
Cancer-Associated Fibroblast-Derived IL-6 Determines Unfavorable Prognosis in Cholangiocarcinoma by Affecting Autophagy-Associated Chemoresponse


 , ,
, ,  and
and 
Abstract
Simple Summary
Abstract
1. Introduction
2. Results
2.1. Clinicopathological Data of CCA Patients
2.2. IL-6 Expression in Fibrotic Stroma Associates with Poor Prognosis in CCA Patients
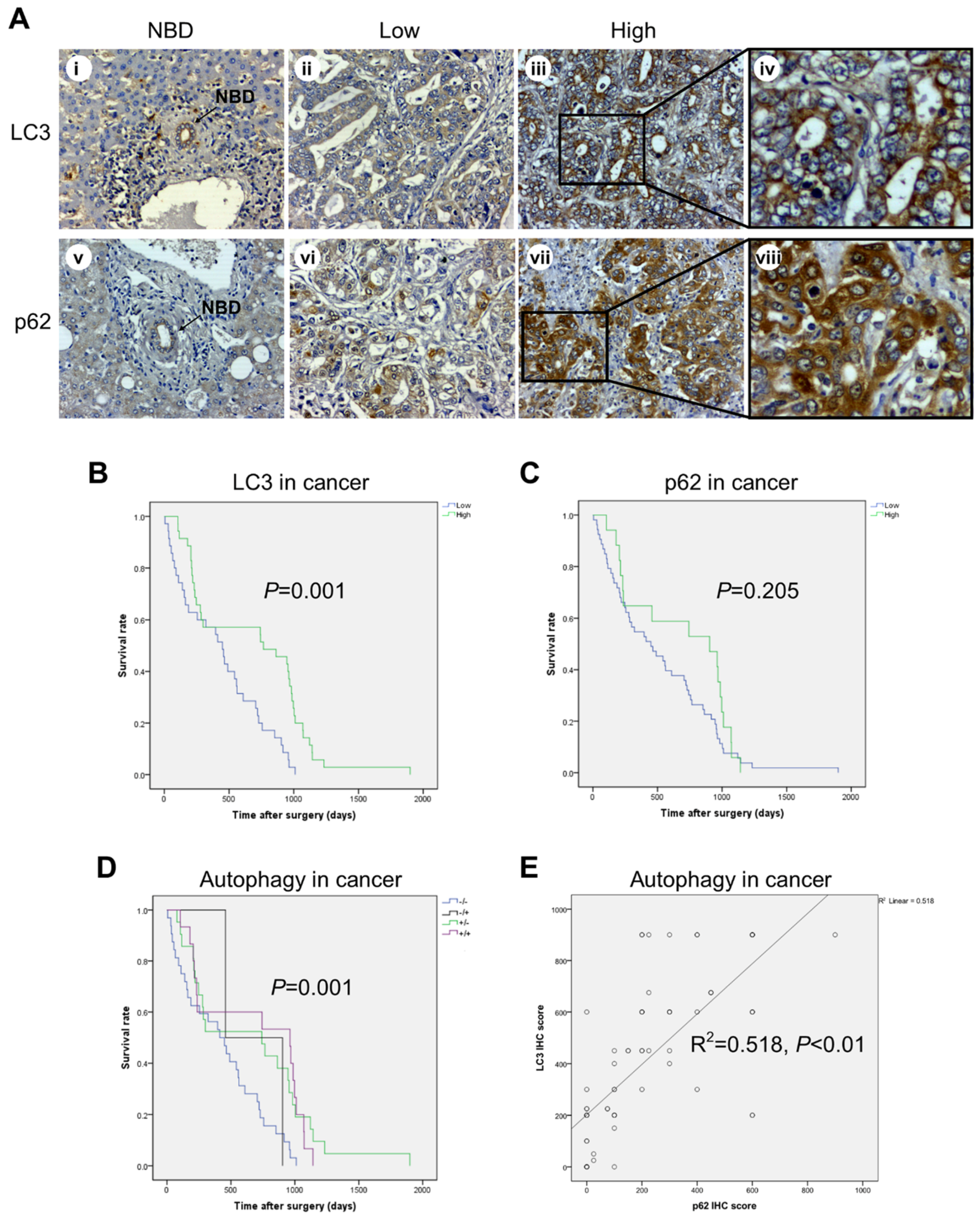
2.3. High Expression of LC3 and Low Expression of p62 in CCA Cells Correlate with Better Prognosis
2.4. Correlation between IL-6, LC3, and p62 Expression and Clinicopathologic Features of CCA Patients
2.5. The Role of Adjuvant Chemotherapy and of Inflammatory and Autophagy Marker Expression in Patient Survival
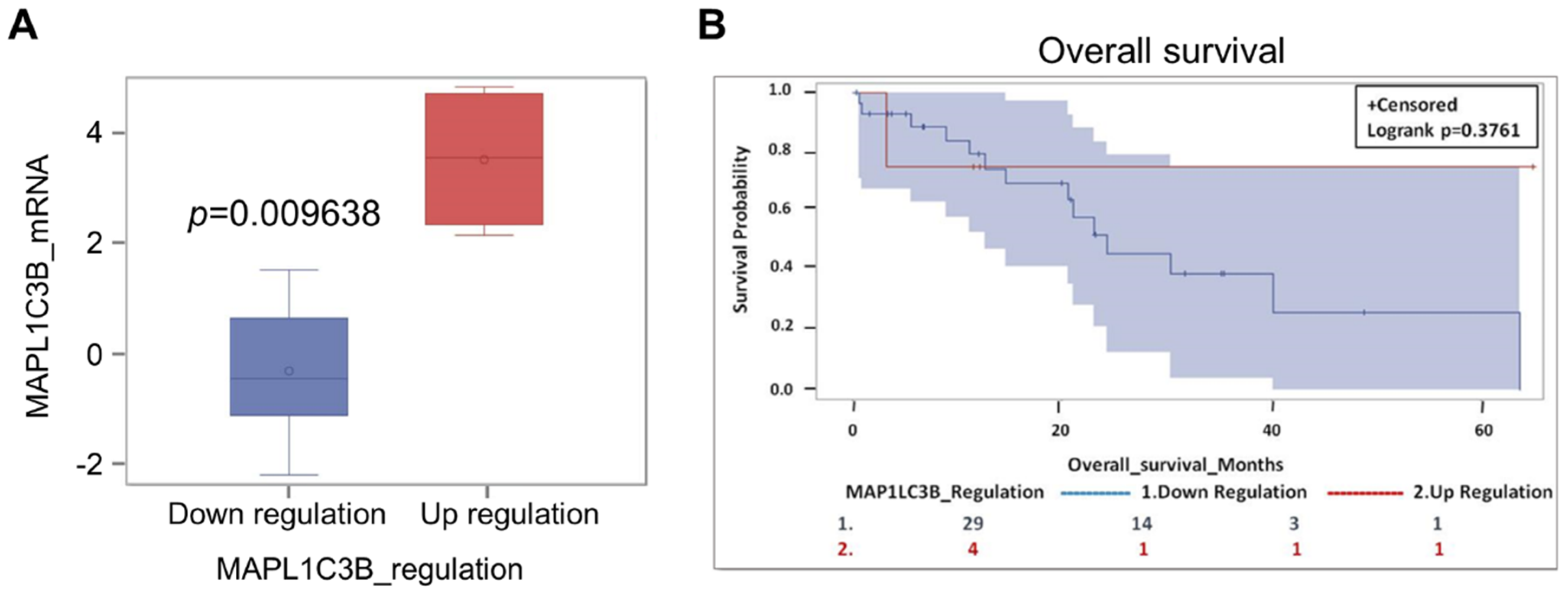
2.6. Expression of Autophagy Markers and Clinical Outcome in CCA-Bearing Patients from the TCGA Database
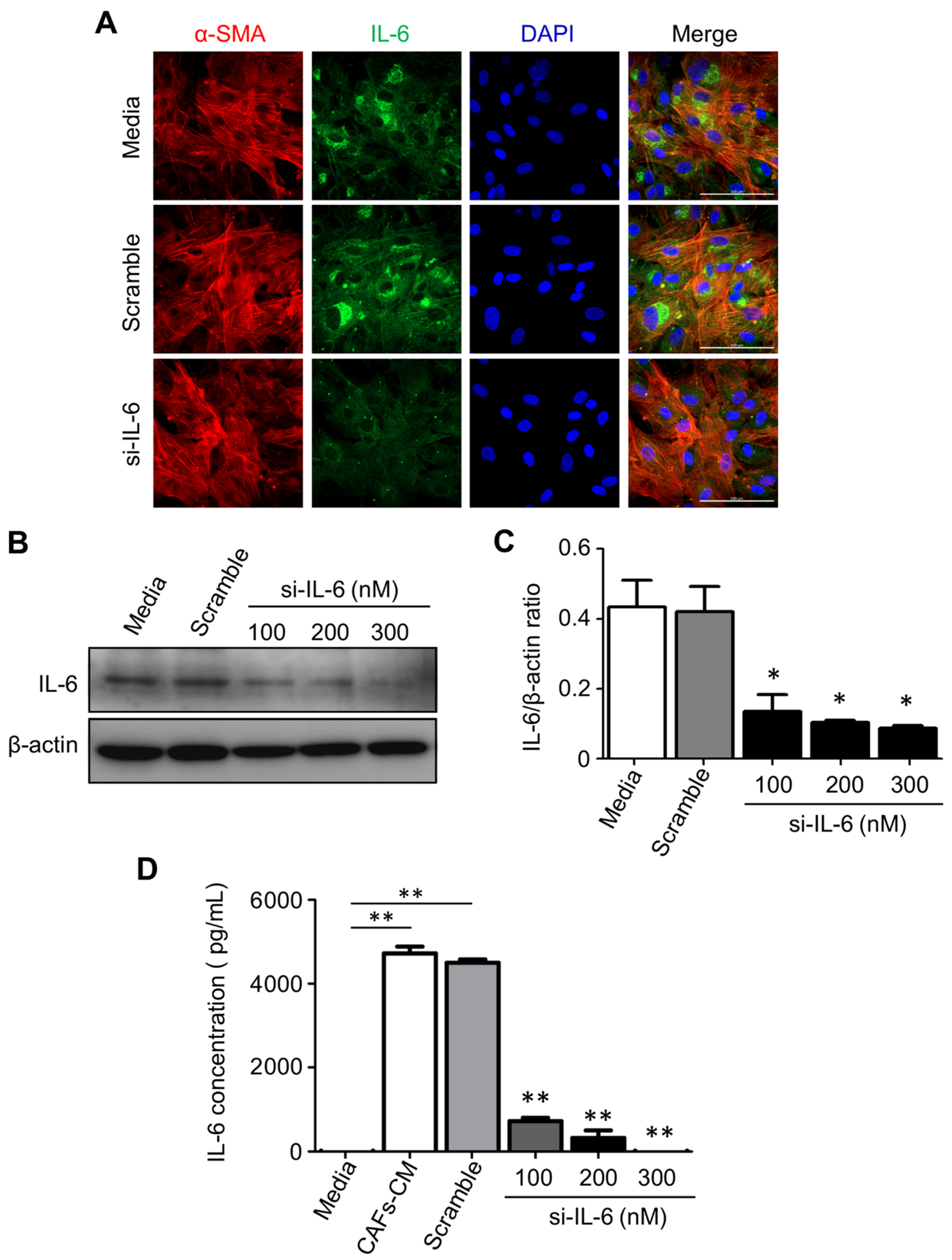
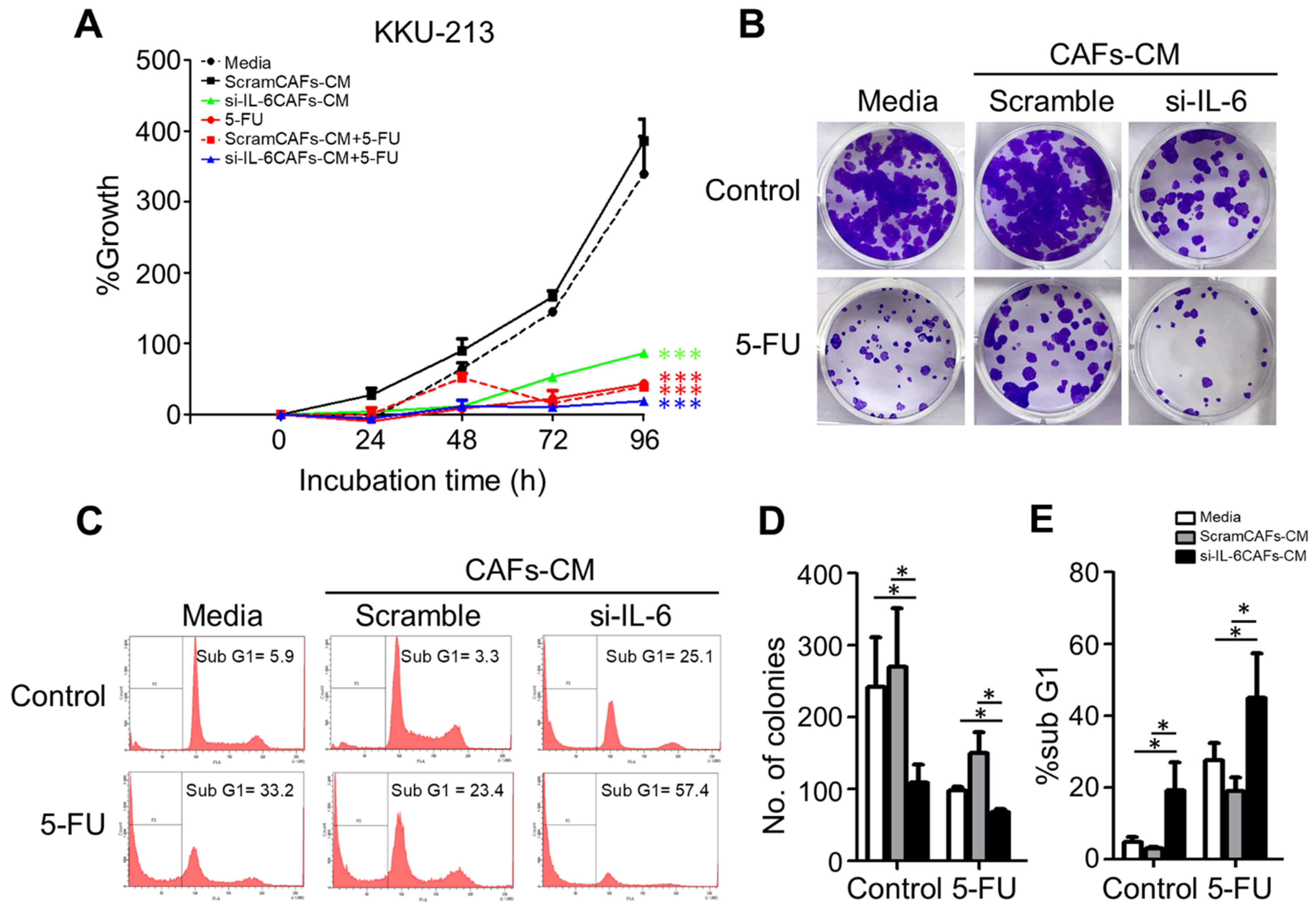
2.7. IL-6 Secreted by CAFs Inhibits Autophagy and Reduces the Chemosensitivity of CCA Cells
2.8. Impaired Autophagy by Stromal Inflammation in an Experimental Animal Model of Cholangiocarcinoma
3. Discussion
4. Materials and Methods
4.1. Patients, Samples, and Ethical Issues
4.2. Cell Lines and Primary Cancer-Associated Fibroblasts
4.3. Antibodies and Reagents
4.4. Cytotoxicity Assay
4.5. Cell Viability
4.6. Immunohistochemical Analysis
4.7. Western Blot Analysis
4.8. Immunofluorescence Staining
4.9. Transient Knockdown of Small Interfering RNA for IL-6 into CAFs
4.10. The Cancer Genome Atlas (TCGA)
4.11. Animal Model, Treatments, and Histological Analyses
4.12. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Abbreviations
| CAFs | cancer-associated fibroblasts |
| CCA | cholangiocarcinoma |
| CoM | control medium |
| CI | confidence interval |
| HR | Hazard ratio |
| IL-6 | interleukin-6 |
| NDMA | N-nitrosodimethylamine |
| Ov | Opisthorchis viverrini |
| TCGA | the cancer genome altos |
| TMA | tumor microarray |
References
- Blechacz, B.; Gores, G.J. Cholangiocarcinoma: Advances in pathogenesis, diagnosis, and treatment. Hepatology 2008, 48, 308–321. [Google Scholar] [CrossRef] [PubMed]
- Doherty, B.; Nambudiri, V.E.; Palmer, W.C. Update on the diagnosis and treatment of cholangiocarcinoma. Curr. Gastroenterol. Rep. 2017, 19, 2. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.F.; Dong, M.; Pan, Y.H.; Chen, J.N.; Huang, X.Q.; Jin, Y.; Shao, C.K. Expression pattern of cancer-associated fibroblast and its clinical relevance in intrahepatic cholangiocarcinoma. Hum. Pathol. 2017, 65, 92–100. [Google Scholar] [CrossRef] [PubMed]
- Prueksapanich, P.; Piyachaturawat, P.; Aumpansub, P.; Ridtitid, W.; Chaiteerakij, R.; Rerknimitr, R. Liver fluke-associated biliary tract cancer. Gut Liver 2018, 12, 236–245. [Google Scholar] [CrossRef]
- Franken, L.C.; Schreuder, A.M.; Roos, E.; Dieren, S.V.; Busch, O.R.; Besselink, M.G.; Gulik, T.M. Morbidity and mortality after major liver resection in patients with perihilar cholangiocarcinoma: A systematic review and meta-analysis. Surgery 2019, 165, 918–928. [Google Scholar] [CrossRef] [PubMed]
- Moris, D.; Kostakis, I.D.; Machairas, N.; Prodromidou, A.; Tsilimigras, D.I.; Ravindra, K.V.; Sudan, D.L.; Knechtle, S.J.; Barbas, A.S. Comparison between liver transplantation and resection for hilar cholangiocarcinoma: A systematic review and meta-analysis. PLoS ONE 2019, 14, e0220527. [Google Scholar] [CrossRef]
- Kennedy, L.; Hargrove, L.; Demieville, J.; Francis, N.; Seils, R.; Villamaria, S.; Villamaria, S.; Francis, H. Recent advances in understanding cholangiocarcinoma. F1000Research 2017, 6, 1818. [Google Scholar] [CrossRef]
- Valle, J.; Wasan, H.; Palmer, D.H.; Cunningham, D.; Anthoney, A.; Maraveyas, A.; Madhusudan, S.; Iveson, T.; Hughes, S.; Pereira, S.P.; et al. Cisplatin plus gemcitabine versus gemcitabine for biliary tract cancer. N. Eng. J. Med. 2010, 362, 1273–1281. [Google Scholar] [CrossRef]
- Luvira, V.; Eurboonyanun, C.; Bhudhisawasdi, V.; Pugkhem, A.; Pairojkul, C.; Luvira, V.; Sathitkarnmanee, E.; Somsap, K.; Kamsa-ard, S. Patterns of recurrence after resection of mass-forming type intrahepatic Cholangiocarcinomas. Asian Pac. J. Cancer Prev. 2017, 17, 4735–4739. [Google Scholar]
- Watanabe, A.; Suzuki, H.; Yokobori, T.; Tsukagoshi, M.; Altan, B.; Kubo, N.; Suzuki, S.; Araki, K.; Wada, S.; Kashiwabara, K.; et al. Stathmin1 regulates p27 expression, proliferation and drug resistance, resulting in poor clinical prognosis in cholangiocarcinoma. Cancer Sci. 2014, 105, 690–696. [Google Scholar] [CrossRef]
- Wirasorn, K.; Ngamprasertchai, T.; Khuntikeo, N.; Pakkhem, A.; Ungarereevittaya, P.; Chindaprasirt, J.; Sookprasert, A. Adjuvant chemotherapy in resectable cholangiocarcinoma patients. J. Gastroenterol. Hepatol. 2013, 28, 1885–1891. [Google Scholar] [CrossRef]
- Wardell, C.P.; Fujita, M.; Yamada, T.; Simbolo, M.; Fassan, M.; Karlic, R.; Polak, P.; Kim, J.; Hatanaka, Y.; Maejima, K. Genomic characterization of biliary tract cancers identifies driver genes and predisposing mutations. J. Hepatol. 2018, 68, 959–969. [Google Scholar] [CrossRef] [PubMed]
- Zhong, W.; Dai, L.; Liu, J.; Zhou, S. Cholangiocarcinoma‑associated genes identified by integrative analysis of gene expression data. Mol. Med. Rep. 2018, 17, 5744–5753. [Google Scholar] [CrossRef]
- Zhang, C.; Zhang, B.; Meng, D.; Ge, C. Comprehensive analysis of DNA methylation and gene expression profiles in cholangiocarcinoma. Cancer Cell Int. 2019, 19, 352. [Google Scholar] [CrossRef]
- Lowery, M.A.; Ptashkin, R.; Jordan, E.; Berger, M.F.; Zehir, A.; Capanu, M.; Kemeny, N.E.; O’Reilly, E.M.; El-Dika, I.; Jarnagin, W.R. Comprehensive molecular profiling of intrahepatic and extrahepatic cholangiocarcinomas: Potential targets for intervention. Clin. Cancer Res. 2018, 24, 4154–4161. [Google Scholar] [CrossRef] [PubMed]
- Ueasilamongkol, P.; Khamphaya, T.; Guerra, M.T.; Rodrigues, M.A.; Gomes, D.A.; Kong, Y.; Wei, W.; Jai, D.; Trampert, D.C.; Ananthanarayanan, M. Type 3 inositol 1, 4, 5-trisphosphate receptor is increased and enhances malignant properties in cholangiocarcinoma. Hepatology 2020, 71, 583–599. [Google Scholar] [CrossRef] [PubMed]
- Sittithumcharee, G.; Suppramote, O.; Vaeteewoottacharn, K.; Sirisuksakun, C.; Jamnongsong, S.; Laphanuwat, P.; Suntiparpluacha, M.; Matha, A.; Chuson, P.; Buraphat, P.; et al. Dependency of Cholangiocarcinoma on Cyclin D–Dependent Kinase Activity. Hepatology 2019, 70, 1614–1630. [Google Scholar] [CrossRef]
- Ishida, K.; Osakabe, M.; Eizuka, M.; Tai, S.; Sugimoto, R.; Fujita, Y.; Katagiri, H.; Takahara, T.; Uesugi, N.; Nitta, H.; et al. The expression of gastrointestinal differentiation markers in extrahepatic cholangiocarcinoma: Clinicopathological significance based on tumor location. Hum. Pathol. 2019, 92, 91–100. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Li, J.; Xia, Y.; Gong, R.; Wang, K.; Yan, Z.; Wan, X.; Liu, G.; Wu, D.; Shi, L.; et al. Prognostic nomogram for intrahepatic cholangiocarcinoma after partial hepatectomy. J. Clin. Oncol. 2013, 31, 1188–1195. [Google Scholar] [CrossRef]
- Sasaki, M.; Matsubara, T.; Kakuda, Y.; Sato, Y.; Nakanuma, Y. Immunostaining for polycomb group protein EZH2 and senescent marker p16INK4a may be useful to differentiate cholangiolocellular carcinoma from ductular reaction and bile duct adenoma. Am. J. Surg. Pathol. 2014, 38, 364–369. [Google Scholar] [CrossRef]
- Sohal, D.P.; Shrotriya, S.; Abazeed, M.; Cruise, M.; Khorana, A. Molecular characteristics of biliary tract cancer. Crit. Rev. Oncol Hematol. 2016, 107, 111–118. [Google Scholar] [CrossRef]
- Simile, M.M.; Bagella, P.; Vidili, G.; Spanu, A.; Manetti, R.; Seddaiu, M.A.; Babudieri, S.; Madeddu, G.; Serra, P.A.; Altana, M.; et al. Targeted therapies in cholangiocarcinoma: Emerging evidence from clinical trials. Medicina 2019, 55, 42. [Google Scholar] [CrossRef]
- Mertens, J.C.; Fingas, C.D.; Christensen, J.D.; Smoot, R.L.; Bronk, S.F.; Werneburg, N.W.; Gustafson, M.P.; Dietz, A.B.; Roberts, L.R.; Sirica, A.E.; et al. Therapeutic effects of deleting cancer-associated fibroblasts in cholangiocarcinoma. Cancer Res. 2013, 73, 897–907. [Google Scholar] [CrossRef]
- Wang, T.T.; Cao, Q.H.; Chen, M.Y.; Xia, Q.; Fan, X.J.; Ma, X.K.; Lin, Q.; Jia, C.C.; Dong, M.; Ruan, D.Y.; et al. Beclin 1 deficiency correlated with lymph node metastasis, predicts a distinct outcome in intrahepatic and extrahepatic cholangiocarcinoma. PLoS ONE 2013, 8, e80317. [Google Scholar] [CrossRef] [PubMed]
- Sasaki, M.; Nitta, T.; Sato, Y.; Nakanuma, Y. Autophagy may occur at an early stage of cholangiocarcinogenesis via biliary intraepithelial neoplasia. Hum. Pathol. 2015, 46, 202–209. [Google Scholar] [CrossRef] [PubMed]
- Hou, Y.J.; Dong, W.L.; Tan, Y.X.; Yang, G.Z.; Pan, Y.F.; Li, Z.; Tang, L.; Wang, M.; Wang, Q.; Wang, H.Y. Inhibition of active autophagy induces apoptosis and increases chemosensitivity in cholangiocarcinoma. Lab. Investig. 2011, 91, 1146–1157. [Google Scholar] [CrossRef] [PubMed]
- Folkerts, H.; Hilgendorf, S.; Vellenga, E.; Bremer, E.; Wiersma, V.R. The multifaceted role of autophagy in cancer and the microenvironment. Med. Res. Rev. 2019, 39, 517–560. [Google Scholar] [CrossRef]
- Mowers, E.E.; Sharifi, M.N.; Macleod, K.F. Functions of autophagy in the tumor microenvironment and cancer metastasis. FEBS J. 2018, 285, 1751–1766. [Google Scholar] [CrossRef] [PubMed]
- Thuwajit, C.; Ferraresi, A.; Titone, R.; Thuwajit, P.; Isidoro, C. The metabolic cross-talk between epithelial cancer cells and stromal fibroblasts in ovarian cancer progression: Autophagy plays a role. Med. Res. Rev. 2018, 38, 1235–1254. [Google Scholar] [CrossRef] [PubMed]
- Thongchot, S.; Vidoni, C.; Ferraresi, A.; Loilome, W.; Yongvanit, P.; Namwat, N.; Isidoro, C. Dihydroartemisinin induces apoptosis and autophagy-dependent cell death in cholangiocarcinoma through a DAPK1-BECLIN1 pathway. Mol. Carcinog. 2018, 57, 1735–1750. [Google Scholar] [CrossRef] [PubMed]
- Dong, L.W.; Hou, Y.J.; Tan, Y.X.; Tang, L.; Pan, Y.F.; Wang, M.; Wang, H.Y. Prognostic significance of Beclin 1 in intrahepatic cholangiocellular carcinoma. Autophagy 2011, 7, 1222–1229. [Google Scholar] [CrossRef]
- Huang, J.L.; Hezel, A.F. Autophagy in intra-hepatic cholangiocarcinoma. Autophagy 2012, 8, 1148–1149. [Google Scholar] [CrossRef]
- Nitta, T.; Sato, Y.; Ren, X.S.; Harada, K.; Sasaki, M.; Hirano, S.; Nakanuma, Y. Autophagy may promote carcinoma cell invasion and correlate with poor prognosis in cholangiocarcinoma. Int. J. Clin. Exp. Pathol. 2014, 7, 4913–4921. [Google Scholar]
- Qu, X.; Yu, J.; Bhagat, G.; Furuya, N.; Hibshoosh, H.; Troxel, A.; Rosen, J.; Eskelinen, E.L.; Mizushima, N.; Ohsumi, Y.; et al. Promotion of tumorigenesis by heterozygous disruption of the beclin 1 autophagy gene. J. Clin. Investig. 2003, 112, 1809–1820. [Google Scholar] [CrossRef]
- Jamnongkan, W.; Thanee, M.; Yongvanit, P.; Loilome, W.; Thanan, R.; Kimawaha, P.; Boonmars, T.; Silakit, R.; Namwat, N.; Techasen, A. Antifibrotic effect of xanthohumol in combination with praziquantel is associated with altered redox status and reduced iron accumulation during liver fluke-associated cholangiocarcinogenesis. Peer J. 2018, 6, e4281. [Google Scholar] [CrossRef] [PubMed]
- Chuaysri, C.; Thuwajit, P.; Paupairoj, A.; Chau-In, S.; Suthiphongchai, T.; Thuwajit, C. Alpha-smooth muscle actin-positive fibroblasts promote biliary cell proliferation and correlate with poor survival in cholangiocarcinoma. Oncol. Rep. 2009, 21, 957–969. [Google Scholar] [PubMed]
- Vaquero, J.; Aoudjehane, L.; Fouassier, L. Cancer-associated fibroblasts in cholangiocarcinoma. Curr. Opin. Gastroenterol. 2020, 36, 63–69. [Google Scholar] [CrossRef] [PubMed]
- Utispan, K.; Thuwajit, P.; Abiko, Y.; Charngkaew, K.; Paupairoj, A.; Chau-in, S.; Thuwajit, C. Gene expression profiling of cholangiocarcinoma-derived fibroblast reveals alterations related to tumor progression and indicates periostin as a poor prognostic marker. Mol. Cancer 2010, 9, 13. [Google Scholar] [CrossRef] [PubMed]
- Johnson, C.; Han, Y.; Hughart, N.; McCarra, J.; Alpini, G.; Meng, F. Interleukin-6 and its receptor, key players in hepatobiliary inflammation and cancer. Transl. Gastrointest. Cancer 2012, 1, 58–70. [Google Scholar] [PubMed]
- Goydos, J.S.; Brumfield, A.M.; Frezza, E.; Booth, A.; Lotze, M.T.; Carty, S.E. Marked elevation of serum interleukin-6 in patients with cholangiocarcinoma: Validation of utility as a clinical marker. Ann. Surg. 1998, 227, 398–404. [Google Scholar] [CrossRef] [PubMed]
- Thongchot, S.; Ferraresi, A.; Vidoni, C.; Loilome, W.; Yongvanit, P.; Namwat, N.; Isidoro, C. Resveratrol interrupts the pro-invasive communication between cancer associated fibroblasts and cholangiocarcinoma cells. Cancer Lett. 2018, 430, 160–171. [Google Scholar] [CrossRef]
- Tangkijvanich, P.; Thong-ngam, D.; Theamboonlers, A.; Hanvivatvong, O.; Kullavanijaya, P.; Poovorawan, Y. Diagnostic role of serum interleukin 6 and CA 19-9 in patients with cholangiocarcinoma. Hepatogastroenterology 2004, 51, 15–19. [Google Scholar] [PubMed]
- Frampton, G.; Invernizzi, P.; Bernuzzi, F.; Pae, H.Y.; Quinn, M.; Horvat, D.; Galindo, C.; Huang, L.; McMillin, M.; Cooper, B.; et al. Interleukin-6-driven progranulin expression increases cholangiocarcinoma growth by an Akt-dependent mechanism. Gut 2012, 61, 268–277. [Google Scholar] [CrossRef] [PubMed]
- Park, J.; Tadlock, L.; Gores, G.J.; Patel, T. Inhibition of interleukin 6–mediated mitogen-activated protein kinase activation attenuates growth of a cholangiocarcinoma cell line. Hepatology 1999, 30, 1128–1133. [Google Scholar] [CrossRef] [PubMed]
- Daya, M.; Loilome, W.; Techasen, A.; Thanee, M.; Sa-Ngiamwibool, P.; Titapun, A.; Yongvanit, P.; Namwat, N. Progranulin modulates cholangiocarcinoma cell proliferation, apoptosis, and motility via the PI3K/pAkt pathway. Oncol. Targets Ther. 2018, 11, 395–408. [Google Scholar] [CrossRef]
- Yu, L.; Chen, Y.; Tooze, S.A. Autophagy pathway: Cellular and molecular mechanisms. Autophagy 2018, 14, 207–215. [Google Scholar] [CrossRef]
- Kroemer, G.; Mariño, G.; Levine, B. Autophagy and the integrated stress response. Mol. Cell. 2010, 40, 280–293. [Google Scholar] [CrossRef]
- Chavez-Dominguez, R.; Perez-Medina, M.; Lopez-Gonzalez, J.S.; Galicia-Velasco, M.; Aguilar-Cazares, D. The double-edge sword of autophagy in cancer: From tumor suppression to pro-tumor activity. Front. Oncol. 2020, 10, 578418. [Google Scholar] [CrossRef] [PubMed]
- Pérez-Hernández, M.; Arias, A.; Martínez-García, D.; Pérez-Tomás, R.; Quesada, R.; Soto-Cerrato, V. Targeting autophagy for cancer treatment and tumor chemosensitization. Cancers 2019, 11, 1599. [Google Scholar] [CrossRef]
- Ferraresi, A.; Girone, C.; Esposito, A.; Vidoni, C.; Vallino, L.; Secomandi, E.; Dhanasekaran, D.N.; Isidoro, C. How autophagy shapes the tumor microenvironment in ovarian cancer. Front. Oncol. 2020, 10, 599915. [Google Scholar] [CrossRef]
- Valente, G.; Morani, F.; Nicotra, G.; Fusco, N.; Peracchio, C.; Titone, R.; Alabiso, O.; Arisio, R.; Katsaros, D.; Benedetto, C.; et al. Expression and clinical significance of the autophagy proteins BECLIN 1 and LC3 in ovarian cancer. Biomed. Res. Int. 2014, 2014, 462658. [Google Scholar] [CrossRef]
- Thongchot, S.; Thanee, M.; Loilome, W.; Techasen, A.; Boonmars, T.; Sa-Ngiamwibool, P.; Titapun, A.; Yongvanit, P.; Isidoro, C.; Namwat, N. Curative effect of xanthohumol supplementation during liver fluke-associated cholangiocarcinogenesis: Potential involvement of autophagy. J. Tradit. Complement. Med. 2019, 10, 230–235. [Google Scholar] [CrossRef]
- Castino, R.; Isidoro, C.; Murphy, D. Autophagy-dependent cell survival and cell death in an autosomal dominant familial neurohypophyseal diabetes insipidus in vitro model. FASEB J. 2005, 19, 1024–1026. [Google Scholar] [CrossRef] [PubMed]
- Sun, J.M.; Xu, H.T.; Zhao, L.; Zhang, Y.B.; Kang, P.C.; Song, Z.F.; Lui, H.S.; Cui, Y.F. Induction of cell-cycle arrest and apoptosis in human cholangiocarcinoma cells by pristimerin. J. Cell Biochem. 2019, 120, 12002–12009. [Google Scholar] [CrossRef]
- Wang, D.; Guo, H.; Yang, H.; Wang, D.; Gao, P.; Wei, W. Pterostilbene, an active constituent of blueberries, suppresses proliferation potential of human cholangiocarcinoma via enhancing the autophagic flux. Front. Pharmacol. 2019, 10, 1238. [Google Scholar] [CrossRef] [PubMed]
- Ferraresi, A.; Phadngam, S.; Morani, F.; Galetto, A.; Alabio, O.; Chiorino, G.; Isidoro, C. Resveratrol inhibits IL-6-induced ovarian cancer cell migration through epigenetic up-regulation of autophagy. Mol. Carcinog. 2017, 56, 1164–1181. [Google Scholar] [CrossRef] [PubMed]
- Namwat, N.; Puetkasichonpasutha, J.; Loilome, W.; Yongvanit, P.; Techasen, A.; Puapairoj, A.; Sripa, B.; Tassaneeyakul, W.; Khuntikeo, N.; Wongkhan, S. Downregulation of reversion-inducing-cysteine-rich protein with Kazal motifs (RECK) is associated with enhanced expression of matrix metalloproteinases and cholangiocarcinoma metastases. J. Gastroenterol. 2011, 46, 664–675. [Google Scholar] [CrossRef]
- Yongvanit, P.; Pinlaor, S.; Loilome, W. Risk biomarkers for assessment and chemoprevention of liver fluke-associated cholangiocarcinoma. Hepatobiliary Pancreat. Sci. 2014, 21, 309–331. [Google Scholar] [CrossRef][Green Version]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
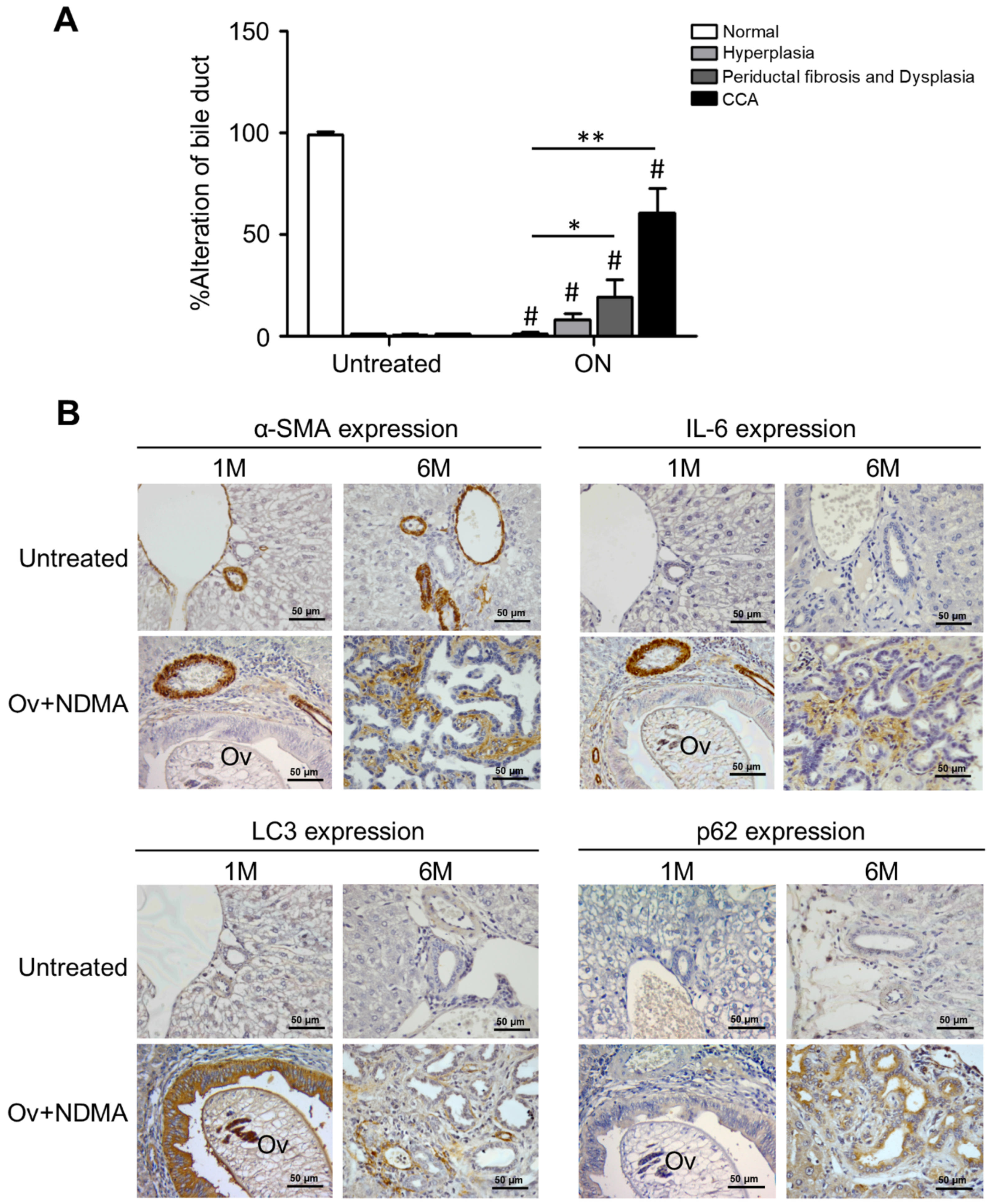{kind=link}
| Factor | n | LC3 | p | P62 | p | IL-6 (in CAFs) | p | |||
|---|---|---|---|---|---|---|---|---|---|---|
| Low | High | Low | High | Low | High | |||||
| Age (years) | ||||||||||
| ≤55 | 35 | 16 | 19 | 0.316 | 27 | 8 | 0.500 | 20 | 15 | 0.500 |
| >55 | 35 | 19 | 16 | 26 | 9 | 19 | 16 | |||
| Sex | ||||||||||
| Female | 27 | 13 | 14 | 0.500 | 22 | 5 | 0.275 | 16 | 11 | 0.412 |
| Male | 43 | 22 | 21 | 31 | 12 | 23 | 20 | |||
| Histological types | ||||||||||
| Non-pap | 33 | 18 | 15 | 0.316 | 26 | 7 | 0.388 | 17 | 16 | 0.335 |
| Papillary | 37 | 17 | 20 | 27 | 10 | 22 | 15 | |||
| Tumor staging | ||||||||||
| I/II | 39 | 17 | 22 | 0.168 | 29 | 10 | 0.496 | 22 | 17 | 0.544 |
| III/IV | 31 | 18 | 13 | 24 | 9 | 17 | 14 | |||
| Tumor size (T stage) | ||||||||||
| T1/T2 | 40 | 18 | 22 | 0.235 | 32 | 8 | 0.246 | 22 | 18 | 0.542 |
| T3/T4 | 30 | 17 | 13 | 8 | 9 | 17 | 13 | |||
| Metastasis to lymph nodes (N stage) | ||||||||||
| N0 | 44 | 23 | 21 | 0.342 | 33 | 11 | 0.099 | 25 | 19 | 0.412 |
| N1/N2 | 26 | 12 | 14 | 20 | 6 | 14 | 12 | |||
| Metastasis to organs (M stage) | ||||||||||
| M0 | 65 | 34 | 31 | 0.178 | 51 | 14 | 0.088 | 37 | 28 | 0.391 |
| M1 | 5 | 1 | 4 | 2 | 3 | 2 | 3 | |||
| Resection status | ||||||||||
| R0 | 49 | 24 | 25 | 0.500 | 37 | 12 | 0.604 | 26 | 23 | 0.339 |
| R1 | 21 | 11 | 10 | 16 | 5 | 13 | 8 | |||
| Drug regimen after surgical resection | ||||||||||
| No | 46 | 24 | 22 | 0.401 | 38 | 8 | 0.604 | 30 | 16 | 0.025 * |
| Yes | 24 | 11 | 13 | 15 | 9 | 9 | 15 | |||
| Recurrence after surgical resection | ||||||||||
| No | 51 | 23 | 28 | 0.141 | 36 | 15 | 0.088 | 29 | 22 | 0.480 |
| Yes | 19 | 12 | 7 | 17 | 2 | 10 | 9 | |||
| Variable (No. of Patients) | No. of Patients | Hazard Ratio (HR) | 95% Confidence Interval (CI) | p-Value |
|---|---|---|---|---|
| LC3 (in cancer) IHC score | ||||
| Low | 35 | 1 | ||
| High | 35 | 0.401 | 0.236–0.681 | 0.001 ** |
| p62 (in cancer) IHC score | ||||
| Low | 53 | 1 | ||
| High | 17 | 0.699 | 0.400–1.221 | 0.208 |
| IL-6 (in CAFs) IHC score | ||||
| Low | 39 | 1 | ||
| High | 31 | 2.004 | 1.138–3.527 | 0.016 * |
| Age (years) | ||||
| ≤55 | 35 | 1 | ||
| >55 | 35 | 0.505 | 0.308–0.828 | 0.252 |
| Sex | ||||
| Female | 27 | 1 | ||
| Male | 43 | 0.844 | 0.507–1.407 | 0.516 |
| Histological types | ||||
| Non-papillary | 33 | 1 | ||
| Papillary | 37 | 1.174 | 0.725–1.903 | 0.514 |
| Tumor stages | ||||
| I–II | 39 | 1 | ||
| III–IV | 31 | 0.535 | 0.330–0.869 | 0.012 * |
| T stages | ||||
| T1/T2 | 40 | 1 | ||
| T3/T4 | 30 | 1.217 | 0.954–1.553 | 0.114 |
| N stages | ||||
| N0 | 44 | 1 | ||
| N1/N2 | 26 | 0.792 | 0.494–1.269 | 0.332 |
| M stages | ||||
| M0 | 65 | 1 | ||
| M1 | 5 | 0.604 | 0.226–1.612 | 0.314 |
| R stages | ||||
| R0 | 41 | 1 | ||
| R1 | 29 | 1.298 | 0.874–1.928 | 0.196 |
| Drug regimen after surgical resection | ||||
| No | 34 | 1 | ||
| Yes | 36 | 1.192 | 0.551–2.581 | 0.655 |
| Drug Regimen | Low IL-6/High LC3/Low p62 | |
|---|---|---|
| Drug regimen Pearson Correlation | 1 | 0.898 ** |
| Sig. (2-tailed) | 0.000 | |
| N | 70 | 70 |
| Low IL-6/High LC3/Low p62 Pearson Correlation | 0.898 ** | 1 |
| Sig. (2-tailed) | 0.000 | |
| N | 70 | 70 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Thongchot, S.; Vidoni, C.; Ferraresi, A.; Loilome, W.; Khuntikeo, N.; Sangkhamanon, S.; Titapun, A.; Isidoro, C.; Namwat, N. Cancer-Associated Fibroblast-Derived IL-6 Determines Unfavorable Prognosis in Cholangiocarcinoma by Affecting Autophagy-Associated Chemoresponse. Cancers 2021, 13, 2134. https://doi.org/10.3390/cancers13092134
Thongchot S, Vidoni C, Ferraresi A, Loilome W, Khuntikeo N, Sangkhamanon S, Titapun A, Isidoro C, Namwat N. Cancer-Associated Fibroblast-Derived IL-6 Determines Unfavorable Prognosis in Cholangiocarcinoma by Affecting Autophagy-Associated Chemoresponse. Cancers. 2021; 13(9):2134. https://doi.org/10.3390/cancers13092134
Chicago/Turabian StyleThongchot, Suyanee, Chiara Vidoni, Alessandra Ferraresi, Watcharin Loilome, Narong Khuntikeo, Sakkarn Sangkhamanon, Attapol Titapun, Ciro Isidoro, and Nisana Namwat. 2021. "Cancer-Associated Fibroblast-Derived IL-6 Determines Unfavorable Prognosis in Cholangiocarcinoma by Affecting Autophagy-Associated Chemoresponse" Cancers 13, no. 9: 2134. https://doi.org/10.3390/cancers13092134
APA StyleThongchot, S., Vidoni, C., Ferraresi, A., Loilome, W., Khuntikeo, N., Sangkhamanon, S., Titapun, A., Isidoro, C., & Namwat, N. (2021). Cancer-Associated Fibroblast-Derived IL-6 Determines Unfavorable Prognosis in Cholangiocarcinoma by Affecting Autophagy-Associated Chemoresponse. Cancers, 13(9), 2134. https://doi.org/10.3390/cancers13092134