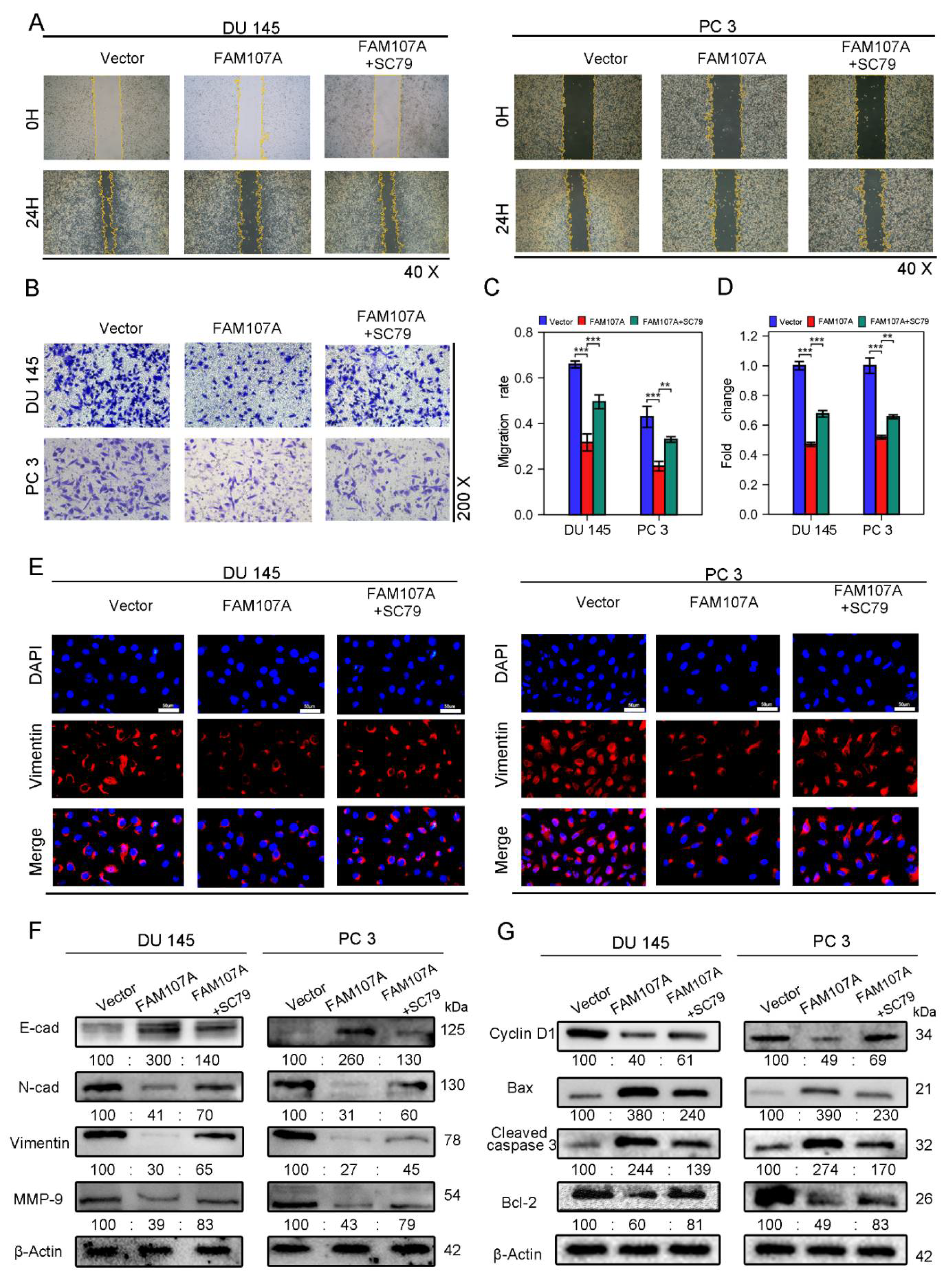
3.5. In PCa Cells, FAM107A Suppressed the FAK/PI3K/AKT Signaling Pathway by Downregulating FAK Expression
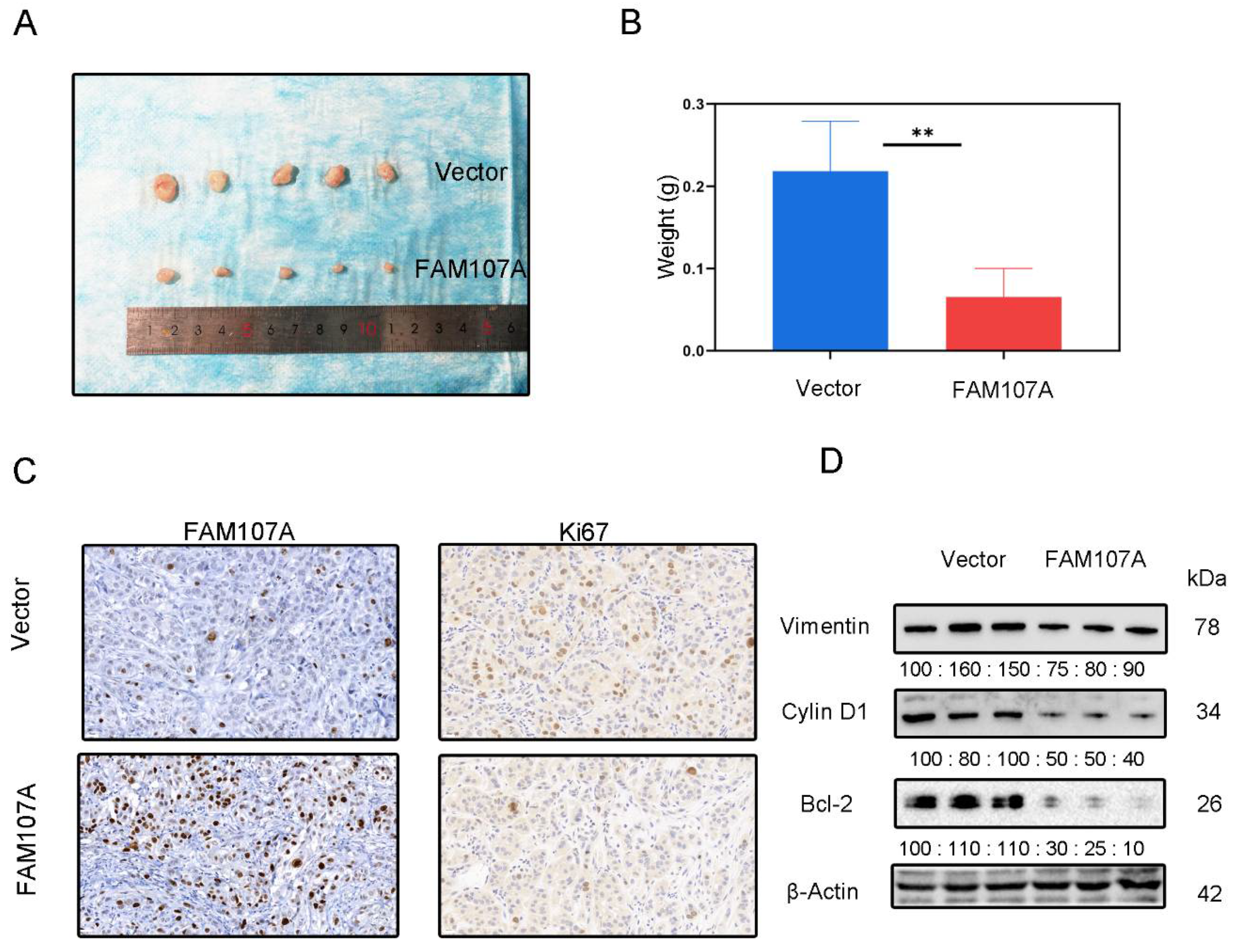
3.6. The Inhibitory Effect of FAM107A on PCa Cells In Vivo
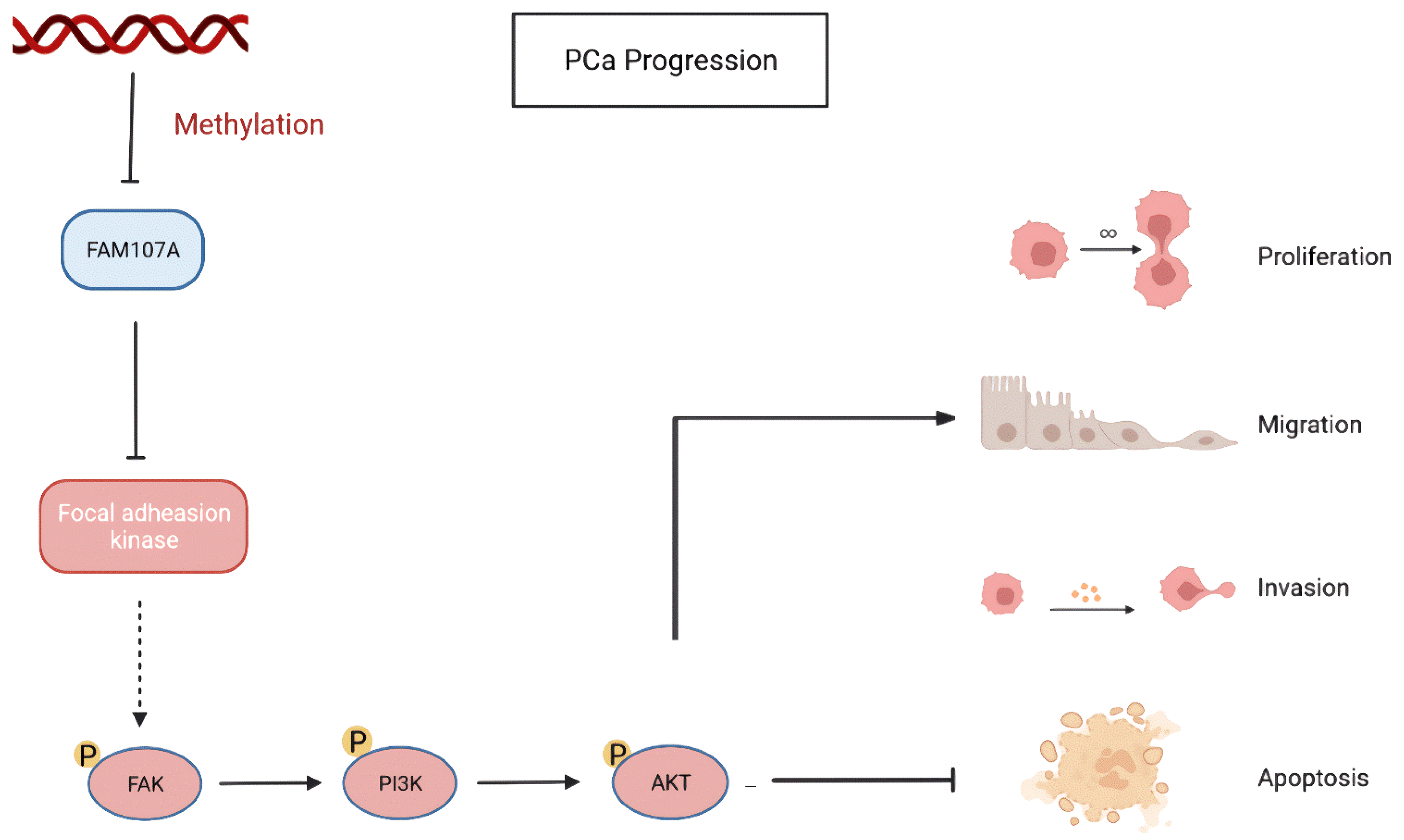
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Siegel, R.L.; Miller, K.D.; Fuchs, H.E.; Jemal, A. Cancer Statistics, 2021. CA Cancer J. Clin. 2021, 71, 7–33. [Google Scholar] [CrossRef] [PubMed]
- Chen, W.; Zheng, R.; Baade, P.D.; Zhang, S.; Zeng, H.; Bray, F.; Jemal, A.; Yu, X.Q.; He, J. Cancer statistics in China, 2015. CA Cancer J. Clin. 2016, 66, 115–132. [Google Scholar] [CrossRef] [PubMed]
- Gamat, M.; McNeel, D.G. Androgen deprivation and immunotherapy for the treatment of prostate cancer. Endocr Relat. Cancer 2017, 24, T297–T310. [Google Scholar] [CrossRef] [PubMed]
- Kok, K.; Naylor, S.L.; Buys, C.H. Deletions of the short arm of chromosome 3 in solid tumors and the search for suppressor genes. Adv. Cancer Res. 1997, 71, 27–92. [Google Scholar]
- Gao, L.; Wu, Z.X.; Assaraf, Y.G.; Chen, Z.S.; Wang, L. Overcoming anti-cancer drug resistance via restoration of tumor suppressor gene function. Drug. Resist Updat 2021, 57, 100770. [Google Scholar] [CrossRef]
- Wang, L.; Darling, J.; Zhang, J.S.; Liu, W.; Qian, J.; Bostwick, D.; Hartmann, L.; Jenkins, R.; Bardenhauer, W.; Schutte, J. Loss of expression of the DRR 1 gene at chromosomal segment 3p21.1 in renal cell carcinoma. Genes Chromosomes Cancer 2000, 27, 1–10. [Google Scholar] [CrossRef]
- Yamato, T.; Orikasa, K.; Fukushige, S.; Orikasa, S.; Horii, A. Isolation and characterization of the novel gene, TU3A, in a commonly deleted region on 3p14.3-->p14.2 in renal cell carcinoma. Cytogenet. Cell Genet. 1999, 87, 291–295. [Google Scholar] [CrossRef]
- Pollen, A.A.; Nowakowski, T.J.; Chen, J.; Retallack, H.; Sandoval-Espinosa, C.; Nicholas, C.R.; Shuga, J.; Liu, S.J.; Oldham, M.C.; Diaz, A. Molecular identity of human outer radial glia during cortical development. Cell 2015, 24, 55–67. [Google Scholar] [CrossRef] [PubMed]
- Manigandan, S.; Mukherjee, S.; Yun, J.W. Loss of family with sequence similarity 107, member A (FAM107A) induces browning in 3T3-L1 adipocytes. Arch Biochem. Biophys. 2021, 704, 108885. [Google Scholar] [CrossRef] [PubMed]
- Kholodnyuk, I.D.; Kozireva, S.; Kost-Alimova, M.; Kashuba, V.; Klein, G.; Imreh, S. Down regulation of 3p genes, LTF, SLC38A3 and DRR1, upon growth of human chromosome 3-mouse fibrosarcoma hybrids in severe combined immunodeficiency mice. Int. J. Cancer 2006, 119, 99–107. [Google Scholar] [CrossRef]
- Ou, L.; Wei, Z.; Xu, J.; Li, W.; Zhou, Y.; Wang, Y.; Shi, B. FAM107A as a Tumor Suppressor in Bladder Cancer Inhibits Cell Proliferation, Migration, and Invasion. Ann. Clin. Lab. Sci. 2022, 52, 260–268. [Google Scholar]
- Mu, P.; Akashi, T.; Lu, F.; Kishida, S.; Kadomatsu, K. A novel nuclear complex of DRR1, F-actin and COMMD1 involved in NF-κB degradation and cell growth suppression in neuroblastoma. Oncogene 2017, 36, 5745–5756. [Google Scholar] [CrossRef]
- Kiwerska, K.; Szaumkessel, M.; Paczkowska, J.; Bodnar, M.; Byzia, E.; Kowal, E.; Kostrzewska-Poczekaj, M.; Janiszewska, J.; Bednarek, K.; Jarmuż-Szymczak, M. Combined deletion and DNA methylation result in silencing of FAM107A gene in laryngeal tumors. Sci. Rep. 2017, 7, 5386. [Google Scholar] [CrossRef]
- Lao, T.D.; Orcid, I.D.; Nguyen, T.N.; Le, T.A.H.; Orcid, I.D. Promoter Hypermethylation of Tumor Suppressor Genes Located on Short Arm of the Chromosome 3 as Potential Biomarker for the Diagnosis of Nasopharyngeal Carcinoma. Diagnostics 2021, 11, 1404. [Google Scholar] [CrossRef]
- Cody, N.A.; Ouellet, V.; Manderson, E.N.; Quinn, M.C.; Filali-Mouhim, A.; Tellis, P.; Zietarska, M.; Provencher, D.M.; Mes-Masson, A.M.; Chevrette, M. Transfer of chromosome 3 fragments suppresses tumorigenicity of an ovarian cancer line monoallelic for chromosome 3p. Oncogene 2007, 26, 618–632. [Google Scholar] [CrossRef]
- Zabarovsky, E.R.; Lerman, M.I.; Minna, J.D. Tumor suppressor genes on chromosome 3p involved in the pathogenesis of lung and and other cancers. Oncogene 2002, 21, 6915–6935. [Google Scholar] [CrossRef]
- Shi, J.; Hu, J.; Zhou, Q.; Du, Y.; Jiang, C. PEpiD: A prostate epigenetic database in mammals. PLoS ONE 2013, 8, e64289. [Google Scholar] [CrossRef]
- Cowell, J.K. Tumour suppressor genes. Ann. Oncol. 1992, 3, 693–698. [Google Scholar] [CrossRef]
- Liang, G.; Weisenberger, D.J. DNA methylation aberrancies as a guide for surveillance and treatment of human cancers. Epigenetics 2017, 12, 416–432. [Google Scholar] [CrossRef]
- Lyko, F. The DNA methyltransferase family: A versatile toolkit for epigenetic regulation. Nat. Rev. Genet. 2018, 19, 81–92. [Google Scholar] [CrossRef]
- Zhang, W.; Xu, J. DNA methyltransferases and their roles in tumorigenesis. Biomark Res. 2017, 5, 1. [Google Scholar] [CrossRef] [PubMed]
- Witte, T.; Plass, C.; Gerhauser, C. Pan-cancer patterns of DNA methylation. Genome. Med. 2014, 6, 66. [Google Scholar] [CrossRef] [PubMed]
- Cheng, H.; Tang, S.; Lian, X.; Meng, H.; Gu, X.; Jiang, J.; Li, X. The Differential Antitumor Activity of 5-Aza-2′-deoxycytidine in Prostate Cancer. J. Cancer 2021, 12, 5593–5604. [Google Scholar] [CrossRef] [PubMed]
- Vanaja, D.K.; Ballman, K.V.; Morlan, B.W.; Cheville, J.C.; Neumann, R.M.; Lieber, M.M.; Tindall, D.J.; Young, C.Y. PDLIM4 repression by hypermethylation as a potential biomarker for prostate cancer. Clin. Cancer Res. 2006, 12, 1128–1136. [Google Scholar] [CrossRef]
- Sun, W.; Yang, J. Functional mechanisms for human tumor suppressors. J. Cancer 2010, 1, 136–140. [Google Scholar] [CrossRef]
- Ma, Y.S.; Wu, Z.J.; Bai, R.Z.; Dong, H.; Xie, B.X.; Wu, X.H.; Hang, X.S.; Liu, A.N.; Jiang, X.H.; Wang, G.R. DRR1 promotes glioblastoma cell invasion and epithelial-mesenchymal transition via regulating AKT activation. Cancer Lett. 2018, 423, 86–94. [Google Scholar] [CrossRef]
- Ross-Adams, H.; Lamb, A.D.; Dunning, M.J.; Halim, S.; Lindberg, J.; Massie, C.M.; Egevad, L.A.; Russell, R.; Ramos-Montoya, A.; Vowler, S.L. Integration of copy number and transcriptomics provides risk stratification in prostate cancer: A discovery and validation cohort study. Ebio Med. 2015, 2, 1133–1144. [Google Scholar] [CrossRef]
- Mortensen, M.M.; Høyer, S.; Lynnerup, A.S.; Ørntoft, T.F.; Sørensen, K.D.; Borre, M.; Dyrskjøt, L. Expression profiling of prostate cancer tissue delineates genes associated with recurrence after prostatectomy. Sci. Rep. 2015, 5, 16018. [Google Scholar] [CrossRef]
- Sboner, A.; Demichelis, F.; Calza, S.; Pawitan, Y.; Setlur, S.R.; Hoshida, Y.; Perner, S.; Adami, H.O.; Fall, K.; Mucci, L.A.; et al. Molecular sampling of prostate cancer: A dilemma for predicting disease progression. BMC Med. Genom. 2010, 3, 8. [Google Scholar] [CrossRef]
- Ma, Y.F.; Li, G.D.; Sun, X.; Li, X.X.; Gao, Y.; Gao, C.; Cao, K.X.; Yang, W.G.; Yu, M.W.; Wang, X.M. Identification of FAM107A as a potential biomarker and therapeutic target for prostate carcinoma. Am. J. Transl. Res. 2021, 13, 10163–10177. [Google Scholar]
- Brabletz, T.; Kalluri, R.; Nieto, M.A.; Weinberg, R.A. EMT in cancer. Nat. Rev. Cancer 2018, 18, 128–134. [Google Scholar] [CrossRef] [PubMed]
- Savagner, P. The epithelial-mesenchymal transition (EMT) phenomenon. Ann. Oncol. 2010, 21 (Suppl. S7), vii89–vii92. [Google Scholar] [CrossRef] [PubMed]
- Lin, C.Y.; Tsai, P.H.; Kandaswami, C.C.; Lee, P.P.; Huang, C.J.; Hwang, J.J.; Lee, M.T. Matrix metalloproteinase-9 cooperates with transcription factor Snail to induce epithelial-mesenchymal transition. Cancer Sci. 2011, 102, 815–827. [Google Scholar] [CrossRef] [PubMed]
- Brullo, C.; Tasso, B. New Insights on Fak and Fak Inhibitors. Curr. Med. Chem. 2021, 28, 3318–3338. [Google Scholar] [CrossRef]
- Wang, Q.; Liu, Z.; Zhai, G.; Yu, X.; Ke, S.; Shao, H.; Guo, J. Overexpression of GATA5 Inhibits Prostate Cancer Progression by Regulating PLAGL2 via the FAK/PI3K/AKT Pathway. Cancers 2022, 14, 2074. [Google Scholar] [CrossRef]
- Benelli, R.; Monteghirfo, S.; Venè, R.; Tosetti, F.; Ferrari, N. The chemopreventive retinoid 4HPR impairs prostate cancer cell migration and invasion by interfering with FAK/AKT/GSK3beta pathway and beta-catenin stability. Mol. Cancer 2010, 9, 142. [Google Scholar] [CrossRef]
- Gao, N.; Zhang, Z.; Jiang, B.H.; Shi, X. Role of PI3K/AKT/mTOR signaling in the cell cycle progression of human prostate cancer. Biochem. Biophys. Res. Commun. 2003, 310, 1124–1132. [Google Scholar] [CrossRef]
- Kim, S.M.; Park, J.H.; Kim, K.D.; Nam, D.; Shim, B.S.; Kim, S.H.; Ahn, K.S.; Choi, S.H.; Ahn, K.S. Brassinin induces apoptosis in PC-3 human prostate cancer cells through the suppression of PI3K/Akt/mTOR/S6K1 signaling cascades. Phytother Res. 2014, 28, 423–431. [Google Scholar] [CrossRef]
- Vo, B.T.; Morton, D., Jr.; Komaragiri, S.; Millena, A.C.; Leath, C.; Khan, S.A. TGF-β effects on prostate cancer cell migration and invasion are mediated by PGE2 through activation of PI3K/AKT/mTOR pathway. Endocrinology 2013, 154, 1768–1779. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Antibody | Specificity | WB | IF | IHC | Company |
|---|---|---|---|---|---|
| FAM107A | Rabbit | 1:500 | - | 1:200 | Proteintech |
| DNMT1 | Rabbit | 1:500 | - | - | Proteintech |
| E-cadherin | Rabbit | 1:1000 | 1:100 | - | Proteintech |
| N-cadherin | Rabbit | 1:1000 | - | - | Proteintech |
| Vimentin | Rabbit | 1:1000 | 1:100 | - | Proteintech |
| MMP-9 | Rabbit | 1:1000 | - | - | Proteintech |
| FAK | Rabbit | 1:1000 | - | - | ABclonal |
| p-FAK | Rabbit | 1:1000 | - | - | ABclonal |
| PI3K | Rabbit | 1:1000 | - | - | ABclonal |
| p-PI3K | Rabbit | 1:1000 | - | - | ABclonal |
| AKT | Mouse | 1:1000 | - | - | ABclonal |
| p-AKT | Mouse | 1:1000 | - | - | ABclonal |
| Cyclin D1 | Rabbit | 1:1000 | - | - | Proteintech |
| Bax | Rabbit | 1:1000 | - | - | CST |
| Bcl-2 | Rabbit | 1:1000 | - | - | CST |
| Cleaved caspase-3 | Rabbit | 1:1000 | - | - | CST |
| Ki67 | Rabbit | - | 1:100 | 1:200 | Abcam |
| β-actin | Rabbit | 1:1000 | - | - | Proteintech |
| Primer Name | Sequence (5′-3′) | Length | Tm | GC Content (%) | Number of Bases |
|---|---|---|---|---|---|
| H-FAM107A (M)-F | TTTGGGATTTGGGGTCGC | 181 | 61.2 | 55.6 | 18 |
| H-FAM107A (M)-R | AACACAACCCGATAAAACCCG | 61.5 | 47.6 | 21 | |
| H-FAM107A (U)-F | GGTTTGGGATTTGGGGTTGT | 60.9 | 50 | 20 | |
| H-FAM107A (U)-R | ACAACACAACCCAATAAAACCCA | 185 | 61.1 | 39.1 | 23 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ke, S.; Liu, Z.; Wang, Q.; Zhai, G.; Shao, H.; Yu, X.; Guo, J. FAM107A Inactivation Associated with Promoter Methylation Affects Prostate Cancer Progression through the FAK/PI3K/AKT Pathway. Cancers 2022, 14, 3915. https://doi.org/10.3390/cancers14163915
Ke S, Liu Z, Wang Q, Zhai G, Shao H, Yu X, Guo J. FAM107A Inactivation Associated with Promoter Methylation Affects Prostate Cancer Progression through the FAK/PI3K/AKT Pathway. Cancers. 2022; 14(16):3915. https://doi.org/10.3390/cancers14163915
Chicago/Turabian StyleKe, Shuai, Zelin Liu, Qinghua Wang, Guanzhong Zhai, Haoren Shao, Xi Yu, and Jia Guo. 2022. "FAM107A Inactivation Associated with Promoter Methylation Affects Prostate Cancer Progression through the FAK/PI3K/AKT Pathway" Cancers 14, no. 16: 3915. https://doi.org/10.3390/cancers14163915
APA StyleKe, S., Liu, Z., Wang, Q., Zhai, G., Shao, H., Yu, X., & Guo, J. (2022). FAM107A Inactivation Associated with Promoter Methylation Affects Prostate Cancer Progression through the FAK/PI3K/AKT Pathway. Cancers, 14(16), 3915. https://doi.org/10.3390/cancers14163915