Fusarium culmorum Produces NX-2 Toxin Simultaneously with Deoxynivalenol and 3-Acetyl-Deoxynivalenol or Nivalenol


 , and
, and 
Abstract
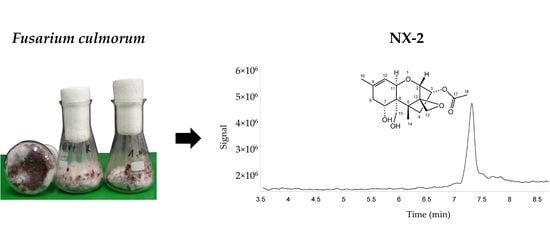
1. Introduction
2. Results
2.1. Mycotoxin Production in Rice Cultures
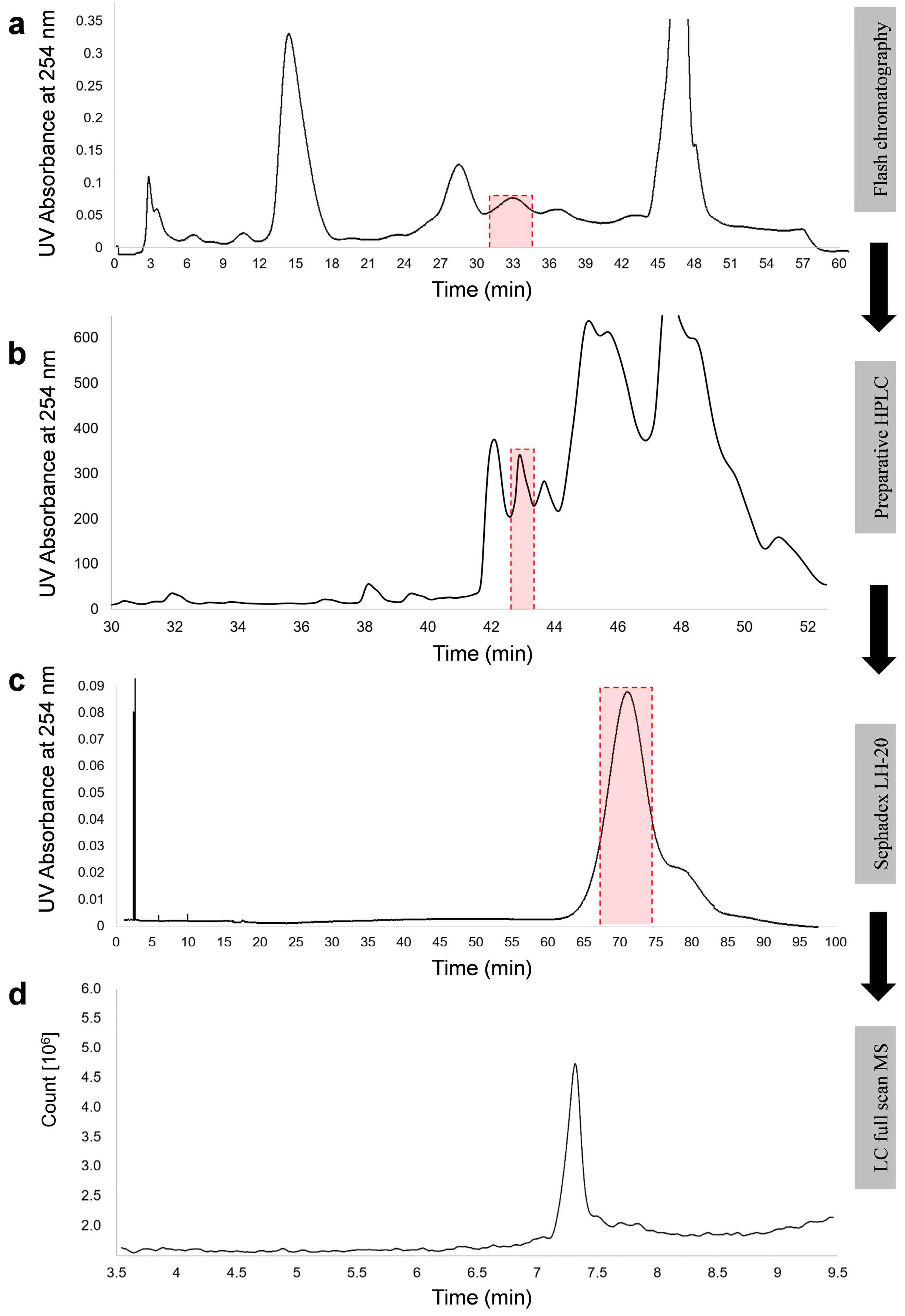
2.2. Confirmation of the Structure of NX-2
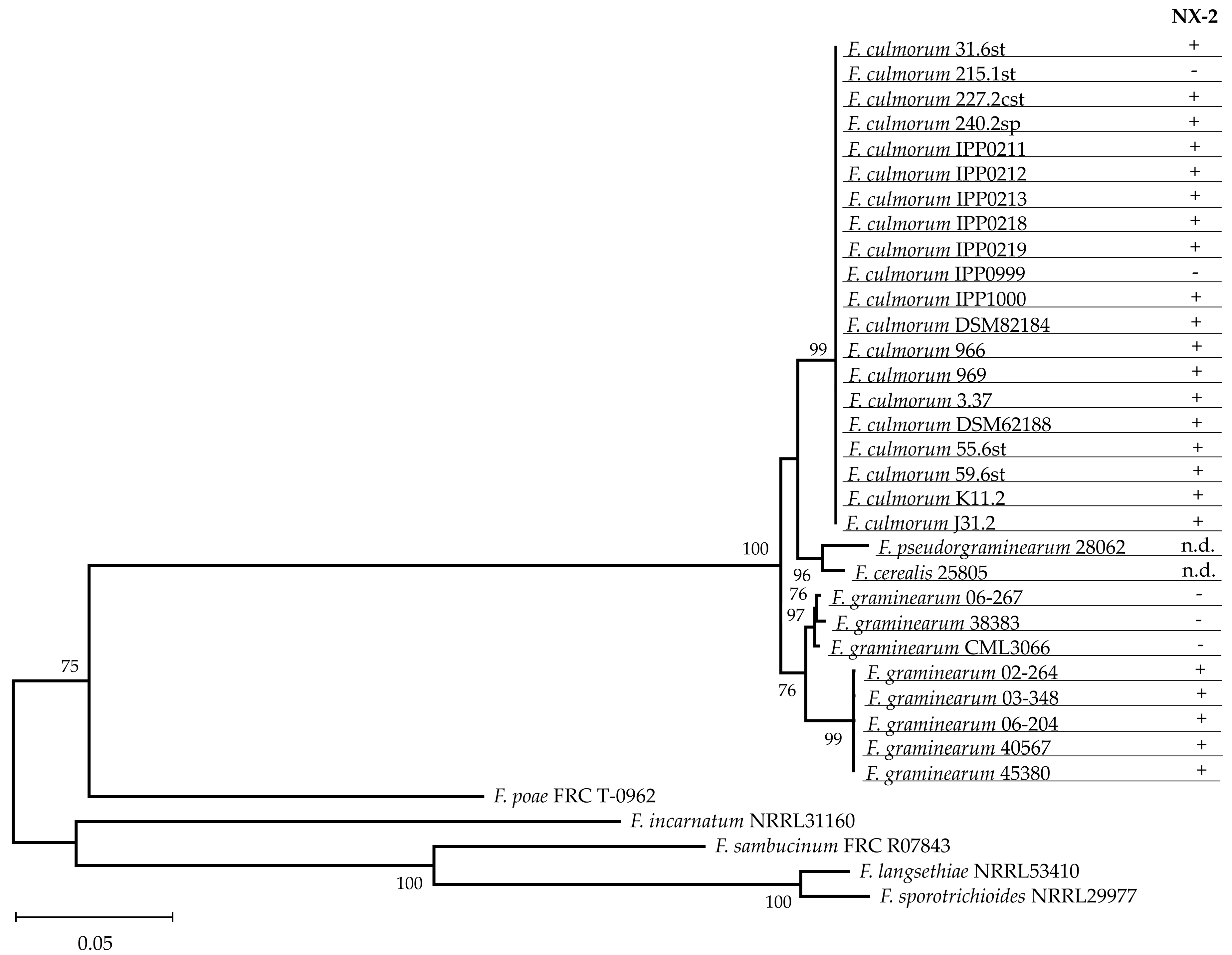
2.3. Species Assignment and Investigation of Polymorphisms in TRI1

3. Discussion
3.1. Production of NX-2 Is a Characteristic Feature of F. culmorum
3.2. Both Chemotypes of F. culmorum Produce NX-2
3.3. Fusarium culmorum Produces NX-2 Simultaneously with DON and 3-ADON or NIV
3.4. NX-2 Production by F. culmorum Is not Caused by a Variant of TRI1
3.5. NX-2 and the Aggressiveness of F. graminearum and F. culmorum
3.6. Do further Fusarium Species Produce NX Toxins?
3.7. Can F. culmorum Produce DON and NIV Simultaneously?
4. Conclusions
5. Materials and Methods
5.1. Fungal Strains

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Species | Origin | Year | Isolate | Host |
|---|---|---|---|---|
| F. culmorum | Germany | 2017 | 31.6st | Maize stalk |
| F. culmorum | Germany | 2018 | 215.1st | Maize stalk |
| F. culmorum | Germany | 2018 | 227.2cst | Maize stalk |
| F. culmorum | Germany | 2018 | 240.2sp | Maize rachis |
| F. culmorum | - | <1984 a | IPP0211 b | Wheat |
| F. culmorum | Italy | <1993 a | IPP0212 b | Wheat |
| F. culmorum | - | 1993 | IPP0213 b | Barley |
| F. culmorum | Hungary | 1991 | IPP0618 b | Wheat |
| F. culmorum | Germany | 1991 | IPP0619 b | Wheat |
| F. culmorum | Germany | 2010 | IPP0999 b | Wheat |
| F. culmorum | Germany | 2010 | IPP1000 b | Wheat |
| F. culmorum | Germany | 1952 | DSM62184 c | Maize grain |
| F. culmorum | Syria | 2009–2010 | 966 d | Wheat |
| F. culmorum | Syria | 2009–2010 | 969 d | Wheat |
| F. culmorum | Germany | 2004 | 3.37 b | Wheat |
| F. culmorum | Germany | 1990 | DSM62188 c | Maize stalk |
| F. culmorum | Germany | 2017 | 55.6st | Maize stalk |
| F. culmorum | Germany | 2017 | 59.6st | Maize stalk |
| F. culmorum | Germany | 2021 | K11.2 | Oat |
| F. culmorum | Germany | 2021 | J31.2 | Oat |
5.2. DNA Methods
5.3. Mycotoxin Extraction and HPLC-MS/MS
5.4. Purification of NX-2
5.5. Nuclear Magnetic Resonance (NMR) Spectroscopy
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Parry, D.W.; Jenkinson, P.; McLeod, L. Fusarium ear blight (scab) in small grain cereals—A review. Plant Pathol. 1995, 44, 207–238. [Google Scholar] [CrossRef]
- Birr, T.; Hasler, M.; Verreet, J.-A.; Klink, H. Composition and predominance of Fusarium species causing fusarium head blight in winter wheat grain depending on cultivar susceptibility and meteorological factors. Microorganisms 2020, 8, 617. [Google Scholar] [CrossRef] [PubMed]
- Foroud, N.A.; Baines, D.; Gagkaeva, T.Y.; Thakor, N.; Badea, A.; Steiner, B.; Bürstmayr, M.; Bürstmayr, H. Trichothecenes in cereal grains—An update. Toxins 2019, 11, 634. [Google Scholar] [CrossRef] [PubMed]
- Polak-Śliwińska, M.; Paszczyk, B. Trichothecenes in food and feed, relevance to human and animal health and methods of detection: A systematic review. Molecules 2021, 26, 454. [Google Scholar] [CrossRef]
- O’Donnell, K.; Rooney, A.P.; Proctor, R.H.; Brown, D.W.; McCormick, S.P.; Ward, T.J.; Frandsen, R.J.N.; Lysøe, E.; Rehner, S.A.; Aoki, T.; et al. Phylogenetic analyses of RPB1 and RPB2 support a middle Cretaceous origin for a clade comprising all agriculturally and medically important Fusaria. Fungal Genet. Biol. 2013, 52, 20–31. [Google Scholar] [CrossRef] [PubMed]
- Scherm, B.; Balmas, V.; Spanu, F.; Pani, G.; Delogu, G.; Pasquali, M.; Migheli, Q. Fusarium culmorum: Causal agent of foot and root rot and head blight on wheat. Mol. Plant Pathol. 2013, 14, 323–341. [Google Scholar] [CrossRef] [PubMed]
- Logrieco, A.; Mulè, G.; Moretti, A.; Bottalico, A. Toxigenic Fusarium species and mycotoxins associated with maize ear rot in Europe. Eur. J. Plant Pathol. 2002, 108, 597–609. [Google Scholar] [CrossRef]
- Pfordt, A.; Ramos Romero, L.; Schiwek, S.; Karlovsky, P.; von Tiedemann, A. Impact of environmental conditions and agronomic practices on the prevalence of Fusarium species associated with ear- and stalk rot in maize. Pathogens 2020, 9, 236. [Google Scholar] [CrossRef]
- Basler, R. Diversity of Fusarium species isolated from UK forage maize and the population structure of F. graminearum from maize and wheat. PeerJ 2016, 4, 2143. [Google Scholar] [CrossRef]
- Sundheim, L.; Brodal, G.; Hofgaard, I.S.; Rafoss, T. Temporal variation of mycotoxin producing fungi in Norwegian cereals. Microorganisms 2013, 1, 188–198. [Google Scholar] [CrossRef]
- Nielsen, L.K.; Jensen, J.D.; Nielsen, G.C.; Jensen, J.E.; Spliid, N.H.; Thomsen, I.K.; Justesen, A.F.; Collinge, D.B.; Jørgensen, L.N. Fusarium head blight of cereals in Denmark: Species complex and related mycotoxins. Phytopathology 2011, 101, 960–969. [Google Scholar] [CrossRef]
- Ropejko, K.; Twarużek, M. Zearalenone and its metabolites—General overview, occurrence, and toxicity. Toxins 2021, 13, 35. [Google Scholar] [CrossRef] [PubMed]
- McCormick, S.P.; Stanley, A.M.; Stover, N.A.; Alexander, N.J. Trichothecenes: From simple to complex mycotoxins. Toxins 2011, 3, 802–814. [Google Scholar] [CrossRef] [PubMed]
- Villafana, R.T.; Ramdass, A.C.; Rampersad, S.N. TRI genotyping and chemotyping: A balance of power. Toxins 2020, 12, 64. [Google Scholar] [CrossRef] [PubMed]
- van der Lee, T.; Zhang, H.; van Diepeningen, A.; Waalwijk, C. Biogeography of Fusarium graminearum species complex and chemotypes: A review. Food Addit. Contam. 2015, 32, 453–460. [Google Scholar] [CrossRef]
- Crippin, T.; Renaud, J.B.; Sumarah, M.W.; Miller, J.D. Comparing genotype and chemotype of Fusarium graminearum from cereals in Ontario, Canada. PLoS ONE 2019, 14, e0216735. [Google Scholar] [CrossRef]
- Gale, L.R.; Ward, T.J.; Balmas, V.; Kistler, H.C. Population subdivision of Fusarium graminearum sensu stricto in the upper midwestern United States. Phytopathology 2007, 97, 1434–1439. [Google Scholar] [CrossRef]
- Gilbert, J.; Clear, R.M.; Ward, T.J.; Gaba, D.; Tekauz, A.; Turkington, T.K.; Woods, S.M.; Nowicki, T.; O’Donnell, K. Relative aggressiveness and production of 3- or 15-acetyl deoxynivalenol and deoxynivalenol by Fusarium graminearum in spring wheat. Can. J. Plant Pathol. 2010, 32, 146–152. [Google Scholar] [CrossRef]
- von der Ohe, C.; Gauthier, V.; Tamburic-Ilincic, L.; Brule-Babel, A.; Fernando, W.G.D.; Clear, R.; Ward, T.J.; Miedaner, T. A comparison of aggressiveness and deoxynivalenol production between Canadian Fusarium graminearum isolates with 3-acetyl and 15-acetyldeoxynivalenol chemotypes in field-grown spring wheat. Eur. J. Plant Pathol. 2010, 127, 407–417. [Google Scholar] [CrossRef]
- Puri, K.D.; Zhong, S. The 3ADON population of Fusarium graminearum found in North Dakota is more aggressive and produces a higher level of DON than the prevalent 15ADON population in spring wheat. Phytopathology 2010, 100, 1007–1014. [Google Scholar] [CrossRef]
- Malihipour, A.; Gilbert, J.; Piercey-Normore, M.; Cloutier, S. Molecular phylogenetic analysis, trichothecene chemotype patterns, and variation in aggressiveness of Fusarium isolates causing head blight in wheat. Plant Dis. 2012, 96, 1016–1025. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.Y.; Sun, H.Y.; Li, W.; Xia, Y.L.; Deng, Y.Y.; Zhang, A.X.; Chen, H.G. Fitness of three chemotypes of Fusarium graminearum species complex in major winter wheat-producing areas of China. PLoS ONE 2017, 12, e0174040. [Google Scholar] [CrossRef] [PubMed]
- Amarasinghe, C.; Sharanowski, B.; Dilantha Fernando, W.G. Molecular phylogenetic relationships, trichothecene chemotype diversity and aggressiveness of strains in a global collection of Fusarium graminearum species. Toxins 2019, 11, 263. [Google Scholar] [CrossRef]
- Poppenberger, B.; Berthiller, F.; Lucyshyn, D.; Sieberer, T.; Schuhmacher, R.; Krska, R.; Kuchler, K.; Glössl, J.; Luschnig, C.; Adam, G. Detoxification of the Fusarium mycotoxin deoxynivalenol by a UDP-glucosyltransferase from Arabidopsis thaliana. J. Biol. Chem. 2003, 278, 47905–47914. [Google Scholar] [CrossRef] [PubMed]
- Pasquali, M.; Beyer, M.; Logrieco, A.; Audenaert, K.; Balmas, V.; Basler, R.; Boutigny, A.L.; Chrpová, J.; Czembor, E.; Gagkaeva, T.; et al. A European database of Fusarium graminearum and F. culmorum trichothecene genotypes. Front. Microbiol. 2016, 7, 406. [Google Scholar] [CrossRef]
- Jennings, P.; Coates, M.E.; Turner, J.A.; Chandler, E.A.; Nicholson, P. Determination of deoxynivalenol and nivalenol chemotypes of Fusarium culmorum isolates from England and Wales by PCR assay. Plant Pathol. 2004, 53, 182–190. [Google Scholar] [CrossRef]
- Quarta, A.; Mita, G.; Haidukowski, M.; Santino, A.; Mulè, G.; Visconti, A. Assessment of trichothecene chemotypes of Fusarium culmorum occurring in Europe. Food Addit. Contam. 2005, 22, 309–315. [Google Scholar] [CrossRef]
- Matny, O.N.; Bates, S.T.; Song, Z. Geographic distribution of Fusarium culmorum chemotypes associated with wheat crown rot in Iraq. J. Plant. Prot. Res. 2017, 57, 43–49. [Google Scholar] [CrossRef]
- Kulik, T.; Buśko, M.; Bilska, K.; Ostrowska-Kołodziejczak, A.; Van Diepeningen, A.D.; Perkowski, J.; Stenglein, S. Depicting the discrepancy between TRI genotype and chemotype on the basis of strain CBS 139514 from a field population of F. graminearum sensu stricto from Argentina. Toxins 2016, 8, 330. [Google Scholar] [CrossRef]
- Varga, E.; Wiesenberger, G.; Hametner, C.; Ward, T.J.; Dong, Y.; Schöfbeck, D.; Mccormick, S.; Broz, K.; Stückler, R.; Schuhmacher, R.; et al. New tricks of an old enemy: Isolates of Fusarium graminearum produce a type A trichothecene mycotoxin. Environ. Microbiol. 2015, 17, 2588–2600. [Google Scholar] [CrossRef]
- Laraba, I.; McCormick, S.P.; Vaughan, M.M.; Geiser, D.M.; O’Donnell, K. Phylogenetic diversity, trichothecene potential, and pathogenicity within Fusarium sambucinum species complex. PLoS ONE 2021, 16, e0250812. [Google Scholar] [CrossRef] [PubMed]
- Lofgren, L.; Riddle, J.; Dong, Y.; Kuhnem, P.R.; Cummings, J.A.; Del Ponte, E.M.; Bergstrom, G.C.; Kistler, H.C. A high proportion of NX-2 genotype strains are found among Fusarium graminearum isolates from northeastern New York State. Eur. J. Plant Pathol. 2018, 150, 791–796. [Google Scholar] [CrossRef]
- Gale, L.R.; Ward, T.J.; Kistler, H.C. A subset of the newly discovered Northland population of Fusarium graminearum from the U.S. does not produce the B-type trichothecenes DON, 15ADON, 3ADON or NIV. In Proceedings of the National Fusarium Head Blight Forum, Milwaukee, WI, USA, 7–9 December 2010; pp. 48–49. [Google Scholar]
- Schiwek, S.; Beule, L.; Vinas, M.; Von Tiedemann, A.; Karlovsky, P. High-resolution melting (HRM) curve assay for the identification of eight Fusarium species causing ear rot in maize. Pathogens 2020, 9, 270. [Google Scholar] [CrossRef]
- Kelly, A.; Proctor, R.H.; Belzile, F.; Chulze, S.N.; Clear, R.M.; Cowger, C.; Elmer, W.; Lee, T.; Obanor, F.; Waalwijk, C.; et al. The geographic distribution and complex evolutionary history of the NX-2 trichothecene chemotype from Fusarium graminearum. Fungal Genet. Biol. 2016, 95, 39–48. [Google Scholar] [CrossRef]
- Kumar, S.; Stecher, G.; Li, M.; Knyaz, C.; Tamura, K. MEGA X: Molecular evolutionary genetics analysis across computing platforms. Mol. Biol. Evol. 2018, 35, 1547–1549. [Google Scholar] [CrossRef] [PubMed]
- Tamura, K.; Nei, M. Estimation of the number of nucleotide substitutions in the control region of mitochondrial DNA in humans and chimpanzees. Mol. Biol. Evol. 1993, 10, 512–526. [Google Scholar] [CrossRef]
- Goswami, R.S.; Kistler, H.C. Heading for Disaster: Fusarium graminearum on cereal crops. Mol. Plant Pathol. 2004, 5, 515–525. [Google Scholar] [CrossRef]
- Oufensou, S.; Balmas, V.; Scherm, B.; Rau, D.; Camilio, S.; Prota, V.A.; Ben-Attia, M.; Gargouri, S.; Pasquali, M.; El-Bok, S. Genetic variability, chemotype distribution, and aggressiveness of Fusarium culmorum on durum wheat in Tunisia. Phytopathol. Mediterr. 2019, 58, 103–113. [Google Scholar] [CrossRef]
- Alkadri, D.; Nipoti, P.; Döll, K.; Karlovsky, P.; Prodi, A.; Pisi, A. Study of fungal colonization of wheat kernels in Syria with a focus on Fusarium species. Int. J. Mol. Sci. 2013, 14, 5938–5951. [Google Scholar] [CrossRef]
- Kelly, A.C.; Clear, R.M.; O’Donnell, K.; McCormick, S.; Turkington, T.K.; Tekauz, A.; Gilbert, J.; Kistler, H.C.; Busman, M.; Ward, T.J. Diversity of Fusarium head blight populations and trichothecene toxin types reveals regional differences in pathogen composition and temporal dynamics. Fungal Genet. Biol. 2015, 82, 22–31. [Google Scholar] [CrossRef]
- Pasquali, M.; Beyer, M.; Bohn, T.; Hoffmann, L. Comparative analysis of genetic chemotyping methods for Fusarium: Tri13 polymorphism does not discriminate between 3- and 15-acetylated deoxynivalenol chemotypes in Fusarium graminearum. J. Phytopathol. 2011, 159, 700–704. [Google Scholar] [CrossRef]
- Proctor, R.H.; McCormick, S.P.; Kim, H.-S.; Cardoza, R.E.; Stanley, A.M.; Lindo, L.; Kelly, A.; Brown, D.W.; Lee, T.; Vaughan, M.M.; et al. Evolution of structural diversity of trichothecenes, a family of toxins produced by plant pathogenic and entomopathogenic fungi. PLoS Pathog. 2018, 14, e1006946. [Google Scholar] [CrossRef] [PubMed]
- McCormick, S.P.; Harris, L.J.; Alexander, N.J.; Ouellet, T.; Saparno, A.; Allard, S.; Desjardins, A.E. Tri1 in Fusarium graminearum encodes a P450 oxygenase. Appl. Environ. Microbiol. 2004, 70, 2044–2051. [Google Scholar] [CrossRef] [PubMed]
- Kimura, M.; Tokai, T.; Takahashi-Ando, N.; Ohsato, S.; Fujimura, M. Molecular and genetic studies of Fusarium trichothecene biosynthesis: Pathways, genes, and evolution. Biosci. Biotechnol. Biochem. 2007, 71, 2105–2123. [Google Scholar] [CrossRef]
- Liang, J.M.; Xayamongkhon, H.; Broz, K.; Dong, Y.; McCormick, S.P.; Abramova, S.; Ward, T.J.; Ma, Z.H.; Kistler, H.C. Temporal dynamics and population genetic structure of Fusarium graminearum in the upper Midwestern United States. Fungal Genet. Biol. 2014, 73, 83–92. [Google Scholar] [CrossRef]
- Ramdass, A.C.; Villafana, R.T.; Rampersad, S.N. Comparative Sequence Analysis of TRI1 of Fusarium. Toxins 2019, 11, 689. [Google Scholar] [CrossRef]
- Davydov, D.R.; Davydova, N.Y.; Sineva, E.V.; Halpert, J.R. Interactions among cytochromes P450 in microsomal membranes. J. Biol. Chem. 2015, 290, 3850–3864. [Google Scholar] [CrossRef]
- Ward, T.J.; Clear, R.M.; Rooney, A.P.; O’Donnell, K.; Gaba, D.; Patrick, S.; Starkey, D.E.; Gilbert, J.; Geiser, D.M.; Nowicki, T.W. An adaptive evolutionary shift in Fusarium head blight pathogen populations is driving the rapid spread of more toxigenic Fusarium graminearum in North America. Fungal Genet. Biol. 2008, 45, 473–484. [Google Scholar] [CrossRef]
- Castiblanco, V.; Castillo, H.E.; Miedaner, T. Candidate genes for aggressiveness in a natural Fusarium culmorum population greatly differ between wheat and rye head blight. J. Fungi 2018, 4, 14. [Google Scholar] [CrossRef]
- Miedaner, T.; Vasquez, A.; Castiblanco, V.; Castillo, H.E.; Foroud, N.; Würschum, T.; Leiser, W. Genome-wide association study for deoxynivalenol production and aggressiveness in wheat and rye head blight by resequencing 92 isolates of Fusarium culmorum. BMC Genom. 2021, 22, 1–16. [Google Scholar] [CrossRef]
- Mueller, B.; Groves, C.L.; Smith, D.L. Chemotype and aggressiveness evaluation of Fusarium graminearum and Fusarium culmorum isolates from wheat fields in Wisconsin. Plant Dis. 2021, 105, 3686–3693. [Google Scholar] [CrossRef] [PubMed]
- Hara-Nishimura, I.; Matsushima, R.; Shimada, T.; Nishimura, M. Diversity and formation of endoplasmic reticulum-derived compartments in plants. are these compartments specific to plant cells? Plant Physiol. 2004, 136, 3435–3439. [Google Scholar] [CrossRef] [PubMed]
- Alexander, N.J.; McCormick, S.P.; Waalwijk, C.; van der Lee, T.; Proctor, R.H. The genetic basis for 3-ADON and 15-ADON trichothecene chemotypes in Fusarium. Fungal Genet. Biol. 2011, 48, 485–495. [Google Scholar] [CrossRef]
- Gang, G.; Miedaner, T.; Schuhmacher, U.; Schollenberger, M.; Geiger, H.H. Deoxynivalenol and nivalenol production by Fusarium culmorum isolates differing in aggressiveness toward winter rye. Phytopathology 1998, 88, 879–884. [Google Scholar] [CrossRef] [PubMed]
- Mateo, J.J.; Mateo, R.; Hinojo, M.J.; Llorens, A.; Jiménez, M. Liquid chromatographic determination of toxigenic secondary metabolites produced by Fusarium strains. J. Chromatogr. A 2002, 955, 245–256. [Google Scholar] [CrossRef]
- Leslie, J.F.; Summerell, B.A. The Fusarium Laboratory Manual; Kansas State University: Manhattan, NY, USA, 2006; ISBN 9780813819198. [Google Scholar] [CrossRef]
- Nirenberg, H. Untersuchungen über die morphologische und biologische Differenzierung in der Fusarium-Sektion Liseola. Mitt. Biol. Bundesanst. Land und Forstwirtsch. Berlin-Dahlem 1976, 169, 1–117. [Google Scholar]
- Brandfass, C.; Karlovsky, P. Upscaled CTAB-based DNA extraction and real-time PCR assays for Fusarium culmorum and F. graminearum DNA in plant material with reduced sampling error. Int. J. Mol. Sci. 2008, 9, 2306–2321. [Google Scholar] [CrossRef] [PubMed]
- Thompson, J.D.; Higgins, D.G.; Gibson, T.J. CLUSTAL W: Improving the sensitivity of progressive multiple sequence alignment through sequence weighting, position-specific gap penalties and weight matrix choice. Nucleic. Acids Res. 1994, 22, 4673–4680. [Google Scholar] [CrossRef]
- Beule, L.; Lehtsaar, E.; Rathgeb, A.; Karlovsky, P. Crop diseases and mycotoxin accumulation in temperate agroforestry systems. Sustainability 2019, 11, 2925. [Google Scholar] [CrossRef]
- Wenzl, T.; Haedrich, J.; Schaechtele, A.; Robouch, P.; Stroka, J. Guidance Document on the Estimation of LOD and LOQ for Measurements in the Field of Contaminants in Feed and Food; EUR 28099; Publications Office of the European Union: Luxembourg, 2016; ISBN 978-92-79-61768-3. [Google Scholar] [CrossRef]



| Isolate | Mycotoxin Content (mg/kg) | ||||
|---|---|---|---|---|---|
| NIV | DON | 15-ADON | 3-ADON | NX-2 | |
| 31.6st | 0.04 | 25.6 | <LOD | 156 | 3.2 |
| 215.1st | <LOD a | 4.2 | <LOD | 90.4 | 2.4 |
| 227.2cst | <LOD | 0.5 | <LOD | 2.1 | <LOD |
| 240.2sp | 0.38 | 355 | <LOD | 552 | 35.6 |
| IPP0211 | 0.31 | 566 | <LOD | 412 | 12.2 |
| IPP0212 | 0.10 | 162 | <LOD | 463 | 16.4 |
| IPP0213 | 0.33 | 639 | <LOD | 388 | 9.4 |
| IPP0618 | <LOD | 20.3 | <LOD | 175 | 3.7 |
| IPP0619 | <LOD | 15.9 | <LOD | 272 | 14.4 |
| IPP0999 | <LOQ b | <LOD | <LOD | <LOD | <LOD |
| IPP1000 | <LOD | 12.6 | <LOD | 132 | 2.7 |
| DSM62184 | 0.05 | 108 | <LOD | 173 | 13.5 |
| 966 | <LOD | 25.7 | <LOD | 118 | 3.2 |
| 969 | <LOQ | 14.3 | <LOD | 147 | 3.2 |
| 3.37 | <LOD | 9.0 | <LOD | 68.6 | 1.4 |
| DSM62188 | <LOQ | 46.5 | <LOD | 294 | 13.6 |
| 55.6st | <LOQ | 31.3 | <LOD | 85.9 | 1.2 |
| 59.6st c | 13.6 | 0.4 | 0.9 | 0.63 | 10.3 |
| K11.2 | <LOD | 1.0 | <LOD | 46.3 | 1.2 |
| J31.2 | 0.08 | 127 | <LOD | 447 | 18.6 |
| Position of Amino Acid Residue | ||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Location in TRI1 | 3 | 5 | 8 | 33 | 35 | 100 | 115 | 134 | 210 | 252 | 254 | 256 | 346 | 361 | 373 | 418 | 430 | 450 |
| F. culmorum NX+ (n = 18) a | I | S | S | A | K | S | F | G | S | R | L | T | F | I | D | Q | T | A |
| F. culmorum NX− (n = 2) b | I | S | S | A | K | S | F | G | S | R | L | T | F | I | D | Q | T | A |
| F. graminearum NX+ (n = 3) c | L | T | Q | T | Q | N | L | A | T | S | M | N | I | F | E | K | P | V |
| F. graminearum NX− (n = 3) d | L | T | Q | A | Q | S | F | A | S | R | L | T | F | I | D | Q | T | A |
| F. cerealis 25805 | I | S | S | A | K | S | F | G | S | R | L | T | F | I | D | Q | T | A |
| F. pseudograminearum 28062 | I | S | S | A | K | S | F | G | S | R | L | T | F | I | D | Q | T | A |
| F. poae FRC T-0962 | I | D | P | T | K | R | F | A | S | N | L | N | F | V | D | Q | T | A |
| F. incarnatum NRRL31160 | M | N | N | A | K | K | F | A | S | H | L | D | K | V | D | D | T | A |
| F. sambucinum FRC R-07843 | L | D | S | R | K | N | F | A | T | R | L | N | K | V | D | Q | T | A |
| F. langsethiae NRRL53410 | F | D | E | R | K | N | F | A | A | R | L | N | K | V | D | L | T | A |
| F. sporotrichioides NRRL29977 | F | D | E | R | K | N | F | A | A | R | L | N | K | V | D | Q | T | A |
| Primer Name | Primer Sequence (5′–3′) | Location a | Source |
|---|---|---|---|
| TRI16IF1 | GCCTSATAGCGACGATCTTGC | 0–485 | [30] |
| Tri1_5prime_RV | GACCGTGAATAACCTCCTTGATCAGT | This study | |
| Tri1_5prime_FW | ACTGATCAAGGAGGTTATTCACGGTC | 460–1225 | This study |
| Tri1_SH_R | CGCTGTCGAGAAGGAACATCTTG | This study | |
| Tri1_SH_F | GGCTATGTACAAGATGTTCCTTCTCG | 1194–1753 | This study |
| FgTRI1-R1 | AACAAGTGGCGAGATCAAACC | [30] | |
| FcTri1F | ATGGCTATCATCAGCAG | 1–1745 | This study |
| Tri1R | CAATTCCAATCGCAGACAA | [46] |
| Toxin | Ioniz. Mode | Parent Ion [m/z] | Fragm. Voltage [V] | Collision Energy [V] | Product Ion [m/z] c | LOD b [mg/kg] | LOQ b [mg/kg] |
|---|---|---|---|---|---|---|---|
| NIV | Pos | 313.1 | 93 | 7 | 205.0 | 0.01 | 0.03 |
| 15 | 175.0 | ||||||
| DON | Pos | 297.1 | 100 | 4 | 249.1 | 0.04 | 0.13 |
| 64 | 91.2 | ||||||
| 3-ADON | Pos | 339.2 | 100 | 8 | 231.1 | 0.02 | 0.08 |
| 8 | 203.0 | ||||||
| 15-ADON | Pos | 339.2 | 90 | 10 | 321.1 | 0.07 | 0.23 |
| 10 | 261.0 | ||||||
| 10 | 137.0 | ||||||
| NX-2 a | Pos | 325.2 | 90 | 10 | 247.2 | 0.04 | 0.12 |
| 10 | 229.2 | ||||||
| 10 | 199.1 | ||||||
| 25 | 121.2 | ||||||
| 35 | 105.1 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Schiwek, S.; Alhussein, M.; Rodemann, C.; Budragchaa, T.; Beule, L.; von Tiedemann, A.; Karlovsky, P. Fusarium culmorum Produces NX-2 Toxin Simultaneously with Deoxynivalenol and 3-Acetyl-Deoxynivalenol or Nivalenol. Toxins 2022, 14, 456. https://doi.org/10.3390/toxins14070456
Schiwek S, Alhussein M, Rodemann C, Budragchaa T, Beule L, von Tiedemann A, Karlovsky P. Fusarium culmorum Produces NX-2 Toxin Simultaneously with Deoxynivalenol and 3-Acetyl-Deoxynivalenol or Nivalenol. Toxins. 2022; 14(7):456. https://doi.org/10.3390/toxins14070456
Chicago/Turabian StyleSchiwek, Simon, Mohammad Alhussein, Charlotte Rodemann, Tuvshinjargal Budragchaa, Lukas Beule, Andreas von Tiedemann, and Petr Karlovsky. 2022. "Fusarium culmorum Produces NX-2 Toxin Simultaneously with Deoxynivalenol and 3-Acetyl-Deoxynivalenol or Nivalenol" Toxins 14, no. 7: 456. https://doi.org/10.3390/toxins14070456
APA StyleSchiwek, S., Alhussein, M., Rodemann, C., Budragchaa, T., Beule, L., von Tiedemann, A., & Karlovsky, P. (2022). Fusarium culmorum Produces NX-2 Toxin Simultaneously with Deoxynivalenol and 3-Acetyl-Deoxynivalenol or Nivalenol. Toxins, 14(7), 456. https://doi.org/10.3390/toxins14070456