Brazilian Table Olives: A Source of Lactic Acid Bacteria with Antimycotoxigenic and Antifungal Activity

 , ,
, ,  ,
,  and
and
Abstract
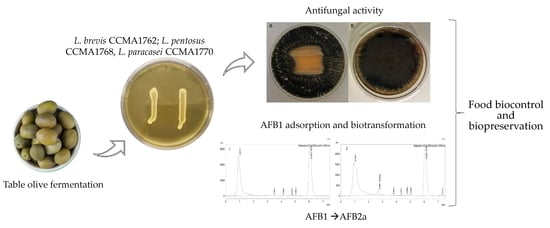
1. Introduction
2. Results
2.1. Screening of LAB for Antifungal Properties
2.2. Plate Inhibition Analysis of Cell-Free Supernatants
2.3. Microplate Inhibition Analysis of Cell-Free Supernatants
2.4. Analyses of Mycotoxins in Inhibition Assays
2.5. Analysis of Cell-Free Supernatant
2.6. Screening of LAB Mycotoxin-Detoxifying Properties
2.7. Evaluation of AFB1 Biotransformation into AFB2a
3. Discussion
4. Conclusions
5. Materials and Methods
5.1. Chemicals and Culture Media
5.2. Lactic acid Bacteria Strains
5.3. Fungal Strain and Inoculum Production
5.4. Screening of LAB for Antifungal Properties
5.5. Antifungal Properties of LAB Cell-Free Supernatants
5.5.1. Plate Inhibition Analysis
5.5.2. Microplate Inhibition Analysis
5.6. HPLC Analysis of Cell-Free Supernatant
5.7. Assessment of LAB Anti-Mycotoxins Properties
5.8. Mycotoxins Analysis
5.9. Evaluation of AFB1 Biotransformation into AFB2a
5.10. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Davies, C.R.; Wohlgemuth, F.; Young, T.; Violet, J.; Dickinson, M.; Sanders, J.W.; Vallieres, C.; Avery, S.V. Evolving challenges and strategies for fungal control in the food supply chain. Fungal Biol. Rev. 2021, 36, 15–26. [Google Scholar] [CrossRef] [PubMed]
- Moretti, A.; Logrieco, A.F.; Susca, A. mycotoxins: An underhand food problem. Mycotoxigenic Fungi 2017, 1541, 3–12. [Google Scholar] [CrossRef]
- Eskola, M.; Kos, G.; Elliott, C.T.; Hajšlová, J.; Mayar, S.; Krska, R. Worldwide contamination of food-crops with mycotoxins: Validity of the widely cited ‘FAO estimate’ of 25%. Crit. Rev. Food Sci. Nutr. 2020, 60, 2773–2789. [Google Scholar] [CrossRef]
- Kumar, A.; Pathak, H.; Bhadauria, S.; Sudan, J. Aflatoxin contamination in food crops: Causes, detection, and management: A review. Food Prod. Process. Nutr. 2021, 3, 17. [Google Scholar] [CrossRef]
- Meneely, J.P.; Kolawole, O.; Haughey, S.A.; Miller, S.J.; Krska, R.; Elliott, C.T. The challenge of global aflatoxins legislation with a focus on peanuts and peanut products: A systematic review. Expo. Health 2022, 1–21. [Google Scholar] [CrossRef]
- Freire, F.D.C.O.; da Rocha, M.E.B. Impact of mycotoxins on human health. In Fungal Metabolites; Springer: Cham, Switzerland, 2017; pp. 239–261. [Google Scholar]
- Niessen, L.; Schmidt, H.; Muhlencoert, E.; Farber, P.; Karolewiez, A.; Geisen, R. Advances in the molecular diagnosis of ochratoxin A-producing fungi. Food Addit. Contam. 2005, 22, 324–334. [Google Scholar] [CrossRef] [PubMed]
- Frisvad, J.C. A critical review of producers of small lactone mycotoxins: Patulin, penicillic acid and moniliformin. World Mycotoxin J. 2018, 11, 73–100. [Google Scholar] [CrossRef]
- Geisen, R.; Schmidt-Heydt, M.; Touhami, N.; Himmelsbach, A. New aspects of ochratoxin A and citrinin biosynthesis in penicillium. Curr. Opin. Food Sci. 2018, 23, 23–31. [Google Scholar] [CrossRef]
- Gourama, H.; Bullerman, L.B. Mycotoxin production by molds isolated from Greek-style black olives. Int. J. Food Microbiol. 1988, 6, 81–90. [Google Scholar] [CrossRef]
- Bavaro, S.L.; Susca, A.; Frisvad, J.C.; Tufariello, M.; Chytiri, A.; Perrone, G.; Mita, G.; Logrieco, A.F.; Bleve, G. Isolation, characterization, and selection of molds associated to fermented black table olives. Front. Microbiol. 2017, 8, 1356. [Google Scholar] [CrossRef] [PubMed]
- Heperkan, D.; Dazkır, G.S.; Kansu, D.Z.; Karbancıoglu Güler, F. Influence of temperature on citrinin accumulation by Penicillium citrinum and Penicillium verrucosum in black table olives. Toxin Rev. 2009, 28, 180–186. [Google Scholar] [CrossRef]
- Guimarães, A.; Santiago, A.; Teixeira, J.A.; Venâncio, A.; Abrunhosa, L. Anti-aflatoxigenic effect of organic acids produced by Lactobacillus plantarum. Int. J. Food Microbiol. 2018, 264, 31–38. [Google Scholar] [CrossRef]
- Simões, L.; Fernandes, N.; de Souza, A.; dos Santos, L.; Magnani, M.; Abrunhosa, L.; Teixeira, J.; Schwan, R.F.; Dias, D.R. Probiotic and antifungal attributes of lactic acid bacteria isolates from naturally fermented brazilian table olives. Fermentation 2022, 8, 277. [Google Scholar] [CrossRef]
- Ngea, G.L.N.; Yang, Q.; Tchabo, W.; Castoria, R.; Zhang, X.; Zhang, H. Leuconostoc mesenteroides subsp. mesenteroides LB7 isolated from apple surface inhibits P. expansum in vitro and reduces patulin in fruit juices. Int. J. Food Microbiol. 2021, 339, 109025. [Google Scholar] [CrossRef] [PubMed]
- Li, L.; Yang, B.; Humza, M.; Geng, H.; Wang, G.; Zhang, C.; Gao, S.; Xing, F.; Liu, Y. A novel strain Lactobacillus brevis 8-2B inhibiting Aspergillus carbonarius growth and ochratoxin A production. LWT 2021, 136, 110308. [Google Scholar] [CrossRef]
- Fabiszewska, A.U.; Zielińska, K.; Wróbel, B. Trends in designing microbial silage quality by biotechnological methods using lactic acid bacteria inoculants: A minireview. World J. Microbiol. Biotechnol. 2019, 35, 1–8. [Google Scholar] [CrossRef]
- Zhao, S.; Hao, X.; Yang, F.; Wang, Y.; Fan, X.; Wang, Y. Antifungal activity of Lactobacillus plantarum ZZUA493 and its application to extend the shelf life of Chinese steamed buns. Foods 2022, 11, 195. [Google Scholar] [CrossRef] [PubMed]
- Corsetti, A.; Gobbetti, M.; Rossi, J.; Damiani, P. Antimould activity of sourdough lactic acid bacteria: Identification of a mixture of organic acids produced by Lactobacillus sanfrancisco CB1. Appl. Microbiol. Biotechnol. 1998, 50, 253–256. [Google Scholar] [CrossRef] [PubMed]
- Siedler, S.; Balti, R.; Neves, A.R. Bioprotective mechanisms of lactic acid bacteria against fungal spoilage of food. Curr. Opin. Biotechnol. 2019, 56, 138–146. [Google Scholar] [CrossRef] [PubMed]
- Heperkan, D.; Meric, B.E.; Sismanoglu, G.; Dalkiliç, G.; Güler, F.K. Mycobiota, mycotoxigenic fungi, and citrinin production in black olives. In Advances in Food Mycology; Springer: Cham, Switzerland, 2006; pp. 203–210. [Google Scholar]
- Pérez-Nevado, F.; Mateos, P.; Hernández, A.; Martín, A.; Benito, M.; Ruiz-Moyano, S.; Córdoba, M. Identification of molds associated with green table olives. In Microorganisms in Industry and Environment: From Scientific and Industrial Research to Consumer Products; World Scientific: Singapore; Shanghai, China, 2011; pp. 398–402. [Google Scholar]
- Khalil, A.M.A.; Hashem, A.H.; Abdelaziz, A.M. Occurrence of toxigenic Penicillium polonicum in retail green table olives from the Saudi Arabia market. Biocatal. Agric. Biotechnol. 2019, 21, 101314. [Google Scholar] [CrossRef]
- El Adlouni, C.; Tozlovanu, M.; Naman, F.; Faid, M.; Pfohl-Leszkowicz, A. Preliminary data on the presence of mycotoxins (ochratoxin A, citrinin and aflatoxin B1) in black table olives “Greek style” of Moroccan origin. Mol. Nutr. Food Res. 2006, 50, 507–512. [Google Scholar] [CrossRef]
- Ghitakou, S.; Koutras, K.; Kanellou, E.; Markaki, P. Study of aflatoxin B1 and ochratoxin A production by natural microflora and Aspergillus parasiticus in black and green olives of Greek origin. Food Microbiol. 2006, 23, 612–621. [Google Scholar] [CrossRef]
- Franzetti, L.; Scarpellini, M.; Vecchio, A.; Planeta, D. Microbiological and safety evaluation of green table olives marketed in Italy. Ann. Microbiol. 2011, 61, 843–851. [Google Scholar] [CrossRef]
- Simões, L.A.; Cristina de Souza, A.; Ferreira, I.; Melo, D.S.; Lopes, L.A.A.; Magnani, M.; Schwan, R.F.; Dias, D.R. Probiotic properties of yeasts isolated from Brazilian fermented table olives. J. Appl. Microbiol. 2021, 131, 1983–1997. [Google Scholar] [CrossRef]
- De Chaves, M.A.; Reginatto, P.; da Costa, B.S.; de Paschoal, R.I.; Teixeira, M.L.; Fuentefria, A.M. Fungicide resistance in Fusarium graminearum species complex. Curr. Microbiol. 2022, 79, 62. [Google Scholar] [CrossRef] [PubMed]
- Magan, N.; Hope, R.; Colleate, A.; Baxter, E.S. Relationship between growth and mycotoxin production by Fusarium species, biocides and environment. Eur. J. Plant Pathol. 2002, 108, 685–690. [Google Scholar] [CrossRef]
- Perpetuini, G.; Prete, R.; Garcia-Gonzalez, N.; Khairul Alam, M.; Corsetti, A. Table Olives More than a Fermented Food. Foods 2020, 9, 178. [Google Scholar] [CrossRef] [PubMed]
- Russo, P.; Arena, M.P.; Fiocco, D.; Capozzi, V.; Drider, D.; Spano, G. Lactobacillus plantarum with broad antifungal activity: A promising approach to increase safety and shelf-life of cereal-based products. Int. J. Food Microbiol. 2017, 247, 48–54. [Google Scholar] [CrossRef]
- Gerez, C.L.; Torino, M.I.; Rollán, G.; de Valdez, G.F. Prevention of bread mould spoilage by using lactic acid bacteria with antifungal properties. Food Control 2009, 20, 144–148. [Google Scholar] [CrossRef]
- Hassan, Y.I.; Bullerman, L.B. Antifungal activity of Lactobacillus paracasei ssp. tolerans isolated from a sourdough bread culture. Int. J. Food Microbiol. 2008, 121, 112–115. [Google Scholar] [CrossRef] [PubMed]
- Ispirli, H.; Dertli, E. Isolation and characterisation of lactic acid bacteria from traditional koumiss and kurut. Int. J. Food Prop. 2017, 20, S2441–S2449. [Google Scholar] [CrossRef]
- Ruggirello, M.; Nucera, D.; Cannoni, M.; Peraino, A.; Rosso, F.; Fontana, M.; Cocolin, L.; Dolci, P. Antifungal activity of yeasts and lactic acid bacteria isolated from cocoa bean fermentations. Food Res. Int. 2019, 115, 519–525. [Google Scholar] [CrossRef] [PubMed]
- Dopazo, V.; Luz, C.; Quiles, J.M.; Calpe, J.; Romano, R.; Manes, J.; Meca, G. Potential application of lactic acid bacteria in the biopreservation of red grape from mycotoxigenic fungi. J. Sci. Food Agric. 2022, 102, 898–907. [Google Scholar] [CrossRef] [PubMed]
- Chen, H.; Ju, H.; Wang, Y.; Du, G.; Yan, X.; Cui, Y.; Yuan, Y.; Yue, T. Antifungal activity and mode of action of Lactic acid bacteria isolated from Kefir against Penicillium expansum. Food Control 2021, 130, 108274. [Google Scholar] [CrossRef]
- Mateo, E.M.; Tarazona, A.; Jiménez, M.; Mateo, F. Lactic acid bacteria as potential agents for biocontrol of aflatoxigenic and ochratoxigenic fungi. Toxins 2022, 14, 807. [Google Scholar] [CrossRef]
- Fernández, A.G.; Adams, M.R.; Fernandez-Diez, M. Table Olives: Production and Processing; Springer Science & Business Media: Berlin/Heidelberg, Germany, 1997. [Google Scholar]
- Abouloifa, H.; Rokni, Y.; Bellaouchi, R.; Ghabbour, N.; Karboune, S.; Brasca, M.; Ben Salah, R.; Chihib, N.E.; Saalaoui, E.; Asehraou, A. Characterization of probiotic properties of antifungal Lactobacillus strains isolated from traditional fermenting green olives. Probiotics Antimicrob. Proteins 2020, 12, 683–696. [Google Scholar] [CrossRef]
- Panebianco, F.; Caridi, A. New insights into the antifungal activity of lactic acid bacteria isolated from different food matrices. Grasas Y Aceites 2021, 72, e400. [Google Scholar] [CrossRef]
- Abouloifa, H.; Gaamouche, S.; Rokni, Y.; Hasnaoui, I.; Bellaouchi, R.; Ghabbour, N.; Karboune, S.; Brasca, M.; D’Hallewin, G.; Salah, R.B. Antifungal activity of probiotic Lactobacillus strains isolated from natural fermented green olives and their application as food bio-preservative. Biol. Control. 2021, 152, 104450. [Google Scholar] [CrossRef]
- Riolo, M.; Luz, C.; Santilli, E.; Meca, G.; Cacciola, S.O. Antifungal activity of selected lactic acid bacteria from olive drupes. BioRxiv 2022, 7, 1–27. [Google Scholar] [CrossRef]
- El Oirdi, S.; Lakhlifi, T.; Bahar, A.A.; Yatim, M.; Rachid, Z.; Belhaj, A. Isolation and identification of Lactobacillus plantarum 4F, a strain with high antifungal activity, fungicidal effect, and biopreservation properties of food. J. Food Process. Preserv. 2021, 45, e15517. [Google Scholar] [CrossRef]
- Sadiq, F.A.; Yan, B.; Tian, F.; Zhao, J.; Zhang, H.; Chen, W. Lactic acid bacteria as antifungal and anti-mycotoxigenic agents: A comprehensive review. Compr. Rev. Food Sci. Food Saf. 2019, 18, 1403–1436. [Google Scholar] [CrossRef] [PubMed]
- De Angelis, M.; Corsetti, A.; Tosti, N.; Rossi, J.; Corbo, M.R.; Gobbetti, M. Characterization of non-starter lactic acid bacteria from Italian ewe cheeses based on phenotypic, genotypic, and cell wall protein analyses. Appl. Environ. Microbiol. 2001, 67, 2011–2020. [Google Scholar] [CrossRef] [PubMed]
- Bleve, G.; Tufariello, M.; Durante, M.; Perbellini, E.; Ramires, F.A.; Grieco, F.; Cappello, M.S.; De Domenico, S.; Mita, G.; Tasioula-Margari, M. Physico-chemical and microbiological characterization of spontaneous fermentation of Cellina di Nardò and Leccino table olives. Front. Microbiol. 2014, 5, 570. [Google Scholar] [CrossRef] [PubMed]
- Chorianopoulos, N.; Boziaris, I.; Stamatiou, A.; Nychas, G.-J. Microbial association and acidity development of unheated and pasteurized green-table olives fermented using glucose or sucrose supplements at various levels. Food Microbiol. 2005, 22, 117–124. [Google Scholar] [CrossRef]
- Nychas, G.J.; Panagou, E.; Parker, M.; Waldron, K.; Tassou, C. Microbial colonization of naturally black olives during fermentation and associated biochemical activities in the cover brine. Lett. Appl. Microbiol. 2002, 34, 173–177. [Google Scholar] [CrossRef]
- Bleve, G.; Tufariello, M.; Durante, M.; Grieco, F.; Ramires, F.A.; Mita, G.; Tasioula-Margari, M.; Logrieco, A.F. Physico-chemical characterization of natural fermentation process of Conservolea and Kalamàta table olives and development of a protocol for the pre-selection of fermentation starters. Food Microbiol. 2015, 46, 368–382. [Google Scholar] [CrossRef] [PubMed]
- Panagou, E.Z.; Schillinger, U.; Franz, C.M.; Nychas, G.J.E. Microbiological and biochemical profile of cv. Conservolea naturally black olives during controlled fermentation with selected strains of lactic acid bacteria. Food Microbiol. 2008, 25, 348–358. [Google Scholar] [CrossRef]
- Garnier, L.; Penland, M.; Thierry, A.; Maillard, M.B.; Jardin, J.; Coton, M.; Salas, M.L.; Coton, E.; Valence, F.; Mounier, J. Antifungal activity of fermented dairy ingredients: Identification of antifungal compounds. Int. J. Food Microbiol. 2020, 322, 108574. [Google Scholar] [CrossRef]
- Luz, C.; D’Opazo, V.; Quiles, J.; Romano, R.; Mañes, J.; Meca, G. Biopreservation of tomatoes using fermented media by lactic acid bacteria. LWT 2020, 130, 109618. [Google Scholar] [CrossRef]
- Nazareth, T.d.M.; Luz, C.; Torrijos, R.; Quiles, J.M.; Luciano, F.B.; Mañes, J.; Meca, G. Potential application of lactic acid bacteria to reduce aflatoxin B1 and fumonisin B1 occurrence on corn kernels and corn ears. Toxins 2020, 12, 21. [Google Scholar] [CrossRef]
- Oude Elferink, S.J.; Krooneman, J.; Gottschal, J.C.; Spoelstra, S.F.; Faber, F.; Driehuis, F. Anaerobic conversion of lactic acid to acetic acid and 1, 2-propanediol by Lactobacillus buchneri. Appl. Environ. Microbiol. 2001, 67, 125–132. [Google Scholar] [CrossRef]
- Palles, T.; Beresford, T.; Condon, S.; Cogan, T. Citrate metabolism in Lactobacillus casei and Lactobacillus plantarum. J. Appl. Microbiol. 1998, 85, 147–154. [Google Scholar] [CrossRef]
- Lahtinen, S.; Ouwehand, A.C.; Salminen, S.; von Wright, A. Lactic Acid Bacteria: Microbiological and Functional Aspects; CRC Press: Boca Raton, FL, USA, 2011. [Google Scholar]
- Guimarães, A.; Venâncio, A.; Abrunhosa, L. Antifungal effect of organic acids from lactic acid bacteria on Penicillium nordicum. Food Addit. Contam. Part A 2018, 35, 1803–1818. [Google Scholar] [CrossRef]
- Liu, A.; Zheng, Y.; Liu, L.; Chen, S.; He, L.; Ao, X.; Yang, Y.; Liu, S. Decontamination of aflatoxins by lactic acid bacteria. Curr. Microbiol. 2020, 77, 3821–3830. [Google Scholar] [CrossRef]
- Zheng, X.; Wei, W.; Rao, S.; Gao, L.; Li, H.; Yang, Z. Degradation of patulin in fruit juice by a lactic acid bacteria strain Lactobacillus casei YZU01. Food Control 2020, 112, 107147. [Google Scholar] [CrossRef]
- Nasrollahzadeh, A.; Mokhtari, S.; Khomeiri, M.; Saris, P. Mycotoxin detoxification of food by lactic acid bacteria. Int. J. Food Contam. 2022, 9, 1–9. [Google Scholar] [CrossRef]
- Bahati, P.; Zeng, X.; Uzizerimana, F.; Tsoggerel, A.; Awais, M.; Qi, G.; Cai, R.; Yue, T.; Yuan, Y. Adsorption mechanism of patulin from apple juice by inactivated lactic acid bacteria isolated from kefir grains. Toxins 2021, 13, 434. [Google Scholar] [CrossRef]
- Møller, C.O.d.A.; Freire, L.; Rosim, R.E.; Margalho, L.P.; Balthazar, C.F.; Franco, L.T.; Sant’Ana, A.d.S.; Corassin, C.H.; Rattray, F.P.; Oliveira, C.A.F.d. Effect of lactic acid bacteria strains on the growth and aflatoxin production potential of Aspergillus parasiticus, and their ability to bind aflatoxin B1, ochratoxin A, and zearalenone in vitro. Front. Microbiol. 2021, 12, 899. [Google Scholar] [CrossRef]
- Wacoo, A.P.; Atukunda, P.; Muhoozi, G.; Braster, M.; Wagner, M.; Van Den Broek, T.J.; Sybesma, W.; Westerberg, A.C.; Iversen, P.O.; Kort, R. Aflatoxins: Occurrence, exposure, and binding to Lactobacillus species from the gut microbiota of rural Ugandan children. Microorganisms 2020, 8, 347. [Google Scholar] [CrossRef]
- Aiko, V.; Edamana, P.; Mehta, A. Decomposition and detoxification of aflatoxin B1 by lactic acid. J. Sci. Food Agric. 2016, 96, 1959–1966. [Google Scholar] [CrossRef]
- Rushing, B.R.; Selim, M.I. Effect of dietary acids on the formation of aflatoxin B2a as a means to detoxify aflatoxin B1. Food Addit. Contam. Part A 2016, 33, 1456–1467. [Google Scholar] [CrossRef] [PubMed]
- Wong, J.J.; Hsieh, D. Mutagenicity of aflatoxins related to their metabolism and carcinogenic potential. Proc. Natl. Acad. Sci. USA 1976, 73, 2241–2244. [Google Scholar] [CrossRef]
- Xu, H.; Wang, L.; Sun, J.; Wang, L.; Guo, H.; Ye, Y.; Sun, X. Microbial detoxification of mycotoxins in food and feed. Crit. Rev. Food Sci. Nutr. 2022, 62, 4951–4969. [Google Scholar] [CrossRef] [PubMed]
- Piotrowska, M. Microbiological Decontamination of Mycotoxins: Opportunities and Limitations. Toxins 2021, 13, 819. [Google Scholar] [CrossRef]
- Peres, C.M.; Peres, C.; Hernández-Mendoza, A.; Malcata, F.X. Review on fermented plant materials as carriers and sources of potentially probiotic lactic acid bacteria–with an emphasis on table olives. Trends Food Sci. Technol. 2012, 26, 31–42. [Google Scholar] [CrossRef]
- Magnusson, J.; Schnürer, J. Lactobacillus coryniformis subsp. coryniformis strain Si3 produces a broad-spectrum proteinaceous antifungal compound. Appl. Environ. Microbiol. 2001, 67, 1–5. [Google Scholar] [CrossRef]
- Taheur, F.B.; Fedhila, K.; Chaieb, K.; Kouidhi, B.; Bakhrouf, A.; Abrunhosa, L. Adsorption of aflatoxin B1, zearalenone and ochratoxin A by microorganisms isolated from Kefir grains. Int. J. Food Microbiol. 2017, 251, 1–7. [Google Scholar] [CrossRef]
- Soares, C.; Rodrigues, P.; Freitas-Silva, O.; Abrunhosa, L.; Venâncio, A. HPLC method for simultaneous detection of aflatoxins and cyclopiazonic acid. World Mycotoxin J. 2010, 3, 225–231. [Google Scholar] [CrossRef]
- Abrunhosa, L.; Inês, A.; Rodrigues, A.I.; Guimarães, A.; Pereira, V.L.; Parpot, P.; Mendes-Faia, A.; Venâncio, A. Biodegradation of ochratoxin A by Pediococcus parvulus isolated from Douro wines. Int. J. Food Microbiol. 2014, 188, 45–52. [Google Scholar] [CrossRef]
- Wang, L.; Yue, T.; Yuan, Y.; Wang, Z.; Ye, M.; Cai, R. A new insight into the adsorption mechanism of patulin by the heat-inactive lactic acid bacteria cells. Food Control 2015, 50, 104–110. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Species | Antifungal Activity | |||
|---|---|---|---|---|
| A. flavus MUM 08.201 | A. carbonarius 01UAs293 | P. nordicum MUM 08.16 | P. expansum MUM 17.86 | |
| L. brevis CCMA 1766 | − | − | +++ | + |
| L. paracasei CCMA 1763 | + | ++ | ++ | + |
| L. paracasei CCMA 1772 | − | − | + | ++ |
| L. pentosus CCMA 1768 | + | ++ | +++ | +++ |
| L. paracasei CCMA1775 | ++ | − | +++ | ++ |
| L. paracasei CCMA 1769 | − | − | + | + |
| L. paracasei CCMA 1770 | − | − | + | + |
| L. brevis CCMA 1762 | + | +++ | ++ | +++ |
| L. paracasei CCMA 1764 | + | +++ | +++ | +++ |
| L. paracasei CCMA 1771 | − | − | +++ | ++ |
| L. brevis CCMA 1765 | − | − | +++ | + |
| L. paracasei CCMA 1767 | − | − | +++ | + |
| L. paracasei CCMA 1773 | − | − | − | − |
| L. paracasei CCMA 1774 | − | − | + | + |
| LAB Strains | Plate Method | Microplate Method | |||||
|---|---|---|---|---|---|---|---|
| Fungi | CCMA 1762 | CCMA 1764 | CCMA 1768 | CCMA 1762 | CCMA 1764 | CCMA 1768 | |
| A. carbonarius 01UAs293 | CFS (300 µL/mL) | 0 | 0 | 0 | 12 b ± 1.2 | 12 b ± 1.4 | 27 c* ± 2.3 |
| A. flavus MUM 08.201 | 4.7 c ± 1.2 | 6.6 c ± 0.6 | 3.5 c ± 1.1 | 18 b ± 0.9 | 15 b ± 0.8 | 14 d ± 1.7 | |
| P. nordicum MUM 08.16 | 39 b ± 1.8 | 37 b ± 1.5 | 40.7 b ± 0.0 | 16 b* ± 0.4 | 95 a ± 1.5 | 95 a ± 1.1 | |
| P. expansum MUM 17.86 | 78 a* ± 1.1 | 92 a ± 1.1 | 92 a ± 0.9 | 87 a ± 2.1 | 93 a* ± 0.9 | 89 b ± 2.4 | |
| A. carbonarius 01UAs293 | CFS (500 µL/mL) | 40 c* ± 1.7 | 100 a | 100 a | 30 c* ± 2.5 | 100 a | 100 a |
| A. flavus MUM 08.201 | 78 b* ± 0.6 | 100 a | 98 a ± 0.6 | 73 b* ± 3.2 | 99 a ± 0.1 | 98 a ± 1.2 | |
| P. nordicum MUM 08.16 | 100 a | 100 a | 100 a | 100 a | 100 a | 100 a | |
| P. expansum MUM 17.86 | 77 b* ± 1.2 | 100 a | 100 a | 96 a ± 3.0 | 100 a | 100 a | |
| Mycotoxins | LAB Strains | Plate Method | Microplate Method | |||
|---|---|---|---|---|---|---|
| Concentration (µg/mL) | Reduction (%) | Concentration (µg/mL) | Reduction (%) | |||
| AFB1 | CCMA 1762 | CFS concentration 300 µL/mL | 4.21 a ± 0.7 | 18% | 3.77 a ± 0.6 | 8% |
| CCMA 1764 | 3.63 a ± 0.1 | 29% | 2.85 a ± 1.0 | 31% | ||
| CCMA 1768 | 3.79 a ± 0.2 | 26% | 3.27 a ± 1.1 | 20% | ||
| Control | 5.13 a ± 0.4 | 4.11 a ± 0.8 | ||||
| OTA | CCMA 1762 | 9.74 a ± 0.1 | - | 5.76 b ± 1.0 | - | |
| CCMA 1764 | 8.49 a ± 1.7 | - | 3.83 b ± 0.7 | - | ||
| CCMA 1768 | 5.31 a* ± 0.9 | - | 4.21 a ± 0.2 | - | ||
| Control | 5.26 a* ± 0.9 | 1.45 b* ± 1.1 | ||||
| PAT | CCMA 1762 | 7.71 a ± 0.9 | 89% | 3.60 b ± 0.37 | 95% | |
| CCMA 1764 | 1.65 a ± 1.2 | 98% | 0 | 100% | ||
| CCMA 1768 | 1.38 a ± 1.46 | 98% | 0 | 100% | ||
| Control | 73.62 a* ± 6.0 | 69.71 b* ± 0.5 | ||||
| AFB1 | CCMA 1762 | CFS concentration 500 µL/mL | 0.24 a ± 0 | 95% | 0.06 a ± 0 | 98% |
| CCMA 1764 | 0 | 100% | 0 | 100% | ||
| CCMA 1768 | 0 | 100% | 0 | 100% | ||
| Control | 5.13 a* ± 0.4 | 4.11 a* ± 0.8 | ||||
| OTA | CCMA 1762 | 1.81 a ± 1.3 | 65% | 0.28 b ± 0.1 | 81% | |
| CCMA 1764 | 0 | 100% | 0 | 100% | ||
| CCMA 1768 | 0 | 100% | 0 | 100% | ||
| Control | 5.26 a* ± 0.9 | 1.45 b ± 1.1 | ||||
| PAT | CCMA 1762 | 5.12 a ± 0.1 | 93% | 3.27 a ± 0.2 | 95% | |
| CCMA 1764 | 0 | 100% | 0 | 100% | ||
| CCMA 1768 | 0 | 100% | 0 | 100% | ||
| Control | 73.62 a* ± 6.0 | 69.71 b* ± 0.5 | ||||
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Simões, L.; Fernandes, N.; Teixeira, J.; Abrunhosa, L.; Dias, D.R. Brazilian Table Olives: A Source of Lactic Acid Bacteria with Antimycotoxigenic and Antifungal Activity. Toxins 2023, 15, 71. https://doi.org/10.3390/toxins15010071
Simões L, Fernandes N, Teixeira J, Abrunhosa L, Dias DR. Brazilian Table Olives: A Source of Lactic Acid Bacteria with Antimycotoxigenic and Antifungal Activity. Toxins. 2023; 15(1):71. https://doi.org/10.3390/toxins15010071
Chicago/Turabian StyleSimões, Luara, Natália Fernandes, José Teixeira, Luís Abrunhosa, and Disney Ribeiro Dias. 2023. "Brazilian Table Olives: A Source of Lactic Acid Bacteria with Antimycotoxigenic and Antifungal Activity" Toxins 15, no. 1: 71. https://doi.org/10.3390/toxins15010071
APA StyleSimões, L., Fernandes, N., Teixeira, J., Abrunhosa, L., & Dias, D. R. (2023). Brazilian Table Olives: A Source of Lactic Acid Bacteria with Antimycotoxigenic and Antifungal Activity. Toxins, 15(1), 71. https://doi.org/10.3390/toxins15010071