Perspectives on the Pseudomonas aeruginosa Type III Secretion System Effector ExoU and Its Subversion of the Host Innate Immune Response to Infection


{kind=link}
{kind=link}
Abstract
1. Introduction
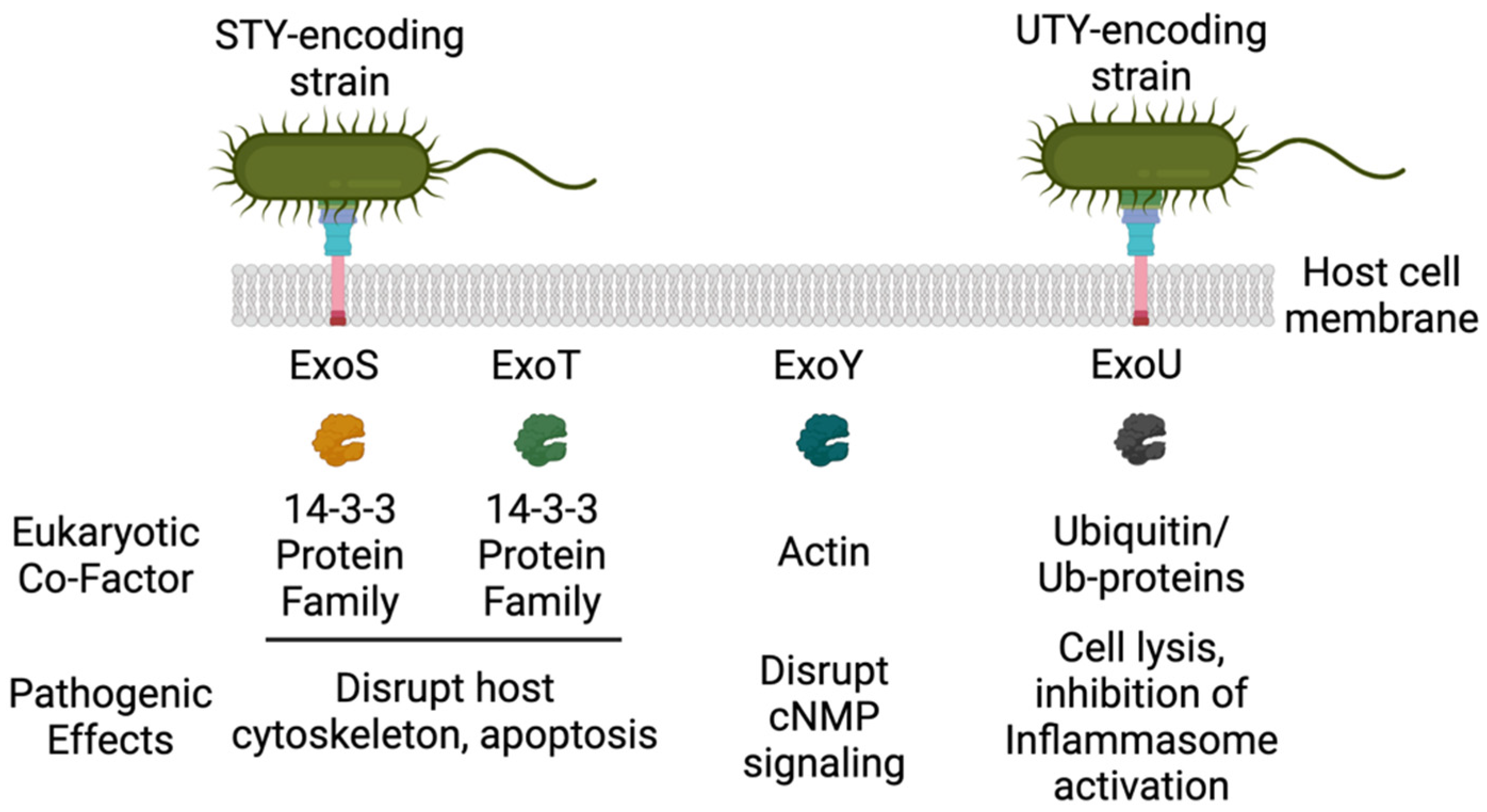
2. ExoU
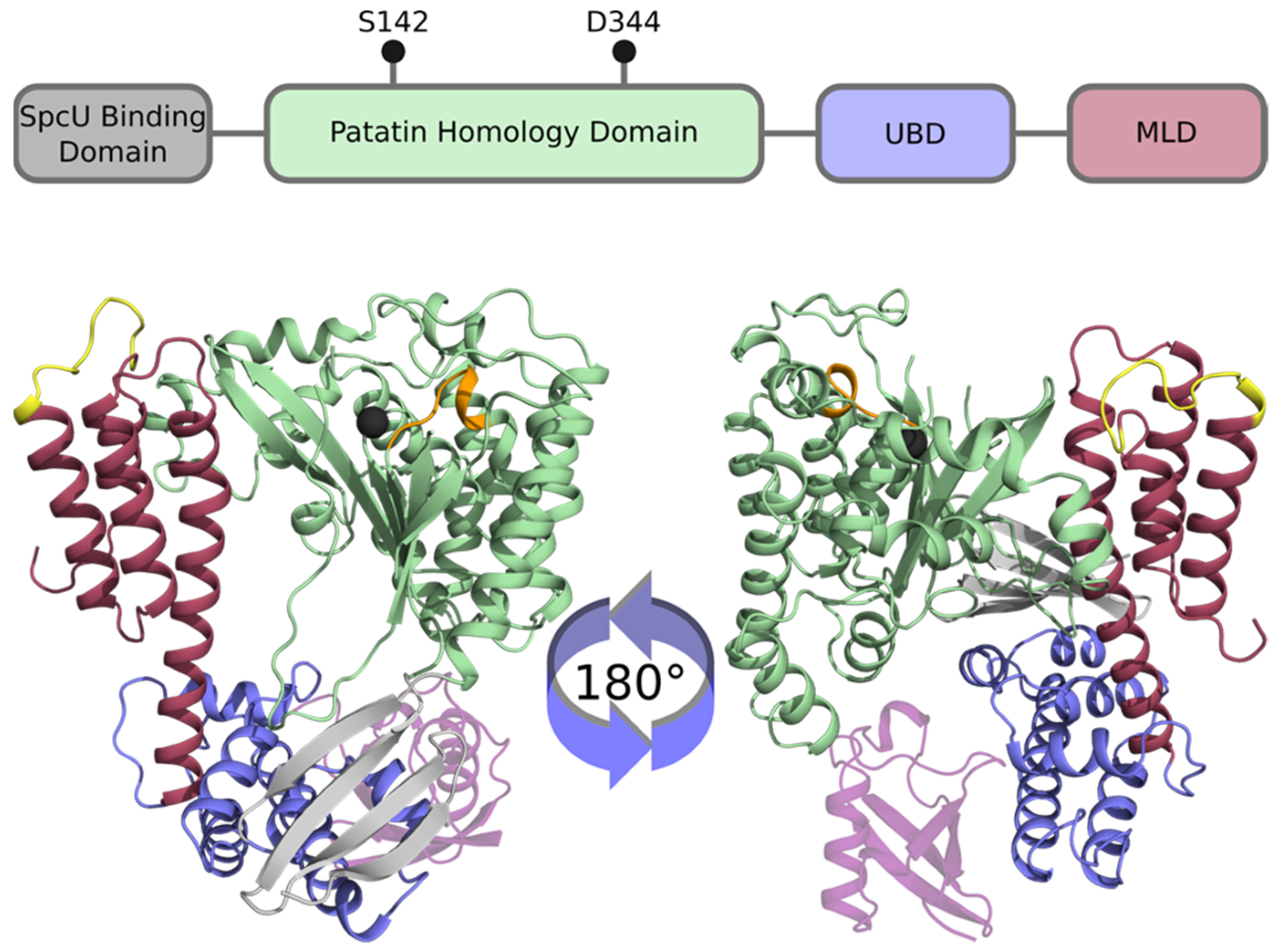
2.1. Structure-Activity Relationships of ExoU
2.2. Crystal Structure of ExoU
2.3. ExoU PLA2 Activity and Regulation by Eukaryotic Co-factors
2.4. ExoU Orthologues
2.5. ExoU PLA Activity and Pathogenesis
2.6. ExoU Subverts the Host Innate Immune Response to Infection
2.7. ExoU Transiently Represses NLRC4 Inflammasome Activation
3. Perspectives
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Feldman, M.; Bryan, R.; Rajan, S.; Scheffler, L.; Brunnert, S.; Tang, H.; Prince, A. Role of flagella in pathogenesis of Pseudomonas aeruginosa pulmonary infection. Infect. Immun. 1998, 66, 43–51. [Google Scholar] [CrossRef]
- Lister, P.D.; Wolter, D.J.; Hanson, N.D. Antibacterial-resistant Pseudomonas aeruginosa: Clinical impact and complex regulation of chromosomally encoded resistance mechanisms. Clin. Microbiol. Rev. 2009, 22, 582–610. [Google Scholar] [CrossRef]
- Global Priority List of Antibiotic-Resistant Bacteria to Guide Research, Discovery, and Development of New Antibiotics. Available online: https://www.who.int/medicines/publications/WHO-PPL-Short_Summary_25Feb-ET_NM_WHO.pdf?ua=1 (accessed on 27 February 2017).
- Pendleton, J.N.; Gorman, S.P.; Gilmore, B.F. Clinical relevance of the ESKAPE pathogens. Expert Rev. Anti-Infect. Ther. 2013, 11, 297–308. [Google Scholar] [CrossRef]
- Boucher, H.W.; Talbot, G.H.; Bradley, J.S.; Edwards, J.E.; Gilbert, D.; Rice, L.B.; Scheld, M.; Spellberg, B.; Bartlett, J. Bad bugs, no drugs: No ESKAPE! An update from the Infectious Diseases Society of America. Clin. Infect. Dis. 2009, 48, 1–12. [Google Scholar] [CrossRef]
- Brewer, S.C.; Wunderink, R.G.; Jones, C.B.; Leeper, K.V., Jr. Ventilator-associated pneumonia due to Pseudomonas aeruginosa. Chest 1996, 109, 1019–1029. [Google Scholar] [CrossRef] [PubMed]
- ElSolh, A.A.; Akinnusi, M.E.; Wiener-Kronish, J.P.; Lynch, S.V.; Pineda, L.A.; Szarpa, K. Persistent infection with Pseudomonas aeruginosa in ventilator-associated pneumonia. Am. J. Respir. Crit. Care Med. 2008, 178, 513–519. [Google Scholar] [CrossRef] [PubMed]
- El-Solh, A.A.; Hattemer, A.; Hauser, A.R.; Alhajhusain, A.; Vora, H. Clinical outcomes of type III Pseudomonas aeruginosa bacteremia. Crit. Care Med. 2012, 40, 1157–1163. [Google Scholar] [CrossRef]
- Hauser, A.R.; Cobb, E.; Bodi, M.; Mariscal, D.; Valles, J.; Engel, J.N.; Rello, J. Type III protein secretion is associated with poor clinical outcomes in patients with ventilator-associated pneumonia caused by Pseudomonas aeruginosa. Crit. Care Med. 2002, 30, 521–528. [Google Scholar] [CrossRef] [PubMed]
- Juan, C.; Pena, C.; Oliver, A. Host and pathogen biomarkers for severe Pseudomonas aeruginosa infections. J. Infect. Dis. 2017, 215 (Suppl. S1), S44–S51. [Google Scholar] [CrossRef]
- Sadikot, R.T.; Blackwell, T.S.; Christman, J.W.; Prince, A.S. Pathogen–Host interactions in Pseudomonas aeruginosa pneumonia. Am. J. Respir. Crit. Care Med. 2005, 171, 1209–1223. [Google Scholar] [CrossRef] [PubMed]
- Williams, B.J.; Dehnbostel, J.; Blackwell, T.S. Pseudomonas aeruginosa: Host defence in lung diseases. Respirology 2010, 15, 1037–1056. [Google Scholar] [CrossRef]
- Gellatly, S.L.; Hancock, R.E. Pseudomonas aeruginosa: New insights into pathogenesis and host defenses. Pathog. Dis. 2013, 67, 159–173. [Google Scholar] [CrossRef] [PubMed]
- Roy-Burman, A.; Savel, R.H.; Racine, S.; Swanson, B.L.; Revadigar, N.S.; Fujimoto, J.; Sawa, T.; Frank, D.W.; Wiener-Kronish, J.P. Type III protein secretion is associated with death in lower respiratory and systemic Pseudomonas aeruginosa infections. J. Infect. Dis. 2001, 183, 1767–1774. [Google Scholar] [CrossRef] [PubMed]
- Tsay, T.B.; Jiang, Y.Z.; Hsu, C.M.; Chen, L.W. Pseudomonas aeruginosa colonization enhances ventilator-associated pneumonia-induced lung injury. Respir. Res. 2016, 17, 101. [Google Scholar] [CrossRef]
- Kudoh, I.; Wiener-Kronish, J.P.; Hashimoto, S.; Pittet, J.F.; Frank, D. Exoproduct secretions of Pseudomonas aeruginosa strains influence severity of alveolar epithelial injury. Am. J. Physiol. 1994, 267, L551–L556. [Google Scholar] [CrossRef] [PubMed]
- Wiener-Kronish, J.P.; Sakuma, T.; Kudoh, I.; Pittet, J.F.; Frank, D.; Dobbs, L.; Vasil, M.L.; Matthay, M.A. Alveolar epithelial injury and pleural empyema in acute P. aeruginosa pneumonia in anesthetized rabbits. J. Appl. Physiol. 1993, 75, 1661–1669. [Google Scholar] [CrossRef] [PubMed]
- Ader, F.; LeBerre, R.; Faure, K.; Gosset, P.; Epaulard, O.; Toussaint, B.; Polack, B.; Nowak, E.; Viget, N.B.; Kipnis, E.; et al. Alveolar response to Pseudomonas aeruginosa: Role of the type III secretion system. Infect. Immun. 2005, 73, 4263–4271. [Google Scholar] [CrossRef] [PubMed]
- Dacheux, D.; Attree, I.; Schneider, C.; Toussaint, B. Cell death of human polymorphonuclear neutrophils induced by a Pseudomonas aeruginosa cystic fibrosis isolate requires a functional type III secretion system. Infect. Immun. 1999, 67, 6164–6167. [Google Scholar] [CrossRef]
- Morin, C.D.; Deziel, E.; Gauthier, J.; Levesque, R.C.; Lau, G.W. An organ system-based synopsis of Pseudomonas aeruginosa virulence. Virulence 2021, 12, 1469–1507. [Google Scholar] [CrossRef] [PubMed]
- Reynolds, D.; Kollef, M. The Epidemiology and Pathogenesis and Treatment of Pseudomonas aeruginosa Infections: An Update. Drugs 2021, 81, 2117–2131. [Google Scholar] [CrossRef]
- Sawa, T. The molecular mechanism of acute lung injury caused by Pseudomonas aeruginosa: From bacterial pathogenesis to host response. J. Intensive Care 2014, 2, 10. [Google Scholar] [CrossRef]
- Schulert, G.S.; Feltman, H.; Rabin, S.D.P.; Martin, C.G.; Battle, S.E.; Rello, J.; Hauser, A.R. Secretion of the Toxin ExoU Is a Marker for Highly Virulent Pseudomonas aeruginosa Isolates Obtained from Patients with Hospital-Acquired Pneumonia. J. Infect. Dis. 2003, 188, 1695–1706. [Google Scholar] [CrossRef]
- Fleiszig, S.M.; Wiener-Kronish, J.P.; Miyazaki, H.; Vallas, V.; Mostov, K.E.; Kanada, D.; Sawa, T.; Yen, T.S.; Frank, D.W. Pseudomonas aeruginosa-mediated cytotoxicity and invasion correlate with distinct genotypes at the loci encoding exoenzyme S. Infect Immun. 1997, 65, 579–586. [Google Scholar] [CrossRef] [PubMed]
- Feltman, H.; Schulert, G.; Khan, S.; Jain, M.; Peterson, L.; Hauser, A.R. Prevalence of type III secretion genes in clinical and environmental isolates of Pseudomonas aeruginosa. Microbiology 2001, 147, 2659–2669. [Google Scholar] [CrossRef] [PubMed]
- Barbieri, J.T.; Sun, J. Pseudomonas aeruginosa ExoS and ExoT. Rev. Physiol. Biochem. Pharmacol. 2004, 152, 79–92. [Google Scholar] [CrossRef]
- Hauser, A.R. The type III secretion system of Pseudomonas aeruginosa: Infection by injection. Nat. Rev. Genet. 2009, 7, 654–665. [Google Scholar] [CrossRef]
- Beckert, U.; Wolter, S.; Hartwig, C.; Bähre, H.; Kaever, V.; Ladant, D.; Frank, D.W.; Seifert, R. ExoY from Pseudomonas aeruginosa is a nucleotidyl cyclase with preference for cGMP and cUMP formation. Biochem. Biophys. Res. Commun. 2014, 450, 870–874. [Google Scholar] [CrossRef]
- Belyy, A.; Raoux-Barbot, D.; Saveanu, C.; Namane, A.; Ogryzko, V.; Worpenberg, L.; David, V.; Henriot, V.; Fellous, S.; Merrifield, C.; et al. Actin activates Pseudomonas aeruginosa ExoY nucleotidyl cyclase toxin and ExoY-like effector domains from MARTX toxins. Nat. Commun. 2016, 7, 13582. [Google Scholar] [CrossRef]
- Morrow, K.A.; Frank, D.W.; Balczon, R.; Stevens, T. The Pseudomonas aeruginosa Exoenzyme Y: A Promiscuous Nucleotidyl Cyclase Edema Factor and Virulence Determinant. Non-Canonical Cycl. Nucleotides 2016, 238, 67–85. [Google Scholar] [CrossRef]
- Morrow, K.A.; Ochoa, C.D.; Balczon, R.; Zhou, C.; Cauthen, L.; Alexeyev, M.; Schmalzer, K.M.; Frank, D.W.; Stevens, T. Pseudomonas aeruginosa exoenzymes U and Y induce a transmissible endothelial proteinopathy. Am. J. Physiol. Cell. Mol. Physiol. 2016, 310, L337–L353. [Google Scholar] [CrossRef] [PubMed]
- Sayner, S.L.; Frank, D.W.; King, J.; Chen, H.; VandeWaa, J.; Stevens, T. Paradoxical cAMP-induced lung endothelial hyperpermeability revealed by Pseudomonas aeruginosa ExoY. Circ. Res. 2004, 95, 196–203. [Google Scholar] [CrossRef] [PubMed]
- Voth, S.; Gwin, M.; Francis, C.M.; Balczon, R.; Frank, D.W.; Pittet, J.; Wagener, B.M.; Moser, S.A.; Alexeyev, M.; Housley, N.; et al. Virulent Pseudomonas aeruginosa infection converts antimicrobial amyloids into cytotoxic prions. FASEB J. 2020, 34, 9156–9179. [Google Scholar] [CrossRef] [PubMed]
- Finck-Barbançon, V.; Goranson, J.; Zhu, L.; Sawa, T.; Wiener-Kronish, J.P.; Fleiszig, S.M.J.; Wu, C.; Mende-Mueller, L.; Frank, D.W. ExoU expression by Pseudomonas aeruginosa correlates with acute cytotoxicity and epithelial injury. Mol. Microbiol. 1997, 25, 547–557. [Google Scholar] [CrossRef] [PubMed]
- Fischer, S.; Dethlefsen, S.; Klockgether, J.; Tummler, B. Phenotypic and Genomic Comparison of the Two Most Common ExoU-Positive Pseudomonas aeruginosa Clones, PA14 and ST235. Msystems 2020, 5, e01007-20. [Google Scholar] [CrossRef]
- Hauser, A.R.; Kang, P.J.; Engel, J.N. PepA, a secreted protein of Pseudomonas aeruginosa, is necessary for cytotoxicity and virulence. Mol. Microbiol. 1998, 27, 807–818. [Google Scholar] [CrossRef] [PubMed]
- Kulasekara, B.R.; Kulasekara, H.D.; Wolfgang, M.C.; Stevens, L.; Frank, D.W.; Lory, S. Acquisition and Evolution of the exoU Locus in Pseudomonas aeruginosa. J. Bacteriol. 2006, 188, 4037–4050. [Google Scholar] [CrossRef] [PubMed]
- Sato, H.; Frank, D.W.; Hillard, C.J.; Feix, J.B.; Pankhaniya, R.R.; Moriyama, K.; Finck-Barbançon, V.; Buchaklian, A.; Lei, M.; Long, R.M.; et al. The mechanism of action of the Pseudomonas aeruginosa-encoded type III cytotoxin, ExoU. EMBO J. 2003, 22, 2959–2969. [Google Scholar] [CrossRef] [PubMed]
- Phillips, R.M.; Six, D.; Dennis, E.A.; Ghosh, P.; Matsumoto, K.; Shionyu, M.; Go, M.; Shimizu, K.; Shinomura, T.; Kimata, K.; et al. In Vivo Phospholipase Activity of the Pseudomonas aeruginosa Cytotoxin ExoU and Protection of Mammalian Cells with Phospholipase A2 Inhibitors. J. Biol. Chem. 2003, 278, 41326–41332. [Google Scholar] [CrossRef]
- Sato, H.; Frank, D.W. ExoU is a potent intracellular phospholipase. Mol. Microbiol. 2004, 53, 1279–1290. [Google Scholar] [CrossRef] [PubMed]
- Finck-Barbançon, V.; Frank, D.W. Multiple Domains Are Required for the Toxic Activity of Pseudomonas aeruginosa ExoU. J. Bacteriol. 2001, 183, 4330–4344. [Google Scholar] [CrossRef]
- Hirschberg, H.J.H.B.; Simons, J.-W.F.A.; Dekker, N.; Egmond, M.R. Cloning, expression, purification and characterization of patatin, a novel phospholipase A. JBIC J. Biol. Inorg. Chem. 2001, 268, 5037–5044. [Google Scholar] [CrossRef]
- Dessen, A.; Tang, J.; Schmidt, H.; Stahl, M.; Clark, J.D.; Seehra, J.; Somers, W.S. Crystal Structure of Human Cytosolic Phospholipase A2 Reveals a Novel Topology and Catalytic Mechanism. Cell 1999, 97, 349–360. [Google Scholar] [CrossRef]
- Rabin, S.D.P.; Hauser, A.R. Functional Regions of the Pseudomonas aeruginosa Cytotoxin ExoU. Infect. Immun. 2005, 73, 573–582. [Google Scholar] [CrossRef] [PubMed]
- Finck-Barbançon, V.; Yahr, T.L.; Frank, D.W. Identification and Characterization of SpcU, a Chaperone Required for Efficient Secretion of the ExoU Cytotoxin. J. Bacteriol. 1998, 180, 6224–6231. [Google Scholar] [CrossRef] [PubMed]
- Rabin, S.D.P.; Veesenmeyer, J.L.; Bieging, K.T.; Hauser, A.R. A C-Terminal Domain Targets the Pseudomonas aeruginosa Cytotoxin ExoU to the Plasma Membrane of Host Cells. Infect. Immun. 2006, 74, 2552–2561. [Google Scholar] [CrossRef]
- Schmalzer, K.M.; Benson, M.A.; Frank, D.W. Activation of ExoU Phospholipase Activity Requires Specific C-Terminal Regions. J. Bacteriol. 2010, 192, 1801–1812. [Google Scholar] [CrossRef] [PubMed]
- Veesenmeyer, J.L.; Howell, H.; Halavaty, A.S.; Ahrens, S.; Anderson, W.F.; Hauser, A.R. Role of the Membrane Localization Domain of the Pseudomonas aeruginosa Effector Protein ExoU in Cytotoxicity. Infect. Immun. 2010, 78, 3346–3357. [Google Scholar] [CrossRef]
- Stirling, F.R.; Cuzick, A.; Kelly, S.M.; Oxley, D.; Evans, T.J. Eukaryotic localization, activation and ubiquitinylation of a bacterial type III secreted toxin. Cell. Microbiol. 2006, 8, 1294–1309. [Google Scholar] [CrossRef]
- Benson, M.A.; Komas, S.M.; Schmalzer, K.M.; Casey, M.S.; Frank, D.W.; Feix, J.B. Induced Conformational Changes in the Activation of the Pseudomonas aeruginosa type III Toxin, ExoU. Biophys. J. 2011, 100, 1335–1343. [Google Scholar] [CrossRef]
- Gendrin, C.; Contreras-Martel, C.; Bouillot, S.; Elsen, S.; Lemaire, D.; Skoufias, D.; Huber, P.; Attree, I.; Dessen, A. Structural Basis of Cytotoxicity Mediated by the Type III Secretion Toxin ExoU from Pseudomonas aeruginosa. PLoS Pathog. 2012, 8, e1002637. [Google Scholar] [CrossRef]
- Halavaty, A.S.; Borek, D.; Tyson, G.; Veesenmeyer, J.L.; Shuvalova, L.; Minasov, G.; Otwinowski, Z.; Hauser, A.R.; Anderson, W.F. Structure of the Type III Secretion Effector Protein ExoU in Complex with Its Chaperone SpcU. PLoS ONE 2012, 7, e49388. [Google Scholar] [CrossRef] [PubMed]
- Akeda, Y.; Galán, J.E. Chaperone release and unfolding of substrates in type III secretion. Nat. Cell Biol. 2005, 437, 911–915. [Google Scholar] [CrossRef]
- Tessmer, M.H.; DeCero, S.A.; Del Alamo, D.; Riegert, M.O.; Meiler, J.; Frank, D.W.; Feix, J.B. Characterization of the ExoU activation mechanism using EPR and integrative modeling. Sci. Rep. 2020, 10, 19700. [Google Scholar] [CrossRef]
- Sato, H.; Feix, J.B.; Frank, D.W. Identification of Superoxide Dismutase as a Cofactor for thePseudomonasType III Toxin, ExoU. Biochem. 2006, 45, 10368–10375. [Google Scholar] [CrossRef]
- Benson, M.A.; Schmalzer, K.M.; Frank, D.W. A sensitive fluorescence-based assay for the detection of ExoU-mediated PLA2 activity. Clin. Chim. Acta 2010, 411, 190–197. [Google Scholar] [CrossRef][Green Version]
- Anderson, D.M.; Schmalzer, K.M.; Sato, H.; Casey, M.; Terhune, S.S.; Haas, A.L.; Feix, J.B.; Frank, D.W. Ubiquitin and ubiquitin-modified proteins activate the Pseudomonas aeruginosa T3SS cytotoxin, ExoU. Mol. Microbiol. 2011, 82, 1454–1467. [Google Scholar] [CrossRef]
- Rechsteiner, M. Ubiquitin-mediated pathways for intracellular proteolysis. Annul. Rev. Cell Biol. 1987, 3, 1–30. [Google Scholar] [CrossRef]
- Anderson, D.M.; Feix, J.B.; Monroe, A.L.; Peterson, F.C.; Volkman, B.F.; Haas, A.L.; Frank, D.W. Identification of the Major Ubiquitin-binding Domain of the Pseudomonas aeruginosa ExoU A2 Phospholipase. J. Biol. Chem. 2013, 288, 26741–26752. [Google Scholar] [CrossRef] [PubMed]
- Tessmer, M.H.; Anderson, D.M.; Buchaklian, A.; Frank, D.W.; Feix, J.B. Cooperative Substrate-Cofactor Interactions and Membrane Localization of the Bacterial Phospholipase A2 (PLA2) Enzyme, ExoU. J. Biol. Chem. 2017, 292, 3411–3419. [Google Scholar] [CrossRef] [PubMed]
- Tessmer, M.H.; Anderson, D.M.; Pickrum, A.M.; Riegert, M.O.; Moretti, R.; Meiler, J.; Feix, J.B.; Frank, D.W. Identification of a ubiquitin-binding interface using Rosetta and DEER. Proc. Natl. Acad. Sci. USA 2018, 115, 525–530. [Google Scholar] [CrossRef]
- Tyson, G.; Halavaty, A.S.; Kim, H.; Geissler, B.; Agard, M.; Satchell, K.J.; Cho, W.; Anderson, W.F.; Hauser, A.R. A Novel Phosphatidylinositol 4,5-Bisphosphate Binding Domain Mediates Plasma Membrane Localization of ExoU and Other Patatin-like Phospholipases. J. Biol. Chem. 2015, 290, 2919–2937. [Google Scholar] [CrossRef]
- Tyson, G.H.; Hauser, A.R. Phosphatidylinositol 4,5-Bisphosphate Is a Novel Coactivator of the Pseudomonas aeruginosa Cytotoxin ExoU. Infect. Immun. 2013, 81, 2873–2881. [Google Scholar] [CrossRef] [PubMed]
- Zhang, A.; Veesenmeyer, J.L.; Hauser, A.R. Phosphatidylinositol 4,5-Bisphosphate-Dependent Oligomerization of the Pseudomonas aeruginosa Cytotoxin ExoU. Infect. Immun. 2018, 86, e00402-17. [Google Scholar] [CrossRef] [PubMed]
- McLaughlin, S.; Wang, J.; Gambhir, A.; Murray, D. PIP2 and Proteins: Interactions, Organization, and Information Flow. Annu. Rev. Biophys. Biomol. Struct. 2002, 31, 151–175. [Google Scholar] [CrossRef] [PubMed]
- Anderson, D.M.; Sato, H.; Dirck, A.T.; Feix, J.B.; Frank, D.W. Ubiquitin Activates Patatin-Like Phospholipases from Multiple Bacterial Species. J. Bacteriol. 2015, 197, 529–541. [Google Scholar] [CrossRef]
- Sato, H.; Frank, D.W. Intoxication of Host Cells by the T3SS Phospholipase ExoU: PI(4,5)P2-Associated, Cytoskeletal Collapse and Late Phase Membrane Blebbing. PLoS ONE 2014, 9, e103127. [Google Scholar] [CrossRef] [PubMed]
- Lee, V.T.; Pukatzki, S.; Sato, H.; Kikawada, E.; Kazimirova, A.A.; Huang, J.; Li, X.; Arm, J.P.; Frank, D.W.; Lory, S. Pseudolipasin A Is a Specific Inhibitor for Phospholipase A 2 Activity of Pseudomonas aeruginosa Cytotoxin ExoU. Infect. Immun. 2007, 75, 1089–1098. [Google Scholar] [CrossRef]
- Kim, D.; Baek, J.; Song, J.; Byeon, H.; Min, H.; Min, K.H. Identification of arylsulfonamides as ExoU inhibitors. Bioorganic Med. Chem. Lett. 2014, 24, 3823–3825. [Google Scholar] [CrossRef] [PubMed]
- Foulkes, D.M.; McLean, K.; Haneef, A.; Fernig, D.G.; Winstanley, C.; Berry, N.; Kaye, S.B. Pseudomonas aeruginosa Toxin ExoU as a Therapeutic Target in the Treatment of Bacterial Infections. Microorganisms 2019, 7, 707. [Google Scholar] [CrossRef]
- Pickrum, A.M.; DeLeon, O.; Dirck, A.; Tessmer, M.H.; Riegert, M.O.; Biller, J.A.; Ledeboer, N.A.; Kirby, J.R.; Frank, D.W. Achromobacter xylosoxidans Cellular Pathology Is Correlated with Activation of a Type III Secretion System. Infect. Immun. 2020, 88. [Google Scholar] [CrossRef]
- Housley, N.A.; Winkler, H.H.; Audia, J.P. The Rickettsia prowazekii ExoU Homologue Possesses Phospholipase A 1 (PLA 1), PLA 2, and Lyso-PLA 2 Activities and Can Function in the Absence of Any Eukaryotic Cofactors In Vitro. J. Bacteriol. 2011, 193, 4634–4642. [Google Scholar] [CrossRef] [PubMed]
- Winkler, H.H.; Miller, E.T. Phospholipase A Activity in the Hemolysis of Sheep and Human Erythrocytes by Rickettsia prowazeki. Infect. Immun. 1980, 29, 316–321. [Google Scholar] [CrossRef]
- Winkler, H.H.; Miller, E.T. Phospholipase A and the interaction of Rickettsia prowazekii and mouse fibroblasts (L-929 cells). Infect. Immun. 1982, 38, 109–113. [Google Scholar] [CrossRef]
- Blanc, G.; Renesto, P.; Raoult, D. Phylogenic Analysis of Rickettsial Patatin-like Protein with Conserved Phospholipase A2 Active Sites. Ann. N. Y. Acad. Sci. 2005, 1063, 83–86. [Google Scholar] [CrossRef] [PubMed]
- Read, T.; Peterson, S.N.; Tourasse, N.; Baillie, L.; Paulsen, I.; Nelson, K.E.; Tettelin, H.; Fouts, D.E.; Eisen, J.A.; Gill, S.R.; et al. The genome sequence of Bacillus anthracis Ames and comparison to closely related bacteria. Nat. Cell Biol. 2003, 423, 81–86. [Google Scholar] [CrossRef] [PubMed]
- Dixon, T.C.; Meselson, M.; Guillemin, J.; Hanna, P.C. Anthrax. N. Engl. J. Med. 1999, 341, 815–826. [Google Scholar] [CrossRef]
- Shin, S.; Kim, Y.-B.; Hur, G.-H. Involvement of phospholipase A2 activation in anthrax lethal toxin-induced cytotoxicity. Cell Biol. Toxicol. 1999, 15, 19–29. [Google Scholar] [CrossRef]
- Howell, H.A.; Logan, L.K.; Hauser, A.R. Type III Secretion of ExoU Is Critical during Early Pseudomonas aeruginosa Pneumonia. MBio 2013, 4, e00032-13. [Google Scholar] [CrossRef]
- Shaver, C.M.; Hauser, A.R. Relative Contributions of Pseudomonas aeruginosa ExoU, ExoS, and ExoT to Virulence in the Lung. Infect. Immun. 2004, 72, 6969–6977. [Google Scholar] [CrossRef]
- Kurahashi, K.; Kajikawa, O.; Sawa, T.; Ohara, M.; Gropper, M.A.; Frank, D.W.; Martin, T.R.; Wiener-Kronish, J.P. Pathogenesis of septic shock in Pseudomonas aeruginosa pneumonia. J. Clin. Investig. 1999, 104, 743–750. [Google Scholar] [CrossRef] [PubMed]
- Diaz, M.H.; Shaver, C.M.; King, J.D.; Musunuri, S.; Kazzaz, J.A.; Hauser, A.R. Pseudomonas aeruginosa Induces Localized Immunosuppression during Pneumonia. Infect. Immun. 2008, 76, 4414–4421. [Google Scholar] [CrossRef] [PubMed]
- Diaz, M.H.; Hauser, A.R. Pseudomonas aeruginosa Cytotoxin ExoU Is Injected into Phagocytic Cells during Acute Pneumonia. Infect. Immun. 2010, 78, 1447–1456. [Google Scholar] [CrossRef] [PubMed]
- Allewelt, M.; Coleman, F.T.; Grout, M.; Priebe, G.P.; Pier, G.B. Acquisition of Expression of the Pseudomonas aeruginosa ExoU Cytotoxin Leads to Increased Bacterial Virulence in a Murine Model of Acute Pneumonia and Systemic Spread. Infect. Immun. 2000, 68, 3998–4004. [Google Scholar] [CrossRef] [PubMed]
- Alvarez, D.F.; Housley, N.; Koloteva, A.; Zhou, C.; O’Donnell, K.; Audia, J.P. Caspase-1 Activation Protects Lung Endothelial Barrier Function during Infection-Induced Stress. Am. J. Respir. Cell Mol. Biol. 2016, 55, 500–510. [Google Scholar] [CrossRef]
- Lindsey, A.S.; Sullivan, L.M.; Housley, N.A.; Koloteva, A.; King, J.A.; Audia, J.P.; Alvarez, D.F. Analysis of pulmonary vascular injury and repair during Pseudomonas aeruginosa infection-induced pneumonia and acute respiratory distress syndrome. Pulm. Circ. 2019, 9, 2045894019826941. [Google Scholar] [CrossRef]
- Thinakaran, G.; Koo, E.H. Amyloid Precursor Protein Trafficking, Processing, and Function. J. Biol. Chem. 2008, 283, 29615–29619. [Google Scholar] [CrossRef]
- Sevigny, J.; Chiao, P.; Bussiere, T.; Weinreb, P.H.; Williams, L.; Maier, M.; Dunstan, R.; Salloway, S.; Chen, T.; Ling, Y.; et al. The antibody aducanumab reduces Abeta plaques in Alzheimer’s disease. Nature 2016, 537, 50–56. [Google Scholar] [CrossRef]
- Inestrosa, N.C.; Soto, C. Molecular biology of the amyloid of Alzheimer’s disease. An overview. Biol. Res. 1992, 25, 63–72. [Google Scholar]
- Du, D.; Murray, A.N.; Cohen, E.; Kim, H.E.; Simkovsky, R.; Dillin, A.; Kelly, J.W. A kinetic aggregation assay allowing selective and sensitive amyloid-beta quantification in cells and tissues. Biochemistry 2011, 50, 1607–1617. [Google Scholar] [CrossRef]
- Parodi-Rullán, R.; Sone, J.Y.; Fossati, S. Endothelial Mitochondrial Dysfunction in Cerebral Amyloid Angiopathy and Alzheimer’s Disease. J. Alzheimer’s Dis. 2019, 72, 1019–1039. [Google Scholar] [CrossRef]
- Dempsey, C.; Rubio Araiz, A.; Bryson, K.J.; Finucane, O.; Larkin, C.; Mills, E.L.; Robertson, A.A.B.; Cooper, M.A.; O’Neill, L.A.J.; Lynch, M.A. Inhibiting the NLRP3 inflammasome with MCC950 promotes non-phlogistic clearance of amyloid-beta and cognitive function in APP/PS1 mice. Brain Behav. Immun. 2017, 61, 306–316. [Google Scholar] [CrossRef] [PubMed]
- Price, J.M.; Chi, X.; Hellermann, G.; Sutton, E.T. Physiological levels of beta-amyloid induce cerebral vessel dysfunction and reduce endothelial nitric oxide production. Neurol. Res. 2001, 23, 506–512. [Google Scholar] [CrossRef]
- Lin, M.T.; Balczon, R.; Pittet, J.-F.; Wagener, B.M.; Moser, S.A.; Morrow, K.A.; Voth, S.; Francis, C.M.; Leavesley, S.; Bell, J.; et al. Nosocomial Pneumonia Elicits an Endothelial Proteinopathy: Evidence for a Source of Neurotoxic Amyloids in Critically Ill Patients. Am. J. Respir. Crit. Care Med. 2018, 198, 1575–1578. [Google Scholar] [CrossRef] [PubMed]
- Balczon, R.; Morrow, K.A.; Zhou, C.; Edmonds, B.; Alexeyev, M.; Pittet, J.; Wagener, B.M.; Moser, S.A.; Leavesley, S.; Zha, X.; et al. Pseudomonas aeruginosa infection liberates transmissible, cytotoxic prion amyloids. FASEB J. 2017, 31, 2785–2796. [Google Scholar] [CrossRef]
- Stevens, T.C.; Ochoa, C.D.; Morrow, K.A.; Robson, M.J.; Prasain, N.; Zhou, C.; Alvarez, D.F.; Frank, D.W.; Balczon, R.; Stevens, T. The Pseudomonas aeruginosa exoenzyme Y impairs endothelial cell proliferation and vascular repair following lung injury. Am. J. Physiol. Cell. Mol. Physiol. 2014, 306, L915–L924. [Google Scholar] [CrossRef]
- Kumar, D.K.V.; Choi, S.H.; Washicosky, K.J.; Eimer, W.A.; Tucker, S.; Ghofrani, J.; Lefkowitz, A.; McColl, G.; Goldstein, L.E.; Tanzi, R.E.; et al. Amyloid-β peptide protects against microbial infection in mouse and worm models of Alzheimer’s disease. Sci. Transl. Med. 2016, 8, 340ra72. [Google Scholar] [CrossRef] [PubMed]
- Kumar, D.K.; Eimer, W.A.; Tanzi, R.E.; Moir, R.D. Alzheimer’s disease: The potential therapeutic role of the natural antibiotic amyloid-β peptide. Neurodegener. Dis. Manag. 2016, 6, 345–348. [Google Scholar] [CrossRef] [PubMed]
- Soscia, S.J.; Kirby, J.E.; Washicosky, K.J.; Tucker, S.M.; Ingelsson, M.; Hyman, B.; Burton, M.A.; Goldstein, L.E.; Duong, S.; Tanzi, R.E.; et al. The Alzheimer’s disease-associated amyloid β-protein is an antimicrobial peptide. PLoS ONE 2010, 5, e9505. [Google Scholar] [CrossRef] [PubMed]
- Martinon, F.; Burns, K.; Tschopp, J. The inflammasome: A molecular platform triggering activation of inflammatory caspases and processing of proIL-beta. Mol. Cell 2002, 10, 417–426. [Google Scholar] [CrossRef]
- Grandjean, T.; Boucher, A.; Thepaut, M.; Monlezun, L.; Guery, B.; Faudry, E.; Kipnis, E.; Dessein, R. The human NAIP-NLRC4-inflammasome senses the Pseudomonas aeruginosa T3SS inner-rod protein. Int. Immunol. 2017, 29, 377–384. [Google Scholar] [CrossRef]
- Kofoed, E.M.; Vance, R.E. Innate immune recognition of bacterial ligands by NAIPs determines inflammasome specificity. Nat. Cell Biol. 2011, 477, 592–595. [Google Scholar] [CrossRef]
- Rayamajhi, M.; Zak, D.E.; Chavarria-Smith, J.; Vance, R.E.; Miao, E.A. Cutting Edge: Mouse NAIP1 Detects the Type III Secretion System Needle Protein. J. Immunol. 2013, 191, 3986–3989. [Google Scholar] [CrossRef] [PubMed]
- Zhao, Y.; Yang, J.; Shi, J.; Gong, Y.-N.; Lu, Q.; Xu, H.; Liu, L.; Shao, F. The NLRC4 inflammasome receptors for bacterial flagellin and type III secretion apparatus. Nat. Cell Biol. 2011, 477, 596–600. [Google Scholar] [CrossRef] [PubMed]
- Dolinay, T.; Kim, Y.S.; Howrylak, J.; Hunninghake, G.M.; An, C.H.; Fredenburgh, L.; Massaro, A.F.; Rogers, A.; Gazourian, L.; Nakahira, K.; et al. Inflammasome-regulated Cytokines Are Critical Mediators of Acute Lung Injury. Am. J. Respir. Crit. Care Med. 2012, 185, 1225–1234. [Google Scholar] [CrossRef] [PubMed]
- Singer, M.; Deutschman, C.S.; Seymour, C.W.; Shankar-Hari, M.; Annane, D.; Bauer, M.; Bellomo, R.; Bernard, G.R.; Chiche, J.-D.; Coopersmith, C.M.; et al. The Third International Consensus Definitions for Sepsis and Septic Shock (Sepsis-3). JAMA 2016, 315, 801–810. [Google Scholar] [CrossRef]
- Sutterwala, F.S.; Mijares, L.A.; Li, L.; Ogura, Y.; Kazmierczak, B.I.; Flavell, R.A. Immune recognition of Pseudomonas aeruginosa mediated by the IPAF/NLRC4 inflammasome. J. Exp. Med. 2007, 204, 3235–3245. [Google Scholar] [CrossRef]
- Ullah, I.; Bayes, H.K.; Li, D.; Simon, A.K.; Jabir, M.S.; Hopkins, L.; Ritchie, N.D.; Tourlomousis, P.; Lupton, A.; Puleston, D.; et al. Mitochondrial damage contributes to Pseudomonas aeruginosa activation of the inflammasome and is downregulated by autophagy. Autophagy 2015, 11, 166–182. [Google Scholar] [CrossRef]


Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hardy, K.S.; Tessmer, M.H.; Frank, D.W.; Audia, J.P. Perspectives on the Pseudomonas aeruginosa Type III Secretion System Effector ExoU and Its Subversion of the Host Innate Immune Response to Infection. Toxins 2021, 13, 880. https://doi.org/10.3390/toxins13120880
Hardy KS, Tessmer MH, Frank DW, Audia JP. Perspectives on the Pseudomonas aeruginosa Type III Secretion System Effector ExoU and Its Subversion of the Host Innate Immune Response to Infection. Toxins. 2021; 13(12):880. https://doi.org/10.3390/toxins13120880
Chicago/Turabian StyleHardy, Kierra S., Maxx H. Tessmer, Dara W. Frank, and Jonathon P. Audia. 2021. "Perspectives on the Pseudomonas aeruginosa Type III Secretion System Effector ExoU and Its Subversion of the Host Innate Immune Response to Infection" Toxins 13, no. 12: 880. https://doi.org/10.3390/toxins13120880
APA StyleHardy, K. S., Tessmer, M. H., Frank, D. W., & Audia, J. P. (2021). Perspectives on the Pseudomonas aeruginosa Type III Secretion System Effector ExoU and Its Subversion of the Host Innate Immune Response to Infection. Toxins, 13(12), 880. https://doi.org/10.3390/toxins13120880