
The Cytotoxic Effect of Curcumin in Rhabdomyosarcoma Is Associated with the Modulation of AMPK, AKT/mTOR, STAT, and p53 Signaling

 ,
,  , , , ,
, , , ,  ,
,  ,
,  and
and
Abstract
1. Introduction
2. Materials and Methods
2.1. Cell Lines and Reagents
2.2. Cell Proliferation Assay
2.3. Trypan Blue Dye Exclusion Assay
2.4. Cell Cycle Analysis
2.5. Apoptosis Assay
2.6. Colony Formation Assay
2.7. Antibody Arrays
2.8. Statistical Analysis
3. Results
3.1. Curcumin Rapidly Kills Rhabdomyosarcoma Cells
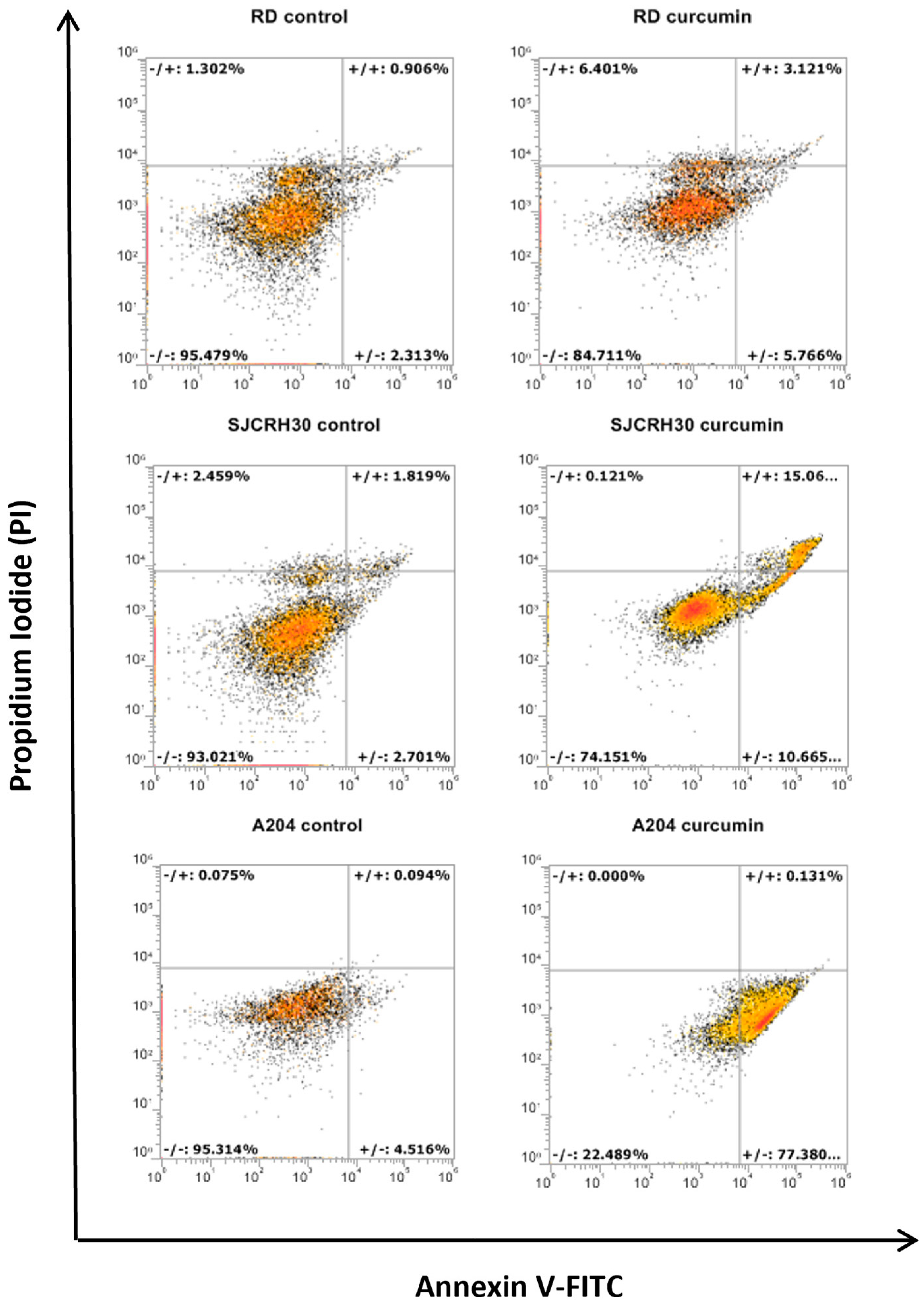
3.2. Curcumin Induces Apoptotic Cell Death in Rhabdomyosarcoma, Regardless of the Status of p53
3.3. Curcumin Inhibits Cell Migration and Colony Forming Ability
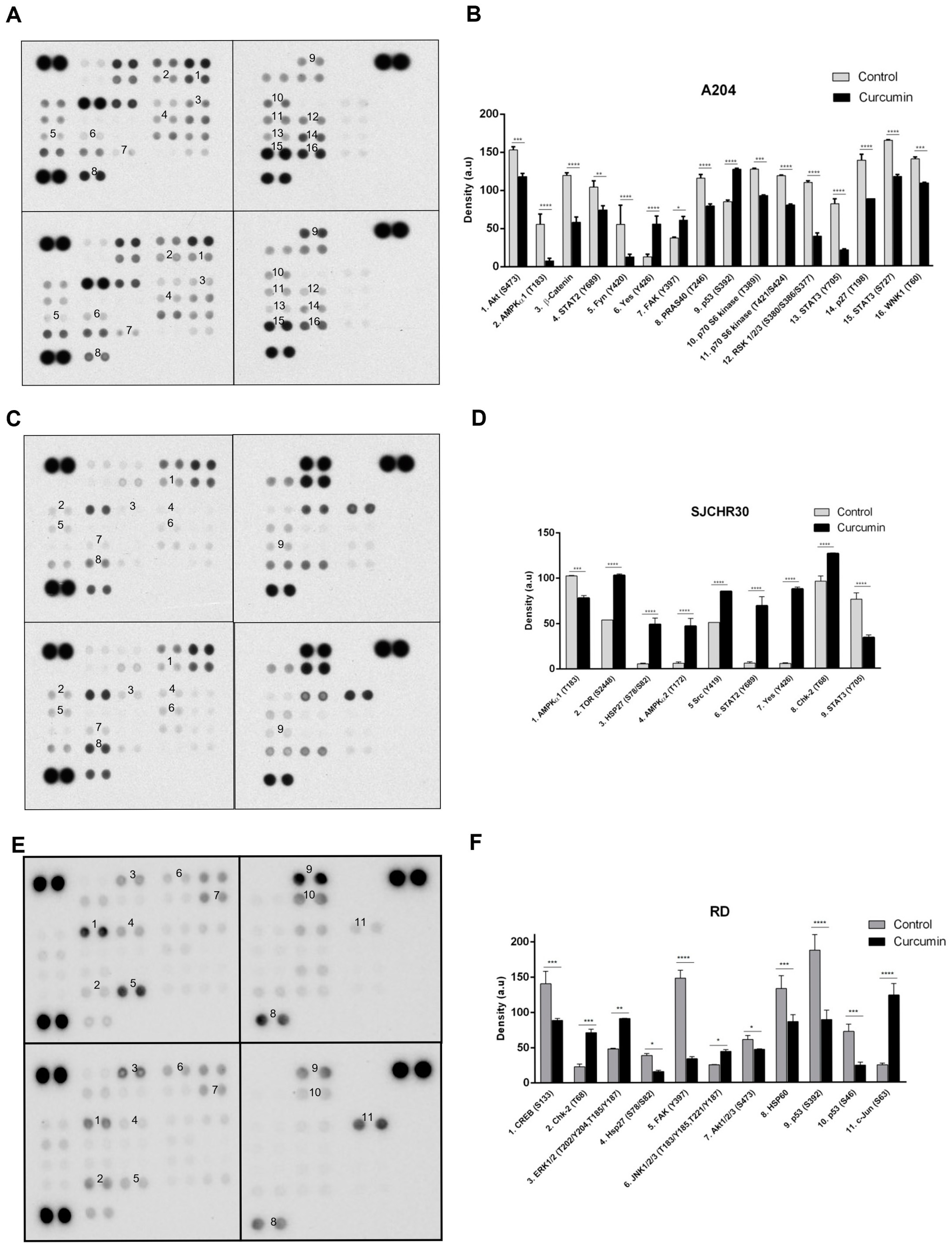
3.4. Curcumin Treatment Alters Diverse Signal Transduction Pathways Important to Tumor Growth
4. Discussion
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Skapek, S.X.; Ferrari, A.; Gupta, A.A.; Lupo, P.J.; Butler, E.; Shipley, J.; Barr, F.G.; Hawkins, D.S. Rhabdomyosarcoma. Nat. Rev. Dis. Prim. 2019, 5, 1. [Google Scholar] [CrossRef] [PubMed]
- Gallego Melcon, S.; Sanchez de Toledo Codina, J. Molecular biology of rhabdomyosarcoma. Clin. Transl. Oncol. 2007, 9, 415–419. [Google Scholar] [CrossRef]
- Scrable, H.; Witte, D.; Shimada, H.; Seemayer, T.; Sheng, W.W.; Soukup, S.; Koufos, A.; Houghton, P.; Lampkin, B.; Cavenee, W. Molecular differential pathology of rhabdomyosarcoma. Genes Chromosomes Cancer 1989, 1, 23–35. [Google Scholar] [CrossRef] [PubMed]
- Nguyen, T.H.; Barr, F.G. Therapeutic Approaches Targeting PAX3-FOXO1 and Its Regulatory and Transcriptional Pathways in Rhabdomyosarcoma. Molecules 2018, 23, 2798. [Google Scholar] [CrossRef] [PubMed]
- Davis, R.J.; Barr, F.G. Fusion genes resulting from alternative chromosomal translocations are overexpressed by gene-specific mechanisms in alveolar rhabdomyosarcoma. Proc. Natl. Acad. Sci. USA 1997, 94, 8047–8051. [Google Scholar] [CrossRef]
- Soleimani, V.D.; Rudnicki, M.A. New insights into the origin and the genetic basis of rhabdomyosarcomas. Cancer Cell 2011, 19, 157–159. [Google Scholar] [CrossRef]
- Burattini, S.; Battistelli, M.; Codenotti, S.; Falcieri, E.; Fanzani, A.; Salucci, S. Melatonin action in tumor skeletal muscle cells: An ultrastructural study. Acta Histochem. 2016, 118, 278–285. [Google Scholar] [CrossRef]
- Codenotti, S.; Battistelli, M.; Burattini, S.; Salucci, S.; Falcieri, E.; Rezzani, R.; Faggi, F.; Colombi, M.; Monti, E.; Fanzani, A. Melatonin decreases cell proliferation, impairs myogenic differentiation and triggers apoptotic cell death in rhabdomyosarcoma cell lines. Oncol. Rep. 2015, 34, 279–287. [Google Scholar] [CrossRef]
- Aslam, M.I.; Hettmer, S.; Abraham, J.; Latocha, D.; Soundararajan, A.; Huang, E.T.; Goros, M.W.; Michalek, J.E.; Wang, S.; Mansoor, A.; et al. Dynamic and nuclear expression of PDGFRalpha and IGF-1R in alveolar Rhabdomyosarcoma. Mol. Cancer Res. 2013, 11, 1303–1313. [Google Scholar] [CrossRef]
- McKinnon, T.; Venier, R.; Yohe, M.; Sindiri, S.; Gryder, B.E.; Shern, J.F.; Kabaroff, L.; Dickson, B.; Schleicher, K.; Chouinard-Pelletier, G.; et al. Functional screening of FGFR4-driven tumorigenesis identifies PI3K/mTOR inhibition as a therapeutic strategy in rhabdomyosarcoma. Oncogene 2018, 37, 2630–2644. [Google Scholar] [CrossRef]
- Chelsky, Z.L.; Paulson, V.A.; Chen, E.Y. Molecular analysis of 10 pleomorphic rhabdomyosarcomas reveals potential prognostic markers and druggable targets. Genes Chromosomes Cancer 2022, 61, 138–147. [Google Scholar] [CrossRef] [PubMed]
- Crose, L.E.; Linardic, C.M. Receptor tyrosine kinases as therapeutic targets in rhabdomyosarcoma. Sarcoma 2011, 2011, 756982. [Google Scholar] [CrossRef] [PubMed]
- Ramadan, F.; Fahs, A.; Ghayad, S.E.; Saab, R. Signaling pathways in Rhabdomyosarcoma invasion and metastasis. Cancer Metastasis Rev. 2020, 39, 287–301. [Google Scholar] [CrossRef] [PubMed]
- Crist, W.; Gehan, E.A.; Ragab, A.H.; Dickman, P.S.; Donaldson, S.S.; Fryer, C.; Hammond, D.; Hays, D.M.; Herrmann, J.; Heyn, R.; et al. The Third Intergroup Rhabdomyosarcoma Study. J. Clin. Oncol. 1995, 13, 610–630. [Google Scholar] [CrossRef]
- Breneman, J.C.; Lyden, E.; Pappo, A.S.; Link, M.P.; Anderson, J.R.; Parham, D.M.; Qualman, S.J.; Wharam, M.D.; Donaldson, S.S.; Maurer, H.M.; et al. Prognostic factors and clinical outcomes in children and adolescents with metastatic rhabdomyosarcoma--a report from the Intergroup Rhabdomyosarcoma Study IV. J. Clin. Oncol. 2003, 21, 78–84. [Google Scholar] [CrossRef] [PubMed]
- Urosevic, M.; Nikolic, L.; Gajic, I.; Nikolic, V.; Dinic, A.; Miljkovic, V. Curcumin: Biological Activities and Modern Pharmaceutical Forms. Antibiotics 2022, 11, 135. [Google Scholar] [CrossRef]
- Giordano, A.; Tommonaro, G. Curcumin and Cancer. Nutrients 2019, 11, 2376. [Google Scholar] [CrossRef]
- Toden, S.; Goel, A. The Holy Grail of Curcumin and its Efficacy in Various Diseases: Is Bioavailability Truly a Big Concern? J. Restor. Med. 2017, 6, 27–36. [Google Scholar] [CrossRef]
- Beevers, C.S.; Li, F.; Liu, L.; Huang, S. Curcumin inhibits the mammalian target of rapamycin-mediated signaling pathways in cancer cells. Int. J. Cancer 2006, 119, 757–764. [Google Scholar] [CrossRef]
- Hinson, A.R.; Jones, R.; Crose, L.E.; Belyea, B.C.; Barr, F.G.; Linardic, C.M. Human rhabdomyosarcoma cell lines for rhabdomyosarcoma research: Utility and pitfalls. Front. Oncol. 2013, 3, 183. [Google Scholar] [CrossRef]
- Loupe, J.M.; Miller, P.J.; Bonner, B.P.; Maggi, E.C.; Vijayaraghavan, J.; Crabtree, J.S.; Taylor, C.M.; Zabaleta, J.; Hollenbach, A.D. Comparative transcriptomic analysis reveals the oncogenic fusion protein PAX3-FOXO1 globally alters mRNA and miRNA to enhance myoblast invasion. Oncogenesis 2016, 5, e246. [Google Scholar] [CrossRef] [PubMed]
- De Pitta, C.; Tombolan, L.; Albiero, G.; Sartori, F.; Romualdi, C.; Jurman, G.; Carli, M.; Furlanello, C.; Lanfranchi, G.; Rosolen, A. Gene expression profiling identifies potential relevant genes in alveolar rhabdomyosarcoma pathogenesis and discriminates PAX3-FKHR positive and negative tumors. Int. J. Cancer 2006, 118, 2772–2781. [Google Scholar] [CrossRef] [PubMed]
- Neale, G.; Su, X.; Morton, C.L.; Phelps, D.; Gorlick, R.; Lock, R.B.; Reynolds, C.P.; Maris, J.M.; Friedman, H.S.; Dome, J.; et al. Molecular characterization of the pediatric preclinical testing panel. Clin. Cancer Res. 2008, 14, 4572–4583. [Google Scholar] [CrossRef] [PubMed]
- Sakaguchi, K.; Sakamoto, H.; Lewis, M.S.; Anderson, C.W.; Erickson, J.W.; Appella, E.; Xie, D. Phosphorylation of serine 392 stabilizes the tetramer formation of tumor suppressor protein p53. Biochemistry 1997, 36, 10117–10124. [Google Scholar] [CrossRef]
- Cuddihy, A.R.; Wong, A.H.; Tam, N.W.; Li, S.; Koromilas, A.E. The double-stranded RNA activated protein kinase PKR physically associates with the tumor suppressor p53 protein and phosphorylates human p53 on serine 392 in vitro. Oncogene 1999, 18, 2690–2702. [Google Scholar] [CrossRef] [PubMed]
- Cuddihy, A.R.; Li, S.; Tam, N.W.; Wong, A.H.; Taya, Y.; Abraham, N.; Bell, J.C.; Koromilas, A.E. Double-stranded-RNA-activated protein kinase PKR enhances transcriptional activation by tumor suppressor p53. Mol. Cell. Biol. 1999, 19, 2475–2484. [Google Scholar] [CrossRef] [PubMed]
- Cox, M.L.; Meek, D.W. Phosphorylation of serine 392 in p53 is a common and integral event during p53 induction by diverse stimuli. Cell. Signal. 2010, 22, 564–571. [Google Scholar] [CrossRef]
- Dai, Z.; Li, G.; Chen, Q.; Yang, X. Ser392 phosphorylation modulated a switch between p53 and transcriptional condensates. Biochim. Biophys. Acta-Gene Regul. Mech. 2022, 1865, 194827. [Google Scholar] [CrossRef]
- Parrales, A.; Iwakuma, T. Targeting Oncogenic Mutant p53 for Cancer Therapy. Front. Oncol. 2015, 5, 288. [Google Scholar] [CrossRef]
- Hu, J.; Cao, J.; Topatana, W.; Juengpanich, S.; Li, S.; Zhang, B.; Shen, J.; Cai, L.; Cai, X.; Chen, M. Targeting mutant p53 for cancer therapy: Direct and indirect strategies. J. Hematol. Oncol. 2021, 14, 157. [Google Scholar] [CrossRef]
- Gao, G.; Fernandez, C.S.; Stapleton, D.; Auster, A.S.; Widmer, J.; Dyck, J.R.; Kemp, B.E.; Witters, L.A. Non-catalytic beta- and gamma-subunit isoforms of the 5’-AMP-activated protein kinase. J. Biol. Chem. 1996, 271, 8675–8681. [Google Scholar] [CrossRef] [PubMed]
- Dyck, J.R.; Gao, G.; Widmer, J.; Stapleton, D.; Fernandez, C.S.; Kemp, B.E.; Witters, L.A. Regulation of 5’-AMP-activated protein kinase activity by the noncatalytic beta and gamma subunits. J. Biol. Chem. 1996, 271, 17798–17803. [Google Scholar] [CrossRef]
- Woods, A.; Cheung, P.C.; Smith, F.C.; Davison, M.D.; Scott, J.; Beri, R.K.; Carling, D. Characterization of AMP-activated protein kinase beta and gamma subunits. Assembly of the heterotrimeric complex in vitro. J. Biol. Chem. 1996, 271, 10282–10290. [Google Scholar] [CrossRef] [PubMed]
- Hardie, D.G.; Hawley, S.A.; Scott, J.W. AMP-activated protein kinase--development of the energy sensor concept. J. Physiol. 2006, 574, 7–15. [Google Scholar] [CrossRef] [PubMed]
- Lage, R.; Dieguez, C.; Vidal-Puig, A.; Lopez, M. AMPK: A metabolic gauge regulating whole-body energy homeostasis. Trends Mol. Med. 2008, 14, 539–549. [Google Scholar] [CrossRef] [PubMed]
- Stapleton, D.; Mitchelhill, K.I.; Gao, G.; Widmer, J.; Michell, B.J.; Teh, T.; House, C.M.; Fernandez, C.S.; Cox, T.; Witters, L.A.; et al. Mammalian AMP-activated protein kinase subfamily. J. Biol. Chem. 1996, 271, 611–614. [Google Scholar] [CrossRef]
- Wang, B.; Nie, J.; Wu, L.; Hu, Y.; Wen, Z.; Dong, L.; Zou, M.H.; Chen, C.; Wang, D.W. AMPKalpha2 Protects Against the Development of Heart Failure by Enhancing Mitophagy via PINK1 Phosphorylation. Circ. Res. 2018, 122, 712–729. [Google Scholar] [CrossRef]
- Philips, R.L.; Wang, Y.; Cheon, H.; Kanno, Y.; Gadina, M.; Sartorelli, V.; Horvath, C.M.; Darnell, J.E., Jr.; Stark, G.R.; O’Shea, J.J. The JAK-STAT pathway at 30: Much learned, much more to do. Cell 2022, 185, 3857–3876. [Google Scholar] [CrossRef]
- Igelmann, S.; Neubauer, H.A.; Ferbeyre, G. STAT3 and STAT5 Activation in Solid Cancers. Cancers 2019, 11, 1428. [Google Scholar] [CrossRef]
- Shen, S.; Niso-Santano, M.; Adjemian, S.; Takehara, T.; Malik, S.A.; Minoux, H.; Souquere, S.; Marino, G.; Lachkar, S.; Senovilla, L.; et al. Cytoplasmic STAT3 represses autophagy by inhibiting PKR activity. Mol. Cell. 2012, 48, 667–680. [Google Scholar] [CrossRef]
- Fink, K.; Grandvaux, N. STAT2 and IRF9: Beyond ISGF3. JAKSTAT 2013, 2, e27521. [Google Scholar] [CrossRef] [PubMed]
- Guenther, M.K.; Graab, U.; Fulda, S. Synthetic lethal interaction between PI3K/Akt/mTOR and Ras/MEK/ERK pathway inhibition in rhabdomyosarcoma. Cancer Lett. 2013, 337, 200–209. [Google Scholar] [CrossRef] [PubMed]
- Renshaw, J.; Taylor, K.R.; Bishop, R.; Valenti, M.; De Haven Brandon, A.; Gowan, S.; Eccles, S.A.; Ruddle, R.R.; Johnson, L.D.; Raynaud, F.I.; et al. Dual blockade of the PI3K/AKT/mTOR (AZD8055) and RAS/MEK/ERK (AZD6244) pathways synergistically inhibits rhabdomyosarcoma cell growth in vitro and in vivo. Clin. Cancer Res. 2013, 19, 5940–5951. [Google Scholar] [CrossRef]
- Preuss, E.; Hugle, M.; Reimann, R.; Schlecht, M.; Fulda, S. Pan-mammalian target of rapamycin (mTOR) inhibitor AZD8055 primes rhabdomyosarcoma cells for ABT-737-induced apoptosis by down-regulating Mcl-1 protein. J. Biol. Chem. 2013, 288, 35287–35296. [Google Scholar] [CrossRef]
- Slotkin, E.K.; Patwardhan, P.P.; Vasudeva, S.D.; de Stanchina, E.; Tap, W.D.; Schwartz, G.K. MLN0128, an ATP-competitive mTOR kinase inhibitor with potent in vitro and in vivo antitumor activity, as potential therapy for bone and soft-tissue sarcoma. Mol. Cancer Ther. 2015, 14, 395–406. [Google Scholar] [CrossRef]
- Graab, U.; Hahn, H.; Fulda, S. Identification of a novel synthetic lethality of combined inhibition of hedgehog and PI3K signaling in rhabdomyosarcoma. Oncotarget 2015, 6, 8722–8735. [Google Scholar] [CrossRef] [PubMed]
- Anderson, J.L.; Park, A.; Akiyama, R.; Tap, W.D.; Denny, C.T.; Federman, N. Evaluation of In Vitro Activity of the Class I PI3K Inhibitor Buparlisib (BKM120) in Pediatric Bone and Soft Tissue Sarcomas. PLoS ONE 2015, 10, e0133610. [Google Scholar] [CrossRef]
- Becher, O.J.; Gilheeney, S.W.; Khakoo, Y.; Lyden, D.C.; Haque, S.; De Braganca, K.C.; Kolesar, J.M.; Huse, J.T.; Modak, S.; Wexler, L.H.; et al. A phase I study of perifosine with temsirolimus for recurrent pediatric solid tumors. Pediatr. Blood Cancer 2017, 64, e26409. [Google Scholar] [CrossRef]
- Piazzi, M.; Bavelloni, A.; Cenni, V.; Salucci, S.; Bartoletti Stella, A.; Tomassini, E.; Scotlandi, K.; Blalock, W.L.; Faenza, I. Combined Treatment with PI3K Inhibitors BYL-719 and CAL-101 Is a Promising Antiproliferative Strategy in Human Rhabdomyosarcoma Cells. Molecules 2022, 27, 2742. [Google Scholar] [CrossRef]
- Wei, C.C.; Ball, S.; Lin, L.; Liu, A.; Fuchs, J.R.; Li, P.K.; Li, C.; Lin, J. Two small molecule compounds, LLL12 and FLLL32, exhibit potent inhibitory activity on STAT3 in human rhabdomyosarcoma cells. Int. J. Oncol. 2011, 38, 279–285. [Google Scholar]
- Sohn, S.I.; Priya, A.; Balasubramaniam, B.; Muthuramalingam, P.; Sivasankar, C.; Selvaraj, A.; Valliammai, A.; Jothi, R.; Pandian, S. Biomedical Applications and Bioavailability of Curcumin-An Updated Overview. Pharmaceutics 2021, 13, 2102. [Google Scholar] [CrossRef] [PubMed]
- Sims, T.L.; McGee, M.; Williams, R.F.; Myers, A.L.; Tracey, L.; Hamner, J.B.; Ng, C.; Wu, J.; Gaber, M.W.; McCarville, B.; et al. IFN-beta restricts tumor growth and sensitizes alveolar rhabdomyosarcoma to ionizing radiation. Mol. Cancer Ther. 2010, 9, 761–771. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Protein (Modification) | A204 | RH30 | RD | Result/Process |
|---|---|---|---|---|
| AMPKα1 (T183) | ↓ | ↓ | (Activation, intracellular localization) The catalytic subunit of the cellular energy sensing protein kinase AMP-activated protein kinase (AMPK). Inhibits energy consuming cellular processes (transcription, translation, ribosome biogenesis, etc.) under conditions of low ATP. Enhances glucose uptake and regulates insulin signaling and glycolysis. Enhances transcriptional activity of transcription factors involved in regulating energy metabolism. Directly inhibits the mTORC1 complex under low nutrient conditions. Promotes autophagy. Regulates cell polarity. Regulates lipid biosynthesis. Phosphorylates and stabilizes β-catenin. Phosphorylation at this site leads to activation and can occur through AMP/ATP-dependent means as well as -independent. | |
| AMPKα2 (T172) | ↑ | (Activation, intracellular localization, protein degradation) The catalytic subunit of the cellular energy sensing protein kinase AMP-activated protein kinase (AMPK). Inhibits energy consuming cellular processes (transcription, translation, ribosome biogenesis, etc.) under conditions of low ATP. Enhances glucose uptake and regulates insulin signaling and glycolysis. Enhances transcriptional activity of transcription factors involved in regulating energy metabolism. Directly inhibits the mTORC1 complex under low nutrient conditions. Promotes autophagy. Regulates cell polarity. Regulates lipid biosynthesis. Phosphorylates and stabilizes β-catenin. Phosphorylation at this site leads to activation and can occur through AMP/ATP-dependent means as well as independent. | ||
| Akt (S473) | ↓ | ↓ | (Activation, intracellular localization) Ser/Thr kinase involved in cell proliferation, survival, and metabolism. Regulates glucose uptake. Is activated by insulin and insulin-like growth factor (IGF). Activation and subsequent phosphorylation and inhibition of the downstream kinase GSK3 promotes the storage of glucose in the form of glycogen. AKT kinases are activated in response to a wide range of growth factors, cytokines, and stress leading to enhancement of mTOR-dependent signaling, activation of NF-κB-dependent gene transcription, and stimulation of CREB1-dependent transcription. Regulates Forkhead box protein transcription factors. Promotes β-catenin stabilization through the inhibition of GSK3B. Promotes the synthesis of anti-apoptotic proteins and results in the sequestering and degradation of pro-apoptotic proteins. Phosphorylation on this site is required for full activity. | |
| β-Catenin | ↓ | Transcriptional regulator of the WNT pathway. Under conditions favoring GSK3B activation, β-catenin is phosphorylated, ubiquitinated, and degraded. In the presence of WNT or the inhibition of GSK3B kinase activity, β-catenin is not degraded but accumulates in the nucleus where it serves as a coactivator for transcription. Regulates cell adhesion and anchorage-independent growth, promotes neurogenesis, regulates insulin internalization. | ||
| Chk-2 (T68) | ↑ | ↑ | (Activation, intracellular localization) Ser/Thr kinase required for cell cycle checkpoint arrest primarily at the G2/M transition point. Has a role in DNA repair and apoptotic cell death induced by DNA double-strand breaks. Phosphorylation at this site enhances homodimerization and subsequently full kinase activation. | |
| CREB (S133; i.e., S119) | ↓ | (Transcriptional activation) Transcription factor which binds to the cAMP response element present in many promoters of cellular and viral genes. Involved in diverse cellular processes. | ||
| ERK1/2 (T202/Y204, T185/Y187) | ↑ | (Activation, intracellular localization) Ser/Thr kinases central to the MAP kinase family. Have diverse roles in proliferation, transcription, translation, survival, cell cycle/cell division. Directly phosphorylates and regulates several transcription factors and translation regulatory proteins. | ||
| FAK (Y397) | ↑ | ↓ | (Activation, intracellular localization, protein degradation) Non-receptor protein tyrosine kinase involved in the regulation of cell adhesion, cell spreading, cell migration, cell proliferation, cell cycle progression, actin cytoskeletal rearrangements, and apoptosis. Required for normal embryonic development (angiogenesis, heart development, and nervous system development) and osteogenesis. Acts downstream of membrane receptors (cytokine, chemokine, growth factor, integrin, G-protein coupled, and immune) where it becomes tyrosine phosphorylated and associates in complex with Src tyrosine kinase family members. Additional tyrosine phosphorylation of PYK2 transforms it into a scaffolding protein capable of stimulating PI3K/AKT/mTOR, RAS/RAF/MEK/ERK and SAPK/JNK1 pathways as well as the translocation of MDM2 to the nucleus where it leads to p53 degradation. Phosphorylation or autophosphorylation at Y397 creates a docking site for SRC kinase family members that are then responsible for subsequent phosphorylation of Y576 and Y577 to induce full FAK activity. | |
| Fyn (Y420) | ↓ | (Intermolecular association) Non-receptor protein tyrosine kinase. Has a role in regulating cell growth and survival, immune response, cell motility, remodeling of the cytoskeleton, and integrin signaling. Following phosphorylation at the C-terminus associates with focal adhesion kinase FAK1 allowing for FAK1 phosphorylation and activation. Phosphorylates β- and Δ-catenins to regulate cellular adhesions. Promotes T-cell differentiation following binding of the T-cell receptor (TCR) through a mechanism involving focal adhesion kinase PYK2. Responsible for CD28 stimulation induced VAV1 activation. Activation of FYN is inhibited by phosphorylation at Y531 and activated by its dephosphorylation. Autophosphorylation of Y420 is required for full activation. | ||
| HSP27 (S78/S82) | ↑ | ↓ | (Activation, intracellular localization, protein degradation) Heat shock protein which functions as a molecular chaperone. Has a role in stress resistance by assisting proteins to maintain the correct folded state. Stress-induced phosphorylation at S78 and S82 impairs chaperone activity. | |
| HSP60 | ↓ | A chaperonin involved in mitochondrial protein import. It also serves for correct folding of mitochondrial matrix proteins following stress. | ||
| JNK (T183/Y185, T221/Y223) | ↑ | (Activation) Involved in cell proliferation, transformation, migration, differentiation, and death. Activated in response to proinflammatory cytokines or cellular stress. Through interaction and phosphorylation of AP-1 transcription factor components, leads to activation of AP-1-dependent transcription. Inhibits replication initiation (JNK1). Inhibits rRNA synthesis upon ribotoxic stress by inactivating RNA pol I (JNK2). Promotes stress induced cell apoptosis by phosphorylating p53 and YAP1. Necessary for Th1 polarization of T-helper cells. Promotes the degradation of β-catenin (JNK2). Can promote the activation of autophagic pathways (JNK1). JNK3 is specific to cells of the nervous system. | ||
| c-Jun (S63) | ↑ | (Acetylation, transcriptional activity induced, protein stabilized) Transcription factor that is a component of the AP-1 transcription factor family. Activation stimulates multiple genes regulating diverse cellular processes. | ||
| p27 (T198) | ↓ | (Altered protein stability, altered intracellular localization) Regulates cell cycle progression. Inhibits cyclin A- and cyclin E-CDK2 complexes but promotes cyclin D-CDK4 complexes depending on its phosphorylation (not only at this site). | ||
| p53 (S392) | ↑ | ↓ | (Transcriptional activation, intracellular localization, altered protein stability) Transcription factor involved in DNA repair, apoptosis, and cell cycle regulation. Regulates the circadian clock. Regulates early ribogenesis. Activated in response to stress. Phosphorylation of S392 has been linked to p53 protein stability and transcriptional activity. | |
| p53 (S46) | ↓ | (Transcriptional activation, intracellular localization, altered protein stability) Transcription factor involved in DNA repair, apoptosis, and cell cycle regulation. Regulates the circadian clock. Regulates early ribogenesis. Activated in response to stress. | ||
| p70 (T389) | ↓ | (Activation, intracellular localization) Ser/Thr kinase activated downstream of mTOR in response to growth promoting stimuli. Phosphorylates various downstream targets to regulate protein synthesis at the level of initiation and elongation. Has a feedback regulatory role on mTORC1/2 signaling. Has a role in promoting TNFα-induced insulin resistance. | ||
| p70 (T421/S424; i.e., T444/S447) | ↓ | (Activation) Ser/Thr kinase activated downstream of mTOR in response to growth promoting stimuli. Phosphorylates various downstream targets to regulate protein synthesis at the level of initiation and elongation. Has a feedback regulatory role on mTORC1/2 signaling. Has a role in promoting TNFα-induced insulin resistance. | ||
| PRAS40 (T246) | ↓ | (Activity inhibited) A negative regulatory subunit of the mTORC1 complex. Negative regulatory activity is relieved by phosphorylation on T246. | ||
| RSK1/2/3 (S380/S386/S377) | ↓ | (Activation, protein degradation, ubiquitination) Ser/Thr kinases downstream of ERK1/ERK2 responsible for regulation of proliferation, survival, and differentiation by modifying mTOR-dependent signaling. Directly influences factors regulating transcription and translation in response to mitogenic- or stress-mediated stimulation. Inhibits GSK3β activity by phosphorylating Ser9 of GSK3B. Promotes assembly of the translational preinitiation complex. Enhances CAP-dependent translation through phosphorylation of EIF4B. May suppress mTOR activity by phosphorylating TSC2. Involved in cell cycle regulation. Regulates osteoblast differentiation. Sites S380 (RSK1), S386 (RSK2), and S377 (RSK3) are activation promoting autophosphorylation sites. | ||
| Src (Y419) | ↑ | A proto-oncogenic non-receptor tyrosine kinase which has a role in numerous signaling events in the cell. Is a central component of the Src receptor signaling complex. | ||
| STAT2 (Y689) | ↓ | ↑ | (Intracellular localization) Signal transducer and activator of transcription. Mediates signaling by type I interferons (IFNs). Important for inducing the antiviral state. It also acts as a negative feedback regulator of IFNAR2. Acts as a regulator of mitochondrial fission. Phosphorylation of Y689 favors nuclear localization. | |
| STAT3 (Y705) | ↓ | ↓ | (Activation, intracellular localization, methylation, protein degradation) Signal transducer and activator of transcription. Mediates cellular responses to diverse interleukin (ILs) and growth factor receptor stimulation. Helps recruit coactivators of transcription to target genes. Involved in the T-cell inflammatory response. Cytoplasmic STAT3 can inhibit activation of the integrated stress response kinase PKR. Regulates β-cell insulin secretion. Phosphorylation at Y705 induces nuclear localization and transcriptional activation. | |
| STAT3 (S727) | ↓ | (Activity altered, intracellular localization) Signal transducer and activator of transcription. Mediates cellular responses to diverse interleukin (ILs) and growth factor receptor stimulation. Helps recruit coactivators of transcription to target genes. Involved in the T-cell inflammatory response. Cytoplasmic STAT3 can inhibit activation of the integrated stress response kinase PKR. Regulates β-cell insulin secretion. Phosphorylation at S727 is required for maximal transcriptional activation by enhancing DNA binding. | ||
| TOR (S2448) | ↑ | (Activation; intracellular localization) The central catalytic subunit of the mTORC1 and mTORC2 kinase complex, which act as major regulators of cellular growth, survival, and metabolism in response to nutrients, stress, energy, and growth factor stimulation. Is a downstream target of the PI3K-AKT pathway. Promotes protein synthesis through the phosphorylation of the eukaryotic initiation factor (eIF)-4E binding protein (4EBP) and by promoting the modification of ribosomal S6 protein. Stimulates ribosome biogenesis by enhancing RNA pol III activity. Regulates autophagy through the phosphorylation of ULK1 and DAP. | ||
| WNK1 (T60) | ↓ | (Activation) Ser/Thr kinase, regulates Na++/K++-chloride coupled receptors. Has a role in cytoskeletal reorganization. | ||
| Yes (Y426) | ↑ | ↑ | (Activation) Non-receptor tyrosine kinase (non-RTK) stimulated downstream of various RTKs. Involved in cell growth, survival, apoptosis, cell adhesion, cytoskeletal remodeling, and differentiation. Regulates G1 and G2/M phases of the cell cycle. Required for AKT-mediated cell migration. Phosphorylation at Y426 blocks inhibitory phosphorylation. |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Salucci, S.; Bavelloni, A.; Stella, A.B.; Fabbri, F.; Vannini, I.; Piazzi, M.; Volkava, K.; Scotlandi, K.; Martinelli, G.; Faenza, I.; et al. The Cytotoxic Effect of Curcumin in Rhabdomyosarcoma Is Associated with the Modulation of AMPK, AKT/mTOR, STAT, and p53 Signaling. Nutrients 2023, 15, 740. https://doi.org/10.3390/nu15030740
Salucci S, Bavelloni A, Stella AB, Fabbri F, Vannini I, Piazzi M, Volkava K, Scotlandi K, Martinelli G, Faenza I, et al. The Cytotoxic Effect of Curcumin in Rhabdomyosarcoma Is Associated with the Modulation of AMPK, AKT/mTOR, STAT, and p53 Signaling. Nutrients. 2023; 15(3):740. https://doi.org/10.3390/nu15030740
Chicago/Turabian StyleSalucci, Sara, Alberto Bavelloni, Anna Bartoletti Stella, Francesco Fabbri, Ivan Vannini, Manuela Piazzi, Karyna Volkava, Katia Scotlandi, Giovanni Martinelli, Irene Faenza, and et al. 2023. "The Cytotoxic Effect of Curcumin in Rhabdomyosarcoma Is Associated with the Modulation of AMPK, AKT/mTOR, STAT, and p53 Signaling" Nutrients 15, no. 3: 740. https://doi.org/10.3390/nu15030740
APA StyleSalucci, S., Bavelloni, A., Stella, A. B., Fabbri, F., Vannini, I., Piazzi, M., Volkava, K., Scotlandi, K., Martinelli, G., Faenza, I., & Blalock, W. (2023). The Cytotoxic Effect of Curcumin in Rhabdomyosarcoma Is Associated with the Modulation of AMPK, AKT/mTOR, STAT, and p53 Signaling. Nutrients, 15(3), 740. https://doi.org/10.3390/nu15030740