Combined Action of Shiga Toxin Type 2 and Subtilase Cytotoxin in the Pathogenesis of Hemolytic Uremic Syndrome

 ,
, 
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
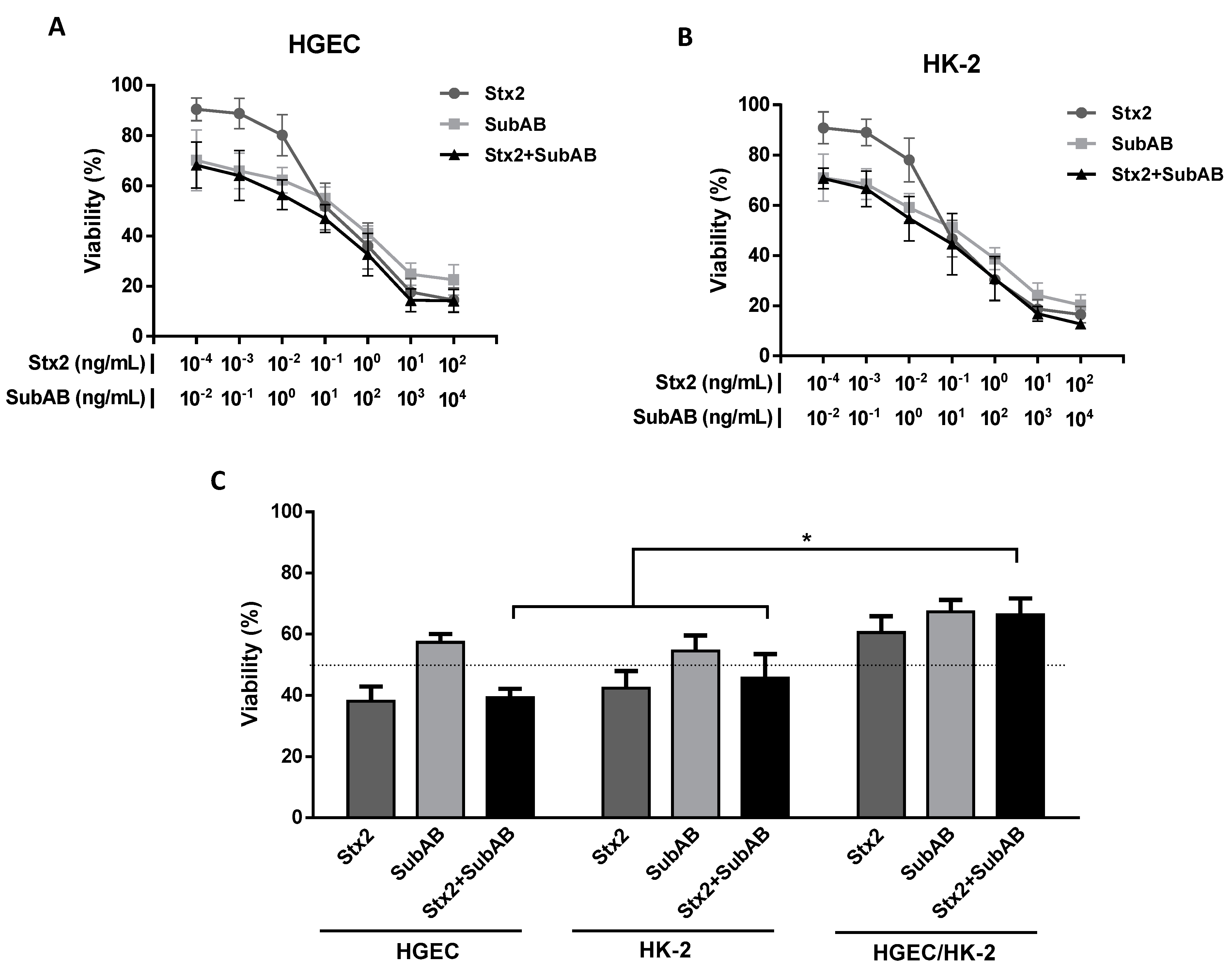
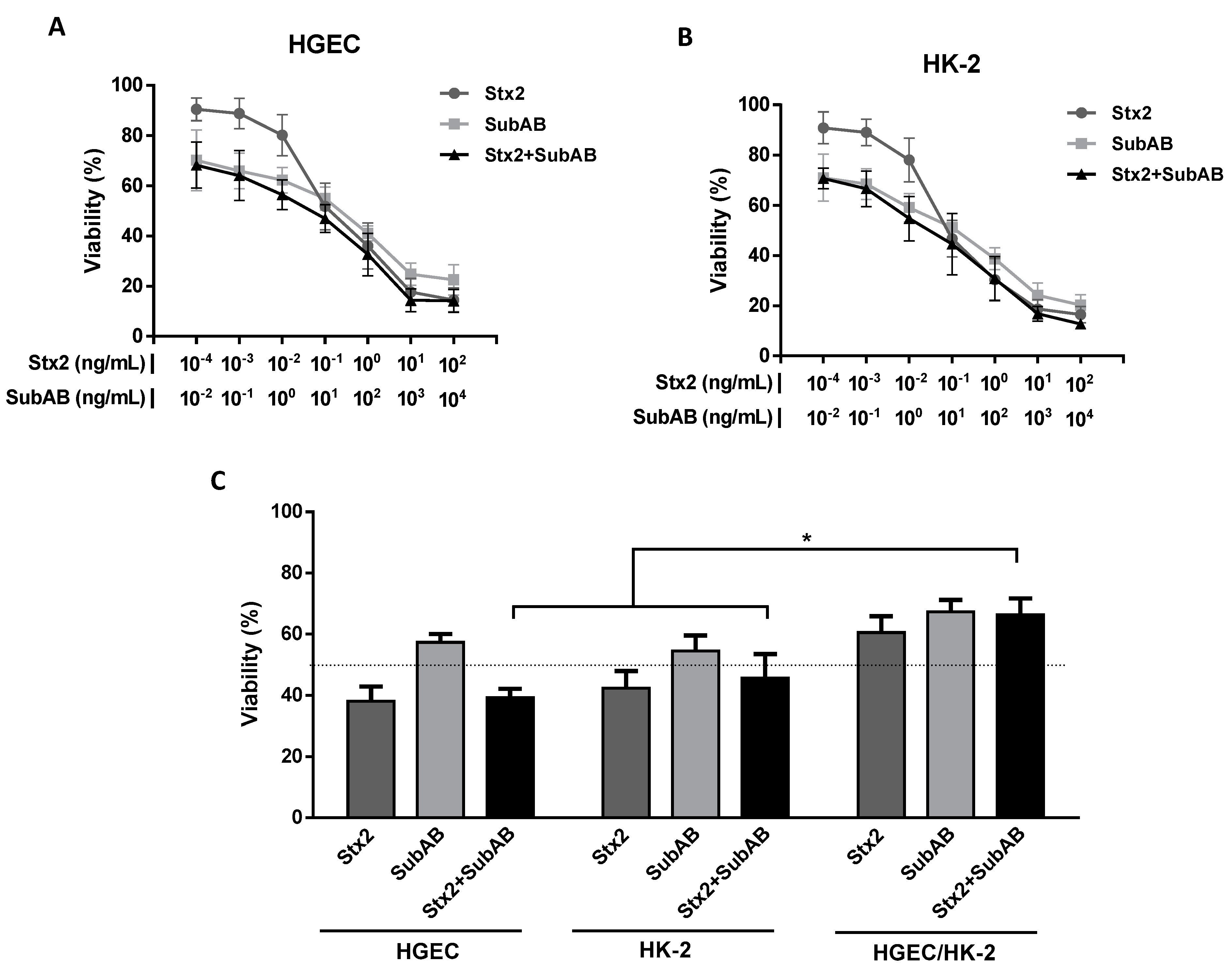
2.1. Co-Incubation (Stx2 + SubAB) Reduced In Vitro Cell Viability without Additive or Synergistic Effects
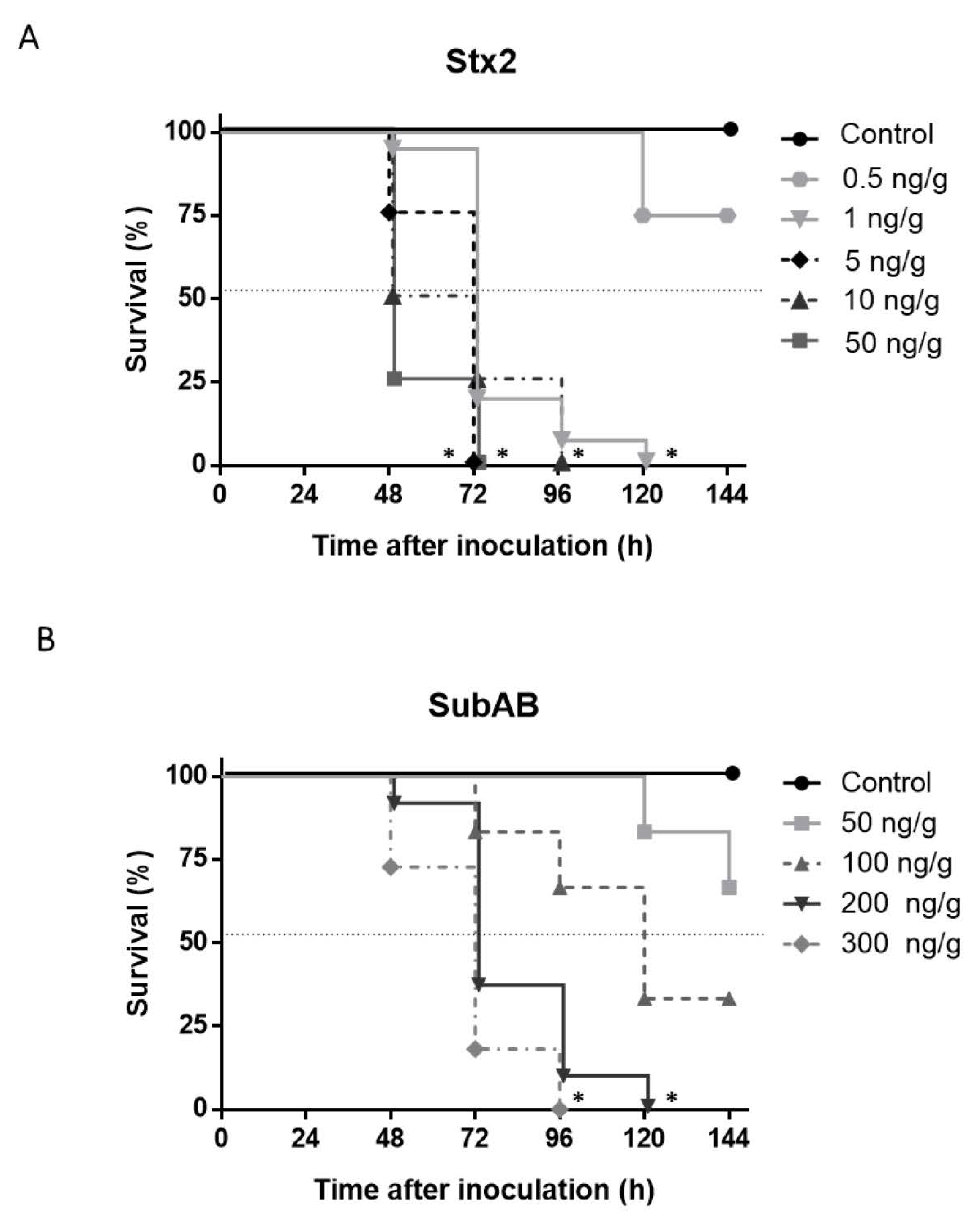
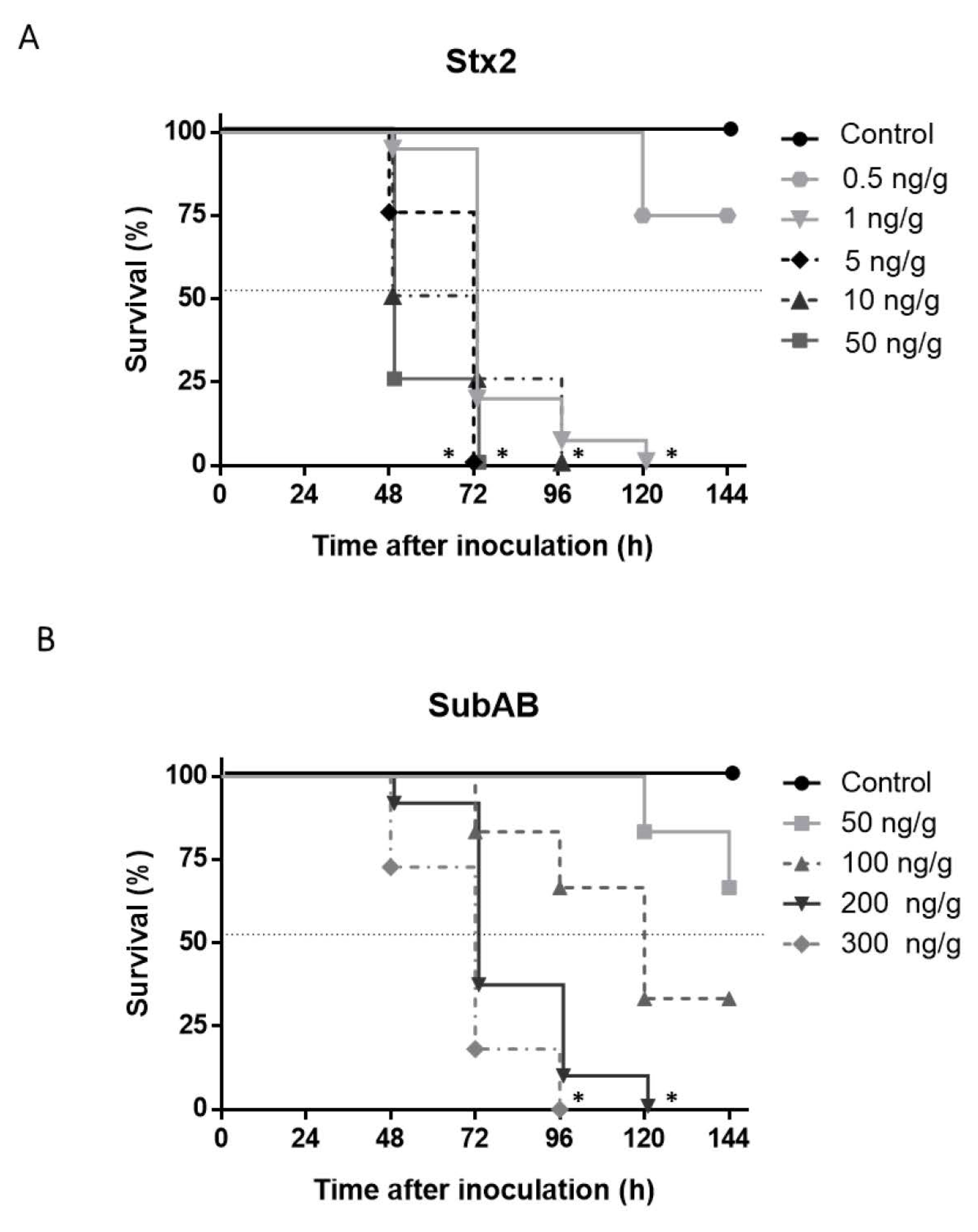
2.2. Stx2 and SubAB Minimum Lethal Dose
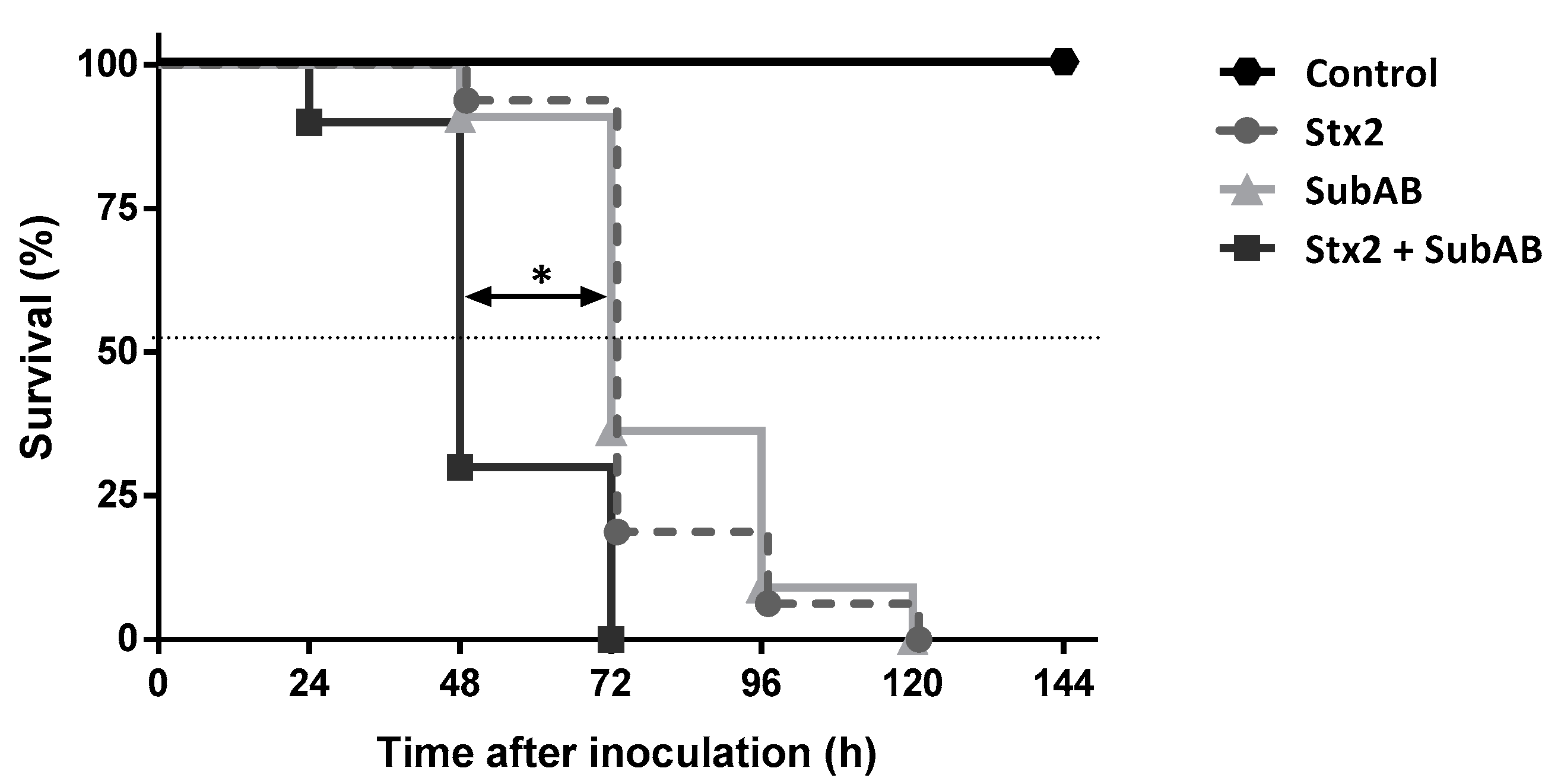
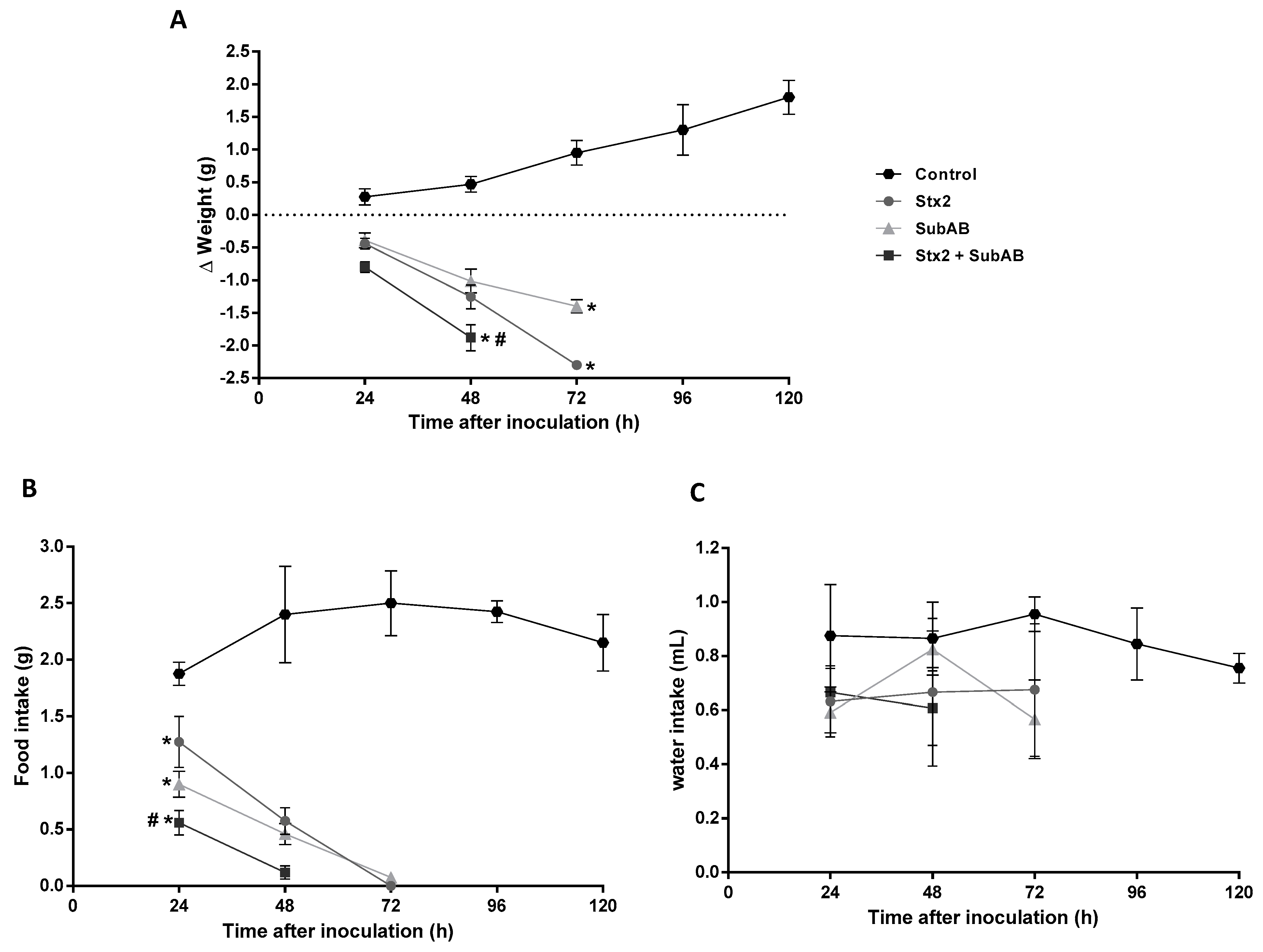
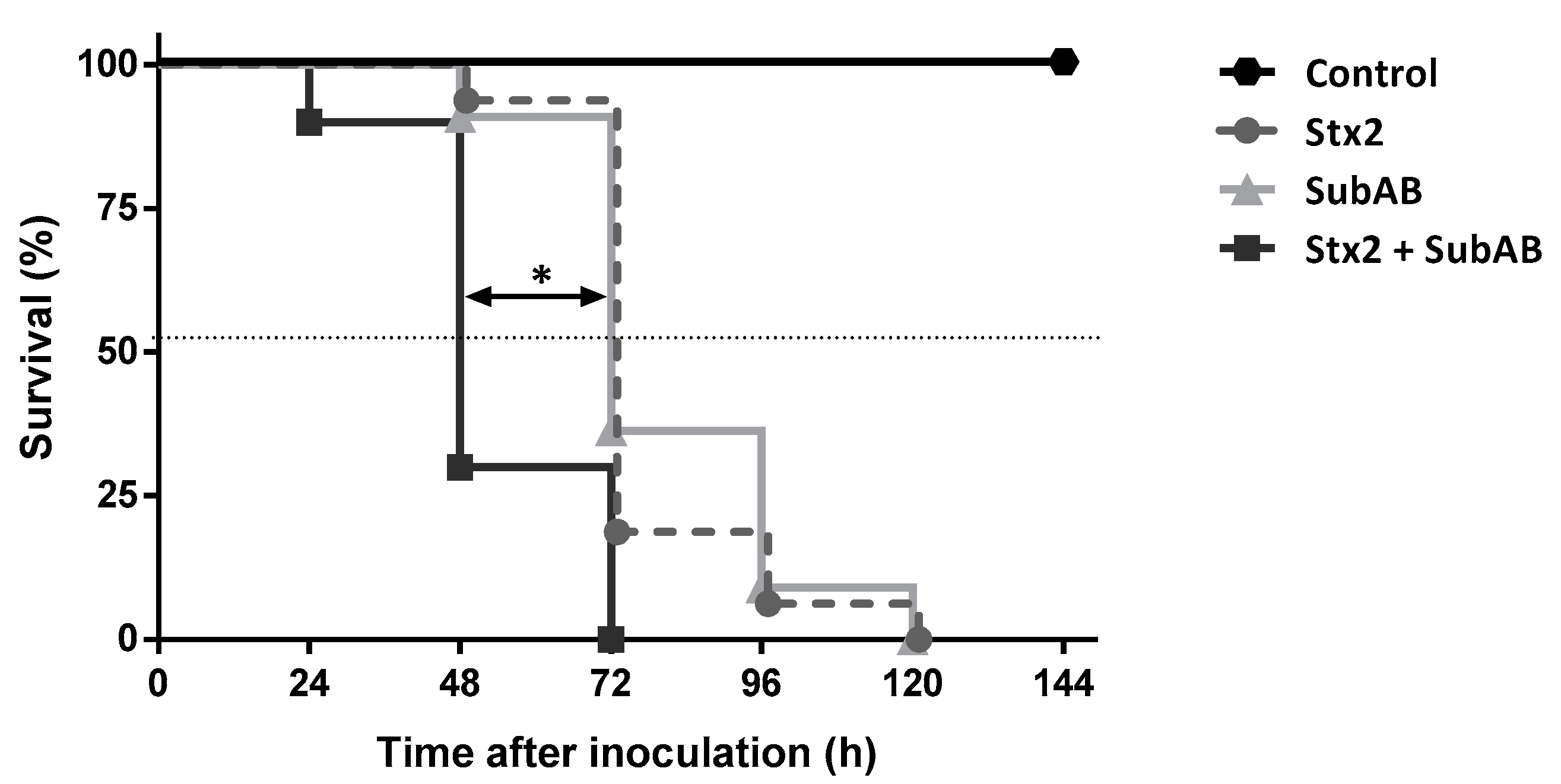
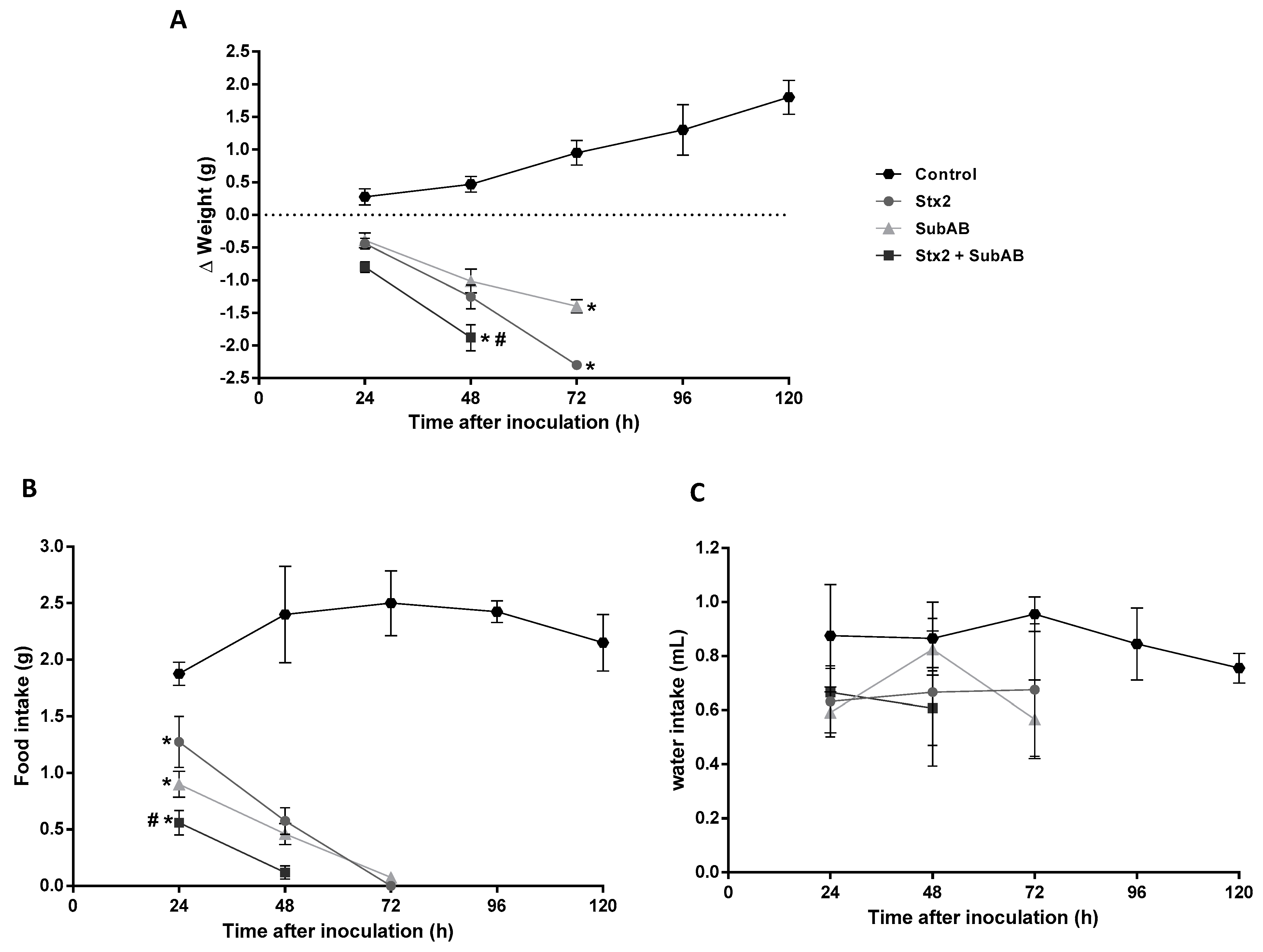
2.3. The Co-Treatment Decreased the Survival Time and the Body Weight in Mice
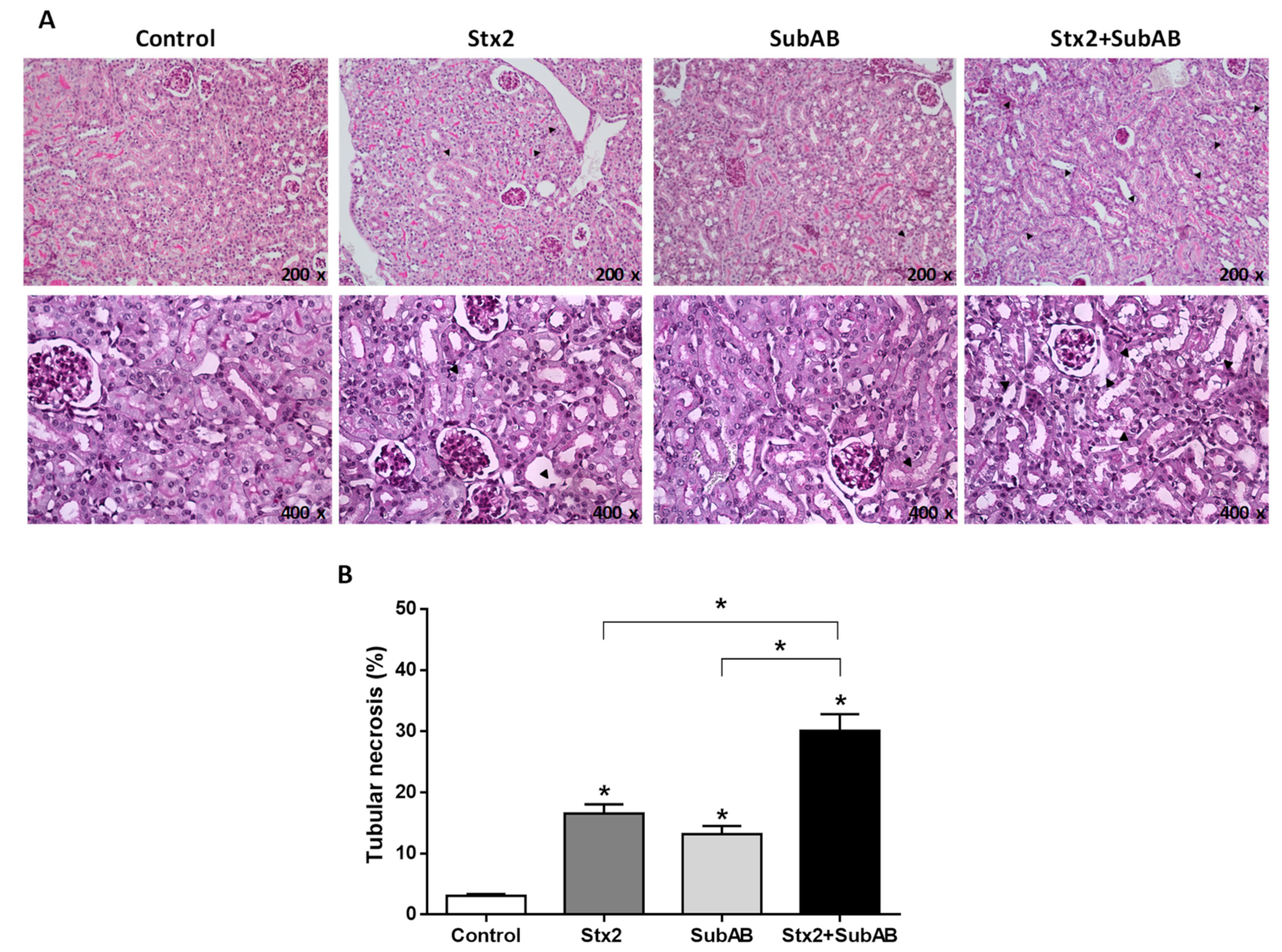
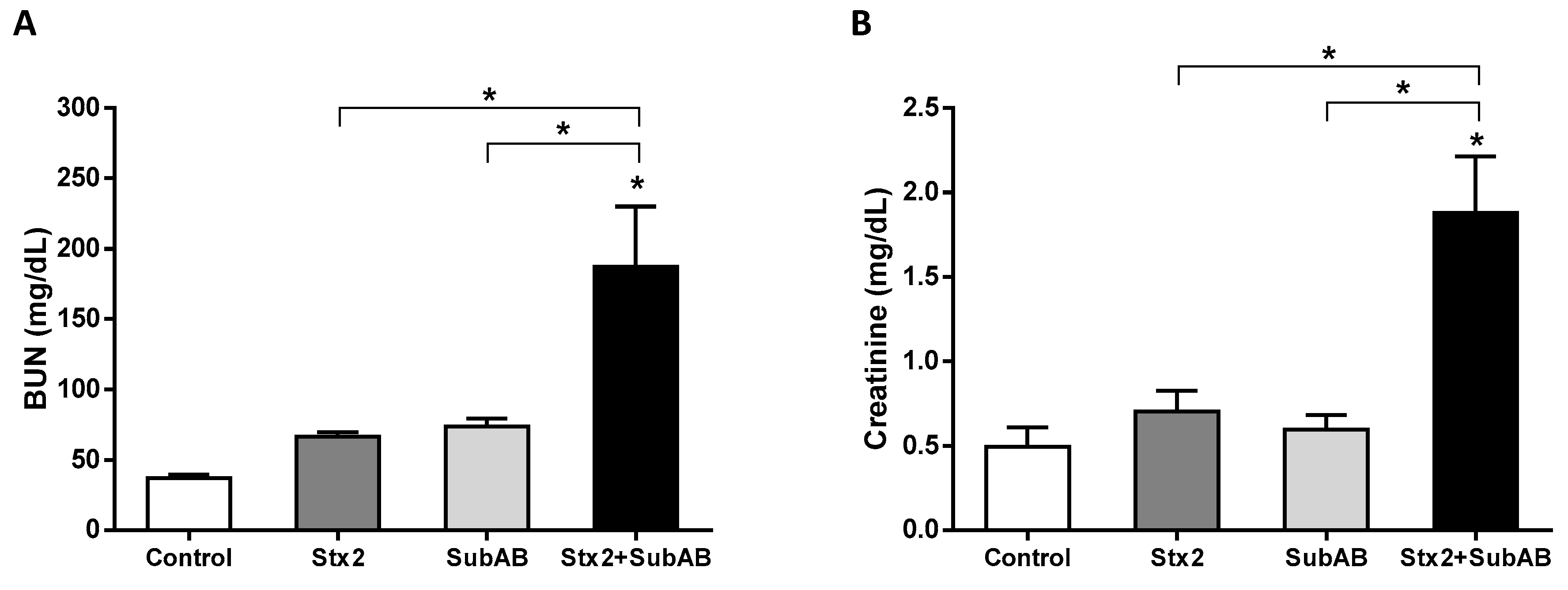
2.4. Renal Damage and Levels of Urea in Serum Are Increased by the Co-Treatment
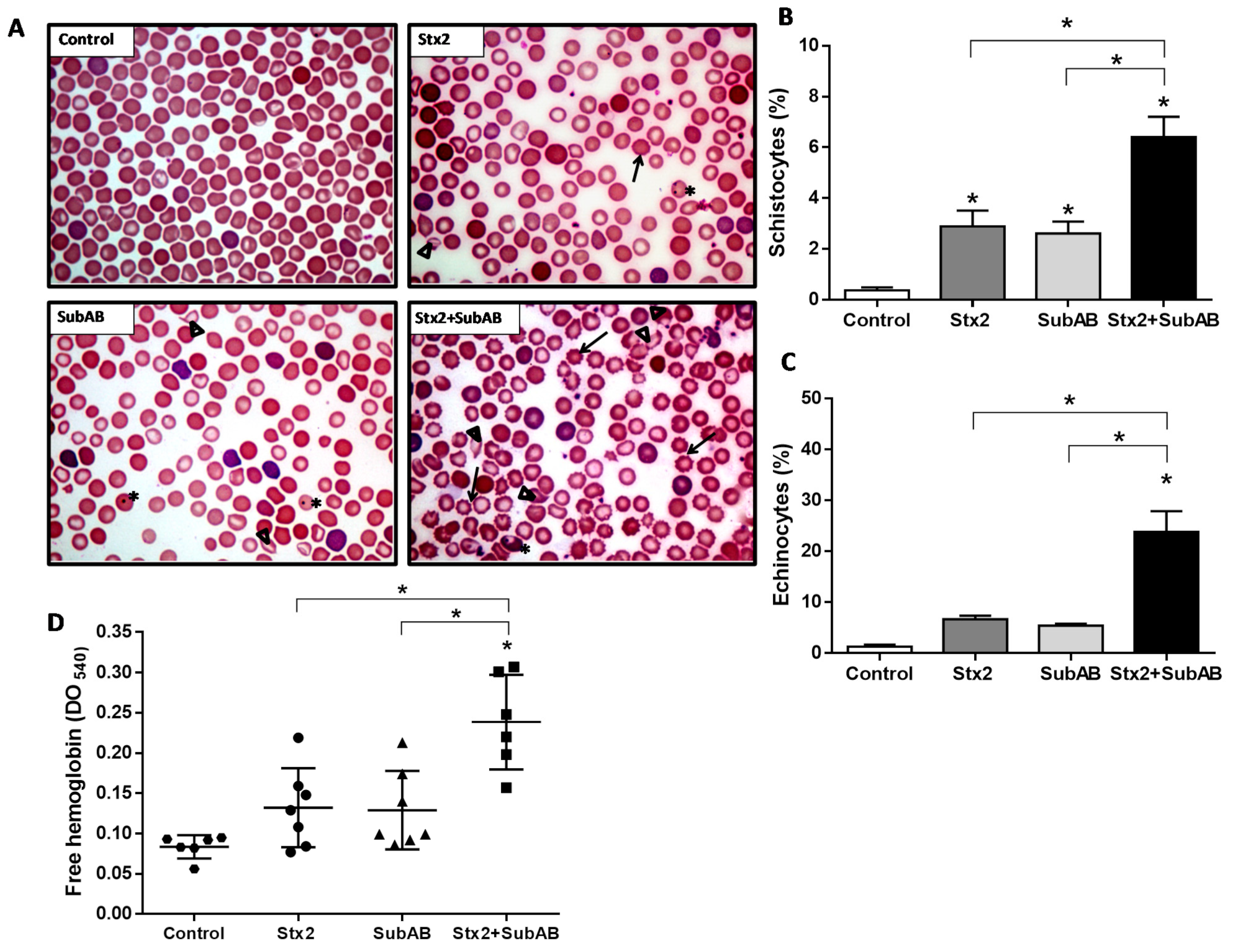
2.5. Co-Treatment Increases Damaged Erythrocytes and Reticulocytes
3. Discussion
4. Conclusions
5. Materials and Methods
5.1. Reagents
5.2. Human Primary Glomerular Microvascular Endothelial Cell Culture
5.3. Tubular Human Epithelial Cell Line Culture
5.4. Monocultures and Co-Cultures of Renal Endothelial and Epithelial Cells
5.5. Cell Viability Evaluation
5.6. Animals
5.7. Setting of Stx2 and SubAB Minimum Lethal Dose
5.8. Evaluation of Mouse Survival and General Status
5.9. Blood Samples and Renal Tissue Collection
5.10. Renal Histoarchitecture
5.11. Renal Function
5.12. Hematological Studies
5.12.1. Blood Smear
5.12.2. Erythrocyte Morphology
5.12.3. Free Hemoglobin
5.13. Data Analysis
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Repetto, H.A. Epidemic hemolytic-uremic syndrome in children. Kidney Int. 1997, 52, 1708–1719. [Google Scholar] [CrossRef] [Green Version]
- Griffin, P.M.; Tauxe, R.V. The epidemiology of infections caused by Escherichia coli O157:H7, other enterohemorrhagic E. coli, and the associated hemolytic uremic syndrome. Epidemiol. Rev. 1991, 13, 60–98. [Google Scholar] [CrossRef] [PubMed]
- Pianciola, L.; Rivas, M. Genotypic Features of Clinical and Bovine Escherichia coli O157 Strains Isolated in Countries with Different Associated-Disease Incidences. Microorganisms 2018, 6, 36. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Alconcher, L.F.; Coccia, P.A.; Suarez, A.; Monteverde, M.L.; y Gutiérrez, M.G.P.; Carlopio, P.M.; Missoni, M.L.; Balestracci, A.; Principi, I.; Ramírez, F.B.; et al. Hyponatremia: A new predictor of mortality in patients with Shiga toxin-producing Escherichia coli hemolytic uremic syndrome. Pediatr. Nephrol. 2018, 33, 1791–1798. [Google Scholar] [CrossRef] [PubMed]
- Alconcher, L.F.; Rivas, M.; Lucarelli, L.I.; Galavotti, J.; Rizzo, M. Shiga toxin-producing Escherichia coli in household members of children with hemolytic uremic syndrome. Eur. J. Clin. Microbiol. Infect. Dis. 2020, 39, 427–432. [Google Scholar] [CrossRef]
- Ministerio de Salud de la Nación Argentina. Boletín Integrado de Vigilancia. 2019. Available online: https://bancos.salud.gob.ar/recurso/boletin-integrado-de-vigilancia-n477-se50-19122019 (accessed on 5 May 2021).
- Repetto, H.A.; Rodríguez de Córdoba, E.; Arrizurieta, E.; Rivas, M.; y Ibarra, C. Microangiopatía trombótica y sindrome hemolítico urémico. In Nefrología Clínica, 3rd ed.; Editorial Médica Panamericana: Buenos Aires, Argentina, 2014; Chapter 25; pp. 352–363. [Google Scholar]
- Luna, M.; Kamariski, M.; Princip, I.; Bocanegra, V.; Vallés, P.G. Severely ill pediatric patients with Shiga toxin-associated hemolytic uremic syndrome (STEC-HUS) who suffered from multiple organ involvement in the early stage. Pediatr. Nephrol. 2020, 36, 1499–1509. [Google Scholar] [CrossRef]
- Weinstein, D.L.; Jackson, M.P.; Samuel, J.E.; Holmes, R.K.; O’Brien, A.D. Cloning and sequencing of a Shiga-like toxin type II variant from Escherichia coli strain responsible for edema disease of swine. J. Bacteriol. 1988, 170, 4223–4230. [Google Scholar] [CrossRef] [Green Version]
- Friedrich, A.W.; Borell, J.; Bielaszewska, M.; Fruth, A.; Tschäpe, H.; Karch, H. Shiga toxin 1c-producing Escherichia coli strains: Phenotypic and genetic characterization and association with human disease. J. Clin. Microbiol. 2003, 41, 2448–2453. [Google Scholar] [CrossRef] [Green Version]
- Kumar, A.; Taneja, N.; Kumar, Y.; Sharma, M. Detection of Shiga toxin variants among Shiga toxin-forming Escherichia coli isolates from animal stool, meat and human stool samples in India. J. Appl. Microbiol. 2012, 113, 1208–1216. [Google Scholar] [CrossRef]
- Probert, W.S.; McQuaid, C.; Schraderet, K. Isolation and identification of an Enterobacter cloacae strain producing a novel subtype of Shiga toxin type 1. J. Clin. Microbiol. 2014, 52, 2346–2351. [Google Scholar] [CrossRef] [Green Version]
- Yang, X.; Bai, X.; Zhange, J.; Sun, H.; Fu, S.; Fan, R.; He, X.; Scheutz, F.; Matussek, A.; Xiong, Y. Escherichia coli strains producing a novel Shiga toxin 2 subtype circulate in China. Int. J. Med. Microbiol. 2020, 310, 151377. [Google Scholar] [CrossRef] [PubMed]
- EFSA Biohaz Panel; Koutsoumanis, K.; Allende, A.; Alvarez-Ordóñez, A.; Bover-Cid, S.; Chemaly, M.; Davies, R.; De Cesare, A.; Herman, L.; Hilbert, F.; et al. Pathogenicity assessment of Shiga toxin-producing Escherichia coli (STEC) and the public health risk posed by contamination of food with STEC. EFSA J. 2020, 18, 5967–6071. [Google Scholar] [CrossRef]
- Rivas, M.; Miliwebsky, E.; Chinen, I.; Deza, N.; Leotta, G.A. The epidemiology of hemolytic uremic syndrome in Argentina. Diagnosis of the etiologic agent, reservoirs and routes of transmission. Medicina 2006, 66, 27–32. [Google Scholar] [PubMed]
- Fitzgerald, S.F.; Beckett, A.E.; Palarea-Albaladejo, J.; McAteer, S.; Shaaban, S.; Morgan, J.; Ahmad, N.I.; Young, R.; Mabbott, N.A.; Morrison, L.; et al. Shiga toxin sub-type 2a increases the efficiency of Escherichia coli O157 transmission between animals and restricts epithelial regeneration in bovine enteroids. PLoS Pathog. 2019, 15, e1008003. [Google Scholar] [CrossRef] [Green Version]
- Paton, A.W.; Srimanote, P.; Talbot, U.M.; Wang, H.; Paton, J.C. A new family of potent AB (5) cytotoxins produced by Shiga toxigenic Escherichia coli. J. Exp. Med. 2004, 200, 35–46. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Michelacci, V.; Tozzoli, R.; Caprioli, A.; Martínez, R.; Scheutz, F.; Grande, L.; Sánchez, S.; Morabito, S. A new pathogenicity island carrying an allelic variant of the subtilase cytotoxin is common among Shiga toxin producing Escherichia coli of human and ovine origin. Clin. Microbiol. Infect. 2013, 19, 149–156. [Google Scholar] [CrossRef] [Green Version]
- Orden, J.A.; Horcajo, P.; De la Fuente, R.; Ruiz-Santa-Quiteria, J.A.; Domínguez-Bernal, G.; Carrión, J. Subtilase cytotoxin-coding genes in verotoxin producing Escherichia coli strains from sheep and goats differ from those from cattle. Appl. Environ. Microbiol. 2011, 77, 8259–8264. [Google Scholar] [CrossRef] [Green Version]
- Funk, J.; Stoeber, H.; Hauser, E.; Schmidt, H. Molecular analysis of subtilase cytotoxin genes of food-borne Shiga toxin-producing Escherichia coli reveals a new allelic subAB variant. BMC Microbiol. 2013, 13, 230. [Google Scholar] [CrossRef] [Green Version]
- Tasara, T.; Fierz, L.; Klumppet, J.; Schmidt, H.; Stephan, R. Draft Genome Sequences of Five Shiga Toxin-Producing Escherichia coli Isolates Harboring the New and Recently Described Subtilase Cytotoxin Allelic Variant subAB 2–3. Genome Announc. 2017, 5, e01582-16. [Google Scholar] [CrossRef] [Green Version]
- Fraser, M.E.; Fujinaga, M.; Cherney, M.M.; Melton-Celsa, A.R.; Twiddy, E.M.; O’Brien, A.D.; James, M.N. Structure of shiga toxin type 2 (stx2) from Escherichia coli O157:H7. J. Biol. Chem. 2004, 279, 27511–27517. [Google Scholar] [CrossRef] [Green Version]
- Jacewicz, M.; Clausen, H.; Nudelman, E.; Donohue-Rolfe, A.; Keusch, G.T. Pathogenesis of shigella diarrhea. XI. Isolation of a shigella toxin-binding glycolipid from rabbit jejunum and hela cells and its identification as globotriaosylceramide. J. Exp. Med. 1986, 163, 1391–1404. [Google Scholar] [CrossRef] [PubMed]
- Lee, M.S.; Kim, M.H.; Tesh, V.L. Shiga toxins expressed by human pathogenic bacteria induce immune responses in host cells. J. Microbiol. 2013, 51, 724–730. [Google Scholar] [CrossRef] [PubMed]
- Byres, E.; Paton, A.W.; Paton, J.C.; Löfling, J.C.; Smith, D.F.; Wilce, M.C.; Talbot, U.M.; Chong, D.C.; Yu, H.; Huang, S.; et al. Incorporation of a non-human glycan mediates human susceptibility to a bacterial toxin. Nature 2008, 456, 648–652. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Paton, A.W.; Beddoe, T.; Thorpee, C.M.; Whisstock, J.C.; Wilce, M.C.; Rossjohn, J.; Talbot, U.M.; Paton, J.C. AB5 subtilase cytotoxin inactivates the endoplasmic reticulum chaperone BiP. Nature 2006, 443, 548–552. [Google Scholar] [CrossRef]
- Wolfson, J.J.; May, K.L.; Thorpe, C.M.; Jandhyala, D.M.; Paton, J.C.; Paton, A.W. Subtilase cytotoxin activates PERK, IRE1 and ATF6 endoplasmic reticulum stress-signalling pathways. Cell Microbiol. 2008, 10, 1775–1786. [Google Scholar] [CrossRef] [Green Version]
- Morinaga, N.; Yahiro, K.; Matsuura, G.; Moss, J.; Noda, M. Subtilase cytotoxin, produced by Shiga-toxigenic Escherichia coli, transiently inhibits protein synthesis of Vero cells via degradation of BiP and induces cell cycle arrest at G1 by downregulation of cyclin D1. Cell Microbiol. 2008, 10, 921–929. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yahiro, K.; Morinaga, N.; Moss, J.; Noda, M. Subtilase cytotoxin induces apoptosis in HeLa cells by mitochondrial permeabilization via activation of Bax/Bak, independent of C/EBF-homologue protein (CHOP), Ire1alpha or JNK signaling. Microb. Pathog. 2010, 49, 153–163. [Google Scholar] [CrossRef] [Green Version]
- Tsutsuki, H.; Yahiro, K.; Suzuki, K.; Suto, A.; Ogura, K.; Nagasawa, S.; Ihara, H.; Shimizu, T.; Nakajima, H.; Moss, J.; et al. Subtilase cytotoxin enhances Escherichia coli survival in macrophages by suppression of nitric oxide production through the inhibition of NF-kappa B activation. Infect. Immun. 2012, 80, 3939–3951. [Google Scholar] [CrossRef] [Green Version]
- Tsutsuki, H.; Yahiro, K.; Ogura, K.; Ichimura, K.; Iyoda, S.; Ohnishi, M.; Nagasawa, S.; Seto, K.; Moss, J.; Noda, M. Subtilase cytotoxin produced by locus of enterocyte effacement-negative Shiga-toxigenic Escherichia coli induces stress granule formation. Cell Microbiol. 2016, 18, 1024–1040. [Google Scholar] [CrossRef]
- Yahiro, K.; Tsutsuki, H.; Ogura, K.; Nagasawa, S.; Moss, J.; Noda, M. DAP1, a negative regulator of autophagy, controls SubAB-mediated apoptosis and autophagy. Infect. Immun. 2014, 82, 4899–4908. [Google Scholar] [CrossRef] [Green Version]
- Galli, L.; Miliwebsky, E.; Irino, K.; Leotta, G.; Rivas, M. Virulence profile comparison between LEE-negative Shiga toxin-producing Escherichia coli (STEC) strains isolated from cattle and humans. Vet. Microbiol. 2010, 143, 307–313. [Google Scholar] [CrossRef] [Green Version]
- Amaral, M.M.; Sacerdoti, F.; Jancic, C.; Repetto, H.A.; Paton, A.W.; Paton, J.C.; Ibarra, C. Action of Shiga toxin type-2 and Subtilase cytotoxin on human microvascular endothelial cells. PLoS ONE 2013, 8, e70431. [Google Scholar] [CrossRef] [Green Version]
- Marquez, L.B.; Velázquez, N.; Repetto, H.A.; Paton, A.W.; Paton, J.C.; Ibarra, C.; Silberstein, C. Effects of Escherichia coli subtilase cytotoxin and Shiga toxin 2 on primary cultures of human renal tubular epithelial cells. PLoS ONE 2014, 9, e87022. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Alvarez, R.S.; Sacerdoti, F.; Jancic, C.; Müthing, J. Comparative characterization of shiga toxin type 2 and subtilase cytotoxin effects on human renal epithelial and endotelial cells grown in monolayer and bilayer conditions. PLoS ONE 2016, 11, e0158180. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Álvarez, R.S.; Jancic, C.; Garimano, N.; Sacerdoti, F.; Paton, A.W.; Paton, J.C.; Ibarra, C.; Amaral, M.M. Crosstalk between Human Microvascular Endothelial Cells and Tubular Epithelial Cells Modulates ProInflammatory Responses Induced by Shiga Toxin Type 2 and Subtilase Cytotoxin. Toxins 2019, 11, 648. [Google Scholar] [CrossRef] [Green Version]
- Wang, H.; Paton, J.C.; Paton, A.W. Pathologic changes in mice induced by subtilase cytotoxin, a potent new Escherichia coli AB5 toxin that targets the endoplasmic reticulum. J. Infect. Dis. 2007, 196, 1093–1101. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Seyahian, A.E.; Oltra, G.; Ochoa, F.; Melendi, S.; Hermes, R.; Paton, J.C.; Paton, A.W.; Lago, N.; Parodi, M.C.; Damiano, A.; et al. Systemic effects of Subtilase cytotoxin produced by Escherichia coli O113:H21. Toxicon 2017, 127, 49–55. [Google Scholar] [CrossRef] [PubMed]
- Velandia, C.V.; Sanso, A.M.; Krügeret, A.; Suárez, L.V.; Lucchesi, P.M.A.; Parma, A.E. Occurrence of subtilase cytotoxin and relation with other virulence factors in verocytotoxigenic Escherichia coli isolated from food and cattle in Argentina. Braz. J. Microbiol. 2011, 42, 711–715. [Google Scholar] [CrossRef] [Green Version]
- Tozzoli, R.; Caprioli, A.; Cappannella, S.; Michelacci, V.; Marziano, M.L.; Morabito, S. Production of the Subtilase AB5 cytotoxin by Shiga toxin-negative Escherichia coli. J. Clin. Microbiol. 2010, 48, 178–183. [Google Scholar] [CrossRef] [Green Version]
- Irino, K.; Midolli Vieira, M.A.; Tardelli Gomes, T.A.; Guth, B.E.C.; Naves, Z.V.F.; Oliveira, M.G.; dos Santos, L.F.; Guirro, M.; Timm, C.D.; Pigatto, C.P.; et al. Subtilase Cytotoxin-Encoding subAB Operon Found Exclusively among Shiga Toxin-Producing Escherichia coli Strains. J. Clin. Microbiol. 2010, 48, 988–990. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, H.; Rogers, T.J.; Paton, J.C.; Paton, A.W. Differential effects of Escherichia coli subtilase cytotoxin and Shiga toxin 2 on chemokine and proinflammatory cytokine expression in human macrophage, colonic epithelial, and brain microvascular endothelial cell lines. Infect. Immun. 2014, 82, 3567–3579. [Google Scholar] [CrossRef] [Green Version]
- Tarr, P.I.; Gordon, C.A.; Chandler, W.L. Shiga-toxin-producing Escherichia coli and haemolytic uraemic syndrome. Lancet 2005, 365, 1073–1086. [Google Scholar] [CrossRef]
- Rivas, M.; Miliwebsky, E.; Chinen, I.; Roldán, C.D.; Balbi, L.; Garcia, B.; Fiorilli, G.; Sosa-Estani, S.; Kincaid, J.; Rangel, J.; et al. Characterization and Epidemiologic Subtyping of Shiga Toxin–Producing Escherichia coli Strains Isolated from Hemolytic Uremic Syndrome and Diarrhea Cases in Argentina. Foodborne Pathog. 2006, 3, 88–96. [Google Scholar] [CrossRef] [Green Version]
- Girard, M.C.; Sacerdoti, F.; Rivera, F.P.; Repetto, H.A.; Ibarra, C.; Amaral, M.M. Prevention of renal damage caused by Shiga toxin type 2: Action of Miglustat on human endothelial and epithelial cells. Toxicon 2015, 105, 27–33. [Google Scholar] [CrossRef] [PubMed]
- Pellizzari, A.; Pang, H.; Lingwoodl, C.A. Binding of verocytotoxin 1 to its receptor is influenced by differences in receptor fatty acid content. Biochemistry 1992, 31, 1363–1370. [Google Scholar] [CrossRef] [PubMed]
- Sandvig, K.; Ryd, M.; Garred, O.; Schweda, E.; Holm, P.K.; van Deurs, B. Retrograde transport from the Golgi complex to the ER of both Shiga toxin and the nontoxic Shiga B-fragment is regulated by butyric acid and cAMP. J. Cell Biol. 1994, 126, 53–64. [Google Scholar] [CrossRef]
- Yahiro, K.; Satoh, M.; Morinaga, N.; Tsutsuki, H.; Ogura, K.; Nagasawa, S.; Nomura, F.; Moss, J.; Noda, M. Identification of subtilase cytotoxin (SubAB) receptors whose signaling, in association with SubAB-induced BiP cleavage, is responsible for apoptosis in HeLa cells. Infect. Immun. 2011, 79, 617–627. [Google Scholar] [CrossRef] [Green Version]
- Wang, H.; Paton, J.C.; Thorpe, C.M.; Bonder, C.S.; Sun, W.Y.; Paton, A.W. Tissue factor-dependent procoagulant activity of subtilase cytotoxin, a potent AB5 toxin produced by shiga toxigenic Escherichia coli. J. Infect. Dis. 2010, 202, 1415–1423. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Keepers, T.R.; Psotka, M.A.; Grosse, L.K.; Obrig, T.G. A murine model of HUS: Shiga toxin with lipopolysaccharide mimics the renal damage and physiologic response of human disease. J. Am. Soc. Nephrol. 2006, 17, 3404–3414. [Google Scholar] [CrossRef] [Green Version]
- Rasooly, R.; Do, P.M.; Griffey, S.M.; Vilches-Moure, J.G.; Friedman, M. Ingested Shiga Toxin 2 (Stx2) Causes Histopathological Changes in Kidney, Spleen, and Thymus Tissues and Mortality in Mice. J. Agric. Food Chem. 2010, 58, 9281–9286. [Google Scholar] [CrossRef] [PubMed]
- Kaplan, B.S. Shiga toxin-induced tubular injury in hemolytic uremic syndrome. Kidney Int. 1998, 54, 648–649. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Exeni, R.; Grimold, I.; Amore, A.; Antonuccio, M. Función tubular en niños con SUH. Arch. Argent. Pediatr. 1977, 75, 149–153. [Google Scholar]
- Richardson, S.E.; Karmali, M.A.; Becker, L.E.; Smith, C.R. The histopathology of the hemolytic uremic syndrome associated with verocytotoxin-producing Escherichia coli infections. Hum. Pathol. 1988, 19, 1102–1108. [Google Scholar] [CrossRef]
- Mohawk, K.L.; O’Brien, A.D. Mouse Models of Escherichia coli O157:H7 Infection and Shiga Toxin Injection. J. Biomed. Biotechnol. 2011, 2011, 258185. [Google Scholar] [CrossRef] [Green Version]
- González, A.M. Experto en Citología de Sangre Periférica, Líquidos Biológicos y Métodos de Análisis de Imágenes. Módulo 2: Citología de Sangre Periférica. Parte II: Diagnóstico Diferencial de las Anemias; Editorial Médica Panamericana: Madrid, Spain, 2021; pp. 1–20. [Google Scholar]
- Repetto, H.A. Clinical management of Escherichia coli cases (The latin America Experience). Pathog. Escherichia coli Lat. Am. 2010, 116, 116–121. [Google Scholar]



Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Álvarez, R.S.; Gómez, F.D.; Zotta, E.; Paton, A.W.; Paton, J.C.; Ibarra, C.; Sacerdoti, F.; Amaral, M.M. Combined Action of Shiga Toxin Type 2 and Subtilase Cytotoxin in the Pathogenesis of Hemolytic Uremic Syndrome. Toxins 2021, 13, 536. https://doi.org/10.3390/toxins13080536
Álvarez RS, Gómez FD, Zotta E, Paton AW, Paton JC, Ibarra C, Sacerdoti F, Amaral MM. Combined Action of Shiga Toxin Type 2 and Subtilase Cytotoxin in the Pathogenesis of Hemolytic Uremic Syndrome. Toxins. 2021; 13(8):536. https://doi.org/10.3390/toxins13080536
Chicago/Turabian StyleÁlvarez, Romina S., Fernando D. Gómez, Elsa Zotta, Adrienne W. Paton, James C. Paton, Cristina Ibarra, Flavia Sacerdoti, and María M. Amaral. 2021. "Combined Action of Shiga Toxin Type 2 and Subtilase Cytotoxin in the Pathogenesis of Hemolytic Uremic Syndrome" Toxins 13, no. 8: 536. https://doi.org/10.3390/toxins13080536
APA StyleÁlvarez, R. S., Gómez, F. D., Zotta, E., Paton, A. W., Paton, J. C., Ibarra, C., Sacerdoti, F., & Amaral, M. M. (2021). Combined Action of Shiga Toxin Type 2 and Subtilase Cytotoxin in the Pathogenesis of Hemolytic Uremic Syndrome. Toxins, 13(8), 536. https://doi.org/10.3390/toxins13080536