Deciphering of Pod Borer [Helicoverpa armigera (Hübner)] Resistance in Cajanus platycarpus (Benth.) Offers Novel Insights on the Reprogramming and Role of Flavonoid Biosynthesis Pathway

 , , ,
, , ,  and
and 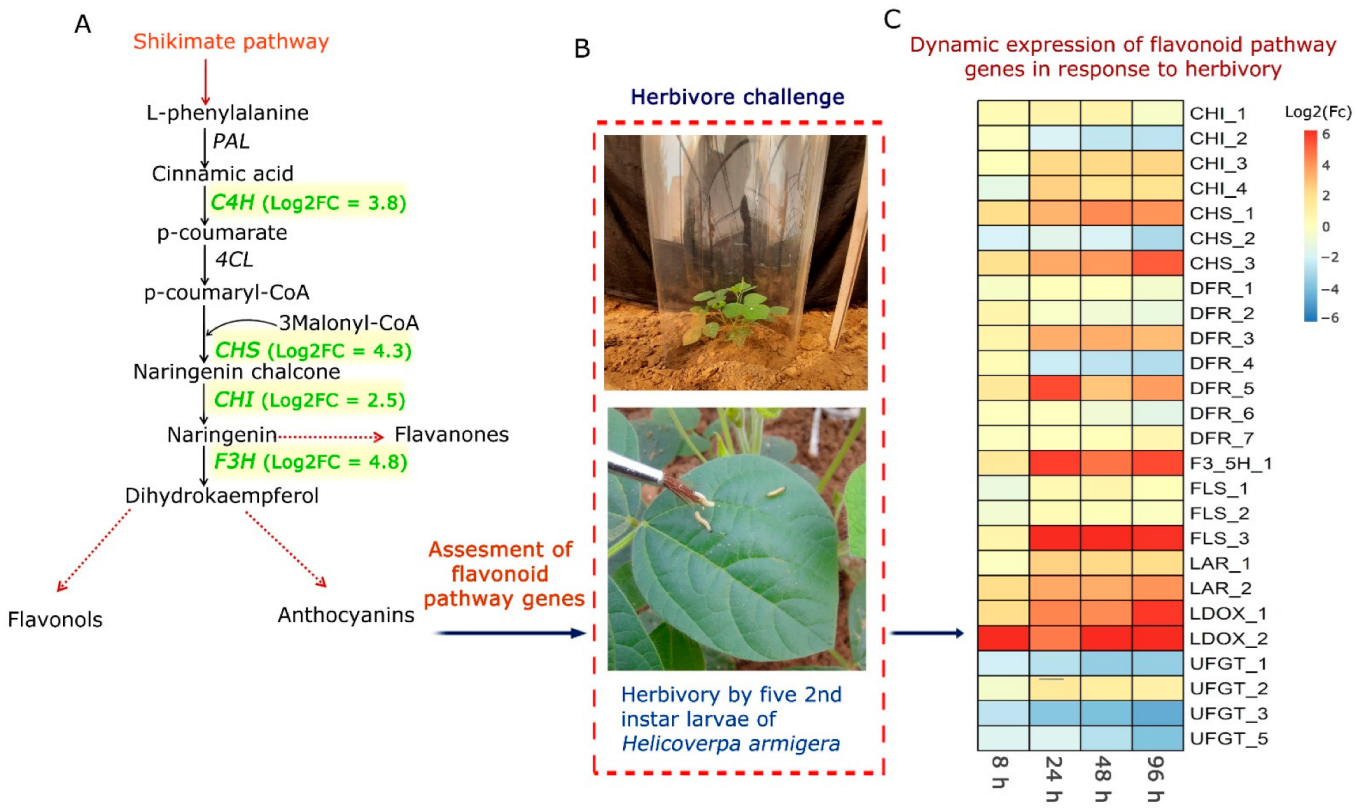{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
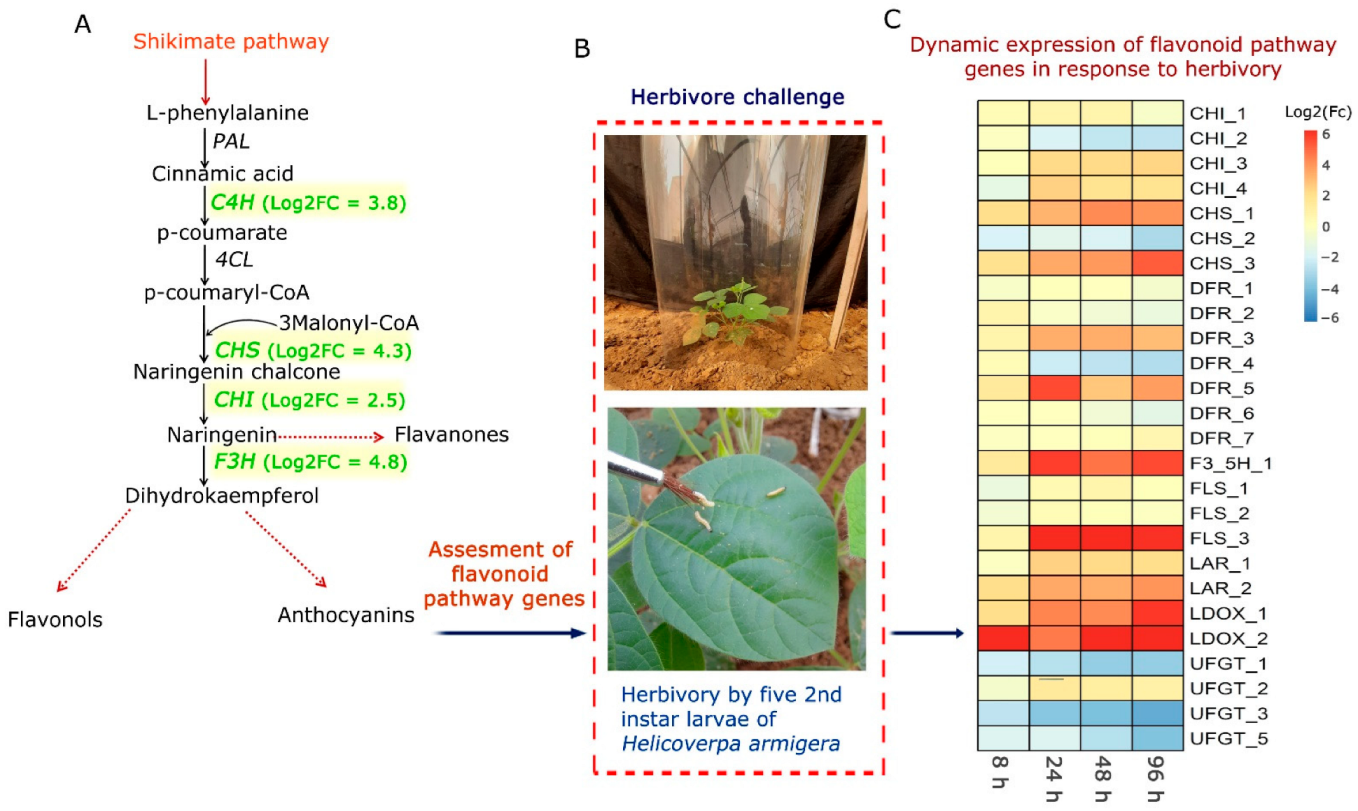
Plant Material and Herbivore Challenge by H. armigera
3. Expression Analysis
3.1. Identification of Flavonoid Pathway Genes from C. platycarpus Transcriptome Data
3.2. Total RNA Isolation and cDNA Synthesis
3.3. Real-Time PCR
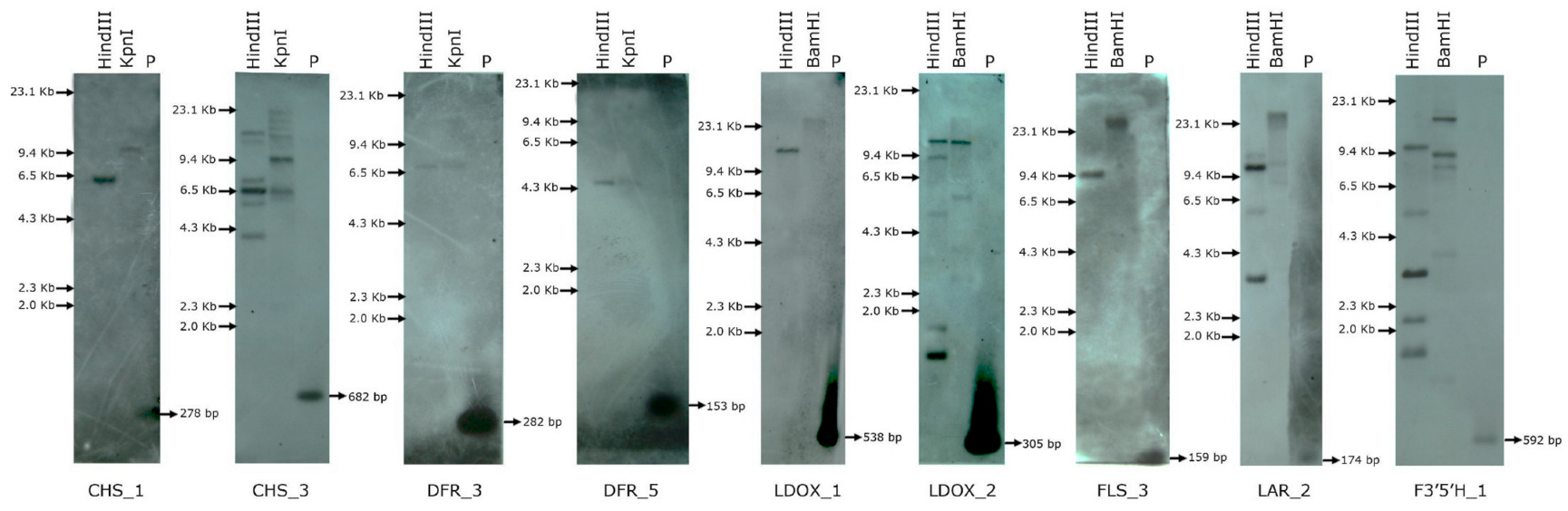
4. Copy Number Assessment of the Selected Flavonoid Biosynthesis Genes in C. platycarpus
4.1. Probe Designing
4.2. Genomic Southern Analysis
4.3. Targeted Quantitative Estimation of Flavonoids and Phenylpropanoids in C. platycarpus during H. armigera Infestation
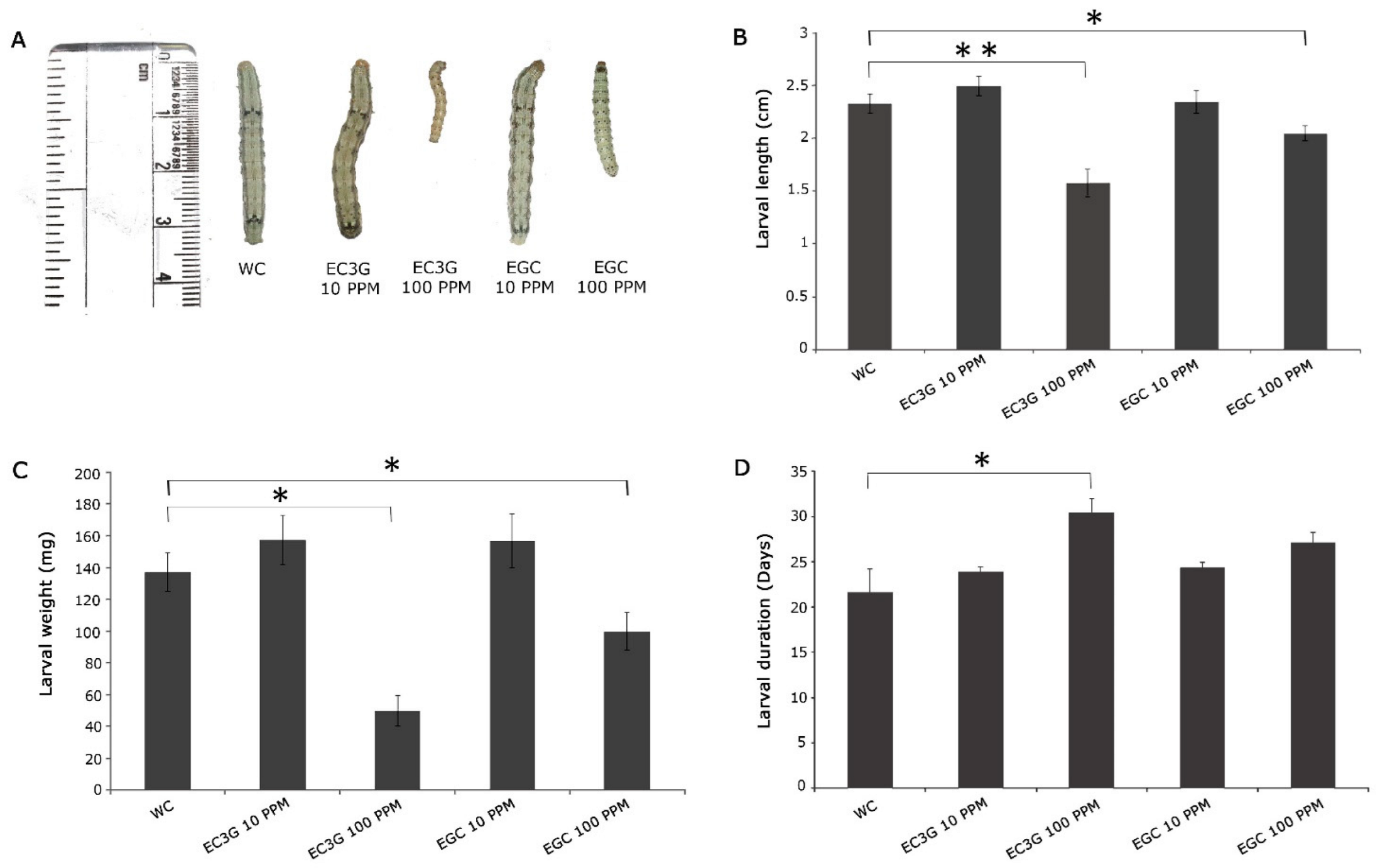
4.4. Assessment of the Biological Activities of Selected Flavonoids on the Growth and Development of H. armigera
5. Results and Discussion
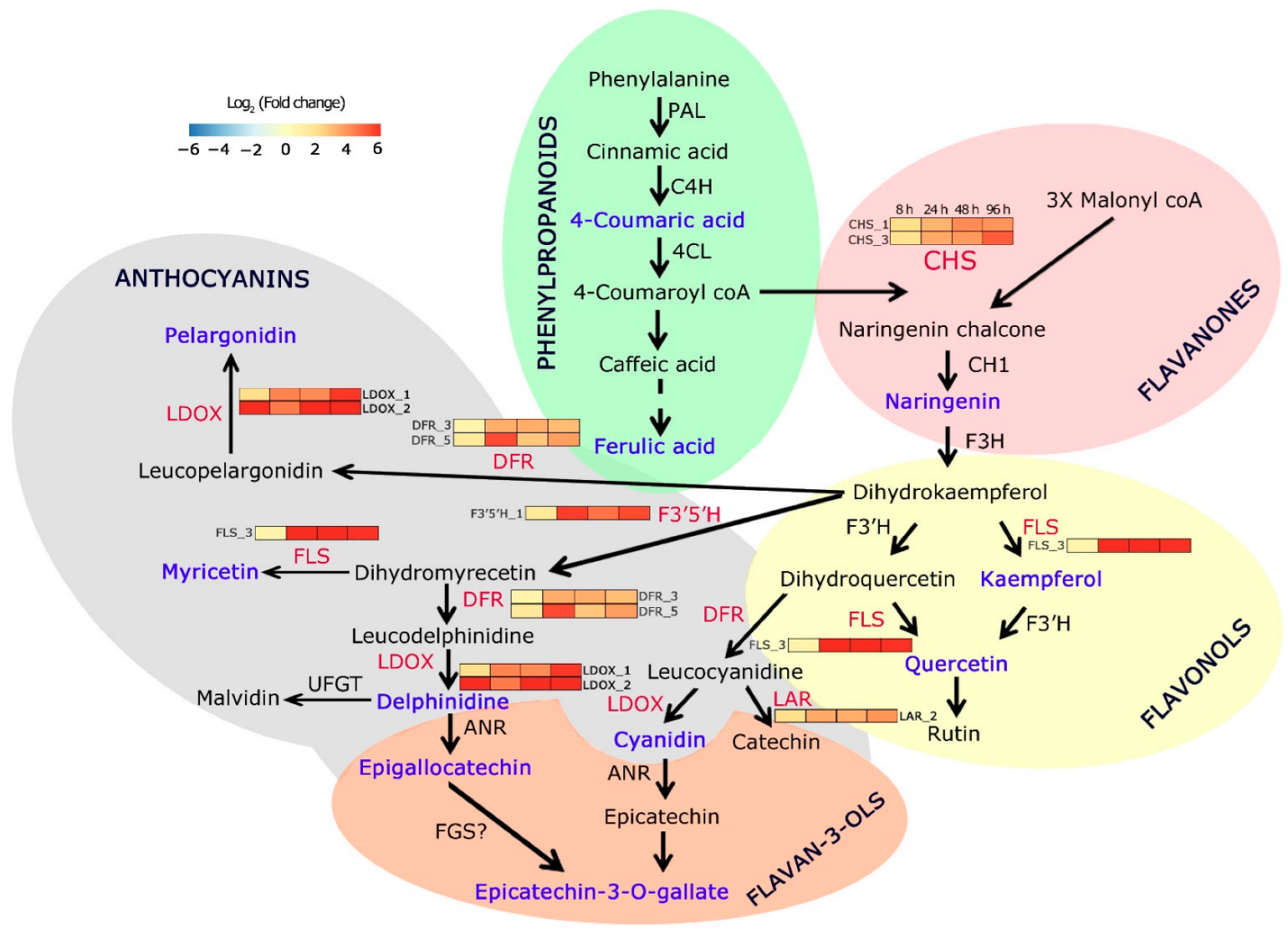
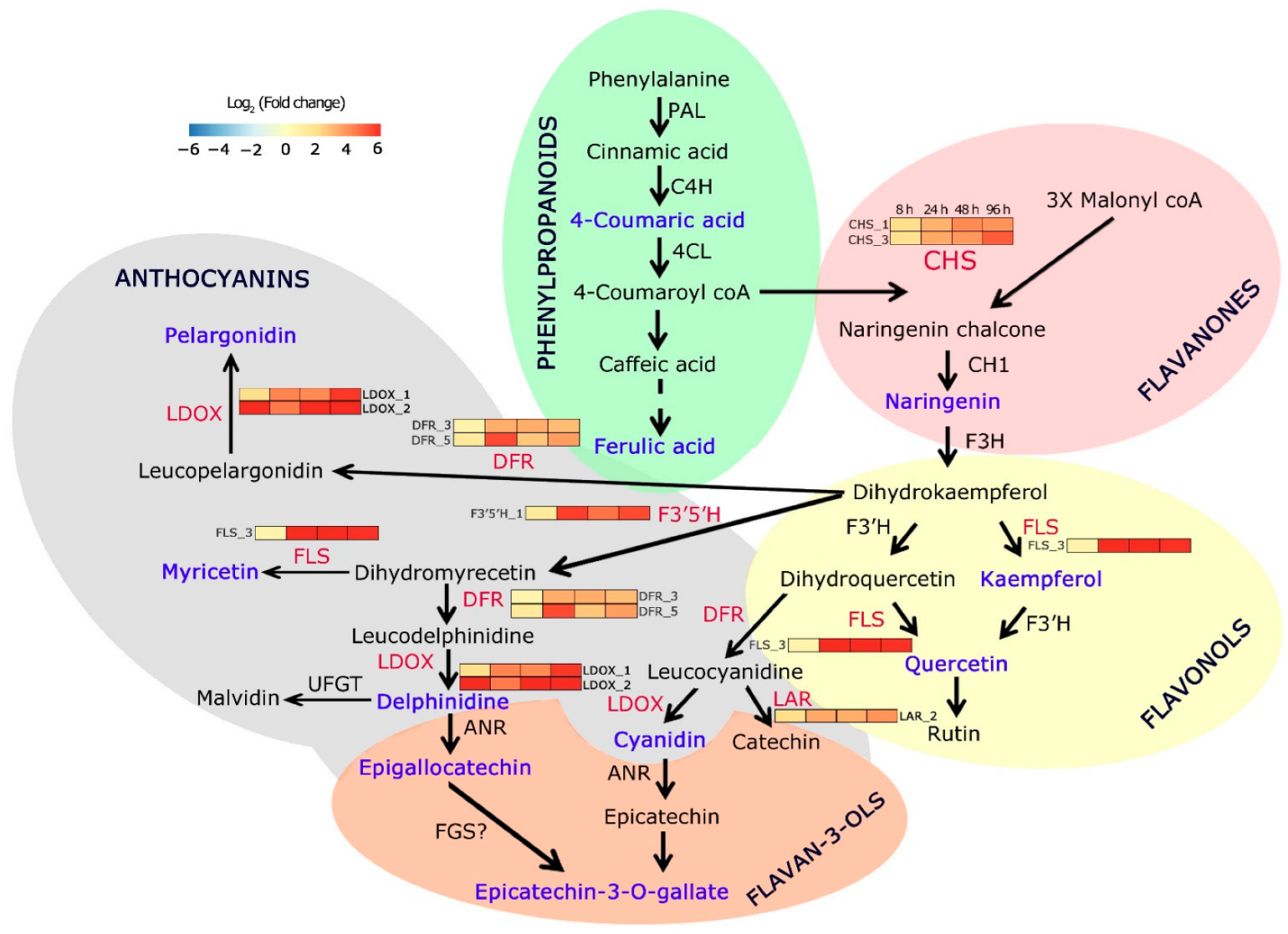
6. Dynamic Expression of Flavonoid Pathway Genes in C. platycarpus in Response to H. armigera
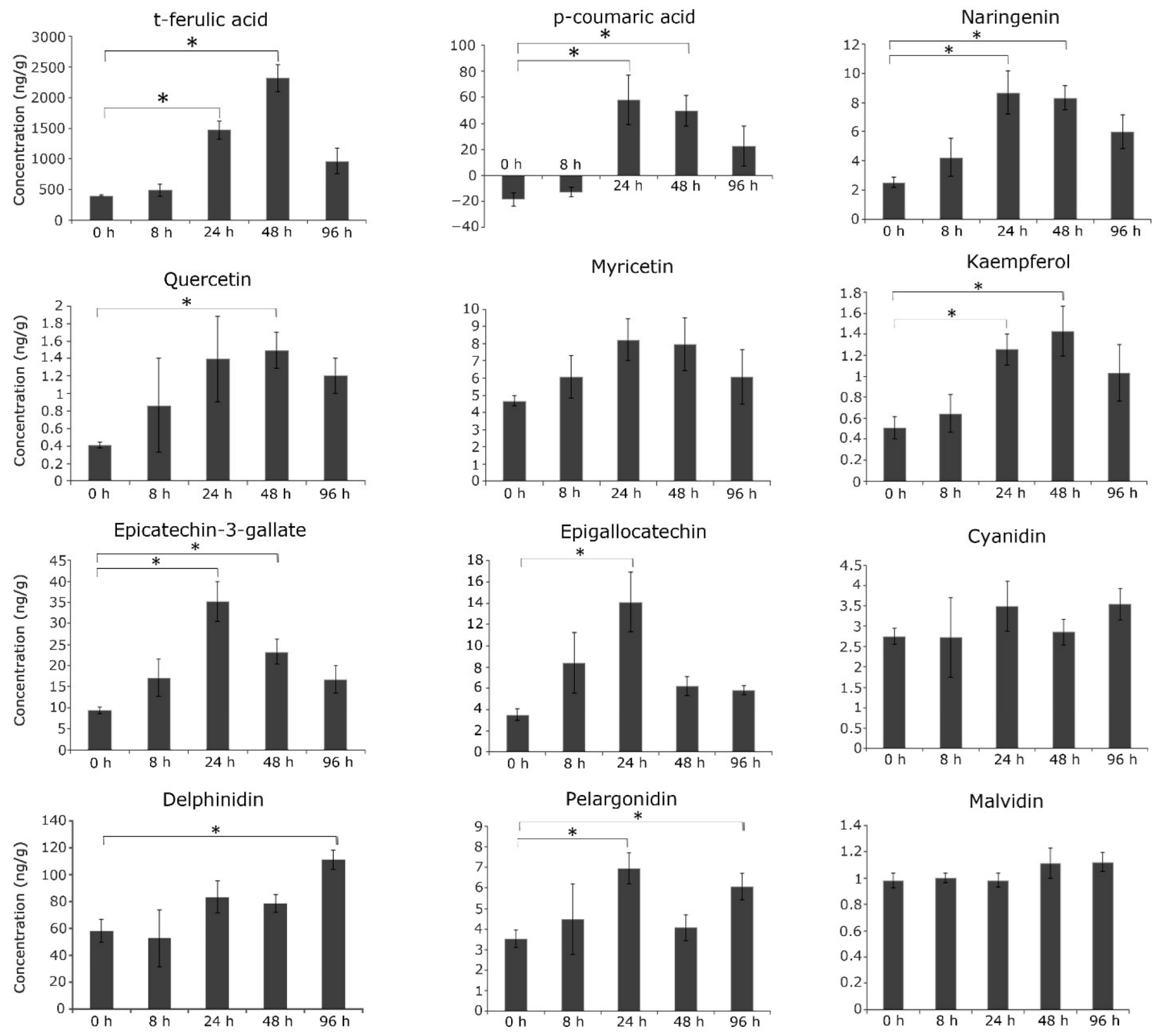
7. Flavonoids and Phenylpropanoids Are Differentially Accumulated during the Dynamic Response of C. platycarpus to H. armigera
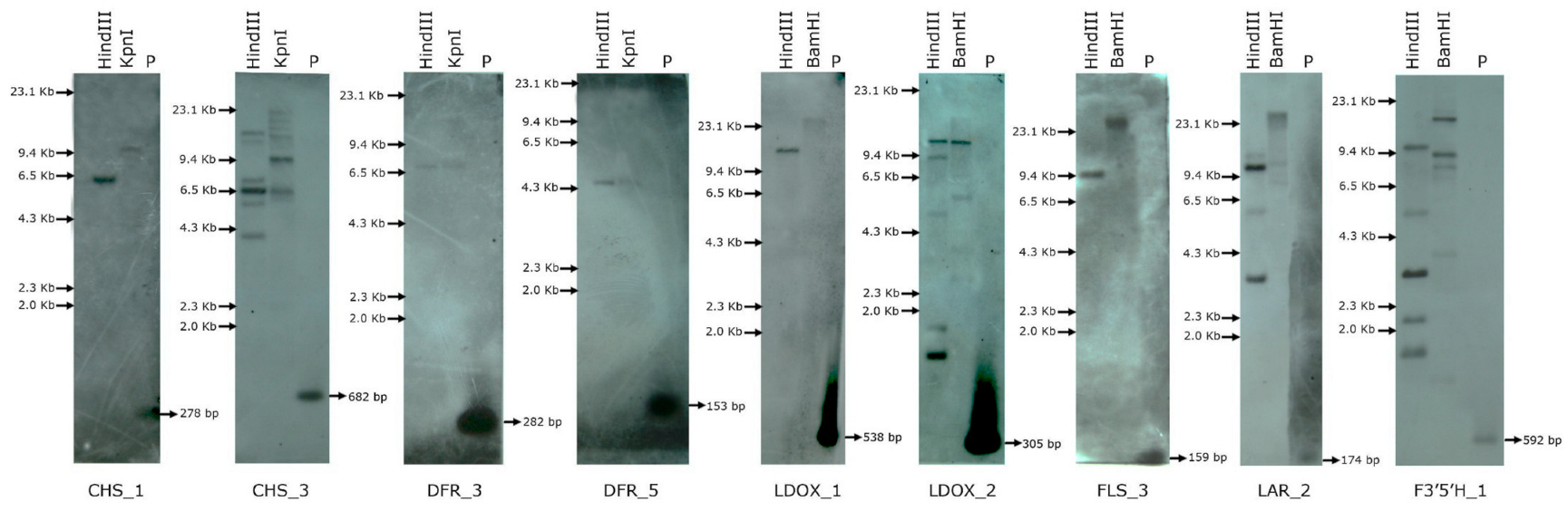
8. Assessment of the Copy Number of Flavonoid Biosynthesis Genes in C. platycarpus
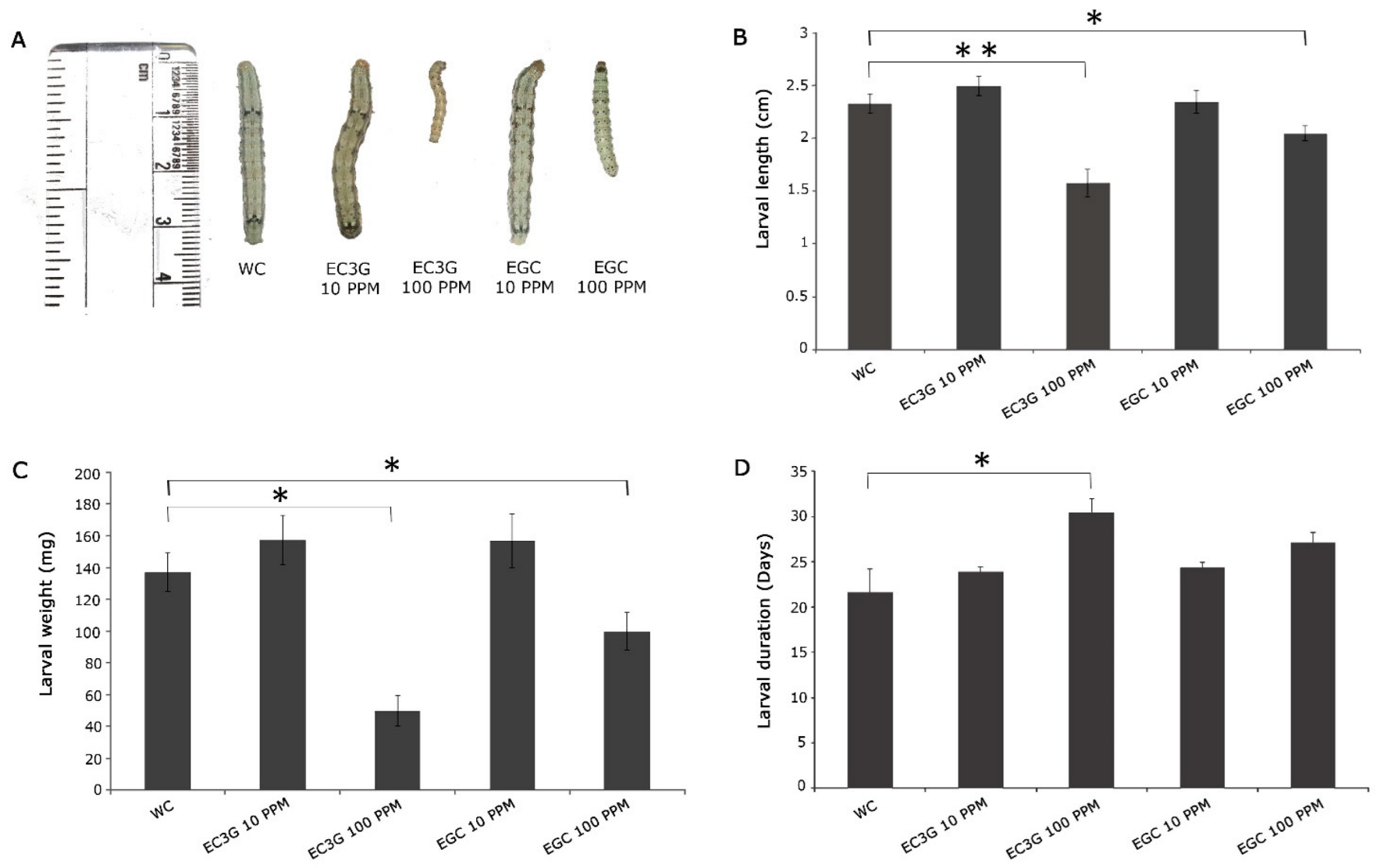
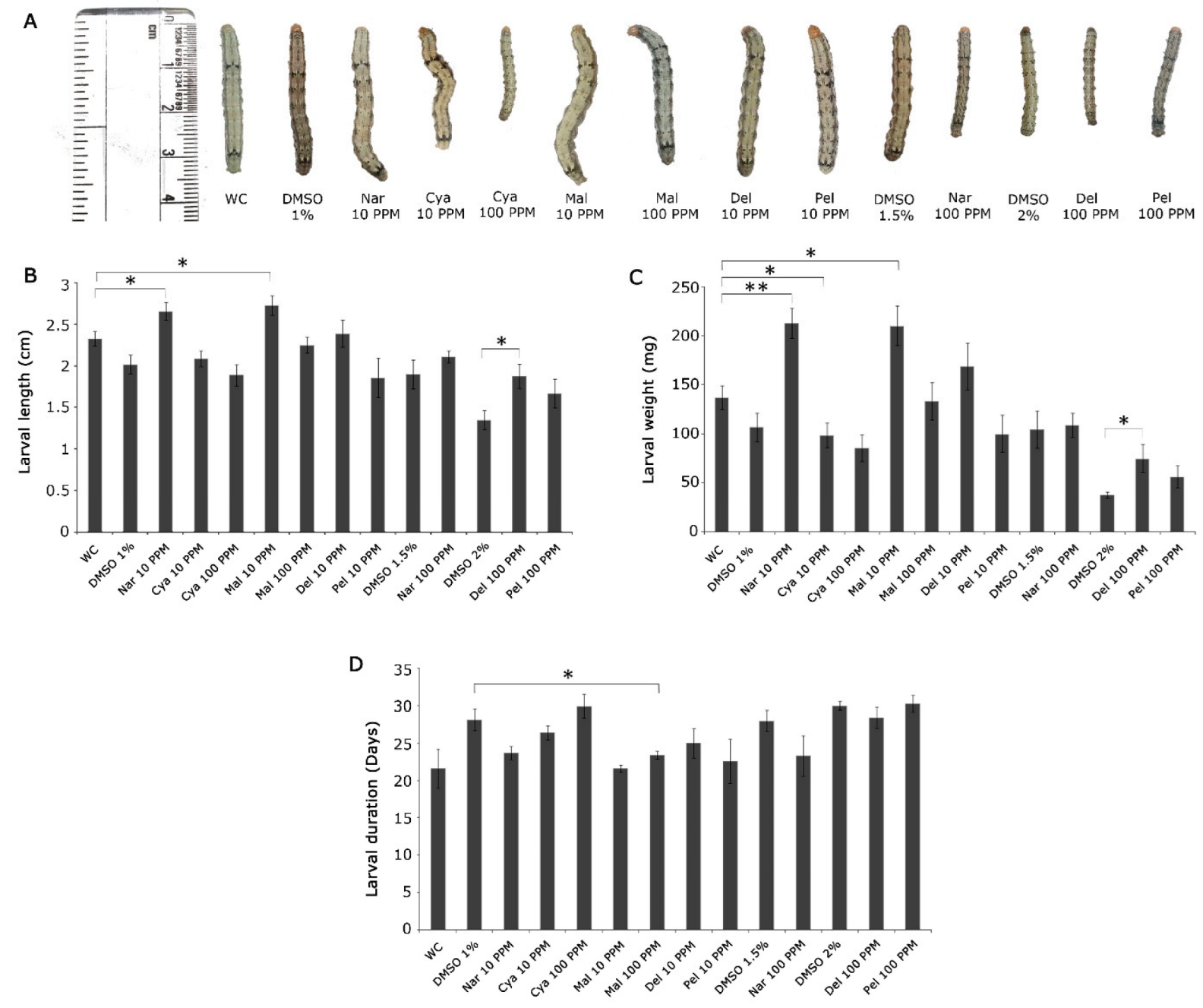
9. Validation of Differentially Produced Flavonoids on the Growth and Development of H. armigera
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Acknowledgments
Conflicts of Interest
References
- Negi, J.; Rathinam, M.; Sreevathsa, R.; Kumar, P.A. Transgenic Pigeonpea [Cajanus cajan (L.) Millsp.]. In Genetically Modified Crops; Springer: Singapore, 2021; pp. 79–96. [Google Scholar]
- Sharma, D.; Reddy, L.J.; Srivastava, R.K.; Saxena, K.B. A unique pigeonpea landrace with multiple properties. J. Food Leg. 2021, 34, 132–135. [Google Scholar]
- Sultana, R.; Saxena, K.B.; Kumar, R.R.; Kumar, D.; Kirti, M. Pigeonpea. In The Beans and the Peas; Woodhead Publishing: Cambridge, UK, 2021; pp. 217–240. [Google Scholar]
- Zhang, H.; Yasmin, F.; Song, B.H. Neglected treasures in the wild—legume wild relatives in food security and human health. Curr. Opin. Plant Biol. 2019, 49, 17–26. [Google Scholar] [CrossRef]
- Pratap, A.; Das, A.; Kumar, S.; Gupta, S. Current perspectives on introgression breeding in food legumes. Front. Plant Sci. 2021, 11, 2118. [Google Scholar] [CrossRef]
- Jadhav, D.R.; Mallikarjuna, N.; Sharma, H.C.; Saxena, K.B. Introgression of Helicoverpa armigera resistance from Cajanus acutifolius-a wild relative from secondary gene pool of pigeon pea (Cajanus cajan). Asian J. Agric. Sci. 2012, 4, 242–248. [Google Scholar]
- Vanambathina, P.; Henry, R.J.; Rachaputi, R.C.; Furtado, A. Secondary genepool of Australian Cajanus species contains sources of resistance to Helicoverpa armigera (Hübner). Ann. Appl. Biol. 2021, 180, 259–272. [Google Scholar] [CrossRef]
- Mallikarjuna, N.; Senapathy, S.; Jadhav, D.R.; Saxena, K.; Sharma, H.C.; Upadhyaya, H.D.; Rathore, A.; Varshney, R. Progress in the utilization of Cajanus platycarpus (Benth.) Maesen in pigeonpea improvement. Plant Breed. 2011, 130, 507–514. [Google Scholar] [CrossRef] [Green Version]
- Mallikarjuna, N.; Jadhav, D.; Reddy, P. Introgression of Cajanus platycarpus genome into cultivated pigeonpea, C. cajan. Euphytica 2006, 149, 161–167. [Google Scholar] [CrossRef] [Green Version]
- Sharma, S.; Paul, P.J.; Kumar, C.V.S.; Nimje, C. Utilizing wild Cajanus platycarpus, a tertiary genepool species for enriching variability in the primary genepool for pigeonpea improvement. Front. Plant Sci. 2020, 11, 1055. [Google Scholar] [CrossRef]
- Mallikarjuna, N.; Sharma, H.C.; Upadhyaya, H.D. Exploitation of wild relatives of pigeonpea and chickpea for resistance to Helicoverpa armigera. J. SAT Agric. Res. 2007, 3, 4. [Google Scholar]
- War, A.R.; Paulraj, M.G.; Ahmad, T.; Buhroo, A.A.; Hussain, B.; Ignacimuthu, S.; Sharma, H.C. Mechanisms of plant defense against insect herbivores. Plant Signal. Behav. 2012, 7, 1306–1320. [Google Scholar] [CrossRef] [Green Version]
- Dong, N.Q.; Lin, H.X. Contribution of phenylpropanoid metabolism to plant development and plant–environment interactions. J. Integr. Plant Biol. 2021, 63, 180–209. [Google Scholar] [CrossRef]
- Sharma, A.; Shahzad, B.; Rehman, A.; Bhardwaj, R.; Landi, M.; Zheng, B. Response of phenylpropanoid pathway and the role of polyphenols in plants under abiotic stress. Molecules 2019, 24, 2452. [Google Scholar] [CrossRef] [Green Version]
- Simmonds, M.S.; Stevenson, P.C. Effects of isoflavonoids from Cicer on larvae of Heliocoverpa armigera. J. Chem. Ecol. 2001, 27, 965–977. [Google Scholar] [CrossRef]
- Goławska, S.; Sprawka, I.; Łukasik, I.; Goławski, A. Are naringenin and quercetin useful chemicals in pest-management strategies? J. Pest Sci. 2014, 7, 173–180. [Google Scholar] [CrossRef] [Green Version]
- Wittstock, U.; Gershenzon, J. Constitutive plant toxins and their role in defense against herbivores and pathogens. Curr. Opin. Plant Biol. 2002, 5, 300–307. [Google Scholar] [CrossRef]
- Erb, M.; Reymond, P. Molecular interactions between plants and insect herbivores. Annu. Rev. Plant Biol. 2019, 70, 527–557. [Google Scholar] [CrossRef] [Green Version]
- Babu, C.; Sharma, H.C.; Madhumati, T.; Raghavaiah, G.; Rao, V.S. A semi-synthetic chickpea flour based diet for long-term maintenance of laboratory culture of Helicoverpa armigera. Indian J. Entomol. 2014, 76, 336–340. [Google Scholar]
- Ramkumar, N.; Rathinam, M.; Singh, S.; Kesiraju, K.; Muniyandi, V.; Singh, N.K.; Dash, P.K.; Sreevathsa, R. Assessment of Pigeonpea (Cajanus cajan L.) transgenics expressing Bt ICPs, Cry2Aa and Cry1AcF under nethouse containment implicated an effective control against herbivory by Helicoverpa armigera (Hübner). Pest Manag. Sci. 2020, 76, 1902–1911. [Google Scholar] [CrossRef]
- Rathinam, M.; Mishra, P.; Mahato, A.K.; Singh, N.K.; Rao, U.; Sreevathsa, R. Comparative transcriptome analyses provide novel insights into the differential response of Pigeonpea (Cajanus cajan L.) and its wild relative (Cajanus platycarpus (Benth.) Maesen) to herbivory by Helicoverpa armigera (Hübner). Plant Mol. Biol. 2019, 101, 163–182. [Google Scholar] [CrossRef]
- Sinha, P.; Singh, V.K.; Suryanarayana, V.; Krishnamurthy, L.; Saxena, R.K.; Varshney, R.K. Evaluation and validation of housekeeping genes as reference for gene expression studies in pigeonpea (Cajanus cajan) under drought stress conditions. PLoS ONE 2015, 10, e0122847. [Google Scholar] [CrossRef]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2−ΔΔCT method. Methods 2001, 5, 402–408. [Google Scholar] [CrossRef]
- Porebski, S.; Bailey, L.G.; Baum, B.R. Modification of a CTAB DNA extraction protocol for plants containing high polysaccharide and polyphenol components. Plant Mol. Biol. Rep. 1997, 15, 8–15. [Google Scholar] [CrossRef]
- Naik, J.; Rajput, R.; Pucker, B.; Stracke, R.; Pandey, A. The R2R3-MYB transcription factor MtMYB134 orchestrates flavonol biosynthesis in Medicago truncatula. Plant Mol. Biol. 2021, 106, 157–172. [Google Scholar] [CrossRef]
- Su, J.; Lai, T.; Li, J. Susceptibility of field populations of Spodoptera litura (Fabricius) (Lepidoptera: Noctuidae) in China to chlorantranilip role and the activities of detoxification enzymes. Crop Prot. 2012, 42, 217–222. [Google Scholar] [CrossRef]
- Rothwell, J.A.; Perez-Jimenez, J.; Neveu, V.; Medina-Remon, A.; M'hiri, N.; García-Lobato, P.; Manach, C.; Knox, C.; Eisner, R.; Wishart, D.S.; et al. Phenol-Explorer 3.0: A major update of the Phenol-Explorer database to incorporate data on the effects of food processing on polyphenol content. Database 2013, 2013, bat070. [Google Scholar] [CrossRef]
- Shah, A.; Tyagi, S.; Saratale, G.D.; Guzik, U.; Hu, A.; Sreevathsa, R.; Reddy, V.D.; Rai, V.; Mulla, S.I. A comprehensive review on the influence of light on signaling cross-talk and molecular communication against phyto-microbiome interactions. Crit. Rev. Biotechnol. 2021, 41, 370–393. [Google Scholar] [CrossRef]
- Jan, R.; Asaf, S.; Numan, M.; Kim, K.M. Plant secondary metabolite biosynthesis and transcriptional regulation in response to biotic and abiotic stress conditions. Agronomy 2021, 11, 968. [Google Scholar] [CrossRef]
- Ahmad, M.A.; Prakash, P.; Singh, H.B. Role of secondary metabolites and biostimulants in conferring biotic and abiotic stress tolerance in crop plants: An emerging application in sustainable agriculture. In New and Future Developments in Microbial Biotechnology and Bioengineering; Elsevier: Amsterdam, The Netherlands, 2022; pp. 355–360. [Google Scholar]
- Sharma, H.C.; Sujana, G.; Manohar Rao, D. Morphological and chemical components of resistance to pod borer, Helicoverpa armigera in wild relatives of pigeonpea. Arthropod-Plant Interact. 2009, 3, 151–161. [Google Scholar] [CrossRef] [Green Version]
- Sujana, G.; Sharma, H.C.; Rao, D.M. Antixenosis and antibiosis components of resistance to pod borer Helicoverpa armigera in wild relatives of pigeonpea. Int. J. Trop. Insect Sci. 2008, 28, 191–200. [Google Scholar]
- Rathinam, M.; Roschitzki, B.; Grossmann, J.; Mishra, P.; Kunz, L.; Wolski, W.; Panse, C.; Tyagi, S.; Rao, U.; Schlapbach, R.; et al. Unraveling the proteomic changes involved in the resistance response of Cajanus platycarpus to herbivory by Helicoverpa armigera. Appl. Microbiol. Biotechnol. 2020, 104, 7603–7618. [Google Scholar] [CrossRef]
- Ngugi-Dawit, A.; Hoang, T.M.L.; Williams, B.; Higgins, T.J.; Mundree, S.G. A wild Cajanus scarabaeoides (L.), Thouars, IBS 3471, for improved insect-resistance in cultivated pigeonpea. Agronomy 2020, 10, 517. [Google Scholar] [CrossRef] [Green Version]
- Zhang, H.; Mittal, N.; Leamy, L.J.; Barazani, O.; Song, B.H. Back into the wild—Apply untapped genetic diversity of wild relatives for crop improvement. Evol. Appl. 2017, 10, 5–24. [Google Scholar] [CrossRef]
- Rathinam, M.; Marimuthu, S.K.; Tyagi, S.; Kesiraju, K.; Alagiamanavalan, L.P.; Rao, U.; Sreevathsa, R. Characterization and in planta validation of a CHI4 chitinase from Cajanus platycarpus (Benth.) Maesen for its efficacy against pod borer, Helicoverpa armigera (Hübner). Pest Manag. Sci. 2021, 77, 2337–2349. [Google Scholar] [CrossRef]
- Lu, Y.; Chen, Q.; Bu, Y.; Luo, R.; Hao, S.; Zhang, J.; Tian, J.; Yao, Y. Flavonoid accumulation plays an important role in the rust resistance of Malus plant leaves. Front. Plant Sci. 2017, 8, 1286. [Google Scholar] [CrossRef] [Green Version]
- Dai, Z.; Tan, J.; Zhou, C.; Yang, X.; Yang, F.; Zhang, S.; Sun, S.; Miao, X.; Shi, Z. The OsmiR396–Os GRF 8–OsF3H-flavonoid pathway mediates resistance to the brown planthopper in rice (Oryza sativa). Plant Biotechnol. J. 2019, 17, 1657–1669. [Google Scholar] [CrossRef] [Green Version]
- Trujillo-Pahua, V.; Vargas-Ponce, O.; Rodríguez-Zaragoza, F.A.; Ordaz-Ortiz, J.J.; Délano-Frier, J.P.; Winkler, R.; Sánchez-Hernández, C.V. Metabolic response to larval herbivory in three Physalis species. Plant Signal. Behav. 2021, 16, 1962050. [Google Scholar] [CrossRef]
- Wang, Y.R.; Zhang, J.S.; Wang, R.; Hou, Y.M.; Fu, H.A.; Xie, Y.; Gao, S.J.; Wang, J.D. Unveiling sugarcane defense response to Mythimna separata herbivory by a combination of transcriptome and metabolic analyses. Pest Manag. Sci. 2021, 77, 4799–4809. [Google Scholar] [CrossRef]
- Simmonds, M.S. Flavonoid–insect interactions: Recent advances in our knowledge. Phytochemistry 2003, 64, 21–30. [Google Scholar] [CrossRef]
- Mithöfer, A.; Boland, W. Plant defense against herbivores: Chemical aspects. Annu. Rev. Plant Biol. 2012, 63, 431–450. [Google Scholar] [CrossRef] [Green Version]
- Shah, A.; Smith, D.L. Flavonoids in agriculture: Chemistry and roles in, biotic and abiotic stress responses, and microbial associations. Agronomy 2020, 10, 1209. [Google Scholar] [CrossRef]
- Alamgir, K.M.; Hojo, Y.; Christeller, J.T.; Fukumoto, K.; Isshiki, R.; Shinya, T.; Baldwin, I.T.; Galis, I. Systematic analysis of rice (Oryza sativa) metabolic responses to herbivory. Plant Cell Environ. 2016, 39, 453–466. [Google Scholar] [CrossRef]
- Khizar, M.; Shi, J.; Saleem, S.; Liaquat, F.; Ashraf, M.; Latif, S.; Haroon, U.; Hassan, S.W.; Rehman, S.U.; Chaudhary, H.J.; et al. Resistance associated metabolite profiling of Aspergillus leaf spot in cotton through non-targeted metabolomics. PLoS ONE 2020, 15, e0228675. [Google Scholar] [CrossRef]
- Baozhu, L.; Ruonan, F.; Yanting, F.; Runan, L.; Hui, Z.; Tingting, C.; Jiong, L.; Han, L.; Xiang, Z.; Chunpeng, S. The flavonoid biosynthesis regulator PFG3 confers drought stress tolerance in plants by promoting flavonoid accumulation. Environ. Exp. Bot. 2022, 196, 104792. [Google Scholar] [CrossRef]
- Johnson, E.T.; Berhow, M.A.; Dowd, P.F. Constitutive expression of the maize genes B1 and C1 in transgenic Hi II maize results in differential tissue pigmentation and generates resistance to Helicoverpa zea. J. Agric. Food Chem. 2010, 58, 2403–2409. [Google Scholar] [CrossRef]
- Luo, P.; Ning, G.; Wang, Z.; Shen, Y.; Jin, H.; Li, P.; Huang, S.; Zhao, J.; Bao, M. Disequilibrium of flavonol synthase and dihydroflavonol-4-reductase expression associated tightly to white vs. red color flower formation in plants. Front. Plant Sci. 2016, 6, 1257. [Google Scholar] [CrossRef] [Green Version]
- Jan, R.; Aaqil Khan, M.; Asaf, S.; Park, J.R.; Lee, I.J.; Kim, K.M. Flavonone 3-hydroxylase Relieves Bacterial Leaf Blight Stress in Rice via Over accumulation of Antioxidant Flavonoids and Induction of Defense Genes and Hormones. Int. J. Mol. Sci. 2021, 22, 6152. [Google Scholar] [CrossRef]
- Gómez, J.D.; Vital, C.E.; Oliveira, M.G.; Ramos, H.J. Broad range flavonoid profiling by LC/MS of soybean genotypes contrasting for resistance to Anticarsia gemmatalis (Lepidoptera: Noctuidae). PLoS ONE 2018, 13, e0205010. [Google Scholar] [CrossRef]
- Golla, S.K.; Rajasekhar, P.; Sharma, S.P.; Hari Prasad, K.V.; Sharma, H.C. Antixenosis and antibiosis mechanisms of resistance to pod borer, Helicoverpa armigera in wild relatives of chickpea, Cicer arietinum. Euphytica 2018, 214, 88. [Google Scholar] [CrossRef]
- Onkokesung, N.; Reichelt, M.; van Doorn, A.; Schuurink, R.C.; van Loon, J.J.; Dicke, M. Modulation of flavonoid metabolites in Arabidopsis thaliana through overexpression of the MYB75 transcription factor: Role of kaempferol-3, 7-dirhamnoside in resistance to the specialist insect herbivore Pieris brassicae. J. Exp. Bot. 2014, 65, 2203–2217. [Google Scholar] [CrossRef] [Green Version]
- Chacón-Fuentes, M.; Parra, L.; Rodriguez-Saona, C.; Seguel, I.; Ceballos, R.; Quiroz, A. Domestication in murtilla (Ugni molinae) reduced defensive flavonol levels but increased resistance against a native herbivorous insect. Environ. Entomol. 2015, 44, 627–637. [Google Scholar] [CrossRef]
- Su, Q.; Zhou, Z.; Zhang, J.; Shi, C.; Zhang, G.; Jin, Z.; Wang, W.; Li, C. Effect of plant secondary metabolites on common cutworm, Spodoptera litura (Lepidoptera: Noctuidae). Entomol. Res. 2018, 48, 8–26. [Google Scholar] [CrossRef]
- Mikani, A. Effect of kaempferol on ecdysteroid titer and oocyte size via tachykinin-4 in cotton bollworm, Helicoverpa armigera (Lepidoptera: Noctuidae). J. Crop Prot. 2019, 8, 153–162. [Google Scholar]
- Hu, Q.; Min, L.; Yang, X.; Jin, S.; Zhang, L.; Li, Y.; Ma, Y.; Qi, X.; Li, D.; Liu, H.; et al. Laccase GhLac1 modulates broad-spectrum biotic stress tolerance via manipulating phenylpropano, d pathway and jasmonic acid synthesis. Plant Physiol. 2018, 176, 1808–1823. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ngugi-Dawit, A.; Njaci, I.; Higgins, T.J.; Williams, B.; Ghimire, S.R.; Mundree, S.G.; Hoang, L.T.M. Comparative TMT Proteomic Analysis Unveils Unique Insights into Helicoverpa armigera (Hübner) Resistance in Cajanus scarabaeoides (L.) Thouars. Int. J. Mol. Sci. 2021, 22, 5941. [Google Scholar] [CrossRef]
- Karageorgou, P.; Manetas, Y. The importance of being red when young: Anthocyanins and the protection of young leaves of Quercus coccifera from insect herbivory and excess light. Tree Physiol. 2006, 26, 613–621. [Google Scholar] [CrossRef] [Green Version]
- Schlindwein, C.C.D.; Feet-Neto, A.G.; Dillenburg, L.R. Chemical and mechanical changes during leaf expansion of four woody species of a dry Restinga woodland. Plant Biol. 2006, 8, 430–438. [Google Scholar] [CrossRef]
- Hedin, P.A.; Jenkin, J.N.; Collum, D.H.; White, W.H.; Parrott, W.L.; MacGown, M.W. Cyanidin-3-β-glucoside, a newly recognized basis for resistance in cotton to the tobacco budworm Heliothis virescens (Fab.) (Lepidoptera: Noctuidae). Experientia 1983, 39, 799–801. [Google Scholar] [CrossRef]
- Jenkins, J.N.; Hedin, P.A.; Parrott, W.L.; McCarty, J.C.; White, W.H. Cotton allelochemics and growth of tobacco budworm larvae. Crop Sci. 1983, 23, 1195–1198. [Google Scholar] [CrossRef]
- Johnson, E.T.; Berhow, M.A.; Dowd, P.F. Colored and white sectors from star-patterned petunia flowers display differential resistance to corn earworm and cabbage looper larvae. J. Chem. Ecol. 2008, 34, 757–765. [Google Scholar] [CrossRef]
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tyagi, S.; Rathinam, M.; Shashank, P.R.; Chaudhary, N.; Shasany, A.K.; Sreevathsa, R. Deciphering of Pod Borer [Helicoverpa armigera (Hübner)] Resistance in Cajanus platycarpus (Benth.) Offers Novel Insights on the Reprogramming and Role of Flavonoid Biosynthesis Pathway. Toxins 2022, 14, 455. https://doi.org/10.3390/toxins14070455
Tyagi S, Rathinam M, Shashank PR, Chaudhary N, Shasany AK, Sreevathsa R. Deciphering of Pod Borer [Helicoverpa armigera (Hübner)] Resistance in Cajanus platycarpus (Benth.) Offers Novel Insights on the Reprogramming and Role of Flavonoid Biosynthesis Pathway. Toxins. 2022; 14(7):455. https://doi.org/10.3390/toxins14070455
Chicago/Turabian StyleTyagi, Shaily, Maniraj Rathinam, Pathour Rajendra Shashank, Nidhee Chaudhary, Ajit Kumar Shasany, and Rohini Sreevathsa. 2022. "Deciphering of Pod Borer [Helicoverpa armigera (Hübner)] Resistance in Cajanus platycarpus (Benth.) Offers Novel Insights on the Reprogramming and Role of Flavonoid Biosynthesis Pathway" Toxins 14, no. 7: 455. https://doi.org/10.3390/toxins14070455
APA StyleTyagi, S., Rathinam, M., Shashank, P. R., Chaudhary, N., Shasany, A. K., & Sreevathsa, R. (2022). Deciphering of Pod Borer [Helicoverpa armigera (Hübner)] Resistance in Cajanus platycarpus (Benth.) Offers Novel Insights on the Reprogramming and Role of Flavonoid Biosynthesis Pathway. Toxins, 14(7), 455. https://doi.org/10.3390/toxins14070455