Effect of Fumonisin B1 on Proliferation and Apoptosis of Intestinal Porcine Epithelial Cells

Abstract
:1. Introduction
2. Results
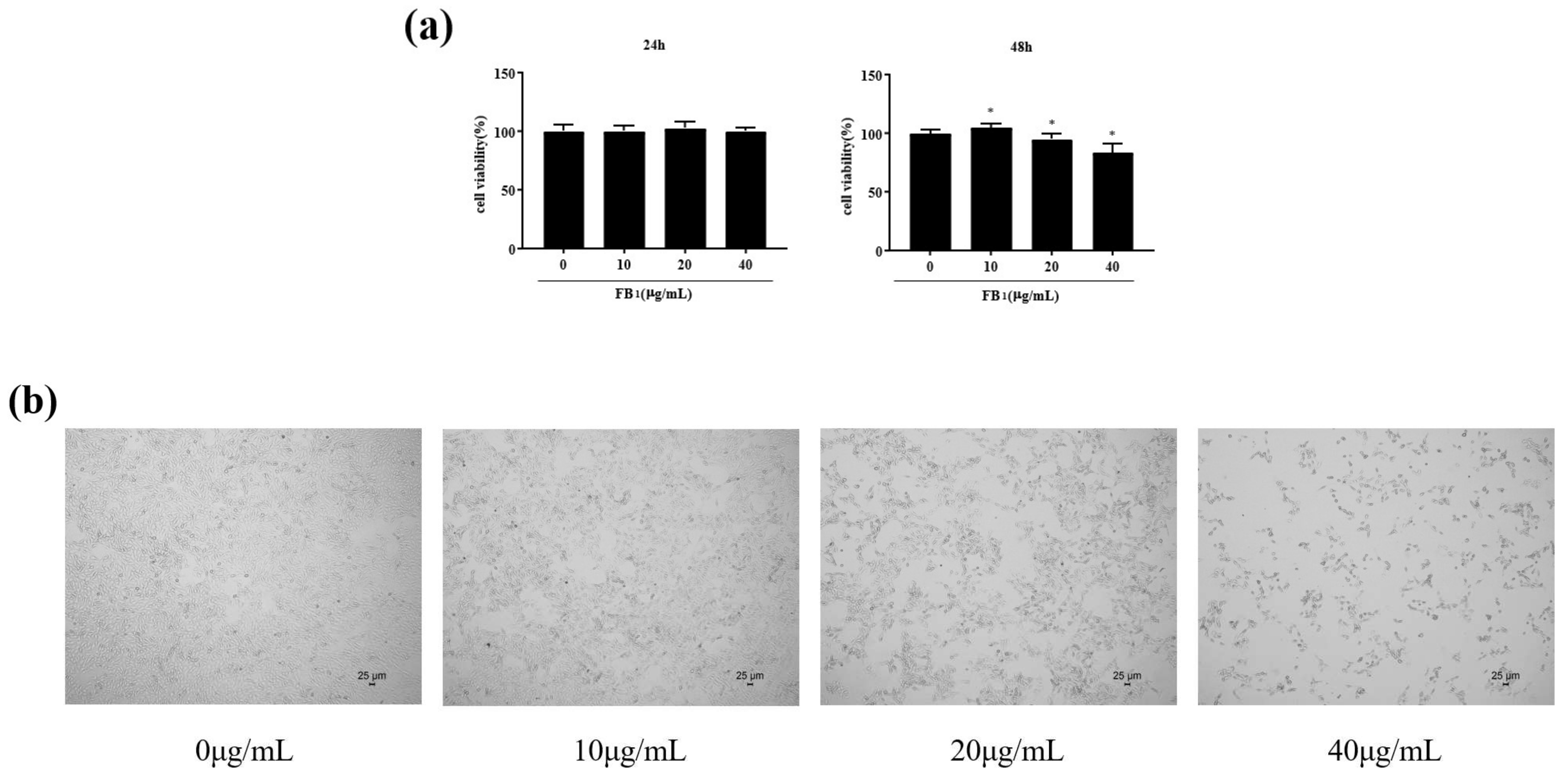
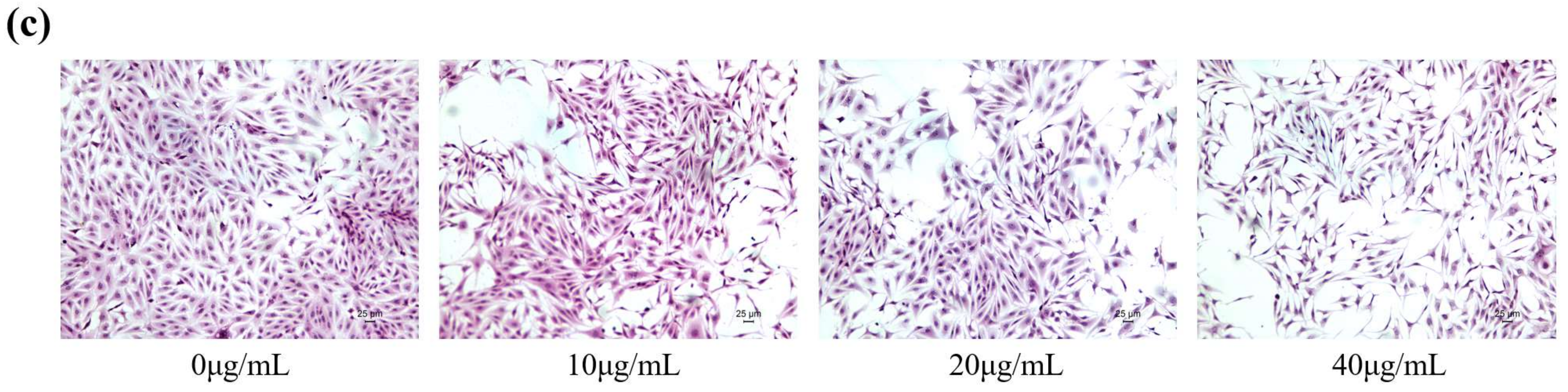
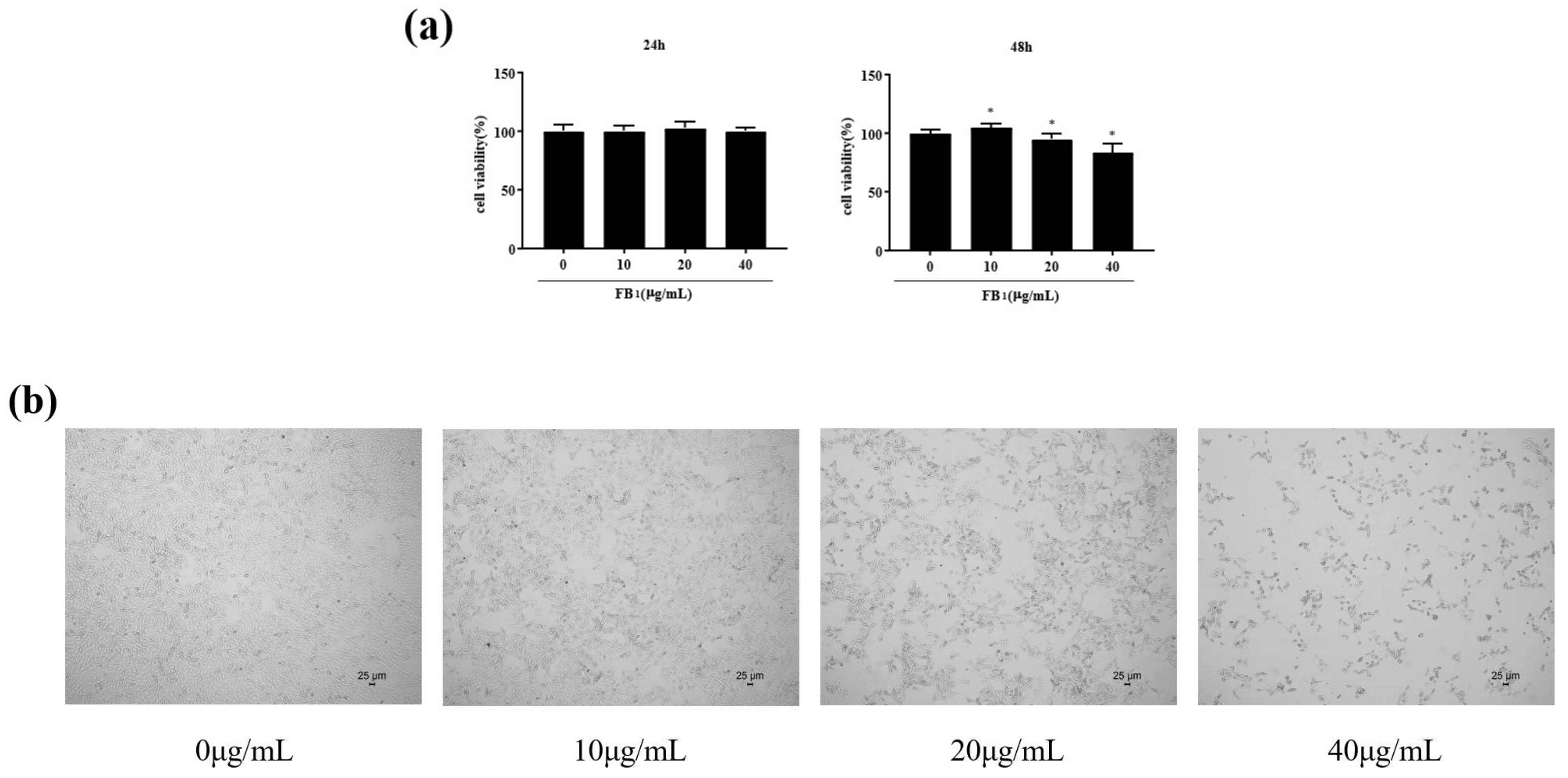
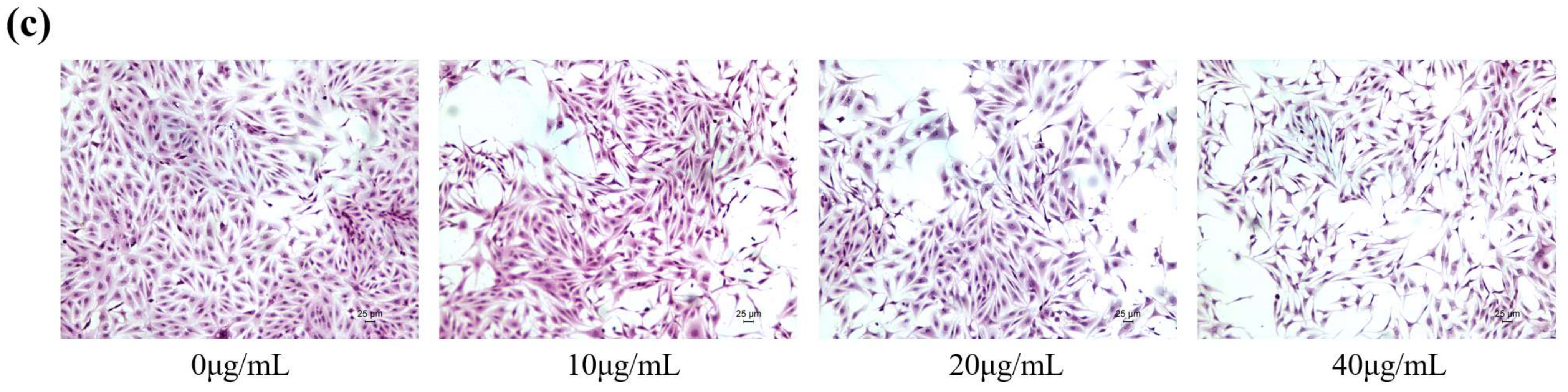
2.1. FB1 Reduced the Viability of IPEC-J2 Cells and Caused Cell Damage
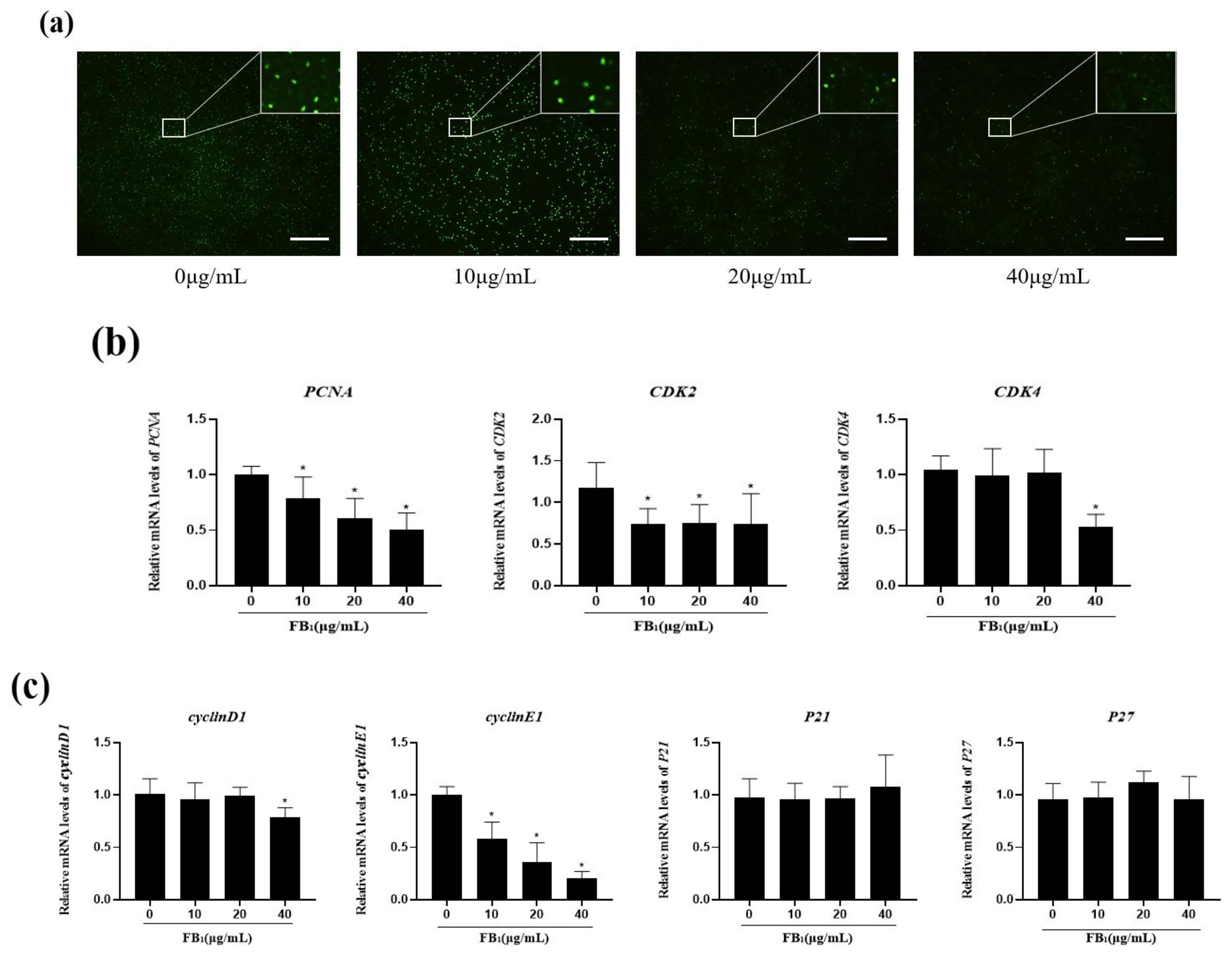
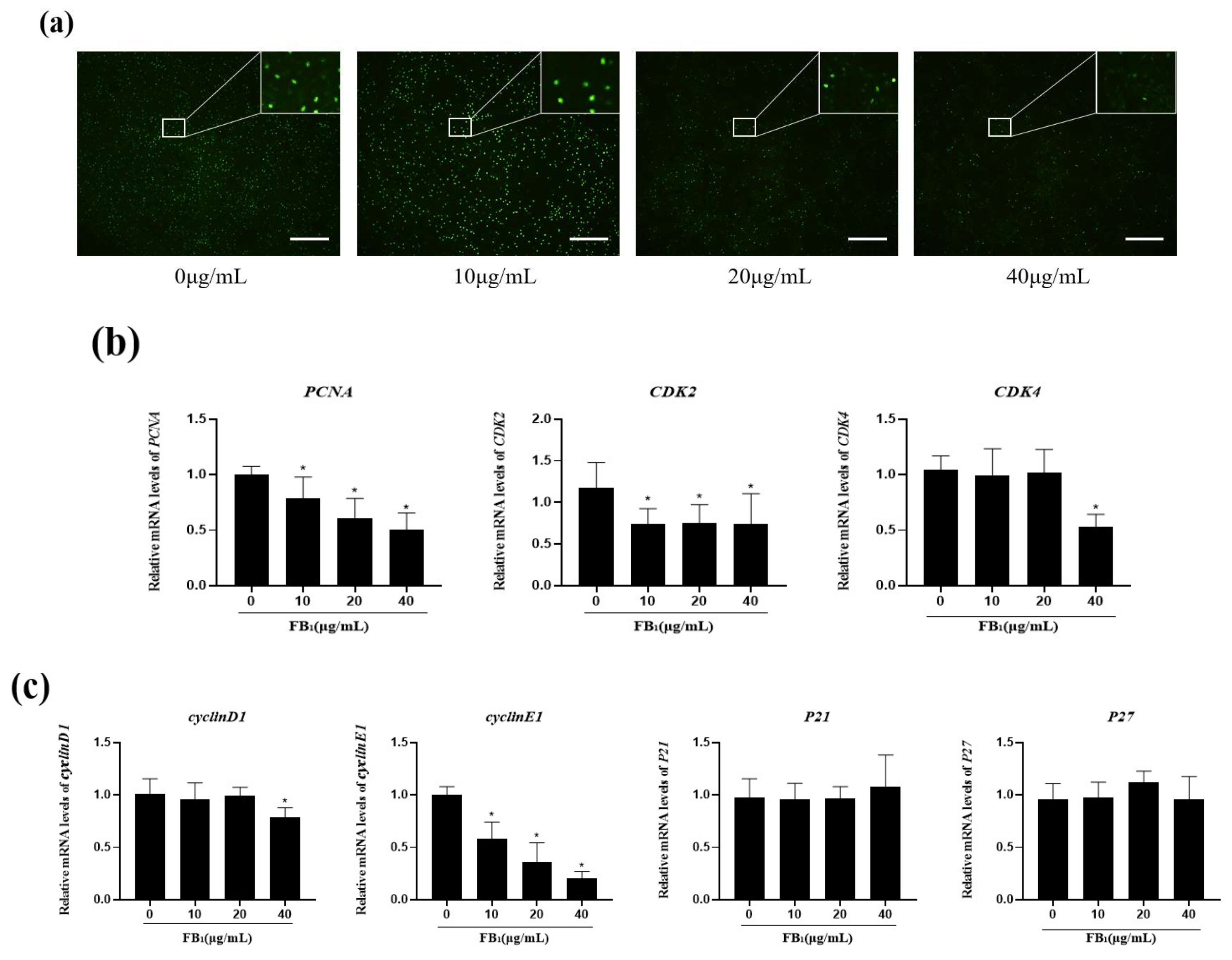
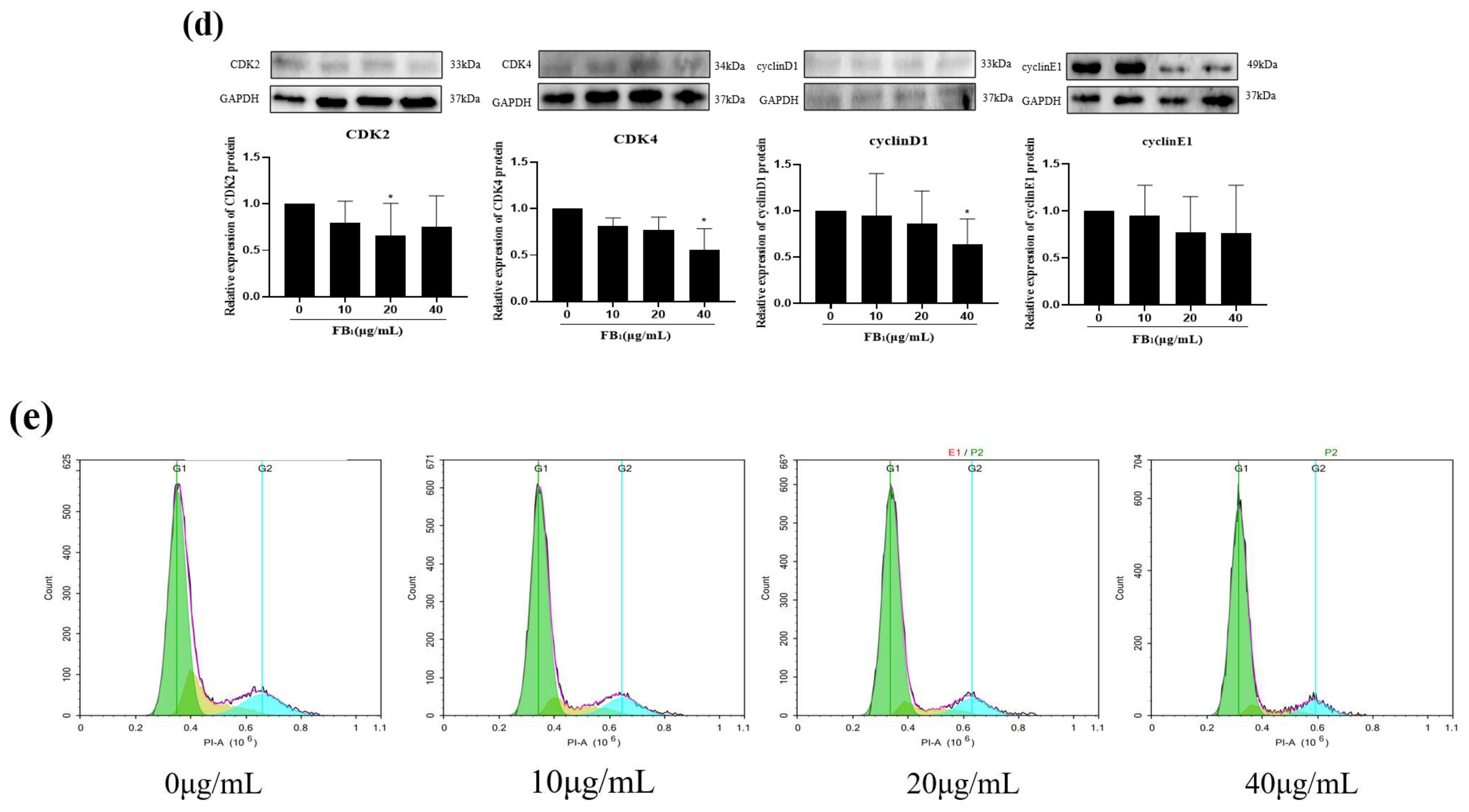
2.2. FB1 Decreased Cell Proliferation and Arrested the Cell Cycle
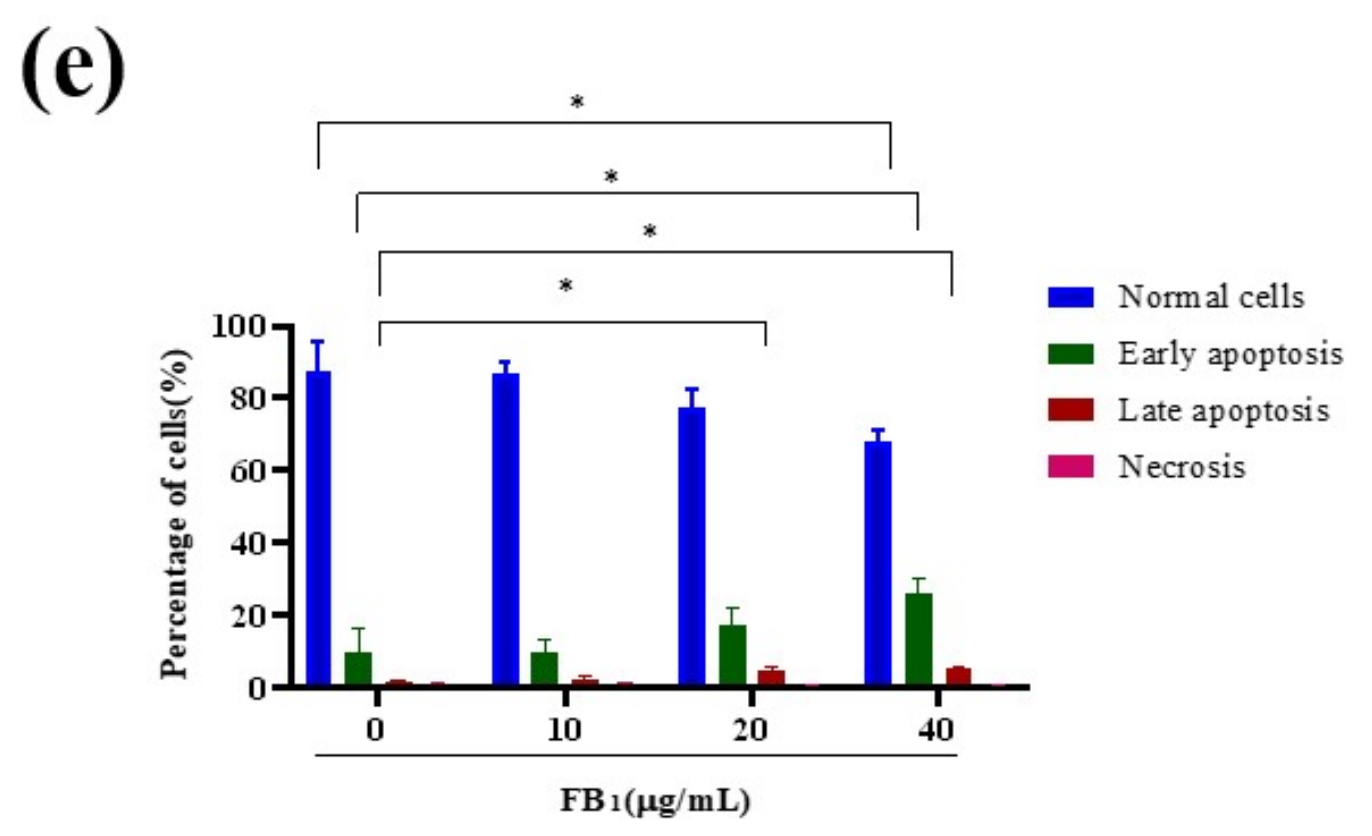
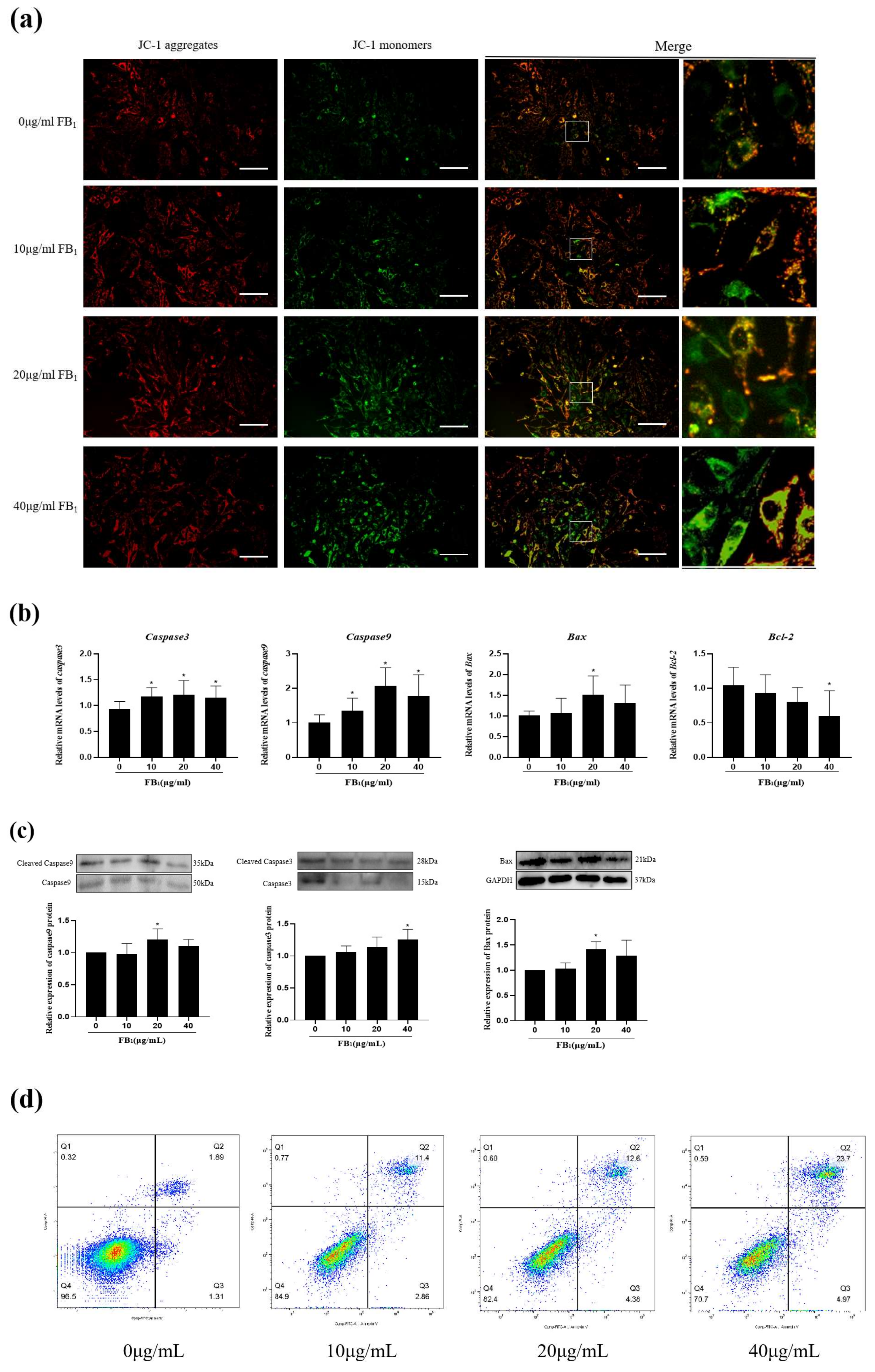
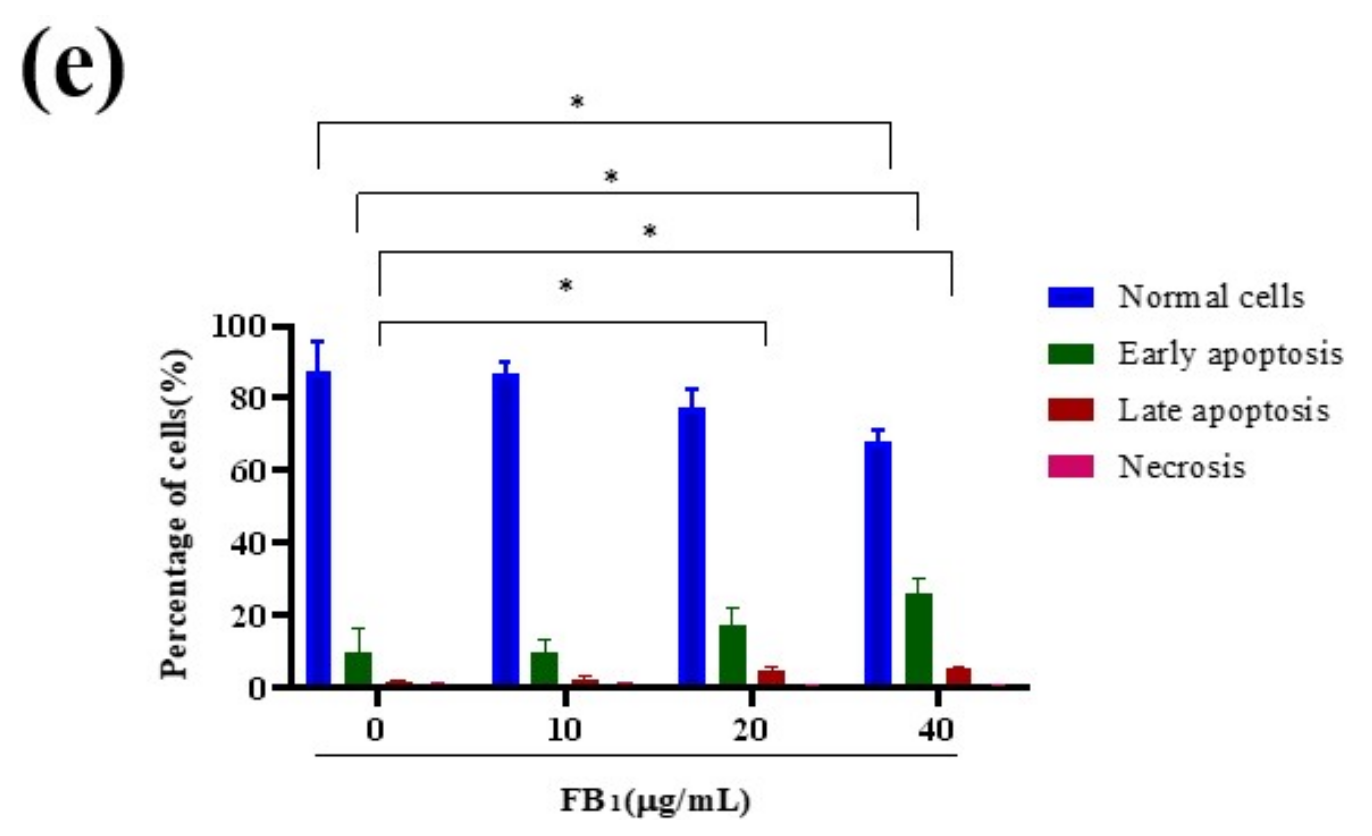
2.3. FB1 Reduced Mitochondrial Membrane Potential and Promoted Cell Apoptosis
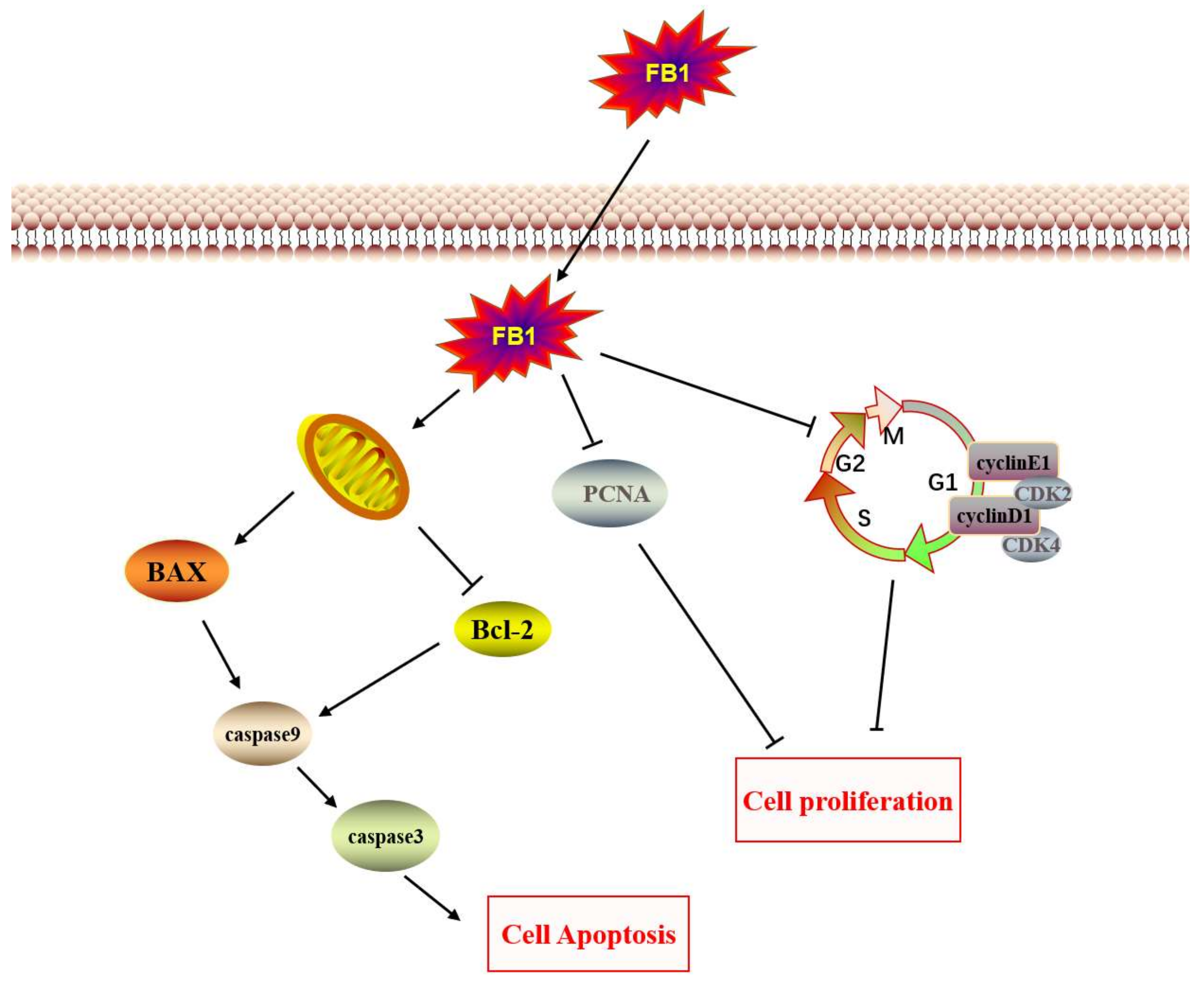
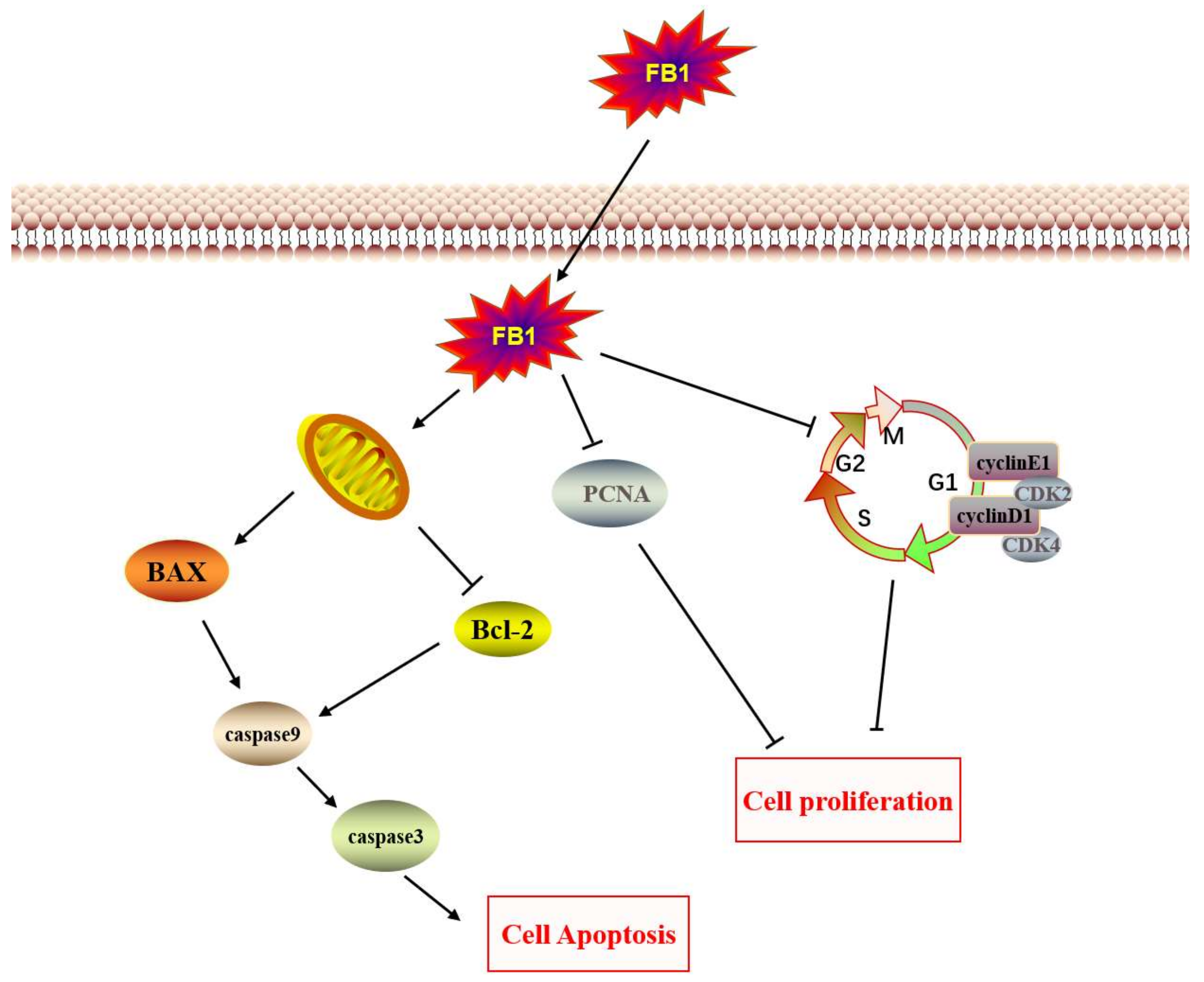
3. Discussion
4. Conclusions
5. Materials and Methods
5.1. Cells and Cell Culture
5.2. Cell Viability Assay
5.3. Observation of Cell Morphology
5.4. Cell Proliferation Assay
5.5. RT-qPCR Analysis
5.6. Western Blot
5.7. Cell Cycle Assay
5.8. Cell Apoptosis Assay
5.9. ∆ψm Determination
5.10. Statistical Analysis
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Yang, C.; Song, G.; Lim, W. Effects of mycotoxin-contaminated feed on farm animals. J. Hazard. Mater. 2020, 389, 122087. [Google Scholar] [CrossRef] [PubMed]
- Cao, C.; Zhu, X.; Li, X.; Ouyang, H.; Wang, K.; Li, X. Assessment of ionic homeostasis imbalance and cytochrome P450 system disturbance in mice during fumonisin B1 (FB1) exposure. Chemosphere 2020, 251, 126393. [Google Scholar] [CrossRef] [PubMed]
- Mirón-Mérida, V.A.; González-Espinosa, Y.; Collado-González, M.; Gong, Y.Y.; Guo, Y.; Goycoolea, F.M. Aptamer–target–gold nanoparticle conjugates for the quantification of fumonisin B1. Biosensors 2021, 11, 18. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Wu, Q.; Wan, D.; Liu, Q.; Chen, D.; Liu, Z.; Martínez-Larrañaga, M.R.; Martínez, M.A.; Anadón, A.; Yuan, Z. Fumonisins: Oxidative stress-mediated toxicity and metabolism in vivo and in vitro. Arch. Toxicol. 2016, 90, 81–101. [Google Scholar] [CrossRef] [PubMed]
- Bouhet, S.; Oswald, I.P. The intestine as a possible target for fumonisin toxicity. Mol. Nutr. Food Res. 2007, 51, 925–931. [Google Scholar] [CrossRef] [PubMed]
- Prelusky, D.B.; Trenholm, H.L.; Rotter, B.A.; Miller, J.D.; Savard, M.E.; Yeung, J.M.; Scott, P.M. Biological fate of fumonisin B1 in food-producing animals. Adv. Exp. Med. Biol. 1996, 392, 265–278. [Google Scholar] [PubMed]
- Bouhet, S.; Hourcade, E.; Loiseau, N.; Fikry, A.; Martinez, S.; Roselli, M.; Galtier, P.; Mengheri, E.; Oswald, I.P. The Mycotoxin fumonisin B1 alters the proliferation and the barrier function of porcine intestinal epithelial cells. Toxicol. Sci. 2003, 77, 165–171. [Google Scholar] [CrossRef] [Green Version]
- Gao, S.; Liu, F. Novel insights into cell cycle regulation of cell fate determination. J. Zhejiang Univ. Sci. B 2019, 20, 467–475. [Google Scholar] [CrossRef]
- Orford, K.W.; Scadden, D.T. Deconstructing stem cell self-renewal: Genetic insights into cell-cycle regulation. Nat. Rev. Genet. 2008, 9, 115–128. [Google Scholar] [CrossRef]
- Strzalka, W.; Ziemienowicz, A. Proliferating cell nuclear antigen (PCNA): A key factor in DNA replication and cell cycle regulation. Ann. Bot. 2011, 10, 1127–1140. [Google Scholar] [CrossRef] [Green Version]
- Gali-Muhtasib, H.; Bakkar, N. Modulating cell cycle: Current applications and prospects for future drug development. Curr. Cancer Drug Targets 2002, 2, 309–336. [Google Scholar] [CrossRef]
- Wang, Z.; Wang, Y.; Wang, S.; Meng, X.; Song, F.; Huo, W.; Zhang, S.; Chang, J.; Li, J.; Zheng, B.; et al. Coxsackievirus A6 induces cell cycle arrest in G0/G1 phase for viral production. Front. Cell. Infect. Microbiol. 2018, 8, 279. [Google Scholar] [CrossRef] [Green Version]
- Gao, X.; Leone, G.W.; Wang, H. Cyclin D-CDK4/6 functions in cancer. Adv. Cancer Res. 2020, 148, 147–169. [Google Scholar]
- Li, Q.; Yuan, Q.; Wang, T.; Zhan, Y.; Yang, L.; Fan, Y.; Lei, H.; Su, J. Fumonisin B1 inhibits cell proliferation and decreases barrier function of swine umbilical vein endothelial cells. Toxins 2021, 13, 863. [Google Scholar] [CrossRef]
- Gudipaty, S.A.; Conner, C.M.; Rosenblatt, J.; Montell, D.J. Unconventional ways to live and die: Cell death and survival in development, homeostasis, and disease. Annu. Rev. Cell Dev. Biol. 2018, 34, 311–332. [Google Scholar] [CrossRef]
- Martin, S.J.; Green, D.R. Protease activation during apoptosis: Death by a thousand cuts? Cell 1995, 82, 349–352. [Google Scholar] [CrossRef] [Green Version]
- Del Re, D.P.; Amgalan, D.; Linkermann, A.; Liu, Q.; Kitsis, R.N. Fundamental mechanisms of regulated cell death and implications for heart disease. Physiol. Rev. 2019, 99, 1765–1817. [Google Scholar] [CrossRef]
- Chen, Z.; Zhou, L.; Yuan, Q.; Chen, H.; Lei, H.; Su, J. Effect of fumonisin B 1 on oxidative stress and gene expression alteration of nutrient transporters in porcine intestinal cells. J. Biochem. Mol. Toxicol. 2021, 35, e22706. [Google Scholar] [CrossRef]
- Chen, Z.; Chen, H.; Li, X.; Yuan, Q.; Su, J.; Yang, L.; Ning, L.; Lei, H. Fumonisin B1 damages the barrier functions of porcine intestinal epithelial cells in vitro. J. Biochem. Mol. Toxicol. 2019, 33, e22397. [Google Scholar] [CrossRef]
- Reisinger, N.; Schürer-Waldheim, S.; Mayer, E.; Debevere, S.; Antonissen, G.; Sulyok, M.; Nagl, V. Mycotoxin occurrence in maize silage—A neglected risk for bovine gut health? Toxins 2019, 11, 577. [Google Scholar] [CrossRef] [Green Version]
- Akbari, P.; Braber, S.; Varasteh, S.; Alizadeh, A.; Garssen, J.; Fink-Gremmels, J. The intestinal barrier as an emerging target in the toxicological assessment of mycotoxins. Arch. Toxicol. 2017, 91, 1007–1029. [Google Scholar] [CrossRef] [Green Version]
- Sun, Y.; Liu, Y.; Ma, X.; Hu, H. The influence of cell cycle regulation on chemotherapy. Int. J. Mol. Sci. 2021, 22, 6923. [Google Scholar] [CrossRef]
- Clark, A.S.; Karasic, T.B.; DeMichele, A.; Vaughn, D.J.; O’Hara, M.; Perini, R.; Zhang, P.; Lal, P.; Feldman, M.; Gallagher, M.; et al. Palbociclib (PD0332991)—A selective and potent cyclin-dependent kinase inhibitor: A review of pharmacodynamics and clinical development. JAMA Oncol. 2016, 2, 253–260. [Google Scholar] [CrossRef]
- Diehl, J.A. Cycling to cancer with cyclin D1. Cancer Biol. Ther. 2002, 1, 226–231. [Google Scholar] [CrossRef] [Green Version]
- Xia, P.; Liu, Y.; Chen, J.; Cheng, Z. Cell cycle proteins as key regulators of postmitotic cell death. Yale J. Biol. Mol. 2019, 92, 641–650. [Google Scholar]
- Taranu, I.; Marin, D.E.; Burlacu, R.; Pinton, P.; Damian, V.; Oswald, I.P. Comparative aspects of in vitro proliferation of human and porcine lymphocytes exposed to mycotoxins. Arch. Anim. Nutr. 2010, 64, 383–393. [Google Scholar] [CrossRef]
- Shin, K.-T.; Guo, J.; Niu, Y.-J.; Cui, X.-S. The toxic effect of aflatoxin B1 on early porcine embryonic development. Theriogenology 2018, 118, 157–163. [Google Scholar] [CrossRef]
- Diesing, A.K.; Nossol, C.; Panther, P.; Walk, N.; Post, A.; Kluess, J.; Kreutzmann, P.; Dänicke, S.; Rothkötter, H.J.; Kahlert, S. Mycotoxin deoxynivalenol (DON) mediates biphasic cellular response in intestinal porcine epithelial cell lines IPEC-1 and IPEC-J2. Toxicol. Lett. 2011, 200, 8–18. [Google Scholar] [CrossRef]
- Cheng, X.; Ferrell, J.E. Apoptosis propagates through the cytoplasm as trigger waves. Science 2018, 361, 607–612. [Google Scholar] [CrossRef] [Green Version]
- Schwabe, R.F.; Luedde, T. Apoptosis and necroptosis in the liver: A matter of life and death. Nat. Rev. Gastroenterol. Hepatol. 2018, 15, 738–752. [Google Scholar] [CrossRef]
- Czabotar, P.E.; Lessene, G.; Strasser, A.; Adams, J.M. Control of apoptosis by the BCL-2 protein family: Implications for physiology and therapy. Mol. Cell Biol. 2014, 15, 49–63. [Google Scholar] [CrossRef] [PubMed]
- Song, Y.; Liu, W.; Zhao, Y.; Zang, J.; Gao, H. Fumonisin B1 exposure induces apoptosis of human kidney tubular epithelial cells through regulating PTEN/PI3K/AKT signaling pathway via disrupting lipid raft formation. Toxicon 2021, 204, 31–36. [Google Scholar] [CrossRef] [PubMed]
- Paul, S.; Jakhar, R.; Bhardwaj, M.; Chauhan, A.K.; Kang, S.C. Fumonisin B1 induces poly (ADP-ribose) (PAR) polymer-mediated cell death (parthanatos) in neuroblastoma. Food Chem. Toxicol. 2021, 154, 112026. [Google Scholar] [CrossRef] [PubMed]
- Cao, C.; Xian, R.; Lin, F.; Li, X.; Li, X.; Qiang, F.; Li, X. Fumonisin B1 induces hepatotoxicity in mice through the activation of oxidative stress, apoptosis and fibrosis. Chemosphere 2022, 296, 133910. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Primer Name | Primer Sequences (5′-3′) | Primer Sequences |
|---|---|---|
| P21 | F: 5′-3′ACCCCTTCCCCATACCC | XM_013977858.2 |
| R: 5′-3′TTCCTAACACCCATGAAACTG | ||
| P27 | F: 5′-3′GTCCCTTTCAGTGAGAACCG ATAC | NM_214316.1 |
| R: 5′-3′TTGCTGCCACATAACGGAATCAT | ||
| PCNA | F: 5′-3′GTGATTCCACCACCATGTTC | NM_001291925.1 |
| R: 5′-3′TGAGACGACTCCATGCTCTG | ||
| cyclin D1 | F: 5′-3′GCGAGGAACAGAAGTGCG | XM_021082686.1 |
| R: 5′-3′TGGAGTTGTCGGTGTAGATGC | ||
| cyclin E1 | F: 5′-3′CTCGCCACTGCCTATACTGA | XM_005653265.2 |
| R: 5′-3′GGTGCCGCTGCATAAGGT | ||
| CDK2 | F: 5′-3′GCGAGGAACAGAAGTGCG | XM_013977858.2 |
| R: 5′-3′TGGAGTTGTCGGTGTAGATGC | ||
| CDK4 | F: 5′-3′GCGGAGATTGGTGTTGGTG | NM_001123097.1 |
| R: 5′-3′CATTGGGGACTCTTACGCTCTT | ||
| Caspase3 | F: 5′-3′TGCTGCAAATCTCAGGGAGACCT | NM_214131.1 |
| R: 5′-3′GTGCCTCGGCAGGCCTGAAT | ||
| Caspase9 | F: 5′-3′TGGCCTCGCTCTGGGATGCT | NM_02107526.7 |
| R: 5′-3′TGGCCTCGCTCTGGGATGCT | ||
| Bcl-2 | F: 5′-3′CTGCGAACCCGGTCTGCCTG | XM_005664627.3 |
| R: 5′-3′TCTCGGGCCCACTGCTCCTC | ||
| Bax | F: 5′-3′CCGAGTGGCGGCCGAAATGT | XM_013998624.2 |
| R: 5′-3′TCCAGCCCAGCAGCCGATCTG | ||
| GAPDH | F: 5′-3′GTGATTCCACCACCATGTTC | XM_021091114.1 |
| R: 5′-3′TGAGACGACTCCATGCTCTG |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, T.; Lei, H.; Zhou, L.; Tang, M.; Liu, Q.; Long, F.; Li, Q.; Su, J. Effect of Fumonisin B1 on Proliferation and Apoptosis of Intestinal Porcine Epithelial Cells. Toxins 2022, 14, 471. https://doi.org/10.3390/toxins14070471
Wang T, Lei H, Zhou L, Tang M, Liu Q, Long F, Li Q, Su J. Effect of Fumonisin B1 on Proliferation and Apoptosis of Intestinal Porcine Epithelial Cells. Toxins. 2022; 14(7):471. https://doi.org/10.3390/toxins14070471
Chicago/Turabian StyleWang, Tianjie, Hongyu Lei, Lihua Zhou, Meiwen Tang, Qing Liu, Feng Long, Qing Li, and Jianming Su. 2022. "Effect of Fumonisin B1 on Proliferation and Apoptosis of Intestinal Porcine Epithelial Cells" Toxins 14, no. 7: 471. https://doi.org/10.3390/toxins14070471
APA StyleWang, T., Lei, H., Zhou, L., Tang, M., Liu, Q., Long, F., Li, Q., & Su, J. (2022). Effect of Fumonisin B1 on Proliferation and Apoptosis of Intestinal Porcine Epithelial Cells. Toxins, 14(7), 471. https://doi.org/10.3390/toxins14070471