
Metabolomic Comparison of Guava (Psidium guajava L.) Leaf Extracts Fermented by Limosilactobacillus fermentum and Lactiplantibacillus plantarum and Their Antioxidant and Antiglycation Activities

 , , and
, , and 
Abstract
1. Introduction
2. Materials and Methods
2.1. Chemicals and Reagents
2.2. Lactobacillus Strains and Culture Conditions
2.3. Sample Preparation and Extraction
2.4. GC-TOF-MS Analysis
2.5. UHPLC–Orbitrap–MS/MS Analysis
2.6. Data Processing and Statistical Analysis
2.7. Determination of the Antioxidant Activities and Antiglycation Activities
3. Results
3.1. Metabolic Profiling of the Guava Leaf Groups (0%, 0.5%, and 2%) Fermented by L. fermentum and L. plantarum
3.2. Metabolic Pathway Comparison of L. fermentum and L. plantarum Fermentation in the Guava Leaf Extract Groups (0%, 0.5%, and 2%)
3.3. Bioconversion of the Guava-Derived Metabolites Fermented by L. fermentum and L. plantarum
3.4. Bioactivity in the Groups Supplemented with the Guava Leaf Extracts (0%, 0.5%, and 2%) and Fermented by L. fermentum and L. plantarum
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Şanlier, N.; Gökcen, B.B.; Sezgin, A.C. Health Benefits of Fermented Foods. Crit. Rev. Food Sci. Nutr. 2019, 59, 506–527. [Google Scholar] [CrossRef]
- Wang, Y.; Wu, J.; Lv, M.; Shao, Z.; Hungwe, M.; Wang, J.; Bai, X.; Xie, J.; Wang, Y.; Geng, W. Metabolism Characteristics of Lactic Acid Bacteria and the Expanding Applications in Food Industry. Front. Bioeng. Biotechnol. 2021, 9, 612285. [Google Scholar] [CrossRef]
- Tang, H.; Huang, W.; Yao, Y.F. The metabolites of lactic acid bacteria: Classification, biosynthesis and modulation of gut microbiota. Microb. Cell 2023, 10, 49–62. [Google Scholar] [CrossRef]
- Leonard, W.; Zhang, P.; Ying, D.; Adhikari, B.; Fang, Z. Fermentation transforms the phenolic profiles and bioactivities of plant-based foods. Biotechnol. Adv. 2021, 49, 107763. [Google Scholar] [CrossRef] [PubMed]
- Abdul Hakim, B.N.; Xuan, N.J.; Oslan, S.N.H. A Comprehensive Review of Bioactive Compounds from Lactic Acid Bacteria: Potential Functions as Functional Food in Dietetics and the Food Industry. Foods 2023, 12, 2850. [Google Scholar] [CrossRef] [PubMed]
- Escrivá, L.; Manyes, L.; Vila-Donat, P.; Font, G.; Meca, G.; Lozano, M. Bioaccessibility and bioavailability of bioactive compounds from yellow mustard flour and milk whey fermented with lactic acid bacteria. Food Funct. 2021, 12, 11250–11261. [Google Scholar] [CrossRef] [PubMed]
- Gao, B.; Wang, J.; Wang, Y.; Xu, Z.; Li, B.; Meng, X.; Sun, X.; Zhu, J. Influence of fermentation by lactic acid bacteria and in vitro digestion on the biotransformations of blueberry juice phenolics. Food Control. 2022, 133, 108603. [Google Scholar] [CrossRef]
- Wu, Y.; Li, S.; Tao, Y.; Li, D.; Han, Y.; Show, P.L.; Wen, G.; Zhou, J. Fermentation of blueberry and blackberry juices using Lactobacillus plantarum, Streptococcus thermophilus and Bifidobacterium bifidum: Growth of probiotics, metabolism of phenolics, antioxidant capacity in vitro and sensory evaluation. Food Chem. 2021, 348, 129083. [Google Scholar] [CrossRef] [PubMed]
- Yang, J.S.; Cho, H.J.; Gil, M.C.; Kim, K.E. Anti-Inflammation and Anti-Melanogenic Effects of Maca Root Extracts Fermented Using Lactobacillus Strains. Antioxidants 2023, 12, 798. [Google Scholar] [CrossRef] [PubMed]
- Lee, M.E.; Yun, Y.R.; Choi, E.J.; Song, J.H.; Kang, J.Y.; Kim, D.U.; Lee, K.W.; Chang, J.Y. Anti-obesity effect of vegetable juice fermented with lactic acid bacteria isolated from kimchi in C57BL/6J mice and human mesenchymal stem cells. Food Funct. 2023, 14, 1349–1356. [Google Scholar] [CrossRef]
- Ai, X.; Wu, C.; Yin, T.; Zhur, O.; Liu, C.; Yan, X.; Yi, C.P.; Liu, D.; Xiao, L.; Li, W.; et al. Antidiabetic Function of Lactobacillus fermentum MF423-Fermented Rice Bran and Its Effect on Gut Microbiota Structure in Type 2 Diabetic Mice. Front. Microbiol. 2021, 12, 682290. [Google Scholar] [CrossRef]
- Algonaiman, R.; Alharbi, H.F.; Barakat, H. Antidiabetic and Hypolipidemic Efficiency of Lactobacillus plantarum Fermented Oat (Avena sativa) Extract in Streptozotocin-Induced Diabetes in Rats. Fermentation 2022, 8, 267. [Google Scholar] [CrossRef]
- Kim, S.H.; Singh, D.; Kim, S.A.; Kwak, M.J.; Cho, D.H.; Kim, J.W.; Roh, J.H.; Kim, W.G.; Han, N.S.; Lee, C.H. Strain-specific metabolomic diversity of Lactiplantibacillus plantarum under aerobic and anaerobic conditions. Food Microbiol. 2023, 116, 104364. [Google Scholar] [CrossRef] [PubMed]
- Yang, K.; Xu, M.; Zhong, F.; Zhu, J. Rapid differentiation of Lactobacillus species via metabolic profiling. J. Microbiol. Methods 2018, 154, 147–155. [Google Scholar] [CrossRef] [PubMed]
- Filannino, P.; Bai, Y.; Di Cagno, R.; Gobbetti, M.; Gänzle, M.G. Metabolism of phenolic compounds by Lactobacillus spp. during fermentation of cherry juice and broccoli puree. Food Microbiol. 2015, 46, 272–279. [Google Scholar] [CrossRef] [PubMed]
- Zhao, X.; Tang, F.; Cai, W.; Peng, B.; Zhang, P.; Shan, C. Effect of fermentation by lactic acid bacteria on the phenolic composition, antioxidant activity, and flavor substances of jujube–wolfberry composite juice. LWT 2023, 184, 114884. [Google Scholar] [CrossRef]
- Filannino, P.; Di Cagno, R.; Gobbetti, M. Metabolic and functional paths of lactic acid bacteria in plant foods: Get out of the labyrinth. Curr. Opin. Biotechnol. 2018, 49, 64–72. [Google Scholar] [CrossRef] [PubMed]
- Kumar, M.; Tomar, M.; Amarowicz, R.; Saurabh, V.; Sneha Nair, M.; Maheshwari, C.; Sasi, M.; Prajapati, U.; Hasan, M.; Singh, S.; et al. Guava (Psidium guajava L.) Leaves: Nutritional Composition, Phytochemical Profile, and Health-Promoting Bioactivities. Foods 2021, 10, 752. [Google Scholar] [CrossRef]
- Ashraf, A.; Sarfraz, R.A.; Rashid, M.A.; Mahmood, A.; Shahid, M.; Noor, N. Chemical composition, antioxidant, antitumor, anticancer and cytotoxic effects of Psidium guajava leaf extracts. Pharm. Biol. 2016, 54, 1971–1981. [Google Scholar] [CrossRef]
- Chen, H.Y.; Yen, G.C. Antioxidant activity and free radical-scavenging capacity of extracts from guava (Psidium guajava L.) leaves. Food Chem. 2007, 101, 686–694. [Google Scholar] [CrossRef]
- Wang, L.; Wei, W.; Tian, X.; Shi, K.; Wu, Z. Improving bioactivities of polyphenol extracts from Psidium guajava L. leaves through co-fermentation of Monascus anka GIM 3.592 and Saccharomyces cerevisiae GIM 2.139. Ind. Crops Prod. 2016, 94, 206–215. [Google Scholar] [CrossRef]
- Choi, S.-Y.; Hwang, J.-H.; Park, S.-Y.; Jin, Y.-J.; Ko, H.-C.; Moon, S.-W.; Kim, S.-J. Fermented guava leaf extract inhibits LPS-induced COX-2 and iNOS expression in Mouse macrophage cells by inhibition of transcription factor NF-κB. Phytother. Res. 2008, 22, 1030–1034. [Google Scholar] [CrossRef]
- Huang, Z.; Luo, Y.; Xia, X.; Wu, A.; Wu, Z. Bioaccessibility, safety, and antidiabetic effect of phenolic-rich extract from fermented Psidium guajava Linn. Leaves. J. Funct. Foods 2021, 86, 104723. [Google Scholar] [CrossRef]
- Wu, J.Q.; Kosten, T.R.; Zhang, X.Y. Free radicals, antioxidant defense systems, and schizophrenia. Prog. Neuropsychopharmacol. Biol. Psychiatry 2013, 46, 200–206. [Google Scholar] [CrossRef] [PubMed]
- Meenatchi, P.; Purushothaman, A.; Maneemegalai, S. Antioxidant, antiglycation and insulinotrophic properties of Coccinia grandis (L.) in vitro: Possible role in prevention of diabetic complications. J. Tradit. Complement. Med. 2017, 7, 54–64. [Google Scholar] [CrossRef] [PubMed]
- De Montijo-Prieto, S.; Razola-Díaz, M.D.C.; Barbieri, F.; Tabanelli, G.; Gardini, F.; Jiménez-Valera, M.; Ruiz-Bravo, A.; Verardo, V.; Gómez-Caravaca, A.M. Impact of Lactic Acid Bacteria Fermentation on Phenolic Compounds and Antioxidant Activity of Avocado Leaf Extracts. Antioxidants 2023, 12, 298. [Google Scholar] [CrossRef] [PubMed]
- Chan, M.Z.A.; Lu, Y.; Liu, S.Q. In Vitro Bioactivities of coffee brews fermented with the probiotics Lacticaseibacillus rhamnosus GG and Saccharomyces boulardii CNCM-I745. Food Res. Int. 2021, 149, 110693. [Google Scholar] [CrossRef] [PubMed]
- Lee, H.J.; Lee, S.M.; Kyung, S.Y.; Ryu, J.J.; Kang, S.H.; Park, M.S.; Lee, C.H. Metabolite Profiling and Anti-Aging Activity of Rice Koji Fermented with Aspergillus oryzae and Aspergillus cristatus: A Comparative Study. Metabolites 2021, 11, 524. [Google Scholar] [CrossRef] [PubMed]
- Kiho, T.; Kato, M.; Usui, S.; Hirano, K. Effect of buformin and metformin on formation of advanced glycation end products by methylglyoxal. Clin. Chim. Acta 2005, 358, 139–145. [Google Scholar] [CrossRef]
- Ortiz, M.E.; Bleckwedel, J.; Raya, R.R.; Mozzi, F. Biotechnological and in situ food production of polyols by lactic acid bacteria. Appl. Microbiol. Biotechnol. 2013, 97, 4713–4726. [Google Scholar] [CrossRef]
- Buron-Moles, G.; Chailyan, A.; Dolejs, I.; Forster, J.; Mikš, M.H. Uncovering carbohydrate metabolism through a genotype-phenotype association study of 56 lactic acid bacteria genomes. Appl. Microbiol. Biotechnol. 2019, 103, 3135–3152. [Google Scholar] [CrossRef]
- Rahman, M.M.; Hossain, K.M.; Rahman, S.M.M. Isolation, characterization, and properties study of probiotic lactic acid bacteria of selected yoghurt from Bangladesh. Afr. J. Microbiol. Res. 2016, 10, 23–31. [Google Scholar] [CrossRef]
- Verce, M.; De Vuyst, L.; Weckx, S. Comparative genomics of Lactobacillus fermentum suggests a free-living lifestyle of this lactic acid bacterial species. Food Microbiol. 2020, 89, 103448. [Google Scholar] [CrossRef]
- Parlindungan, E.; May, B.K.; Jones, O.A.H. Metabolic Insights into the Effects of Nutrient Stress on Lactobacillus plantarum B21. Front. Mol. Biosci. 2019, 6, 75. [Google Scholar] [CrossRef] [PubMed]
- Fernández, M.; Zúñiga, M. Amino Acid Catabolic Pathways of Lactic Acid Bacteria. Crit. Rev. Microbiol. 2006, 32, 155–183. [Google Scholar] [CrossRef]
- Valerio, F.; Lavermicocca, P.; Pascale, M.; Visconti, A. Production of phenyllactic acid by lactic acid bacteria: An approach to the selection of strains contributing to food quality and preservation. FEMS Microbiol. Lett. 2004, 233, 289–295. [Google Scholar] [CrossRef]
- Li, X.; Jiang, B.; Pan, B.; Mu, W.; Zhang, T. Purification and Partial Characterization of Lactobacillus Species SK007 Lactate Dehydrogenase (LDH) Catalyzing Phenylpyruvic Acid (PPA) Conversion into Phenyllactic Acid (PLA). J. Agric. Food Chem. 2008, 56, 2392–2399. [Google Scholar] [CrossRef] [PubMed]
- Méndez, M.G.G.; Martínez, T.K.M.; Valdés, J.A.A.; González, M.L.C.; Gallegos, A.C.F.; Sepúlveda, L. Application of Lactic Acid Bacteria in Fermentation Processes to Obtain Tannases Using Agro-Industrial Wastes. Fermentation 2021, 7, 48. [Google Scholar] [CrossRef]
- Adebo, O.A.; Njobeh, P.B.; Kayitesi, E. Fermentation by Lactobacillus fermentum strains (singly and in combination) enhances the properties of ting from two whole grain sorghum types. J. Cereal Sci. 2018, 82, 49–56. [Google Scholar] [CrossRef]
- Reverón, I.; de las Rivas, B.; Matesanz, R.; Muñoz, R.; López de Felipe, F. Molecular Adaptation of Lactobacillus plantarum WCFS1 to gallic acid revealed by genome-scale transcriptomic signature and physiological analysis. Microb. Cell Fact. 2015, 14, 160. [Google Scholar] [CrossRef]
- Xu, H.; Sun, J.; Zhao, Z.; Ma, X.; Li, C.; Liu, L.; Zhang, G. Lactobacillus plantarum ZLC-18 fermentation improve tyrosinase inhibition activity and antioxidant capacity in soybean hulls. Int. J. Food Sci. Technol. 2022, 57, 4518–4527. [Google Scholar] [CrossRef]
- Gupta, A.K.; Yumnam, M.; Medhi, M.; Koch, P.; Chakraborty, S.; Mishra, P. Isolation and characterization of naringinase enzyme and its application in debittering of Pomelo juice (Citrus grandis): A comparative study with macroporous resin. J. Food Process Preserv. 2021, 45, e15380. [Google Scholar] [CrossRef]
- Nguyen, N.A.; Cao, N.T.; Nguyen, T.H.H.; Ji, J.H.; Cha, G.S.; Kang, H.S.; Yun, C.H. Enzymatic Production of 3-OH Phlorizin, a Possible Bioactive Polyphenol from Apples, by Bacillus megaterium Cyp102a1 via Regioselective Hydroxylation. Antioxidants 2021, 10, 1327. [Google Scholar] [CrossRef]
- Michlmayr, H.; Kneifel, W. β-Glucosidase activities of lactic acid bacteria: Mechanisms, impact on fermented food and human health. FEMS Microbiol. Lett. 2014, 352, 1–10. [Google Scholar] [CrossRef]
- Guo, X.; Guo, A.; Li, E. Biotransformation of two citrus flavanones by lactic acid bacteria in chemical defined Medium. Bioprocess Biosyst. Eng. 2021, 44, 235–246. [Google Scholar] [CrossRef]
- Mueller, M.; Zartl, B.; Schleritzko, A.; Stenzl, M.; Viernstein, H.; Unger, F.M. Rhamnosidase activity of selected probiotics and their ability to hydrolyse flavonoid rhamnoglucosides. Bioprocess Biosyst. Eng. 2018, 41, 221–228. [Google Scholar] [CrossRef]
- Pietta, P.G. Flavonoids as Antioxidants. J. Nat. Prod. 2000, 63, 1035–1042. [Google Scholar] [CrossRef]
- Alqaraleh, M.; Kasabri, V.; Al-Majali, I.; Aljaafreh, A.; Al-Othman, N.; Khleifat, K.; Al-tawarah, N.M.; Qaralleh, H.; Khwaldeh, A.S.; Alalawi, S.; et al. Branched chain amino acids as in vitro and in vivo anti-oxidation compounds. Res. J. Pharm. Technol. 2021, 14, 3899–3904. [Google Scholar] [CrossRef]
- Kahkeshani, N.; Farzaei, F.; Fotouhi, M.; Alavi, S.S.; Bahramsoltani, R.; Naseri, R.; Momtaz, S.; Abbasabadi, Z.; Rahimi, R.; Farzaei, M.H.; et al. Pharmacological effects of gallic acid in health and disease: A mechanistic review. Iran. J. Basic. Med. Sci. 2019, 22, 225–237. [Google Scholar] [CrossRef]
- Khan, A.N.; Singh, R.; Bhattacharya, A.; Chakravarti, R.; Roy, S.; Ravichandiran, V.; Ghosh, D. A Short Review on Glucogallin and Its Pharmacological Activities. Mini-Rev. Med. Chem. 2022, 22, 2820–2830. [Google Scholar] [CrossRef] [PubMed]
- Seckin, B.; Sekmen, A.H.; Türkan, I. An Enhancing Effect of Exogenous Mannitol on the Antioxidant Enzyme Activities in Roots of Wheat under Salt Stress. J. Plant Growth Regul. 2009, 28, 12–20. [Google Scholar] [CrossRef]
- Shao, X.; Chen, H.; Zhu, Y.; Sedighi, R.; Ho, C.T.; Sang, S. Essential Structural Requirements and Additive Effects for Flavonoids to Scavenge Methylglyoxal. J. Agric. Food Chem. 2014, 62, 3202–3210. [Google Scholar] [CrossRef] [PubMed]
- Alhadid, A.; Bustanji, Y.; Harb, A.; Al-Hiari, Y.; Abdalla, S. Vanillic acid Inhibited the Induced Glycation Using in Vitro and in Vivo Models. Evid.-Based Complement. Altern. Med. 2022, 2022, 7119256. [Google Scholar] [CrossRef] [PubMed]
- Zhou, Q.; Gong, J.; Wang, M. Phloretin and its methylglyoxal adduct: Implications against advanced glycation end products-induced inflammation in endothelial cells. Food Chem. Toxicol. 2019, 129, 291–300. [Google Scholar] [CrossRef] [PubMed]
- Teng, J.; Li, Y.; Yu, W.; Zhao, Y.; Hu, X.; Tao, N.P.; Wang, M. Naringenin, a common flavanone, inhibits the formation of AGEs in bread and attenuates AGEs-induced oxidative stress and inflammation in RAW264.7 cells. Food Chem. 2018, 269, 35–42. [Google Scholar] [CrossRef]
- Beloborodova, N.; Bairamov, I.; Olenin, A.; Shubina, V.; Teplova, V.; Fedotcheva, N. Effect of phenolic acids of microbial origin on production of reactive oxygen species in mitochondria and neutrophils. J. Biomed. Sci. 2012, 19, 89. [Google Scholar] [CrossRef]
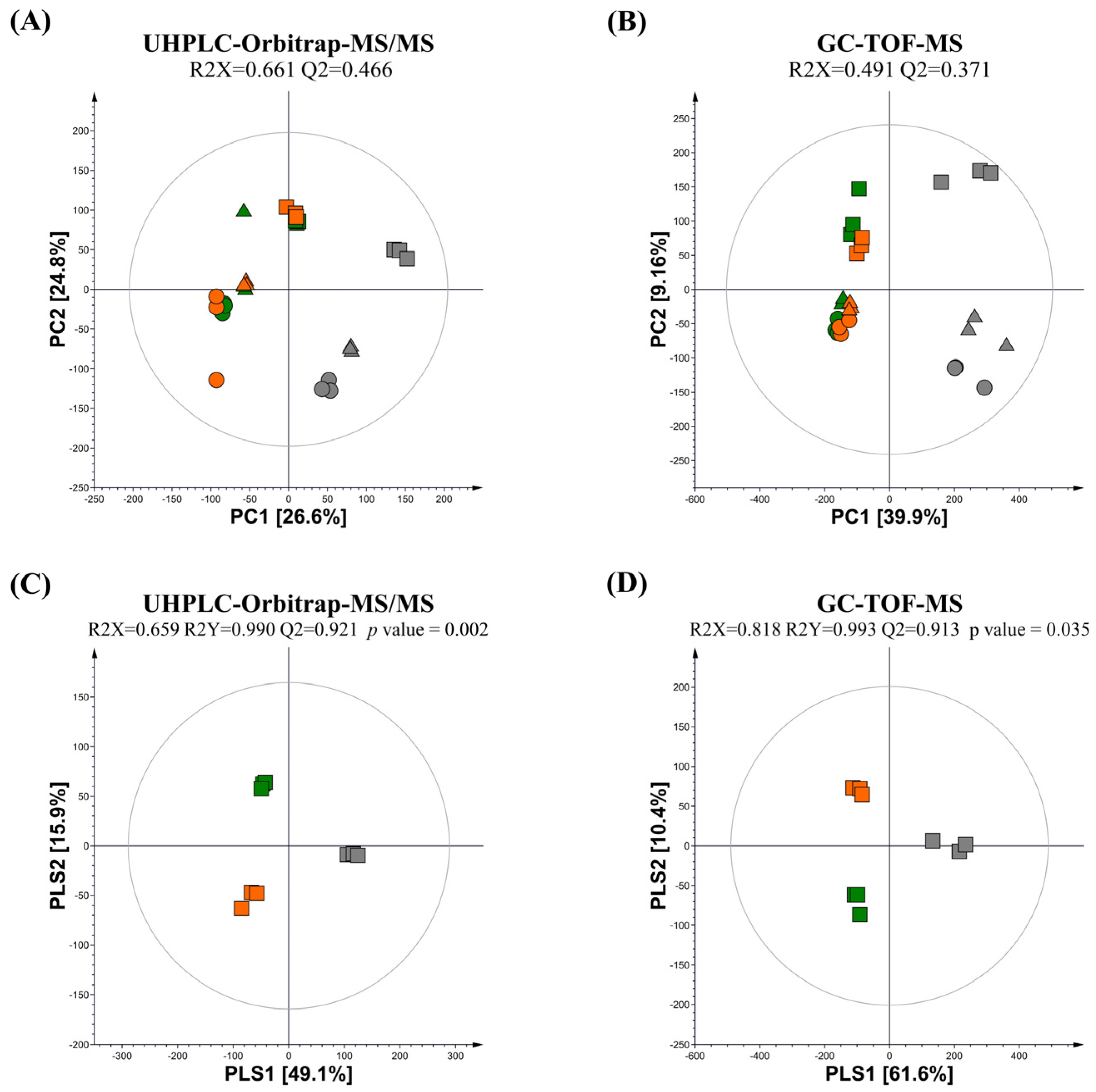

 : non-fermented (NFG);
: non-fermented (NFG);  : fermented by L. fermentum (LFG);
: fermented by L. fermentum (LFG);  : fermented by L. plantarum (LPG)).
: non-fermented (NFG); : fermented by L. fermentum (LFG); : fermented by L. plantarum (LPG)).
: fermented by L. plantarum (LPG)).
: non-fermented (NFG); : fermented by L. fermentum (LFG); : fermented by L. plantarum (LPG)).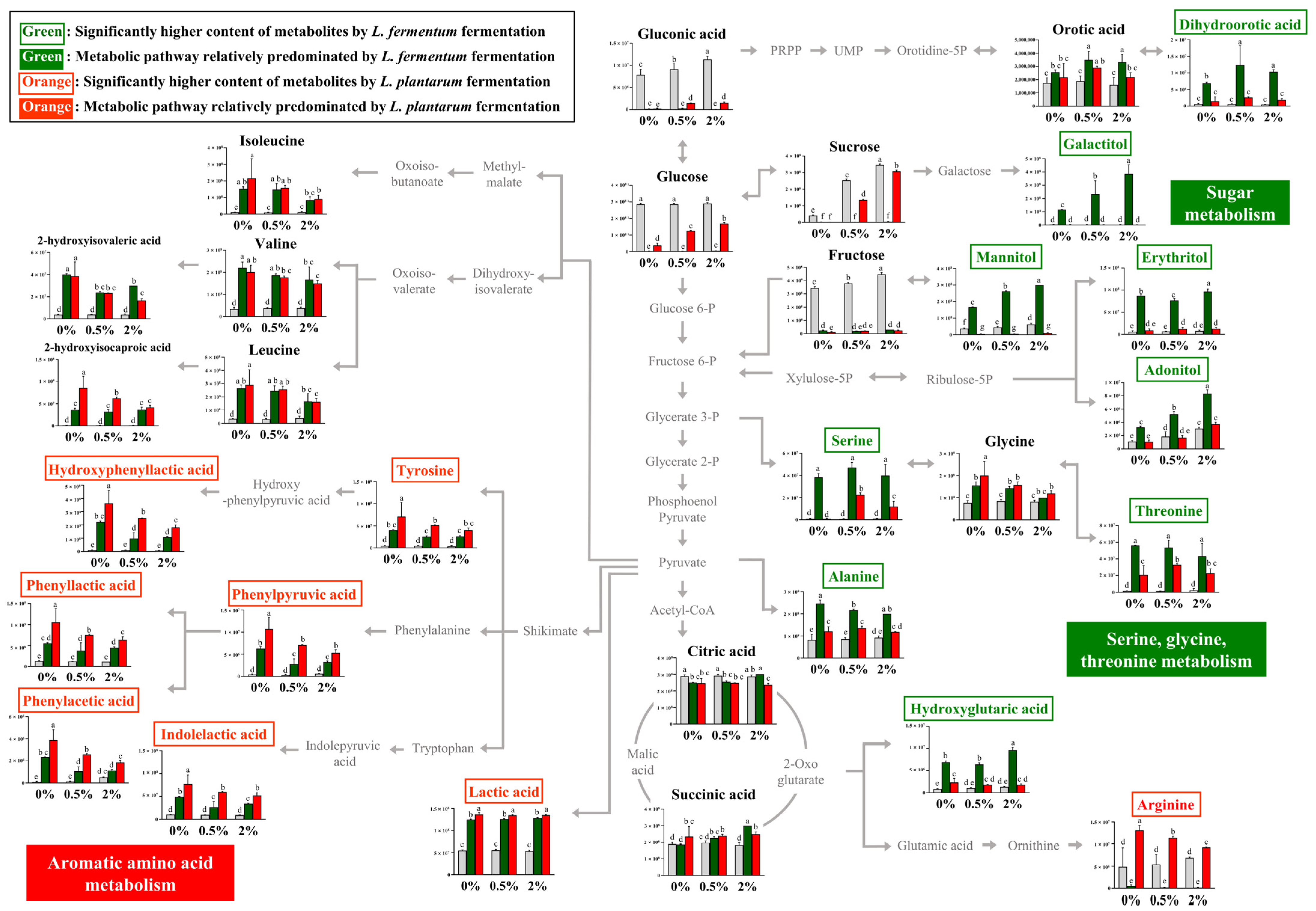 : non-fermented (NFG); : fermented by L. fermentum (LFG); : fermented by L. plantarum (LPG)).
: non-fermented (NFG); : fermented by L. fermentum (LFG); : fermented by L. plantarum (LPG)).
: non-fermented (NFG); : fermented by L. fermentum (LFG); : fermented by L. plantarum (LPG)).
: non-fermented (NFG); : fermented by L. fermentum (LFG); : fermented by L. plantarum (LPG)).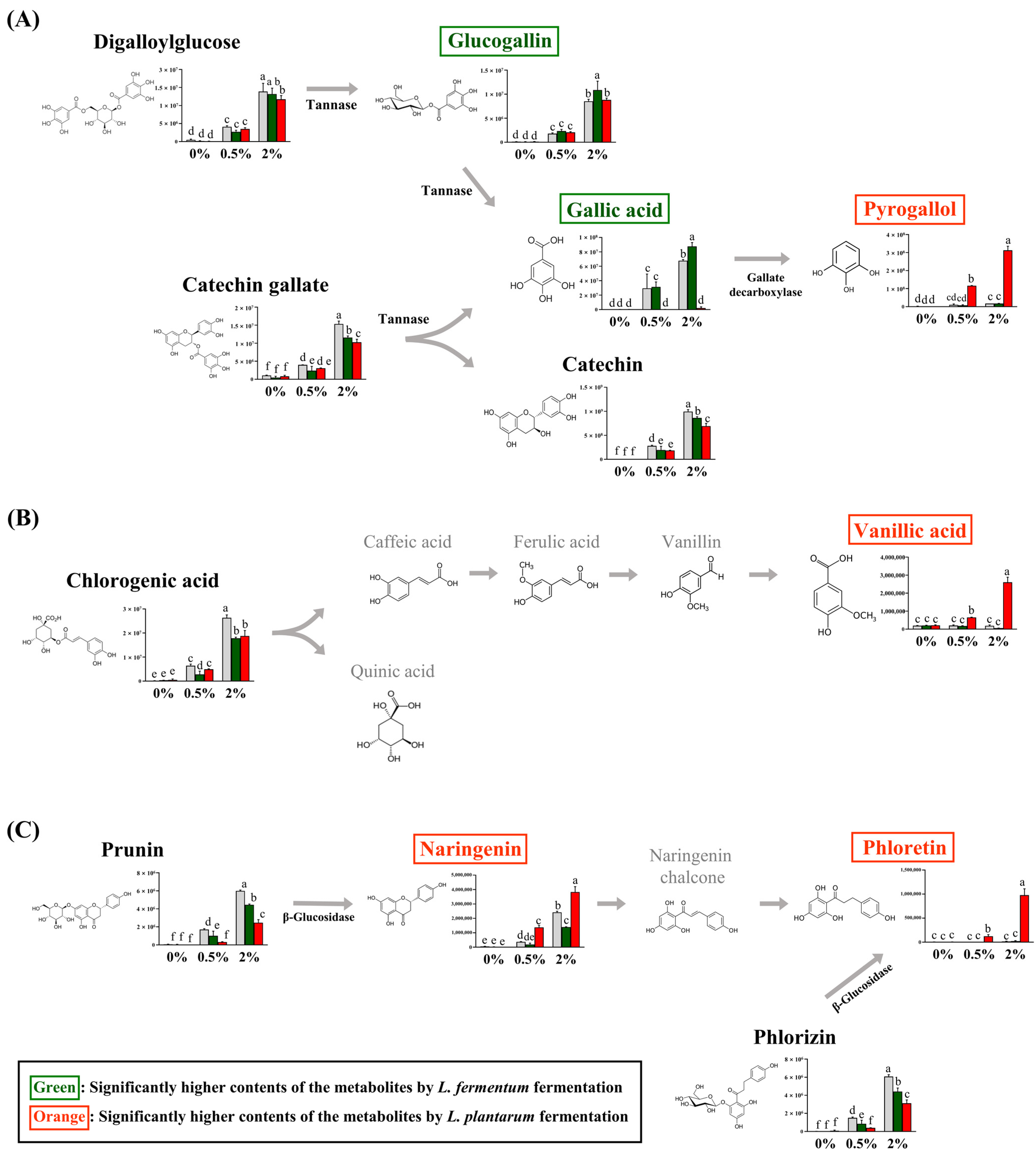 : non-fermented (NFG); : fermented by L. fermentum (LFG); : fermented by L. plantarum (LPG)).
: non-fermented (NFG); : fermented by L. fermentum (LFG); : fermented by L. plantarum (LPG)).
: non-fermented (NFG); : fermented by L. fermentum (LFG); : fermented by L. plantarum (LPG)).
: non-fermented (NFG); : fermented by L. fermentum (LFG); : fermented by L. plantarum (LPG)).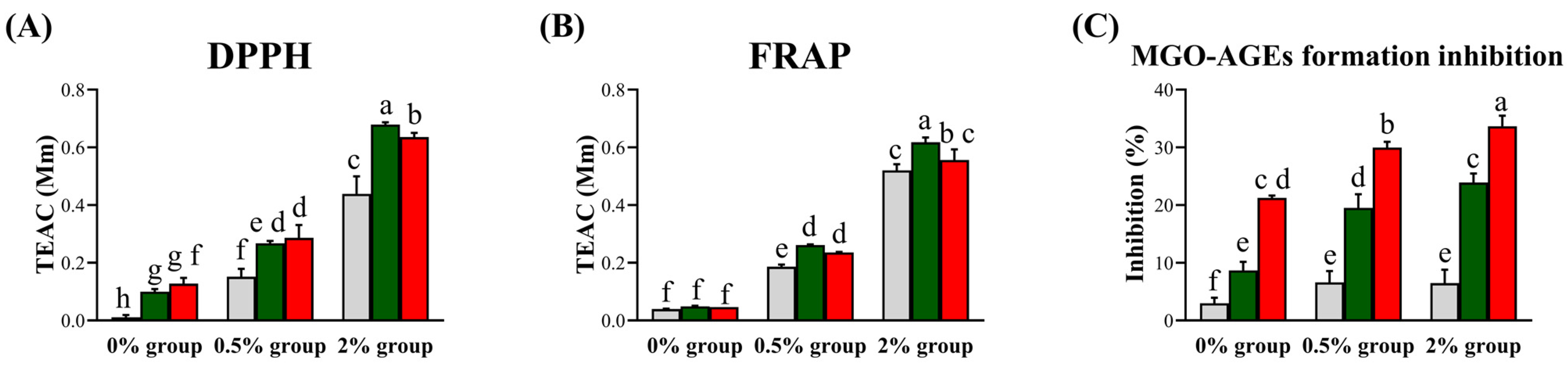
 , LFG-specific compounds;
, LFG-specific compounds;  , LPG-specific compounds;
, LPG-specific compounds;  , non-specific compound).
, LFG-specific compounds; , LPG-specific compounds; , non-specific compound).
, non-specific compound).
, LFG-specific compounds; , LPG-specific compounds; , non-specific compound).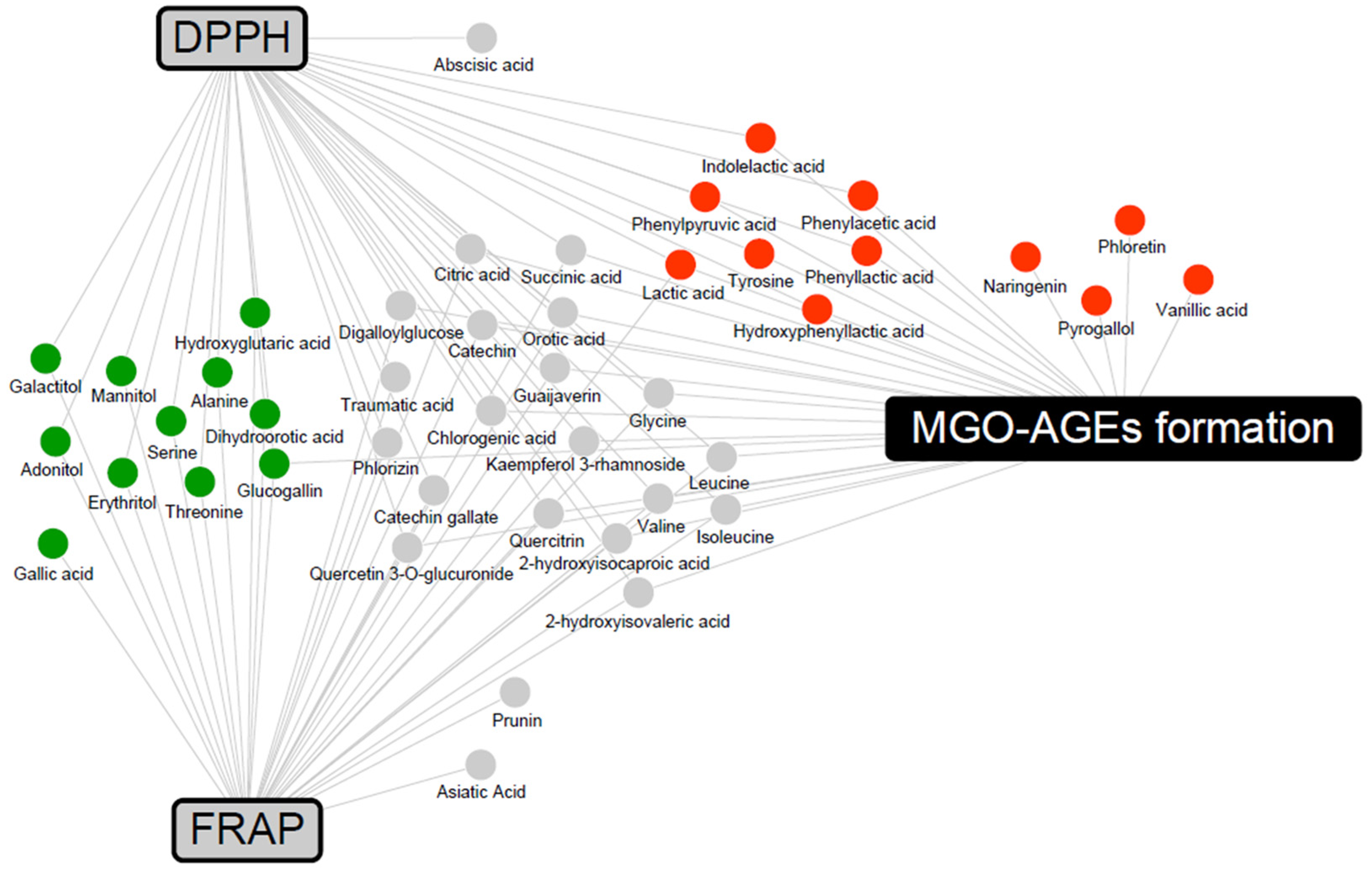

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Substrate | Inoculation | Abb a | Symbol |
|---|---|---|---|
| MRS | - | NFG-0 |  |
| (0% group) | Limosilactobacillus fermentum KCTC15072BP | LFG-0 |  |
| Lactiplantibacillus plantarum KGMB00831 | LPG-0 |  | |
| MRS with 0.5% (w/v) guava leaf extract | - | NFG-0.5 |  |
| (0.5% group) | Limosilactobacillus fermentum KCTC15072BP | LFG-0.5 |  |
| Lactiplantibacillus plantarum KGMB00831 | LPG-0.5 |  | |
| MRS with 2% (w/v) guava leaf extract | - | NFG-2 |  |
| (2% group) | Limosilactobacillus fermentum KCTC15072BP | LPG-2 |  |
| Lactiplantibacillus plantarum KGMB00831 | LFG-2 |  |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Jun, B.-G.; Kim, S.-H.; Kim, S.-H.; Hong, S.-M.; Lee, H.; Lim, Y.; Kim, S.-Y.; Lee, C.-H. Metabolomic Comparison of Guava (Psidium guajava L.) Leaf Extracts Fermented by Limosilactobacillus fermentum and Lactiplantibacillus plantarum and Their Antioxidant and Antiglycation Activities. Nutrients 2024, 16, 841. https://doi.org/10.3390/nu16060841
Jun B-G, Kim S-H, Kim S-H, Hong S-M, Lee H, Lim Y, Kim S-Y, Lee C-H. Metabolomic Comparison of Guava (Psidium guajava L.) Leaf Extracts Fermented by Limosilactobacillus fermentum and Lactiplantibacillus plantarum and Their Antioxidant and Antiglycation Activities. Nutrients. 2024; 16(6):841. https://doi.org/10.3390/nu16060841
Chicago/Turabian StyleJun, Bo-Gyu, Su-Hyun Kim, Seon-Hyeok Kim, Seong-Min Hong, Heaji Lee, Yunsook Lim, Sun-Yeou Kim, and Choong-Hwan Lee. 2024. "Metabolomic Comparison of Guava (Psidium guajava L.) Leaf Extracts Fermented by Limosilactobacillus fermentum and Lactiplantibacillus plantarum and Their Antioxidant and Antiglycation Activities" Nutrients 16, no. 6: 841. https://doi.org/10.3390/nu16060841
APA StyleJun, B.-G., Kim, S.-H., Kim, S.-H., Hong, S.-M., Lee, H., Lim, Y., Kim, S.-Y., & Lee, C.-H. (2024). Metabolomic Comparison of Guava (Psidium guajava L.) Leaf Extracts Fermented by Limosilactobacillus fermentum and Lactiplantibacillus plantarum and Their Antioxidant and Antiglycation Activities. Nutrients, 16(6), 841. https://doi.org/10.3390/nu16060841