Neurophysiological Basis of Deep Brain Stimulation and Botulinum Neurotoxin Injection for Treating Oromandibular Dystonia



{kind=link}
{kind=link}
Abstract
1. Introduction
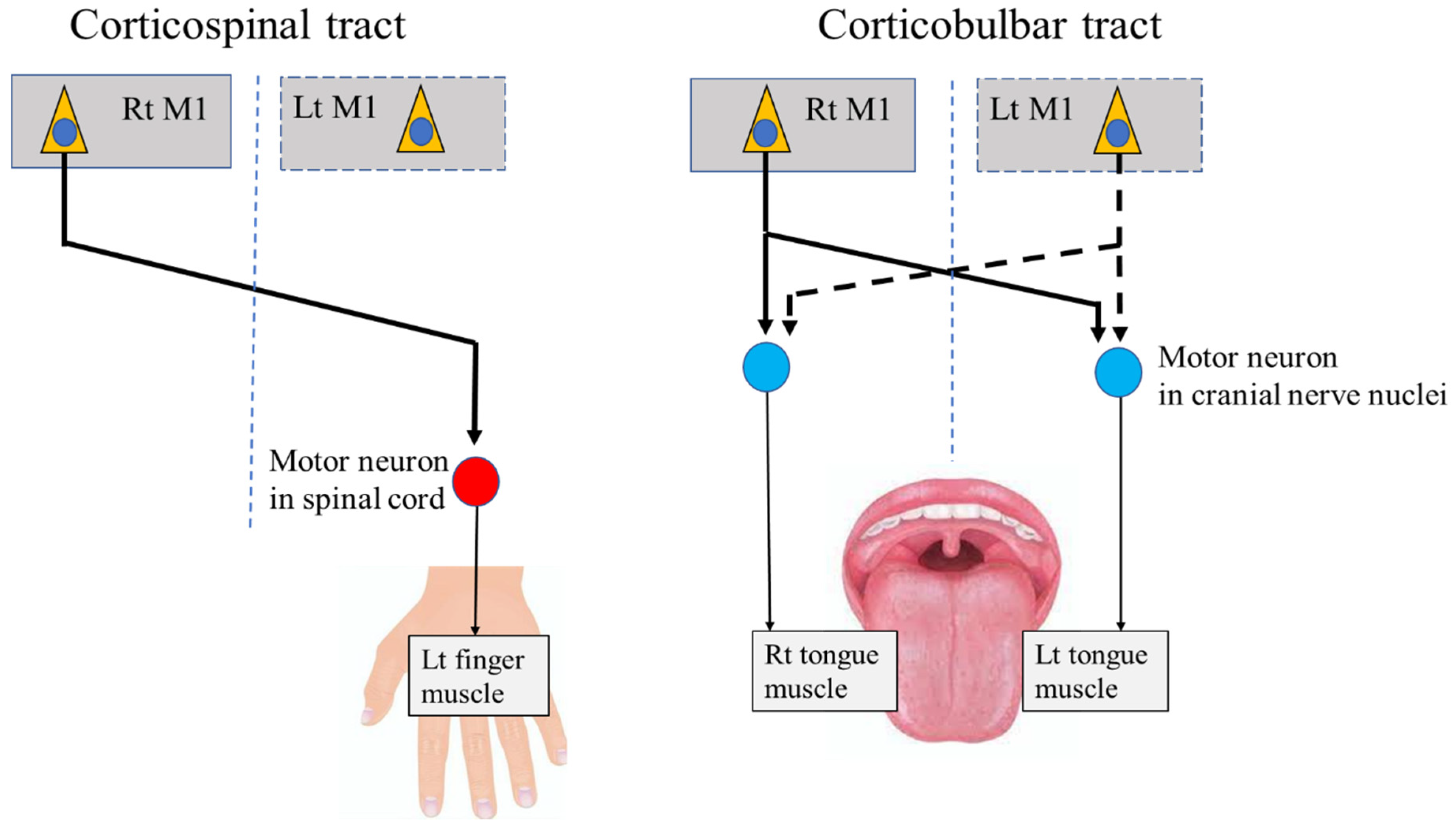
2. Neural Pathways from the Primary Motor Cortex (M1) to Stomatognathic Systems
3. Bilateral Brain Activation Related to Stomatognathic Functions
3.1. Movement-Related Cortical Fields and Potentials
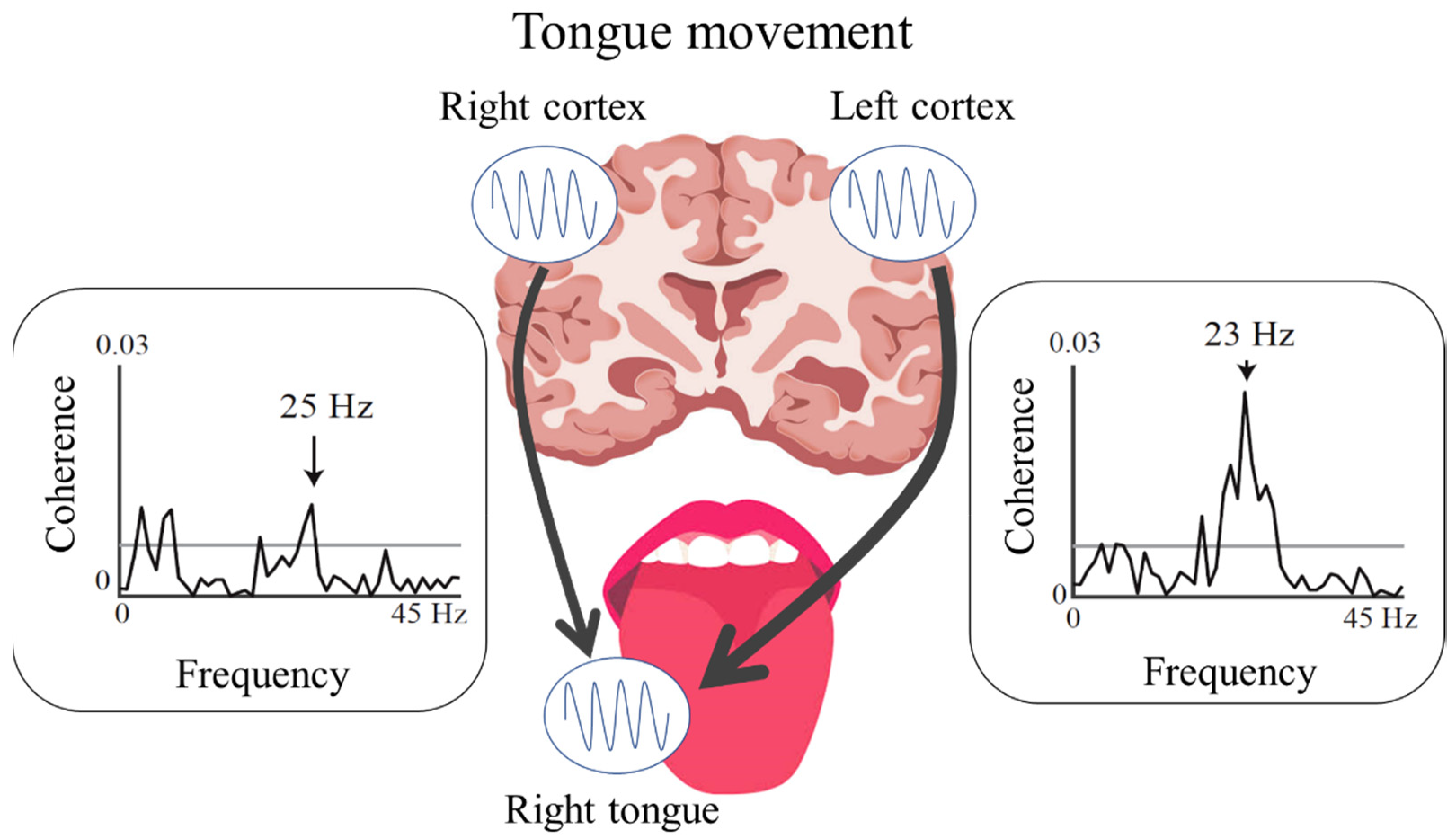
3.2. Cortico-Muscular Coherence and Cortico-Kinematic Coherence during Voluntary Movements
3.3. Somatosensory-Evoked Fields and Potentials
4. Oromandibular Dystonia
4.1. Pathophysiology of OMD
4.2. Treatment Options for OMD
5. Future Directions
6. Conclusions
7. Methods
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Comella, C.L. Systematic review of botulinum toxin treatment for oromandibular dystonia. Toxicon 2018, 147, 96–99. [Google Scholar] [CrossRef] [PubMed]
- Scorr, L.M.; Factor, S.A.; Parra, S.P.; Kaye, R.; Paniello, R.C.; Norris, S.A.; Perlmutter, J.S.; Bäumer, T.; Usnich, T.; Berman, B.D.; et al. Oromandibular dystonia: A clinical examination of 2020 cases. Front. Neurol. 2021, 12, 700714. [Google Scholar] [CrossRef]
- Yoshida, K. Botulinum toxin therapy for oromandibular dystonia and other movement disorders in the stomatognathic system. Toxins 2022, 14, 282. [Google Scholar] [CrossRef] [PubMed]
- Albanese, A.; Bhatia, K.; Bressman, S.B.; Delong, M.R.; Fahn, S.; Fung, V.S.; Hallett, M.; Jankovic, J.; Jinnah, H.A.; Klein, C.; et al. Phenomenology and classification of dystonia: A consensus update. Mov. Disord. 2013, 28, 863–873. [Google Scholar] [CrossRef] [PubMed]
- Colosimo, C.; Berardelli, A. Clinical phenomenology of dystonia. Int. Rev. Neurobiol. 2011, 98, 509–524. [Google Scholar] [CrossRef] [PubMed]
- Manzo, N.; Ginatempo, F.; Belvisi, D.; Defazio, G.; Conte, A.; Deriu, F.; Berardelli, A. Pathophysiological mechanisms of oromandibular dystonia. Clin. Neurophysiol. 2022, 134, 73–80. [Google Scholar] [CrossRef] [PubMed]
- Yoshida, K. Development and validation of a disease-specific oromandibular dystonia rating scale (OMDRS). Front. Neurol. 2020, 11, 583177. [Google Scholar] [CrossRef]
- Meige, H. Les convulsions de la face: Une forme clinique de convulsion faciale, bilatérale et médiane. Rev. Neurol. 1910, 10, 437–443. [Google Scholar]
- Weiss, E.M.; Hershey, T.; Karimi, M.; Racette, B.; Tabbal, S.D.; Mink, J.W.; Paniello, R.C.; Perlmutter, J.S. Relative risk of spread of symptoms among the focal onset primary dystonias. Mov. Disord. 2006, 21, 1175–1181. [Google Scholar] [CrossRef]
- Abbruzzese, G.; Berardelli, A.; Girlanda, P.; Marchese, R.; Martino, D.; Morgante, F.; Avanzino, L.; Colosimo, C.; Defazio, G. Long-term assessment of the risk of spread in primary late-onset focal dystonia. J. Neurol. Neurosurg. Psychiatry 2008, 79, 392–396. [Google Scholar] [CrossRef]
- Pandey, S.; Sharma, S. Meige’s syndrome: History, epidemiology, clinical features, pathogenesis and treatment. J. Neurol. Sci. 2017, 15, 162–170. [Google Scholar] [CrossRef] [PubMed]
- Pirio Richardson, S.; Wegele, A.R.; Skipper, B.; Deligtisch, A.; Jinnah, H.A.; Dystonia Coalition Investigators. Dystonia treatment: Patterns of medication use in an international cohort. Neurology 2017, 88, 543–550. [Google Scholar] [CrossRef] [PubMed]
- Steeves, T.D.; Day, L.; Dykeman, J.; Jette, N.; Pringsheim, T. The prevalence of primary dystonia: A systematic review and meta-analysis. Mov. Disord. 2012, 27, 1789–1796. [Google Scholar] [CrossRef] [PubMed]
- Nutt, J.G.; Muenter, M.D.; Aronson, A.; Kurland, L.T.; Melton, L.J. Epidemiology of focal and generalized dystonia in Rochester, Minnesota. Mov. Disord. 1988, 3, 188–194. [Google Scholar] [CrossRef]
- Cardoso, F.; Jankovic, J. Dystonia and dyskinesia. Psychiatr. Clin. N. Am. 1997, 20, 821–838. [Google Scholar] [CrossRef]
- Yoshida, K. Prevalence and incidence of oromandibular dystonia: An oral and maxillofacial surgery service-based study. Clin. Oral Investig. 2021, 25, 5755–5764. [Google Scholar] [CrossRef]
- Hallett, M.; Benecke, R.; Blitzer, A.; Comella, C.L. Treatment of focal dystonias with botulinum neurotoxin. Toxicon 2009, 54, 628–633. [Google Scholar] [CrossRef]
- Tisch, S. Deep brain stimulation in dystonia: Factors contributing to variability in outcome in short and long term follow-up. Curr. Opin. Neurol. 2022, 35, 510–517. [Google Scholar] [CrossRef]
- Schneider, S.A.; Aggarwal, A.; Bhatt, M.; Dupont, E.; Tisch, S.; Limousin, P.; Lee, P.; Quinn, N.; Bhatia, K.P. Severe tongue protrusion dystonia: Clinical syndromes and possible treatment. Neurology 2006, 67, 940–943. [Google Scholar] [CrossRef]
- Chung, J.C.; Kim, J.P.; Chang, W.S.; Kim, H.Y.; Chang, J.W. Bilateral pallidal stimulation for “sticking-out tongue” feature in patients with primary focal tongue protrusion dystonia. Neuromodulation 2014, 17, 133–137. [Google Scholar] [CrossRef]
- Wu, Y.; Pan, Y.; Huang, P.; Wang, T.; Zhang, C.; Sun, B.; Li, D.; Li, H.; Wu, Y. Subthalamic deep brain stimulation in lingual dystonia: A case series study. Park. Relat. Disord. 2021, 88, 114–115. [Google Scholar] [CrossRef] [PubMed]
- Asahi, T.; Ikeda, K.; Yamamoto, J.; Tsubono, H.; Muro, Y.; Sato, S. Bilateral pallidal stimulation with directional leads for primary focal lingual dystonia. Stereotact. Funct. Neurosurg. 2021, 99, 207–211. [Google Scholar] [CrossRef]
- Hari, R.; Salmelin, R. Magnetoencephalography: From SQUIDs to neuroscience. Neuroimage 20th anniversary special edition. Neuroimage 2012, 61, 386–396. [Google Scholar] [CrossRef] [PubMed]
- Penfield, W.; Boldrey, E. Somatic motor and sensory representation in the cerebral cortex of man as studied by electrical stimulation. Brain 1937, 60, 389–443. [Google Scholar] [CrossRef]
- Kubota, K.; Masegi, T. Muscle spindle supply to the human jaw muscle. J. Dent. Res. 1977, 56, 901–909. [Google Scholar] [CrossRef] [PubMed]
- Nakamura, A.; Yamada, T.; Goto, A.; Kato, T.; Ito, K.; Abe, Y.; Kachi, T.; Kakigi, R. Somatosensory homunculus as drawn by MEG. Neuroimage 1998, 7, 377–386. [Google Scholar] [CrossRef]
- Deecke, L.; Weinberg, H.; Brickett, P. Magnetic fields of the human brain accompanying voluntary movement: Bereitschaftsmagnetfeld. Exp. Brain Res. 1982, 48, 144–148. [Google Scholar] [CrossRef]
- Hari, R.; Antervo, A.; Katila, T.; Poutanen, T.; Seppänen, M.; Tuomisto, T.; Varpula, T. Cerebral magnetic fields associated with voluntary limb movements in man. Il Nuovo Cimento D 1983, 2, 484–494. [Google Scholar] [CrossRef]
- Cheyne, D.; Kristeva, R.; Deecke, L. Homuncular organization of human motor cortex as indicated by neuromagnetic recordings. Neurosci. Lett. 1991, 122, 17–20. [Google Scholar] [CrossRef]
- Nakasato, N.; Itoh, H.; Hatanaka, K.; Nakahara, H.; Kanno, A.; Yoshimoto, T. Movement-related magnetic fields to tongue protrusion. Neuroimage 2001, 14, 924–935. [Google Scholar] [CrossRef]
- Maezawa, H.; Oguma, H.; Hirai, Y.; Hisadome, K.; Shiraishi, H.; Funahashi, M. Movement-related cortical magnetic fields associated with self-paced tongue protrusion in humans. Neurosci. Res. 2017, 117, 22–27. [Google Scholar] [CrossRef] [PubMed]
- Nagamine, T.; Kajola, M.; Salmelin, R.; Shibasaki, H.; Hari, R. Movement-related slow cortical magnetic fields and changes of spontaneous MEG- and EEG-brain rhythms. Electroencephalogr. Clin. Neurophysiol. 1996, 99, 274–286. [Google Scholar] [CrossRef]
- Kornhuber, H.H.; Deecke, L. Changes in the brain potential in voluntary movements and passive movements in man: Readiness potential and reafferent potentials. Pflüger’s Arch. Gesamte Physiol. Menschen Tiere 1965, 284, 1–17. [Google Scholar] [CrossRef]
- Yoshida, K.; Kaji, R.; Hamano, T.; Kohara, N.; Kimura, J.; Shibasaki, H.; Iizuka, T. Cortical potentials associated with voluntary mandibular movements. J. Dent. Res. 2000, 79, 1514–1518. [Google Scholar] [CrossRef] [PubMed]
- Yoshida, K.; Kaji, R.; Kohara, N.; Murase, N.; Ikeda, A.; Shibasaki, H.; Iizuka, T. Movement-related cortical potentials before jaw excursions in oromandibular dystonia. Mov. Disord. 2003, 18, 94–100. [Google Scholar] [CrossRef]
- Shibukawa, Y.; Shintani, M.; Kumai, T.; Suzuki, T.; Nakamura, Y. Cortical neuromagnetic fields preceding voluntary jaw movements. J. Dent. Res. 2004, 83, 572–577. [Google Scholar] [CrossRef]
- Conway, B.A.; Halliday, D.M.; Farmer, S.F.; Shahani, U.; Maas, P.; Weir, A.I.; Rosenberg, J.R. Synchronization between motor cortex and spinal motoneuronal pool during the performance of a maintained motor task in man. J. Physiol. 1995, 489, 917–924. [Google Scholar] [CrossRef]
- Mima, T.; Hallett, M. Corticomuscular coherence: A review. J. Clin. Neurophysiol. 1999, 16, 501–511. [Google Scholar] [CrossRef]
- Maezawa, H.; Mima, T.; Yazawa, S.; Matsuhashi, M.; Shiraishi, H.; Hirai, Y.; Funahashi, M. Contralateral dominance of corticomuscular coherence for both sides of the tongue during human tongue protrusion: An MEG study. Neuroimage 2014, 101, 245–255. [Google Scholar] [CrossRef]
- Maezawa, H.; Mima, T.; Yazawa, S.; Matsuhashi, M.; Shiraishi, H.; Funahashi, M. Cortico-muscular synchronization by proprioceptive afferents from the tongue muscles during isometric tongue protrusion. Neuroimage 2016, 128, 284–292. [Google Scholar] [CrossRef]
- Bourguignon, M.; De Tiège, X.; Op de Beeck, M.; Pirotte, B.; Van Bogaert, P.; Goldman, S.; Hari, R.; Jousmäki, V. Functional motor-cortex mapping using corticokinematic coherence. Neuroimage 2011, 55, 1475–1479. [Google Scholar] [CrossRef] [PubMed]
- Bourguignon, M.; Piitulainen, H.; De Tiège, X.; Jousmäki, V.; Hari, R. Corticokinematic coherence mainly reflects movement-induced proprioceptive feedback. Neuroimage 2015, 106, 382–390. [Google Scholar] [CrossRef]
- Maezawa, H.; Fujimoto, M.; Hata, Y.; Matsuhashi, M.; Hashimoto, H.; Kashioka, H.; Yanagida, T.; Hirata, M. Functional cortical localization of tongue movements using corticokinematic coherence with a deep learning-assisted motion capture system. Sci. Rep. 2022, 12, 388. [Google Scholar] [CrossRef] [PubMed]
- Hari, R.; Kaukoranta, E.; Reinikainen, K.; Huopaniemie, T.; Mauno, J. Neuromagnetic localization of cortical activity evoked by painful dental stimulation in man. Neurosci. Lett. 1983, 42, 77–82. [Google Scholar] [CrossRef]
- Karhu, J.; Hari, R.; Lu, S.T.; Paetau, R.; Rif, J. Cerebral magnetic fields to lingual stimulation. Electroencephalogr. Clin. Neurophysiol. 1991, 80, 459–468. [Google Scholar] [CrossRef]
- Maezawa, H.; Yoshida, K.; Nagamine, T.; Matsubayashi, J.; Enatsu, R.; Bessho, K.; Fukuyama, H. Somatosensory evoked magnetic fields following electric tongue stimulation using pin electrodes. Neurosci. Res. 2008, 62, 131–139. [Google Scholar] [CrossRef]
- Sakamoto, K.; Nakata, H.; Kakigi, R. Somatosensory-evoked magnetic fields following stimulation of the tongue in humans. Clin. Neurophysiol. 2008, 119, 1664–1673. [Google Scholar] [CrossRef]
- Tamura, Y.; Shibukawa, Y.; Shintani, M.; Kaneko, Y.; Ichinohe, T. Oral structure representation in human somatosensory cortex. Neuroimage 2008, 43, 128–135. [Google Scholar] [CrossRef]
- Nakahara, H.; Nakasato, N.; Kanno, A.; Murayama, S.; Hatanaka, K.; Itoh, H.; Yoshimoto, T. Somatosensory-evoked fields for gingiva, lip, and tongue. J. Dent. Res. 2004, 83, 307–311. [Google Scholar] [CrossRef]
- Maezawa, H.; Matsuhashi, M.; Yoshida, K.; Mima, T.; Nagamine, T.; Fukuyama, H. Evaluation of lip sensory disturbance using somatosensory evoked magnetic fields. Clin. Neurophysiol. 2014, 125, 363–369. [Google Scholar] [CrossRef]
- Yoshida, K.; Maezawa, H.; Nagamine, T.; Fukuyama, H.; Murakami, K.; Iizuka, T. Somatosensory evoked magnetic fields to air-puff stimulation on the soft palate. Neurosci. Res. 2006, 55, 116–122. [Google Scholar] [CrossRef] [PubMed]
- Maezawa, H.; Hirai, Y.; Shiraishi, H.; Funahashi, M. Somatosensory evoked magnetic fields following tongue and hard palate stimulation on the preferred chewing side. J. Neurol. Sci. 2014, 347, 288–294. [Google Scholar] [CrossRef] [PubMed]
- Yamashita, H.; Kumamoto, Y.; Nakashima, T.; Yamamoto, T.; Inokuchi, A.; Komiyama, S. Magnetic sensory cortical responses evoked by tactile stimulations of the human face, oral cavity and flap reconstructions of the tongue. Eur. Arch. Otorhinolaryngol. 1999, 256 (Suppl. S1), S42–S46. [Google Scholar] [CrossRef] [PubMed]
- Ikeda, A.; Lüders, H.O.; Burgess, R.C.; Sakamoto, A.; Klem, G.H.; Morris, H.H.; Shibasaki, H. Generator locations of movement-related potentials with tongue protrusions and vocalizations: Subdural recording in human. Electroencephalogr. Clin. Neurophysiol. 1995, 96, 310–328. [Google Scholar] [CrossRef]
- Disbrow, E.A.; Hinkley, L.B.; Roberts, T.P. Ipsilateral representation of oral structures in human anterior parietal somatosensory cortex and integration of inputs across the midline. J. Comp. Neurol. 2003, 467, 487–495. [Google Scholar] [CrossRef] [PubMed]
- Maezawa, H.; Yoshida, K.; Matsuhashi, M.; Yokoyama, Y.; Mima, T.; Bessho, K.; Fujita, S.; Nagamine, T.; Fukuyama, H. Evaluation of tongue sensory disturbance by somatosensory evoked magnetic fields following tongue stimulation. Neurosci. Res. 2011, 71, 244–250. [Google Scholar] [CrossRef]
- Maezawa, H.; Tojyo, I.; Yoshida, K.; Fujita, S. Recovery of impaired somatosensory evoked fields after improvement of tongue sensory deficits with neurosurgical reconstruction. J. Oral Maxillofac. Surg. 2016, 74, 1473–1482. [Google Scholar] [CrossRef]
- Hallett, M.; Kanchana, S. Pathophysiology of dystonia. In Clinical Diagnosis and Management of Dystonia; CRC Press: Abingdon, UK, 2007; pp. 43–51. [Google Scholar]
- Vidailhet, M.; Grabli, D.; Roze, E. Pathophysiology of dystonia. Curr. Opin. Neurol. 2009, 22, 406–413. [Google Scholar] [CrossRef]
- Skogseid, I.M. Dystonia—New advances in classification, genetics, pathophysiology and treatment. Acta Neurol. Scand. Suppl. 2014, 198, 13–19. [Google Scholar] [CrossRef]
- Brüggemann, N. Contemporary functional neuroanatomy and pathophysiology of dystonia. J. Neural Transm. 2021, 128, 499–508. [Google Scholar] [CrossRef]
- Shibasaki, H.; Hallett, M. What is the Bereitschaftspotential? Clin. Neurophysiol. 2006, 117, 2341–2356. [Google Scholar] [CrossRef] [PubMed]
- Neshige, R.; Lüders, H.; Shibasaki, H. Recording of movement-related potentials from scalp and cortex in man. Brain 1988, 111, 719–736. [Google Scholar] [CrossRef] [PubMed]
- Ikeda, A.; Lüders, H.O.; Burgess, R.C.; Shibasaki, H. Movement-related potentials recorded from supplementary motor area and primary motor area. Role of supplementary motor area in voluntary movements. Brain 1992, 115, 1017–1043. [Google Scholar] [CrossRef] [PubMed]
- Shibasaki, H.; Sadato, N.; Lyshkow, H.; Yonekura, Y.; Honda, M.; Nagamine, T.; Suwazono, S.; Magata, Y.; Ikeda, A.; Miyazaki, M. Both primary motor cortex and supplementary motor area play an important role in complex finger movement. Brain 1993, 116, 1387–1398. [Google Scholar] [CrossRef]
- Fève, A.; Bathien, N.; Rondot, P. Abnormal movement related potentials in patients with lesions of basal ganglia and anterior thalamus. J. Neurol. Neurosurg. Psychiatry 1994, 57, 100–104. [Google Scholar] [CrossRef]
- Deuschl, G.; Toro, C.; Matsumoto, J.; Hallett, M. Movement-related cortical potentials in writer’s cramp. Ann. Neurol. 1995, 38, 862–868. [Google Scholar] [CrossRef]
- Yazawa, S.; Ikeda, A.; Kaji, R.; Terada, K.; Nagamine, T.; Toma, K.; Kubori, T.; Kimura, J.; Shibasaki, H. Abnormal cortical processing of voluntary muscle relaxation in patients with focal hand dystonia studied by movement-related potentials. Brain 1999, 122, 1357–1366. [Google Scholar] [CrossRef]
- Oga, T.; Honda, M.; Toma, K.; Murase, N.; Okada, T.; Hanakawa, T.; Sawamoto, N.; Nagamine, T.; Konishi, J.; Fukuyama, H.; et al. Abnormal cortical mechanisms of voluntary muscle relaxation in patients with writer’s cramp: An fMRI study. Brain 2002, 125, 895–903. [Google Scholar] [CrossRef]
- Kaji, R.; Ikeda, A.; Ikeda, T.; Kubori, T.; Mezaki, T.; Kohara, N.; Kanda, M.; Nagamine, T.; Honda, M.; Rothwell, J.C. Physiological study of cervical dystonia. Task-specific abnormality in contingent negative variation. Brain 1995, 118, 511–522. [Google Scholar] [CrossRef]
- Ikeda, A.; Shibasaki, H.; Kaji, R.; Terada, K.; Nagamine, T.; Honda, M.; Hamano, T.; Kimura, J. Abnormal sensorimotor integration in writer’s cramp: Study of contingent negative variation. Mov. Disord. 1996, 11, 683–690. [Google Scholar] [CrossRef]
- Murase, N.; Kaji, R.; Shimazu, H.; Katayama-Hirota, M.; Ikeda, A.; Kohara, N.; Kimura, J.; Shibasaki, H.; Rothwell, J.C. Abnormal premovement gating of somatosensory input in writer’s cramp. Brain 2000, 123, 1813–1829. [Google Scholar] [CrossRef] [PubMed]
- Tecchio, F.; Melgari, J.M.; Zappasodi, F.; Porcaro, C.; Milazzo, D.; Cassetta, E.; Rossini, P.M. Sensorimotor integration in focal task-specific hand dystonia: A magnetoencephalographic assessment. Neuroscience 2008, 154, 563–571. [Google Scholar] [CrossRef] [PubMed]
- Ridding, M.C.; Sheean, G.; Rothwell, J.C.; Inzelberg, R.; Kujirai, T. Changes in the balance between motor cortical excitation and inhibition in focal, task specific dystonia. J. Neurol. Neurosurg. Psychiatry. 1995, 59, 493–498. [Google Scholar] [CrossRef] [PubMed]
- Sohn, Y.H.; Hallett, M. Disturbed surround inhibition in focal hand dystonia. Ann. Neurol. 2004, 56, 595–599. [Google Scholar] [CrossRef]
- Münte, T.F.; Altenmüller, E.; Jäncke, L. The musician’s brain as a model of neuroplasticity. Nat. Rev. Neurosci. 2002, 3, 473–478. [Google Scholar] [CrossRef]
- Haslinger, B.; Erhard, P.; Dresel, C.; Castrop, F.; Roettinger, M.; Ceballos-Baumann, A.O. ‘Silent event-related’ fMRI reveals reduced sensorimotor activation in laryngeal dystonia. Neurology 2005, 65, 1562–1569. [Google Scholar] [CrossRef]
- Ceballos-Baumann, A.O.; Passingham, R.E.; Warner, T.; Playford, E.D.; Marsden, C.D.; Brooks, D.J. Overactive prefrontal and underactive motor cortical areas in idiopathic dystonia. Ann. Neurol. 1995, 37, 363–372. [Google Scholar] [CrossRef]
- Tempel, L.W.; Perlmutter, J.S. Abnormal vibration-induced cerebral blood flow responses in idiopathic dystonia. Brain 1990, 113, 691–707. [Google Scholar] [CrossRef]
- Tinazzi, M.; Rosso, T.; Fiaschi, A. Role of the somatosensory system in primary dystonia. Mov. Disord. 2003, 18, 605–622. [Google Scholar] [CrossRef]
- Hallett, M. Is dystonia a sensory disorder? Ann. Neurol. 1995, 38, 139–140. [Google Scholar] [CrossRef]
- Simpson, D.M.; Blitzer, A.; Brashear, A.; Comella, C.; Dubinsky, R.; Hallett, M.; Jankovic, J.; Karp, B.; Ludlow, C.L.; Miyasaki, J.M.; et al. Assessment: Botulinum neurotoxin for the treatment of movement disorders (an evidence-based review): Report of the Therapeutics and Technology Assessment Subcommittee of the American Academy of Neurology. Neurology 2008, 70, 1699–1706. [Google Scholar] [CrossRef] [PubMed]
- Jankovic, J. An update on new and unique uses of botulinum toxin in movement disorders. Toxicon 2018, 147, 84–88. [Google Scholar] [CrossRef] [PubMed]
- Anandan, C.; Jankovic, J. Botulinum toxin in movement disorders: An update. Toxins 2021, 13, 42. [Google Scholar] [CrossRef] [PubMed]
- Lee, H.J.; Kim, J.S.; Youn, K.H.; Lee, J.; Kim, H.-J. Ultrasound-guided botulinum neurotoxin type A injection for correcting asymmetrical smiles. Aesthet. Surg. J. 2018, 38, 130–134. [Google Scholar] [CrossRef] [PubMed]
- Wu, W.T.; Chang, K.V.; Chang, H.C.; Chen, L.R.; Kuan, C.H.; Kao, J.T.; Wei, L.Y.; Chen, Y.J.; Han, D.S.; Özçakar, L. Ultrasound imaging of the facial muscles and relevance with botulinum toxin injections: A pictorial essay and narrative review. Toxins 2022, 27, 101. [Google Scholar] [CrossRef]
- Rossetto, O.; Pirazzini, M.; Montecucco, C. Botulinum neurotoxins: Genetic, structural and mechanistic insights. Nat. Rev. Microbiol. 2014, 12, 535549. [Google Scholar] [CrossRef]
- Pirazzini, M.; Rossetto, O.; Eleopra, R.; Montecucco, C. Botulinum neurotoxins: Biology, pharmacology, and toxicology. Pharmacol. Rev. 2017, 69, 200235. [Google Scholar] [CrossRef]
- Luvisetto, S. Botulinum neurotoxins in central nervous system: An overview from animal models to human therapy. Toxins 2021, 13, 751. [Google Scholar] [CrossRef]
- Giladi, N. The mechanism of action of botulinum toxin type A in focal dystonia is most probably through its dual effect on efferent (motor) and afferent pathways at the injected site. J. Neurol. Sci. 1997, 152, 132135. [Google Scholar] [CrossRef]
- Hok, P.; Veverka, T.; Hlustik, P.; Nevrly, M.; Kanovsky, P. The central effects of botulinum toxin in dystonia and spasticity. Toxins 2021, 13, 155. [Google Scholar] [CrossRef]
- Weise, D.; Weise, C.M.; Naumann, M. Central effects of botulinum neurotoxin-Evidence from human studies. Toxins 2019, 11, 21. [Google Scholar] [CrossRef] [PubMed]
- Ghang, J.Y.; Lee, M.K.; Jun, S.M.; Ghang, C.G. Outcome of pallidal deep brain stimulation in Meige syndrome. J. Korean Neurosurg. Soc. 2010, 48, 134–138. [Google Scholar] [CrossRef] [PubMed]
- Lyons, M.K.; Birch, B.D.; Hillman, R.A.; Boucher, O.K.; Evidente, V.G. Long-term follow-up of deep brain stimulation for Meige syndrome. Neurosurg. Focus 2010, 29, E5. [Google Scholar] [CrossRef]
- Ostrem, J.L.; Marks, W.J.; Volz, M.M.; Heath, S.L.; Starr, P.A. Pallidal deep brain stimulation in patients with cranial-cervical dystonia (Meige syndrome). Mov. Disord. 2007, 22, 1885–1891. [Google Scholar] [CrossRef] [PubMed]
- Cury, R.G.; Kalia, S.K.; Shah, B.B.; Jimenez-Shahed, J.; Prashanth, L.K.; Moro, E. Surgical treatment of dystonia. Expert Rev. Neurother. 2018, 18, 477–492. [Google Scholar] [CrossRef] [PubMed]
- Kupsch, A.; Benecke, R.; Müller, J.; Trottenberg, T.; Schneider, G.H.; Poewe, W.; Eisner, W.; Wolters, A.; Müller, J.U.; Deuschl, G.; et al. Pallidal deep-brain stimulation in primary generalized or segmental dystonia. N. Engl. J. Med. 2006, 355, 1978–1990. [Google Scholar] [CrossRef] [PubMed]
- Albanese, A.; Asmus, F.; Bhatia, K.P.; Elia, A.E.; Elibol, B.; Filippini, G.; Gasser, T.; Krauss, J.K.; Nardocci, N.; Newton, A.; et al. EFNS guidelines on diagnosis and treatment of primary dystonias. Eur. J. Neurol. 2011, 18, 5–18. [Google Scholar] [CrossRef]
- Tagliati, M.; Krack, P.; Volkmann, J.; Aziz, T.; Krauss, J.K.; Kupsch, A.; Vidailhet, A.M. Long-term management of DBS in dystonia: Response to stimulation, adverse events, battery changes, and special considerations. Mov. Disord. 2011, 26 (Suppl. S1), S54–S62. [Google Scholar] [CrossRef]
- Horisawa, S.; Ochiai, T.; Goto, S.; Nakajima, T.; Takeda, N.; Kawamata, T.; Taira, T. Long-term outcome of pallidal stimulation for Meige syndrome. J. Neurosurg. 2018, 130, 84–89. [Google Scholar] [CrossRef]
- Sako, W.; Morigaki, R.; Mizobuchi, Y.; Tsuzuki, T.; Ima, H.; Ushio, Y.; Nagahiro, S.; Kaji, R.; Goto, S. Bilateral pallidal deep brain stimulation in primary Meige syndrome. Parkinsonism Relat. Disord. 2011, 17, 123–125. [Google Scholar] [CrossRef]
- Inoue, N.; Nagahiro, S.; Kaji, R.; Goto, S. Long-term suppression of Meige syndrome after pallidal stimulation: A 10-year follow-up study. Mov. Disord. 2010, 25, 1756–1758. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Zhang, Z.; Mao, Z.; Yu, X. Deep brain stimulation for Meige syndrome: A meta-analysis with individual patient data. J. Neurol. 2019, 266, 2646–2656. [Google Scholar] [CrossRef]
- Limotai, N.; Go, C.; Oyama, G.; Hwynn, N.; Zesiewicz, T.; Foote, K.; Bhidayasiri, R.; Malaty, I.; Zeilman, P.; Rodriguez, R.; et al. Mixed results for GPi-DBS in the treatment of cranio-facial and cranio-cervical dystonia symptoms. J. Neurol. 2011, 258, 2069–2074. [Google Scholar] [CrossRef] [PubMed]
- Vidailhet, M.; Jutras, M.F.; Grabli, D.; Roze, E. Deep brain stimulation for dystonia. J. Neurol. Neurosurg. Psychiatry 2013, 84, 1029–1042. [Google Scholar] [CrossRef] [PubMed]
- Cif, L.; Demailly, D.; Lin, J.P.; Barwick, K.E.; Sa, M.; Abela, L.; Malhotra, S.; Chong, W.K.; Steel, D.; Sanchis-Juan, A.; et al. KMT2B-related disorders: Expansion of the phenotypic spectrum and long-term efficacy of deep brain stimulation. Brain 2020, 143, 3242–3261. [Google Scholar] [CrossRef]
- Yoshida, K. Botulinum neurotoxin therapy for lingual dystonia using an individualized injection method based on clinical features. Toxins 2019, 11, 51. [Google Scholar] [CrossRef]
- Frucht, S.; Fahn, S.; Ford, B.; Gelb, M. A geste antagoniste device to treat jaw-closing dystonia. Mov. Disord. 1999, 14, 883–886. [Google Scholar] [CrossRef]
- Lo, S.E.; Gelb, M.; Frucht, S.J. Geste antagonistes in idiopathic lower cranial dystonia. Mov. Disord. 2007, 22, 1012–1017. [Google Scholar] [CrossRef]
- Satoh, M.; Narita, M.; Tomimoto, H. Three cases of focal embouchure dystonia: Classifications and successful therapy using a dental splint. Eur. Neurol. 2011, 66, 85–90. [Google Scholar] [CrossRef]
- Yoshida, K. Sensory trick splint as a multimodal therapy for oromandibular dystonia. J. Prosthodont. Res. 2018, 62, 239–244. [Google Scholar] [CrossRef]
- De Meyer, M.; Vereecke, L.; Bottenberg, P.; Jacquet, W.; Sims, A.B.; Santens, P. Oral appliances in the treatment of oromandibular dystonia: A systematic review. Acta Neurol. Belg. 2020, 120, 831–836. [Google Scholar] [CrossRef] [PubMed]
- Gonzalez-Alegre, P.; Schneider, R.L.; Hoffman, H. Clinical, etiological, and therapeutic features of jaw-opening and jaw-closing oromandibular dystonias: A decade of experience at a single treatment center. Tremor Other Hyperkinet. Mov. 2014, 4, 231. [Google Scholar] [CrossRef]
- Schramm, A.; Classen, J.; Reiners, K.; Naumann, M. Characteristics of sensory trick-like manoeuvres in jaw-opening dystonia. Mov. Disord. 2007, 22, 430–433. [Google Scholar] [CrossRef] [PubMed]
- Sankhla, C.; Lai, E.C.; Jankovic, J. Peripherally induced oromandibular dystonia. J. Neurol. Neurosurg. Psychiatry 1998, 65, 722–728. [Google Scholar] [CrossRef] [PubMed]
- Schiefer, T.K.; Matsumoto, J.Y.; Lee, K.H. Moving forward: Advances in the treatment of movement disorders with deep brain stimulation. Front. Integr. Neurosci. 2011, 5, 69. [Google Scholar] [CrossRef]
- Marceglia, S.; Mrakic-Sposta, S.; Fumagalli, M.; Ferrucci, R.; Mameli, F.; Vergari, M.; Barbieri, S.; Priori, A. Cathodal transcranial direct current stimulation improves focal hand dystonia in musicians: A two-case study. Front. Neurosci. 2017, 11, 508. [Google Scholar] [CrossRef]
- Young, S.J.; Bertucco, M.; Sheehan-Stross, R.; Sanger, T.D. Cathodal transcranial direct current stimulation in children with dystonia: A pilot open-label trial. J. Child Neurol. 2013, 28, 1238–1244. [Google Scholar] [CrossRef]
- Brittain, J.S.; Probert-Smith, P.; Aziz, T.Z.; Brown, P. Tremor suppression by rhythmic transcranial current stimulation. Curr. Biol. 2013, 23, 436–440. [Google Scholar] [CrossRef]
- Buttkus, F.; Weidenmüller, M.; Schneider, S.; Jabusch, H.C.; Nitsche, M.A.; Paulus, W.; Altenmüller, E. Failure of cathodal direct current stimulation to improve fine motor control in musician’s dystonia. Mov. Disord. 2010, 25, 389–394. [Google Scholar] [CrossRef]
- Benninger, D.H.; Lomarev, M.; Lopez, G.; Pal, N.; Luckenbaugh, D.A.; Hallett, M. Transcranial direct current stimulation for the treatment of focal hand dystonia. Mov. Disord. 2011, 26, 1698–1702. [Google Scholar] [CrossRef]
- Bhanpuri, N.H.; Bertucco, M.; Young, S.J.; Lee, A.A.; Sanger, T.D. Multiday transcranial direct current stimulation causes clinically insignificant changes in childhood dystonia: A pilot study. J. Child Neurol. 2015, 30, 1604–1615. [Google Scholar] [CrossRef] [PubMed]
- Maezawa, H.; Vicario, C.M.; Kuo, M.F.; Hirata, M.; Mima, T.; Nitsche, M.A. Effects of bilateral anodal transcranial direct current stimulation over the tongue primary motor cortex on cortical excitability of the tongue and tongue motor functions. Brain Stimul. 2020, 13, 270–272. [Google Scholar] [CrossRef] [PubMed]


Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Maezawa, H.; Hirata, M.; Yoshida, K. Neurophysiological Basis of Deep Brain Stimulation and Botulinum Neurotoxin Injection for Treating Oromandibular Dystonia. Toxins 2022, 14, 751. https://doi.org/10.3390/toxins14110751
Maezawa H, Hirata M, Yoshida K. Neurophysiological Basis of Deep Brain Stimulation and Botulinum Neurotoxin Injection for Treating Oromandibular Dystonia. Toxins. 2022; 14(11):751. https://doi.org/10.3390/toxins14110751
Chicago/Turabian StyleMaezawa, Hitoshi, Masayuki Hirata, and Kazuya Yoshida. 2022. "Neurophysiological Basis of Deep Brain Stimulation and Botulinum Neurotoxin Injection for Treating Oromandibular Dystonia" Toxins 14, no. 11: 751. https://doi.org/10.3390/toxins14110751
APA StyleMaezawa, H., Hirata, M., & Yoshida, K. (2022). Neurophysiological Basis of Deep Brain Stimulation and Botulinum Neurotoxin Injection for Treating Oromandibular Dystonia. Toxins, 14(11), 751. https://doi.org/10.3390/toxins14110751