Prevalence, Enterotoxigenic Potential and Antimicrobial Resistance of Staphylococcus aureus and Methicillin-Resistant Staphylococcus aureus (MRSA) Isolated from Algerian Ready to Eat Foods

 ,
,  and
and 
Abstract
:1. Introduction
2. Results
2.1. Prevalance of Staphylococcus aureus in Algerian Ready to Eat Foods
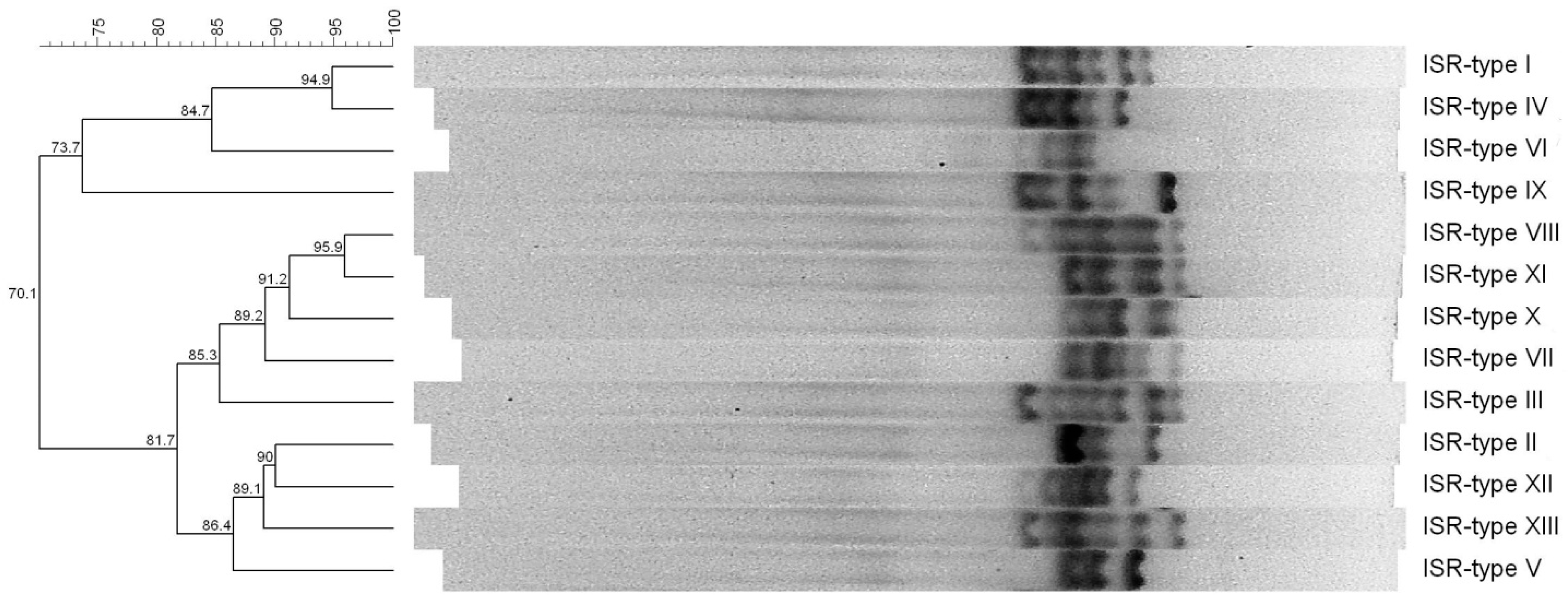
2.2. 16S-23S rDNA Intergenic Spacer Region (ISR)-PCR, Staphylococcal Enterotoxin Gene (SEg)-Typing and Presence of tst and mecA Genes
2.3. Characterization of the Staphylococcal Chromosomal Cassette mec (SCCmec)
2.4. Antimicrobial Resistance
3. Discussion
4. Conclusions
5. Materials and Methods
5.1. Sampling
5.2. Isolation of Presumptive Staphylococcus aureus
5.3. DNA Extraction
5.4. Identification of Staphylococcus aureus
5.5. Staphylococcal Enterotoxin Gene (SEg-) Typing and Real Time PCR of the Enterotoxin Gene Cluster (egc)
5.6. 16S-23S rDNA Intergenic Spacer Region PCR (ISR-PCR)
5.7. Detection of tst and mecA Genes and Characterization of the Staphylococcal Chromosomal Cassette mec (SCCmec)
5.8. Antimicrobial Susceptibility Testing
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Andjelković, U.; Šrajer Gajdošik, M.; Gašo-Sokač, D.; Martinović, T.; Josić, D. Foodomics and Food Safety: Where We Are. Food Technol. Biotechnol. 2017, 55, 290–307. [Google Scholar] [CrossRef]
- Jung, Y.; Jang, H.; Matthews, K.R. Effect of the food production chain from farm practices to vegetable processing on outbreak incidence. Microb. Biotechnol. 2014, 7, 517–527. [Google Scholar] [CrossRef] [PubMed]
- World Health Organization. WHO Estimates of the Global Burden of Food Borne Diseases; Foodborne Disease Burden Epidemiology Reference Group (2007–2015); World Health Organization: Geneva, Switzerland, 2015; 255p. [Google Scholar]
- Flynn, K.; Pérez Villarreal, B.; Barranco, A.; Belc, N.; Björnsdóttir, B.; Fusco, V.; Rainieri, S.; Smaradóttir, S.E.; Smeu, I.; Teixeira, P.; et al. An introduction to current food safety needs. Trends Food Sci. Technol. 2019, 84, 1–3. [Google Scholar] [CrossRef] [Green Version]
- McLinden, T.; Sargeant, J.M.; Thomas, M.K.; Papadopoulos, A.; Fazil, A. Component costs of foodborne illness: A scoping review. BMC Public Health 2014, 14, 509. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Franz, C.M.A.P.; den Besten, H.M.W.; Bohnlein, C.; Gareis, M.; Zwietering, M.H.; Fusco, V. Microbial food safety in the 21st century: Emerging challenges and foodborne pathogenic bacteria. Trends Food Sci Technol. 2018, 81, 155–158. [Google Scholar] [CrossRef]
- Foster, T.J. The Staphylococcus aureus “superbug”. J. Clin. Investig. 2004, 114, 1693–1696. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tong, S.Y.C.; Davis, J.S.; Eichenberger, E.; Holland, T.L.; Fowler, V.G., Jr. Staphylococcus aureus infections: Epidemiology, pathophysiology, clinical manifestations, and management. Clin. Microbiol. Rev. 2015, 28, 603–661. [Google Scholar] [CrossRef] [Green Version]
- Puah, S.M.; Chua, K.H.; Tan, J.A. Virulence factors and antibiotic susceptibility of Staphylococcus aureus isolates in ready-to-eat foods: Detection of S. aureus contamination and a high prevalence of virulence genes. Int. J. Environ. Res. Public Health 2016, 13, 199. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Havelaar, A.H.; Kirk, M.D.; Torgerson, P.R.; Gibb, H.J.; Hald, T.; Lake, R.J.; Praet, N.; Bellinger, D.C.; de Silva, N.R.; Gargouri, N.; et al. World Health Organization global estimates and regional comparisons of the burden of foodborne disease in 2010. PLoS Med. 2015, 12, e1001923. [Google Scholar] [CrossRef] [Green Version]
- Fusco, V.; Quero, G.M.; Morea, M.; Blaiotta, G.; Visconti, A. Rapid and reliable identification of Staphylococcus aureus harbouring the enterotoxin gene cluster (egc) and quantitative detection in raw milk by real time PCR. Int. J. Food Microbiol. 2011, 144, 528–537. [Google Scholar] [CrossRef]
- Leyral, G.; Vierling, E. Microbiologie et Toxicologie des Aliments: Hygiène et Sécurité Alimentaires, 4th ed.; Doin: Bordeaux, France, 2007; 290p. [Google Scholar]
- Chieffi, D.; Fanelli, F.; Cho, G.-S.; Schubert, J.; Blaiotta, G.; Franz, C.M.A.P.; Bania, J.; Fusco, V. Novel insights into the enterotoxigenic potential and genomic background of Staphylococcus aureus isolated from raw milk. Food Microbiol. 2020, 90, 103482. [Google Scholar] [CrossRef]
- Cafini, F.; Le Thuy, N.T.; Román, F.; Prieto, J.; Dubrac, S.; Msadek, T.; Morikawa, K. Methodology for the Study of Horizontal Gene Transfer in Staphylococcus aureus. J. Vis. Exp. 2017, 121, 55087. [Google Scholar] [CrossRef]
- Rajkovic, A.; Jovanovic, J.; Monteiro, S.; Decleer, M.; Andjelkovic, M.; Foubert, A.; Beloglazova, N.; Tsilla, V.; Sas, B.; Madder, A.; et al. Detection of toxins involved in foodborne diseases caused by Gram-positive bacteria. Compr. Rev. Food Sci. Food Saf. 2020, 19, 1605–1657. [Google Scholar] [CrossRef] [PubMed]
- Le Loir, Y.; Baron, F.; Gautier, M. Staphylococcus aureus and food poisoning. Genet. Mol. Res. 2003, 2, 63–76. [Google Scholar] [PubMed]
- Gomes, B.C.; Franco, B.D.G.M.; De Martinis, E.C.P. Microbiological food safety issues in Brazil: Bacterial pathogens. Foodborne Pathog. Dis. 2013, 10, 197–205. [Google Scholar] [CrossRef] [PubMed]
- Chambers, H.F.; Deleo, F.R. Waves of resistance: Staphylococcus aureus in the antibiotic era. Nat. Rev. Microbiol. 2009, 7, 629–641. [Google Scholar] [CrossRef] [PubMed]
- Shlaes, D.M.; Projan, S.J. Antimicrobial resistance versus the discovery and development of newantimicrobials. In BT—Antimicrobial Drug Resistance: Mechanisms of Drug Resistance; Mayers, D.L., Ed.; Humana Press: Totowa, NJ, USA, 2009; pp. 43–50. [Google Scholar]
- Fishovitz, J.; Hermoso, J.A.; Chang, M.; Mobashery, S. Penicillin-binding protein 2a of methicillin-resistant Staphylococcus aureus. IUBMB Life 2014, 66, 572–577. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, X.; Li, G.; Xia, X.; Yang, B.; Xi, M.; Meng, J. Antimicrobial susceptibility and molecular typing of methicillin-resistant Staphylococcus aureus in retail foods in Shaanxi, China. Foodborne Pathog. Dis. 2014, 11, 281–286. [Google Scholar] [CrossRef] [PubMed]
- Islam, M.A.; Parveen, S.; Rahman, M.; Huq, M.; Nabi, A.; Khan, Z.U.M.; Ahmed, N.; Wagenaar, J.A. Occurrence and Characterization of Methicillin Resistant Staphylococcus aureus in Processed Raw Foods and Ready-to-Eat Foods in an Urban Setting of a Developing Country. Front. Microbiol. 2019, 10, 503. [Google Scholar] [CrossRef]
- Fusco, V.; Chieffi, D.; Fanelli, F.; Logrieco, A.F.; Cho, G.-S.; Kabisch, J.; Böhnlein, C.; Franz, C.M.A.P. Microbial quality and safety of milk and milk products in the 21st century. Compr. Rev. Food Sci. Food Saf. 2020, 19, 2013–2049. [Google Scholar] [CrossRef] [PubMed]
- Petinaki, E.; Spiliopoulou, L. Methicillin-resistant Staphylococcus aureus among companion and food-chain animals: Impact of human contacts. Clin. Microbiol. Infect. 2012, 18, 626–634. [Google Scholar] [CrossRef] [Green Version]
- Chiang, Y.C.; Liao, W.W.; Fan, C.M.; Pai, W.Y.; Chiou, C.S.; Tsen, H.Y. PCR detection of Staphylococcal enterotoxins (SEs) N, O, P, Q, R, U, and survey of SE types in Staphylococcus aureus isolates from food-poisoning cases in Taiwan. Int. J. Food Microbiol. 2008, 15, 66–73. [Google Scholar] [CrossRef] [PubMed]
- Chomvarin, C.; Chantarasuk, Y.; Srigulbutr, S.; Chareonsudjai, S.; Chaicumpar, K. Enteropathogenic bacteria and enterotoxin-producing Staphylococcus aureus isolated from ready-to-eat foods in Khon Kaen, Thailand. Southeast Asian J. Trop. Med. Public Health 2006, 37, 983–990. [Google Scholar] [PubMed]
- Oh, S.K.; Lee, N.; Cho, Y.S.; Shin, D.B.; Choi, S.Y.; Koo, M. Occurrence of toxigenic Staphylococcus aureus in ready-to-eat food in Korea. J. Food Prot. 2007, 70, 1153–1158. [Google Scholar] [CrossRef]
- Lozano, C.; Gharsa, H.; Ben Slama, K.; Zarazaga, M.; Torres, C. Staphylococcus aureus in animals and food: Methicillin resistance, prevalence and population structure. A review in the African Continent. Microorganisms 2016, 4, 12. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Paudyal, N.; Anihouvi, V.; Hounhouigan, J.; Matsheka, M.I.; Sekwati-Monang, B.; Amoa-Awua, W.; Atter, A.; Ackah, N.B.; Mbugua, S.; Asagbra, A.; et al. Prevalence of foodborne pathogens in food from selected African countries—A meta-analysis. Int. J. Food Microbiol. 2017, 249, 35–43. [Google Scholar] [CrossRef]
- Oliveira, D.C.; de Lencastre, H. Multiplex PCR strategy for rapid identification of structural types and variants of the mec element in methicillin-resistant Staphylococcus aureus. Antimicrob. Agents Chemother. 2002, 46, 2155–2161. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chaalal, W.; Chaalal, N.; Bourafa, N.; Kihal, M.; Diene, S.M.; Rolain, J.-M. Characterization of Staphylococcus aureus Isolated from Food Products in Western Algeria. Foodborne Pathog. Dis. 2018, 15, 353–360. [Google Scholar] [CrossRef] [PubMed]
- Titouche, Y.; Houali, K.; Ruiz-Ripa, L.; Vingadassalon, N.; Nia, Y.; Fatihi, A.; Cauquil, A.; Bouchez, P.; Bouhier, L.; Torres, C.; et al. Enterotoxin genes and antimicrobial resistance in Staphylococcus aureus isolated from food products in Algeria. J. Appl. Microbiol. 2020, 129, 1043–1052. [Google Scholar] [CrossRef] [PubMed]
- Madoroba, E.; Magwedere, K.; Chaora, N.S.; Matle, I.; Muchadeyi, F.; Mathole, M.A.; Pierneef, R. Microbial communities of meat and meat products: An exploratory analysis of the product quality and safety at selected enterprises in South Africa. Microorganisms 2021, 9, 507. [Google Scholar] [CrossRef]
- Saad, M.S.; Fatin, S.H.; Fahim, A.S.; Marionette, Z.N.; Marwa, Z.S. Prevalence of methicillin-resistant Staphylococcus aureus in some ready-to-eat meat products. Am. J. Biomed. Sci. Res. 2019, 4, 461–465. [Google Scholar] [CrossRef]
- Ire, S.F.; Imuh, V.T. Bacteriological quality evaluation and safety of randomly selected ready-to-eat foods sold in Port Harcourt city, Nigeria. J. Appl. Life Sci. Int. 2016, 7, 1–10. [Google Scholar] [CrossRef]
- Abdeen, E.; Mousa, W.; Abdelsalam, S.; Heikal, H.; Shawish, R.; Nooruzzaman, M.; Soliman, M.; Batiha, G.; Hamad, A.; Abdeen, A. Prevalence and Characterization of Coagulase Positive Staphylococci from Food Products and Human Specimens in Egypt. Antibiotics 2021, 10, 75. [Google Scholar] [CrossRef] [PubMed]
- Argudín, M.Á.; Mendoza, M.C.; Rodicio, M.R. Food Poisoning and Staphylococcus aureus Enterotoxins. Toxins 2010, 2, 1751–1773. [Google Scholar] [CrossRef] [PubMed]
- Sezer, Ç.; Özgür, Ç.; Aksem, A.; Leyla, V. Food handlers: A bridge in the journey of enterotoxigenic MRSA in food. J. Verbr. Lebensm. 2015, 10, 123–129. [Google Scholar] [CrossRef]
- Kadariya, J.; Smith, T.C.; Thapaliya, D. Staphylococcus aureus and staphylococcal food-borne disease: An ongoing challenge in public health. BioMed Res. Int. 2014, 827965. [Google Scholar] [CrossRef] [Green Version]
- Prado Martin, J.G.; de Oliveira, E.; Silva, G.; da Fonseca, C.R.; Morales, C.B.; Souza Pamplona Silva, C.; Miquelluti, D.L.; Porto, E. Efficiency of a cleaning protocol for the removal of enterotoxigenic Staphylococcus aureus strains in dairy plants. Int. J. Food Microbiol. 2016, 238, 295–301. [Google Scholar] [CrossRef] [PubMed]
- Mahami, T.; Amafu-Dey, H.; Odonkor, S. Microbial food safety risk: Cooked and smoked sausages as a potential source. Int. J. Biol. Pharm. Allied Sci. 2012, 1, 99–107. [Google Scholar]
- Oguttu, J.W.; McCrindle, C.M.E.; Makita, K.; Grace, D. Investigation of the food value chain of ready-to-eat chicken and the associated risk for staphylococcal food poisoning in Tshwane Metropole, South Africa. Food Control 2014, 45, 87–94. [Google Scholar] [CrossRef] [Green Version]
- Shiningeni, D.; Chimwamurombe, P.; Shilangale, R.; Misihairabgwi, J. Prevalence of pathogenic bacteria in street vended ready-to-eat meats in Windhoek, Namibia. Meat Sci. 2019, 148, 223–228. [Google Scholar] [CrossRef]
- Bania, J.; Dabrowska, A.; Bystron, J.; Korzekwa, K.; Chrzanowska, J.; Molenda, J. Distribution of newly described enterotoxin-like genes in Staphylococcus aureus from food. Int. J. Food Microbiol. 2006, 108, 36–41. [Google Scholar] [CrossRef] [PubMed]
- Chao, G.; Bao, G.; Cao, Y.; Yan, W.; Wang, Y.; Zhang, X.; Zhou, L.; Wu, Y. Prevalence and diversity of enterotoxin genes with genetic background of Staphylococcus aureus isolates from different origins in China. Int. J. Food Microbiol. 2015, 211, 142–147. [Google Scholar] [CrossRef] [PubMed]
- Aung, M.S.; Urushibara, N.; Kawaguchiya, M.; Ito, M.; Habadera, S.; Kobayashi, N. Prevalence and Genetic Diversity of Staphylococcal Enterotoxin (-Like) Genes sey, selw, selx, selz, sel26 and sel27 in Community-Acquired Methicillin-Resistant Staphylococcus aureus. Toxins 2020, 12, 347. [Google Scholar] [CrossRef] [PubMed]
- Wilson, G.J.; Tuffs, S.W.; Wee, B.A.; Seo, K.S.; Park, N.; Connelley, T.; Guinane, C.M.; Morrison, W.I.; Fitzgerald, J.R. Bovine Staphylococcus aureus superantigens stimulate the entire T cell repertoire of cattle. Infect. Immun. 2018, 86, e00505-18. [Google Scholar] [CrossRef] [Green Version]
- Bianchi, D.; Gallina, S.; Bellio, A.; Chiesa, F.; Civera, T.; Decastelli, L. Enterotoxin gene profiles of Staphylococcus aureus isolated from milk and dairy products in Italy. Lett. Appl. Microbiol. 2014, 58, 190–196. [Google Scholar] [CrossRef]
- Song, M.; Shi, C.; Xu, X.; Shi, X. Molecular typing and virulence gene profiles of enterotoxin gene cluster (egc)-positive Staphylococcus aureus isolates obtained from various food and clinical specimens. Foodborne Pathog. Dis. 2016, 13, 592–601. [Google Scholar] [CrossRef]
- Jarraud, S.; Peyrat, M.A.; Lim, A.; Tristan, A.; Bes, M.; Mougel, C.; Etienne, J.; Vandenesch, F.; Bonneville, M.; Lina, G. egc, a highly prevalent operon of enterotoxin gene, forms a putative nursery of superantigens in Staphylococcus aureus. J. Immunol. 2001, 166, 669–677. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Collery, M.M.; Smyth, C.J. Rapid differentiation of Staphylococcus aureus isolates harbouring egc loci with pseudogenes ψent1 and ψent2 and the selu or seluv gene using PCR-RFLP. J. Med. Microbiol. 2007, 56, 208–216. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Noto, M.J.; Archer, G.L. A subset of Staphylococcus aureus strains harboring staphylococcal cassette chromosome mec (SCCmec) type IV is deficient in CcrAB-mediated SCCmec excision. Antimicrob. Agents Chemother. 2006, 50, 2782–2788. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, X.; Yu, S.; Wu, Q.; Zhang, J.; Wu, S.; Rong, D. Multilocus sequence typing and virulence-associated gene profile analysis of Staphylococcus aureus isolates from retail ready-to-eat food in China. Front. Microbiol. 2018, 9, 197. [Google Scholar] [CrossRef] [PubMed]
- Johler, S.; Giannini, P.; Jermini, M.; Hummerjohann, J.; Baumgartner, A.; Stephan, R. Further evidence for staphylococcal food poisoning outbreaks caused by egc-encoded enterotoxins. Toxins 2015, 7, 997–1004. [Google Scholar] [CrossRef] [PubMed]
- Umeda, K.; Nakamura, H.; Yamamoto, K.; Nishina, N.; Yasufuku, K.; Hirai, Y.; Hirayama, T.; Goto, K.; Hase, A.; Ogasawara, J. Molecular and epidemiological characterization of staphylococcal foodborne outbreak of Staphylococcus aureus harboring seg, sei, sem, sen, seo, and selu genes without production of classical enterotoxins. Int. J. Food Microbiol. 2017, 256, 30–35. [Google Scholar] [CrossRef]
- Ikeda, T.; Tamate, N.; Yamaguchi, K.; Makino, S. Mass outbreak of food poisoning disease caused by small amounts of staphylococcal enterotoxins A and H. Appl. Environ. Microbiol. 2005, 71, 2793–2795. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jørgensen, H.J.; Mathisen, T.; Lovseth, A.; Omoe, K.; Qvale, K.S.; Loncarevic, S. An outbreak of staphylococcal food poisoning caused by enterotoxin H in mashed potato made with raw milk. FEMS Microbiol. Lett. 2005, 252, 67–72. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Benkerroum, N. Staphylococcal enterotoxins and enterotoxin-like toxins with special reference to dairy products: An overview. Crit. Rev. Food Sci. Nutr. 2017, 58, 1943–1970. [Google Scholar] [CrossRef]
- Bissong, M.E.A.; Tahnteng, B.F.; Ateba, C.N.; Akoachere, J.-F.T.K. Pathogenic Potential and Antimicrobial Resistance Profile of Staphylococcus aureus in Milk and Beef from the Northwest and Southwest Regions of Cameroon. BioMed Res. Int. 2020, 2020, 1–12. [Google Scholar] [CrossRef]
- Omwenga, I.; Aboge, G.O.; Mitema, E.S.; Obiero, G.; Ngaywa, C.; Ngwili, N.; Wamwere, G.; Wainaina, M.; Bett, B. Staphylococcus aureus enterotoxin genes detected in milk from various livestock species in northern pastoral region of Kenya. Food Control 2019, 103, 126–132. [Google Scholar] [CrossRef]
- Chen, Q.; Xie, S. Genotypes, enterotoxin gene profiles, and antimicrobial resistance of Staphylococcus aureus associated with foodborne outbreaks in Hangzhou, China. Toxins 2019, 11, 307. [Google Scholar] [CrossRef] [Green Version]
- Ciupescu, L.-M.; Auvray, F.; Nicorescu, I.M.; Meheut, T.; Ciupescu, V.; Lardeux, A.-L.; Tanasuica, R.; Hennekinne, J.-A. Characterization of Staphylococcus aureus strains and evidence for the involvement of non-classical enterotoxin genes in food poisoning outbreaks. FEMS Microbiol. Lett. 2018, 365, fny139. [Google Scholar] [CrossRef]
- Denayer, S.; Delbrassinne, L.; Nia, Y.; Botteldoorn, N. Food-Borne outbreak investigation and molecular typing: High diversity of Staphylococcus aureus strains and importance of toxin detection. Toxins 2017, 9, 407. [Google Scholar] [CrossRef] [Green Version]
- Sato’o, Y.; Omoe, K.; Naito, I.; Ono, H.K.; Nakane, A.; Sugai, M.; Yamagishi, N.; Hu, D. Molecular epidemiology and identification of a Staphylococcus aureus clone causing food poisoning outbreaks in Japan. J. Clin. Microbiol. 2014, 5, 2637–2640. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Omoe, K.; Hu, D.-L.; Ono, H.K.; Shimizu, S.; Takahashi-Omoe, H.; Nakane, A.; Uchiyama, T.; Shinagawa, K.; Imanishi, K. Emetic potentials of newly identified staphylococcal enterotoxin-like toxins. Infect. Immun. 2013, 81, 3627–3631. [Google Scholar] [CrossRef] [Green Version]
- Ono, H.K.; Omoe, K.; Imanishi, K.; Iwakabe, Y.; Hu, D.L.; Kato, H.; Saito, N.; Nakane, A.; Uchiyama, T.; Shinagawa, K. Identification and characterization of two novel staphylococcal enterotoxins, types S and T. Infect. Immun. 2008, 76, 4999–5005. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Aydin, A.; Sudagidan, M.; Muratoglu, K. Prevalence of staphylococcal enterotoxins, toxin genes and genetic-relatedness of foodborne Staphylococcus aureus strains isolated in the Marmara Region of Turkey. Int. J. Food Microbiol. 2011, 148, 99–106. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jans, C.; Merz, A.; Johler, S.; Younan, M.; Tanner, S.A.; Kaindi, D.W.M.; Wangoh, J.; Bonfoh, B.; Meile, L.; Tasara, T. East and West African milk products are reservoirs for human and livestock-associated Staphylococcus aureus. Food Microbiol. 2017, 65, 64–73. [Google Scholar] [CrossRef]
- Wongboot, W.; Chomvarin, C.; Namwat, W. Phenotypic and genotypic detection of enterotoxins, toxic shock syndrome toxin-1 and of methicillin resistance in Staphylococcus aureus isolated from retail ready-to-eat foods in Northeastern Thailand. Southeast Asian J. Trop. Med. Public Health 2015, 46, 97–104. [Google Scholar] [PubMed]
- Durand, G.; Bes, M.; Meugnier, H.; Enright, M.C.; Forey, F.; Liassine, N.; Wenger, A.; Kikuchi, K.; Lina, G.; Vandenesch, F.; et al. Detection of new methicillin-resistant Staphylococcus aureus clones containing the toxic shock syndrome toxin 1 gene responsible for hospital- and community-acquired infections in France. J. Clin. Microbiol. 2006, 44, 847–853. [Google Scholar] [CrossRef] [Green Version]
- Sultan, A.M.; Nabiel, Y. Association of tsst-1 and pvl with mecA genes among clinical Staphylococcus aureus isolates from a tertiary care hospital. J. Pure Appl. Microbiol. 2019, 13, 855–864. [Google Scholar] [CrossRef] [Green Version]
- Achek, R.; Hotzel, H.; Cantekin, Z.; Nabi, I.; Hamdi, T.M.; Neubauer, H.; El-Adawy, H. Emerging of antimicrobial resistance in staphylococci isolated from clinical and food samples in Algeria. BMC Res. Notes 2018, 11, 663. [Google Scholar] [CrossRef]
- Oniciuc, E.-A.; Nicolau, A.I.; Hernández, M.; Rodríguez-Lázaro, D. Presence of methicillin-resistant Staphylococcus aureus in the food chain. Trends Food Sci. Technol. 2017, 61, 49–59. [Google Scholar] [CrossRef]
- Crago, B.; Ferrato, C.; Drews, S.J.; Svenson, L.W.; Tyrrell, G.; Louie, M. Prevalence of Staphylococcus aureus and methicillin-resistant S. aureus (MRSA) in food samples associated with foodborne illness in Alberta, Canada from 2007 to 2010. Food Microbiol. 2012, 32, 202–205. [Google Scholar] [CrossRef] [PubMed]
- Mairi, A.; Touati, A.; Lavigne, J.-P. Methicillin-Resistant Staphylococcus aureus ST80 Clone: A systematic review. Toxins 2020, 12, 119. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Monecke, S.; Slickers, P.; Gawlik, D.; Müller, E.; Reissig, A.; Ruppelt-Lorz, A.; Cortez de Jäckel, S.; Feßler, A.T.; Frank, M.; Hotzel, H.; et al. Variability of SCCmec elements in livestock-associated CC398 MRSA. Vet. Microbiol. 2018, 217, 36–46. [Google Scholar] [CrossRef]
- Aung, K.T.; Hsu, L.Y.; Koh, T.H.; Hapuarachchi, H.C.; Chau, M.L.; Gutiérrez, R.A.; Ng, L.C.; Hapuarachchi, H.C. Prevalence of methicillin-resistant Staphylococcus aureus (MRSA) in retail food in Singapore. Antimicrob. Resist. Infect. Control 2017, 6, 94. [Google Scholar] [CrossRef] [Green Version]
- Liao, F.; Gu, W.; Yang, Z.; Mo, Z.; Fan, L.; Guo, Y.; Fu, X.; Xu, W.; Li, C.; Dai, J. Molecular characteristics of Staphylococcus aureus isolates from food surveillance in southwest China. BMC Microbiol. 2018, 18, 91. [Google Scholar] [CrossRef] [PubMed]
- Rodríguez-Lázaro, D.; Oniciuc, E.-A.; García, P.G.; Gallego, D.; Fernández-Natal, I.; Dominguez-Gil, M.; Eiros-Bouza, J.M.; Wagner, M.; Nicolau, A.I.; Hernández, M. Detection and characterization of Staphylococcus aureus and methicillin-resistant S. aureus in foods confiscated in EU Borders. Front. Microbiol. 2017, 8, 1344. [Google Scholar] [CrossRef]
- Sergelidis, D.; Angelidis, A. Methicillin-resistant Staphylococcus aureus: A controversial food-borne pathogen. Lett. Appl. Microbiol. 2017, 64, 409–418. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jones, T.F.; Kellum, M.E.; Porter, S.S.; Bell, M.; Schaffner, W. An outbreak of community-acquired foodborne illness caused by methicillin-resistant Staphylococcus aureus. Emerg. Infect. Dis. 2002, 8, 82–84. [Google Scholar] [CrossRef] [PubMed]
- Pu, S.; Wang, F.; Ge, B. Characterization of toxin genes and antimicrobial susceptibility of Staphylococcus aureus isolates from Louisiana retail meats. Foodborne Pathog. Dis. 2011, 8, 299–306. [Google Scholar] [CrossRef]
- Cremonesi, P.; Zottola, T.; Locatelli, C.; Pollera, C.; Castiglioni, B.; Scaccabarozzi, L.; Moroni, P. Identification of virulence factors in 16S-23S rRNA intergenic spacer genotyped Staphylococcus aureus isolated from water buffaloes and small ruminants. J. Dairy Sci. 2013, 96, 7666–7674. [Google Scholar] [CrossRef]
- Govender, V.; Madoroba, E.; Magwedere, K.; Fosgate, G.; Kuonza, L. Prevalence and risk factors contributing to antibiotic-resistant Staphylococcus aureus isolates from poultry meat products in South Africa, 2015–2016. J. S. Afr. Vet. Assoc. 2019, 90, a1738. [Google Scholar] [CrossRef] [Green Version]
- Ge, B.; Mukherjee, S.; Hsu, C.-H.; Davis, J.A.; Tran, T.T.T.; Yang, Q.; Abbott, J.W.; Ayers, S.L.; Young, S.R.; Crarey, E.T.; et al. MRSA and multidrug-resistant Staphylococcus aureus in U.S. retail meats, 2010–2011. Food Microbiol. 2017, 62, 289–297. [Google Scholar] [CrossRef]
- Wang, W.; Baloch, Z.; Jiang, T.; Zhang, C.; Peng, Z.; Li, F.; Fanning, S.; Ma, A.; Xu, J. Enterotoxigenicity and antimicrobial resistance of Staphylococcus aureus isolated from retail food in China. Front. Microbiol. 2017, 8, 2256. [Google Scholar] [CrossRef] [PubMed]
- Ma, Y.; Zhao, Y.; Tang, J.; Tang, C.; Chen, J.; Liu, J. Antimicrobial susceptibility and presence of resistance & enterotoxins/enterotoxinlikes genes in Staphylococcus aureus from food. CyTA J. Food 2018, 16, 76–84. [Google Scholar] [CrossRef]
- Abdalrahman, L.S.; Stanley, A.; Wells, H.; Fakhr, M.K. Isolation, Virulence, and Antimicrobial Resistance of Methicillin-Resistant Staphylococcus aureus (MRSA) and Methicillin Sensitive Staphylococcus aureus (MSSA) Strains from Oklahoma Retail Poultry Meats. Int. J. Environ. Res. Public Health 2015, 12, 6148–6161. [Google Scholar] [CrossRef] [PubMed]
- Kim, Y.H.; Kim, H.S.; Kim, S.; Kim, M.; Kwak, H.S. Prevalence and characteristics of antimicrobial-resistant Staphylococcus aureus and methicillin-resistant Staphylococcus aureus from retail meat in Korea. Food Sci. Anim. Resour. 2020, 40, 758–771. [Google Scholar] [CrossRef]
- Foster, T.J. Antibiotic resistance in Staphylococcus aureus. Current status and future prospects. FEMS Microbiol. Rev. 2017, 41, 430–449. [Google Scholar] [CrossRef] [PubMed]
- Anonymous. Microbiology of Food and Animal Feeding Stuffs—Horizontal Method for the Enumeration of Coagulase-Positive Staphylococci (Staphylococcus aureus and Other Species)—Part 1: Technique Using Baird-Parker Agar Medium; ISO: London, UK, 1999; ISO 6888-1. [Google Scholar]
- Brakstad, O.G.; Aasbakk, K.; Maeland, J.A. Detection of Staphylococcus aureus by polymerase chain reaction amplification of the nuc gene. J. Clin. Microbiol. 1992, 30, 1654–1660. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Blaiotta, G.; Ercolini, D.; Pennacchia, C.; Fusco, V.; Casaburi, A.; Pepe, O.; Villani, F. PCR detection of staphylococcal enterotoxin genes in Staphylococcus spp. strains isolated from meat and dairy products. Evidence for new variants of seG and seI in S. aureus AB-8802. J. Appl. Microbiol. 2004, 97, 719–730. [Google Scholar] [CrossRef]
- Sergeev, N.; Volokhov, D.; Chizhikov, V.; Rasooly, A. Simultaneous analysis of multiple staphylococcal enterotoxin genes by an oligonucleotide microarray assay. J. Clin. Microbiol. 2004, 42, 2134–2143. [Google Scholar] [CrossRef] [Green Version]
- Collery, M.M.; Smyth, D.S.; Tumilty, J.J.; Twohig, J.M.; Smyth, C.J. Associations between enterotoxin gene cluster types egc1, egc2 and egc3, agr types, enterotoxin and enterotoxin-like gene profiles, and molecular typing characteristics of human nasal carriage and animal isolates of Staphylococcus aureus. J. Med. Microbiol. 2009, 58, 13–25. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schubert, J.; Podkowik, M.; Bystron, J.; Bania, J. Production of staphylococcal enterotoxins in microbial broth and milk by Staphylococcus aureus strains harboring seh gene. Int. J. Food Microbiol. 2016, 235, 36–45. [Google Scholar] [CrossRef] [PubMed]
- Thomas, D.Y.; Jarraud, S.; Lemercier, B.; Cozon, G.; Echasserieau, K.; Etienne, J.; Gougeon, M.-L.; Lina, G.; Vandenesch, F. Staphylococcal enterotoxin-like toxins U2 and V, two new staphylococcal superantigens arising from recombination within the enterotoxin gene cluster. Infect. Immun. 2006, 74, 4724–4734. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fusco, V.; Quero, G.M.; Stea, G.; Morea, M.; Visconti, A. Novel PCR-based identification of Weissella confusa using an AFLP-derived marker. Int. J. Food Microbiol. 2011, 145, 437–443. [Google Scholar] [CrossRef]
- Di Lena, M.; Quero, G.M.; Santovito, E.; Verran, J.; De Angelis, M.; Fusco, V. A selective medium for isolation and accurate enumeration of Lactobacillus casei-group members in probiotic milks and dairy products. Int. Dairy J. 2015, 47, 27–36. [Google Scholar] [CrossRef]
- Johnson, W.M.; Tyler, S.D.; Ewan, E.P.; Ashton, F.E.; Pollard, D.R.; Rozee, K.R. Detection of genes for enterotoxins, exfoliative toxins, and toxic shock syndrome toxin 1 in Staphylococcus aureus by the polymerase chain reaction. J. Clin. Microbiol. 1991, 29, 426–430. [Google Scholar] [CrossRef] [Green Version]
- CLSI (Clinical and Laboratory Standards Institute). Performance Standards for Antimicrobial Susceptibility Testing; Twenty-Second Informational Supplement; CLSI Document M100-S22; Clinical and Laboratory Standards Institute: Wayne, PA, USA, 2012. [Google Scholar]
- EUCAST (The European Committee on Antimicrobial Susceptibility Testing). Breakpoint Tables for Interpretation of MICs and Zone Diameters, Version 2.0. 2012. Available online: http://www.eucast.org/clinical_breakpoints/ (accessed on 27 October 2021).


{kind=link}
{kind=link}
| Food Category | Number of Samples | S. aureus Positive Samples (% of Positive Samples) | CPS Count a | CPS Count Range b |
|---|---|---|---|---|
| Meat/fish-based foods | 55 | 21 (38.2%) | 3.48 ± 0.86 | 1.40 ± 0.12–4.49 ± 0.26 |
| Vegetable-based foods | 72 | 16 (22.2%) | 3.19 ± 1.03 | 1.00 ± 0.5–4.43 ± 0.15 |
| Pastries | 43 | 7 (16.3%) | 3.57 ± 0.80 | 2.84 ± 0.11–5.11 ± 0.24 |
| Cereals | 17 | 3 (17.6%) | 2.40 ± 1.10 | 2.08 ± 0.16–4.13 ± 0.14 |
| Various foods (milk- and egg-based foods) | 20 | 1 (5.0%) | 1.70 ± 0.10 | – |
| Total | 207 | 48 (23.2%) | 3.38 ± 0.94 | 1.00 ± 0.5–5.11 ± 0.24 |
| Isolate | Origin | ISR-Type | SEg-Type a | tst | mecA (SCCmec Type) |
|---|---|---|---|---|---|
| SA01 | Potato in sauce b | I | 1 | - | - |
| SA06 | Chicken c | I | 1 | - | - |
| SA11 | Couscous with meat c | I | 1 | - | - |
| SA13 | Rice d | I | 1 | - | - |
| SA23 | Minced meat c | I | 1 | - | - |
| SA33 | Mashed potatoes b | I | 1 | - | - |
| SA41 | Salad b | I | 1 | - | - |
| SA43 | Rice d | I | 1 | - | - |
| SA44 | Salad b | I | 1 | - | - |
| SA50 | Salad b | I | 1 | - | - |
| SA53 | Minced meat c | I | 1 | - | - |
| SA54 | Beet salad b | I | 1 | - | - |
| SA58 | Meat c | I | 1 | - | - |
| SA73 | Salad b | I | 1 | - | - |
| SA78 | Lentil soup b | I | 1 | - | - |
| SA83 | Beans b | I | 1 | - | - |
| SA84 | Chicken c | I | 1 | - | - |
| SA86 | Sausages c | I | 1 | - | - |
| SA87 | Pastry e | I | 1 | - | - |
| SA07 | Meat c | IV | 1 | - | - |
| SA22 | Salad b | IV | 1 | - | - |
| SA38 | Spaghetti with tomato sauce b | IV | 1 | - | - |
| SA39 | Vegetable and meat sauce c | IV | 1 | - | - |
| SA49 | Potato b | IV | 1 | - | - |
| SA10 | Meat c | VI | 1 | - | - |
| SA02 | Meat c | II | 2 | - | - |
| SA30 | Meat c | II | 2 | - | - |
| SA08 | Meat c | V | 2 | - | - |
| SA19 | Pastry e | IX | 2 | - | - |
| SA24 | Lentil soup b | IX | 2 | - | - |
| SA34 | Meat c | IX | 2 | - | - |
| SA37 | Meat c | IX | 2 | - | - |
| SA82 | Sautéed beef with potato c | XII | 2 | + | - |
| SA03 | Salad b | III | 3 | - | - |
| SA04 | Meat c | III | 3 | - | - |
| SA09 | Pastry e | III | 3 | - | - |
| SA29 | Meat c | III | 3 | - | - |
| SA31 | Salad b | III | 3 | - | - |
| SA72 | Pastry e | III | 3 | - | - |
| SA05 | Pastry e | VII | 4 | - | - |
| SA18 | Pizza d | VII | 4 | - | - |
| SA55 | Pastry e | VII | 4 | - | - |
| SA56 | Pastry e | VII | 4 | - | - |
| SA59 | Russian salad b | VII | 4 | - | - |
| SA20 | Meat c | VIII | 5 | - | - |
| SA17 | Turkey pieces c | X | 6 | - | + (SCCmec IV) |
| SA46 | Braised beef c | X | 6 | - | + (SCCmec IV) |
| SA51 | Fermented milk f | XI | 7 | - | - |
| DSM 20231T | Human pleural fluid | XIII | 1 | - | - |
| ISR-Type | SEg-Type a | tst | mecA (SCCmec Type) | Origin and Number of S. aureus Isolates (% Out of the Total for Each Genotype) | Total (% of Total Isolates) | ||||
|---|---|---|---|---|---|---|---|---|---|
| Meat/Fish-Based Foods | Vegetable-Based Foods | Pastries | Cereals | Various Foods (Milk- and Egg-Based Foods) | |||||
| I | 1 | - | - | 7 (36.8%) | 9 (47.4%) | 1 (5.3%) | 2 (10.5%) | 19 (39.6%) | |
| IV | 1 | - | - | 2 (40.0%) | 3 (60.0%) | 5 (10.4%) | |||
| VI | 1 | - | - | 1 (100%) | 1 (2.1%) | ||||
| II | 2 | - | - | 2 (100%) | 2 (4.2%) | ||||
| V | 2 | - | - | 1 (100%) | 1 (2.1%) | ||||
| IX | 2 | - | - | 2 (50.0%) | 1 (25.0%) | 1 (25.0%) | 4 (8.3%) | ||
| XII | 2 | + | - | 1 (100%) | 1 (2.1%) | ||||
| III | 3 | - | - | 2 (33.3%) | 2 (33.3%) | 2 (33.3%) | 6 (12.5%) | ||
| VII | 4 | - | - | 1 (20.0%) | 3 (60.0%) | 1 (20.0%) | 5 (10.4%) | ||
| VIII | 5 | - | - | 1 (100%) | 1 (2.1%) | ||||
| X | 6 | - | + (SCCmec IV) | 2 (100%) | 2 (4.2%) | ||||
| XI | 7 | - | - | 1 (100%) | 1 (2.1%) | ||||
| Total (% of total isolates) | 21 (43.8%) | 16 (33.3%) | 7 (14.6%) | 3 (6.2%) | 1 (2.1%) | 48 (100%) | |||
| Class | Antimicrobials | S. aureus Strains | ||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| SA01 | SA02 | SA04 | SA07 | SA08 | SA10 | SA18 | SA20 | SA24 | SA46 | SA51 | SA82 | DSM 20231T | ||
| β-Lactams | P | R | S | R | R | R | R | R | R | R | R | S | R | S |
| OXA | S | S | S | S | S | S | S | S | S | R | S | S | S | |
| FOX | S | S | S | S | S | S | S | S | S | R | S | S | S | |
| Aminoglycosides | GEN | S | S | S | S | S | S | S | S | S | S | S | S | S |
| KA | S | S | S | S | S | S | R | S | S | R | S | S | S | |
| TOB | S | S | S | S | S | S | S | S | S | S | S | S | S | |
| Quinolones | OF | S | R | S | S | S | S | S | S | S | S | S | S | S |
| Macrolides | ERY | S | R | S | S | S | S | S | S | S | S | S | I | S |
| Lincosamides | L | S | R | S | S | S | S | S | S | S | S | S | S | S |
| CLI | S | S | S | S | S | S | S | S | S | S | S | S | S | |
| Streptogramins | PRI | S | S | S | S | S | S | S | S | S | S | S | S | S |
| Oxazolidinones | LZ | S | S | S | S | S | S | S | S | S | S | S | S | S |
| Glycopeptides | TEI | S | S | S | S | S | S | S | S | S | S | S | S | S |
| VAN | S | S | S | S | S | S | S | S | S | S | S | S | S | |
| Tetracyclines | TE | S | R | S | S | S | S | R | S | S | S | S | S | S |
| Fosfomycins | FOS | S | S | S | S | S | S | S | S | S | S | S | S | S |
| Nitrofurans | NIT | S | S | S | S | S | S | S | S | S | S | S | S | S |
| Steroidal | FA | S | S | I | S | S | S | S | S | S | S | S | S | S |
| Ansamycins | RIF | S | S | S | S | S | S | S | S | S | S | S | S | S |
| Folate pathway inhibitors | COT | S | S | S | S | S | S | S | S | S | S | S | S | S |
| Resistance profile a | P | OF, ERY, L, TE | P | P | P | P | P, KA, TE | P | P | P, OXA, FOX, KA | - b | P | - b | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mekhloufi, O.A.; Chieffi, D.; Hammoudi, A.; Bensefia, S.A.; Fanelli, F.; Fusco, V. Prevalence, Enterotoxigenic Potential and Antimicrobial Resistance of Staphylococcus aureus and Methicillin-Resistant Staphylococcus aureus (MRSA) Isolated from Algerian Ready to Eat Foods. Toxins 2021, 13, 835. https://doi.org/10.3390/toxins13120835
Mekhloufi OA, Chieffi D, Hammoudi A, Bensefia SA, Fanelli F, Fusco V. Prevalence, Enterotoxigenic Potential and Antimicrobial Resistance of Staphylococcus aureus and Methicillin-Resistant Staphylococcus aureus (MRSA) Isolated from Algerian Ready to Eat Foods. Toxins. 2021; 13(12):835. https://doi.org/10.3390/toxins13120835
Chicago/Turabian StyleMekhloufi, Omar Amine, Daniele Chieffi, Abdelhamid Hammoudi, Sid Ahmed Bensefia, Francesca Fanelli, and Vincenzina Fusco. 2021. "Prevalence, Enterotoxigenic Potential and Antimicrobial Resistance of Staphylococcus aureus and Methicillin-Resistant Staphylococcus aureus (MRSA) Isolated from Algerian Ready to Eat Foods" Toxins 13, no. 12: 835. https://doi.org/10.3390/toxins13120835
APA StyleMekhloufi, O. A., Chieffi, D., Hammoudi, A., Bensefia, S. A., Fanelli, F., & Fusco, V. (2021). Prevalence, Enterotoxigenic Potential and Antimicrobial Resistance of Staphylococcus aureus and Methicillin-Resistant Staphylococcus aureus (MRSA) Isolated from Algerian Ready to Eat Foods. Toxins, 13(12), 835. https://doi.org/10.3390/toxins13120835