
Dark Chocolate Intake Positively Modulates Gut Permeability in Elite Football Athletes: A Randomized Controlled Study

 , , , ,
, , , ,  ,
,  and
and 
Abstract
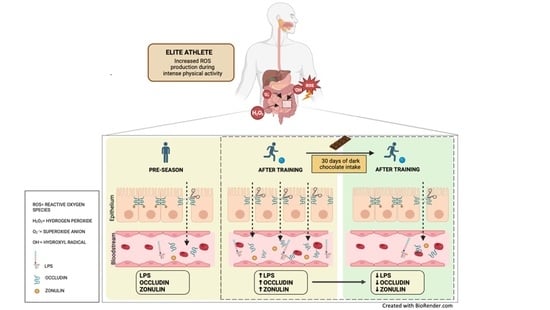
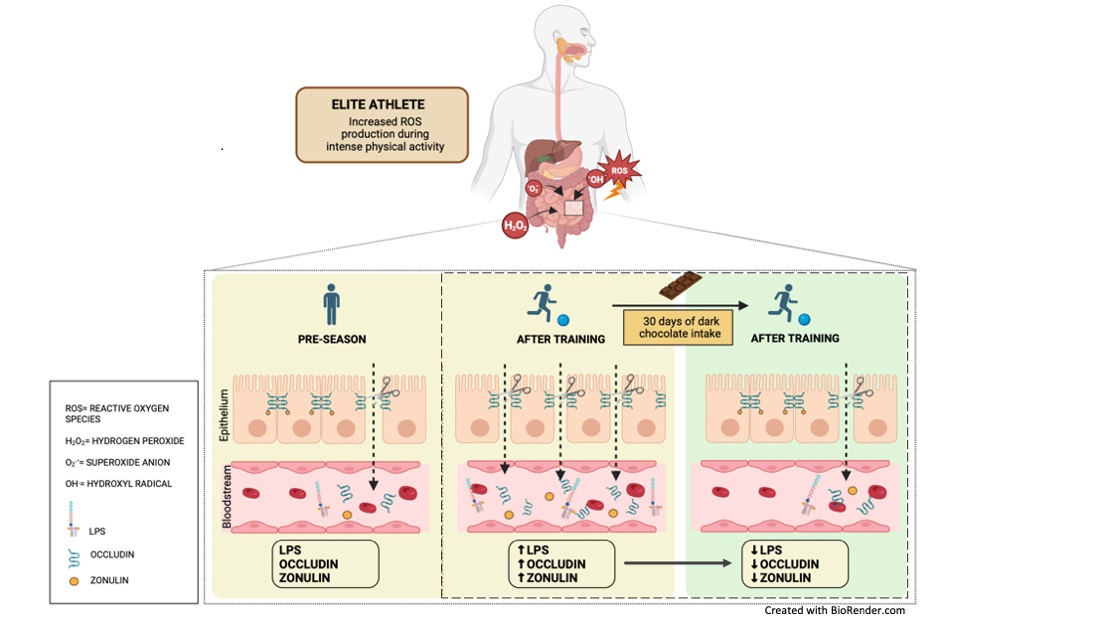
:
1. Introduction
2. Materials and Methods
2.1. Study Participants
2.2. Study Design
2.3. Study Product
2.4. Blood Sampling and Preparations
2.5. Evaluation of Gut Permeability Biomarkers
2.6. Cell Culture
2.7. Extraction of Phenolic Fraction from Chocolate
2.8. Caco-2 Cells Viability
2.9. Evaluation of Oxidative Stress Biomarkers
2.10. Protein Detection, Electrophoresis, and Western Blot Analysis
2.11. Statistical Analysis
3. Results
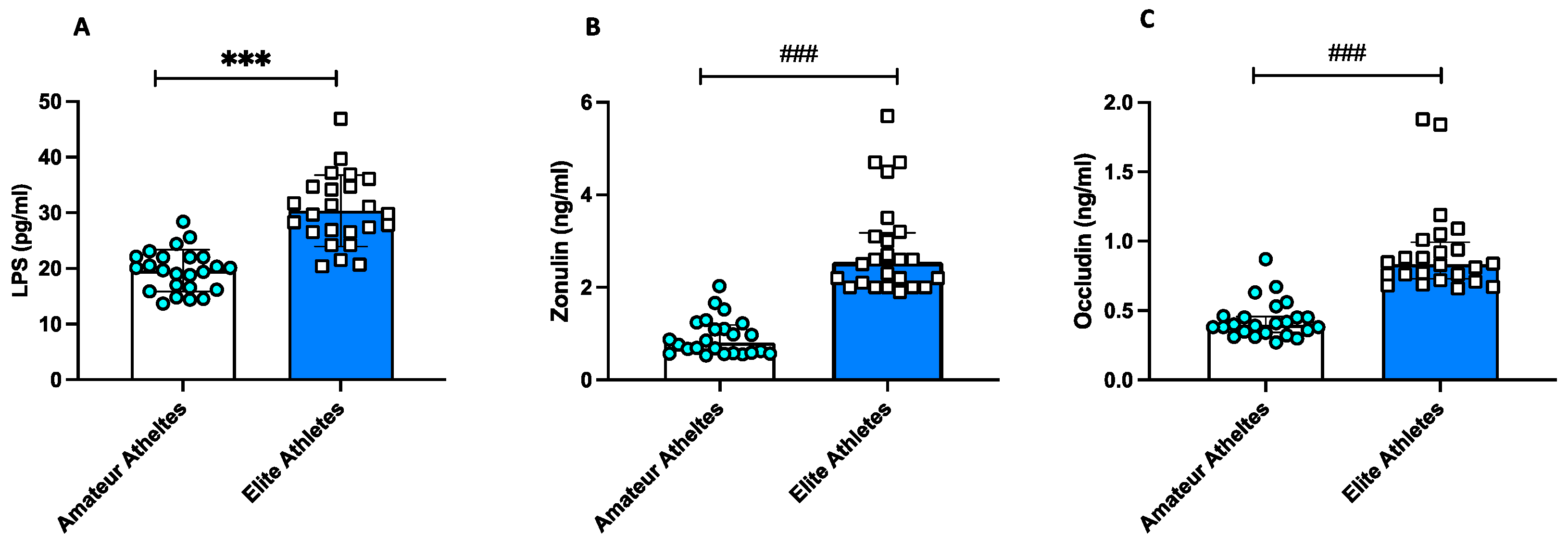
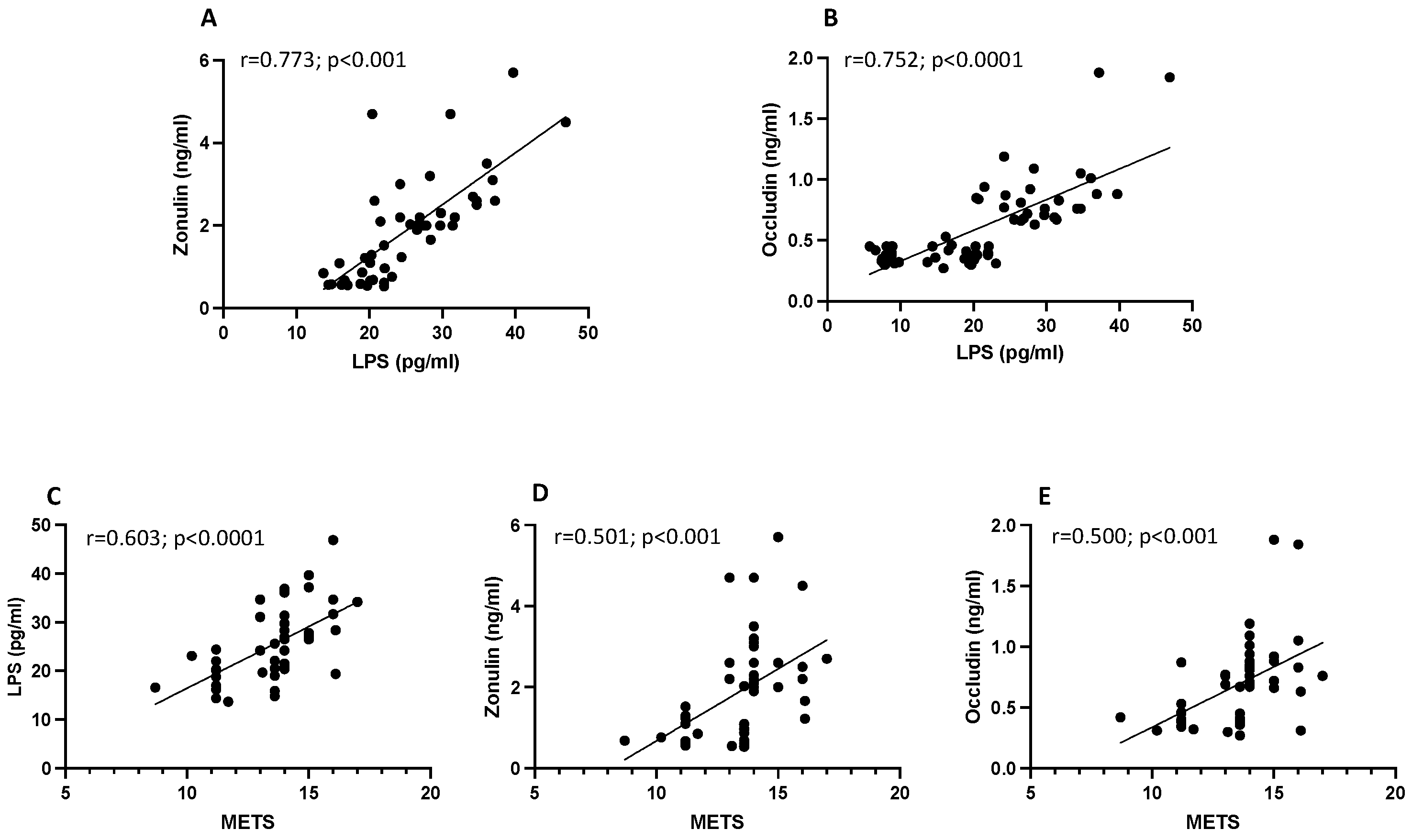
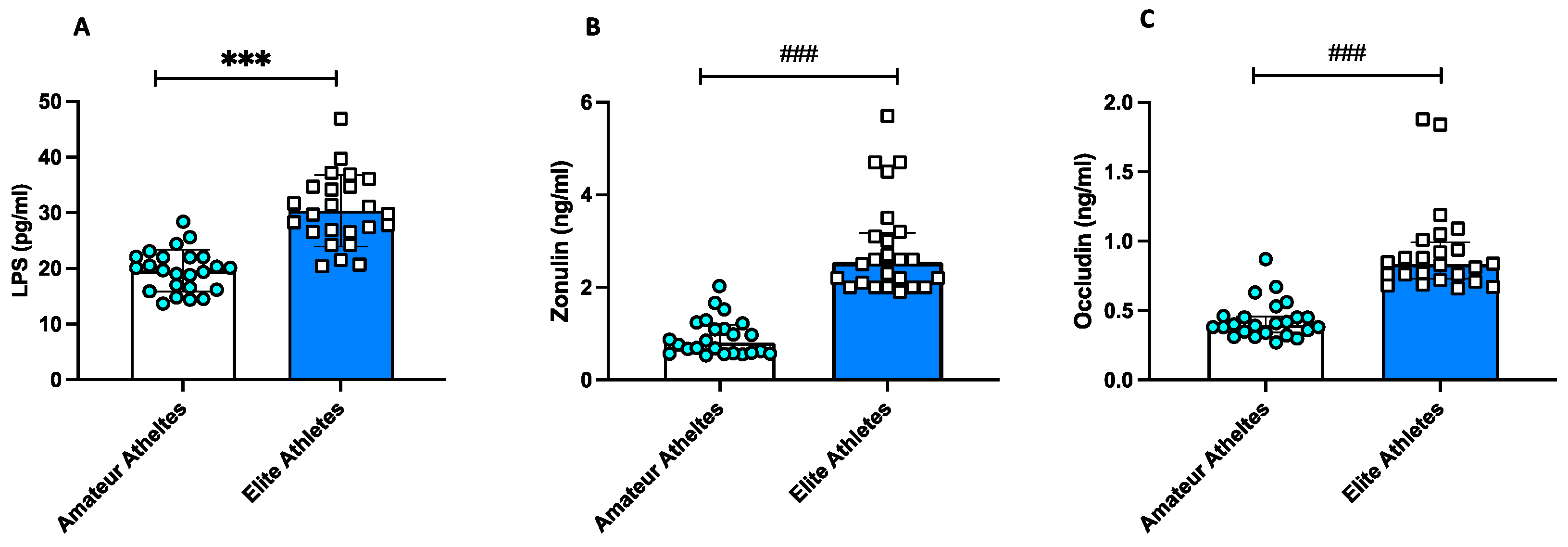
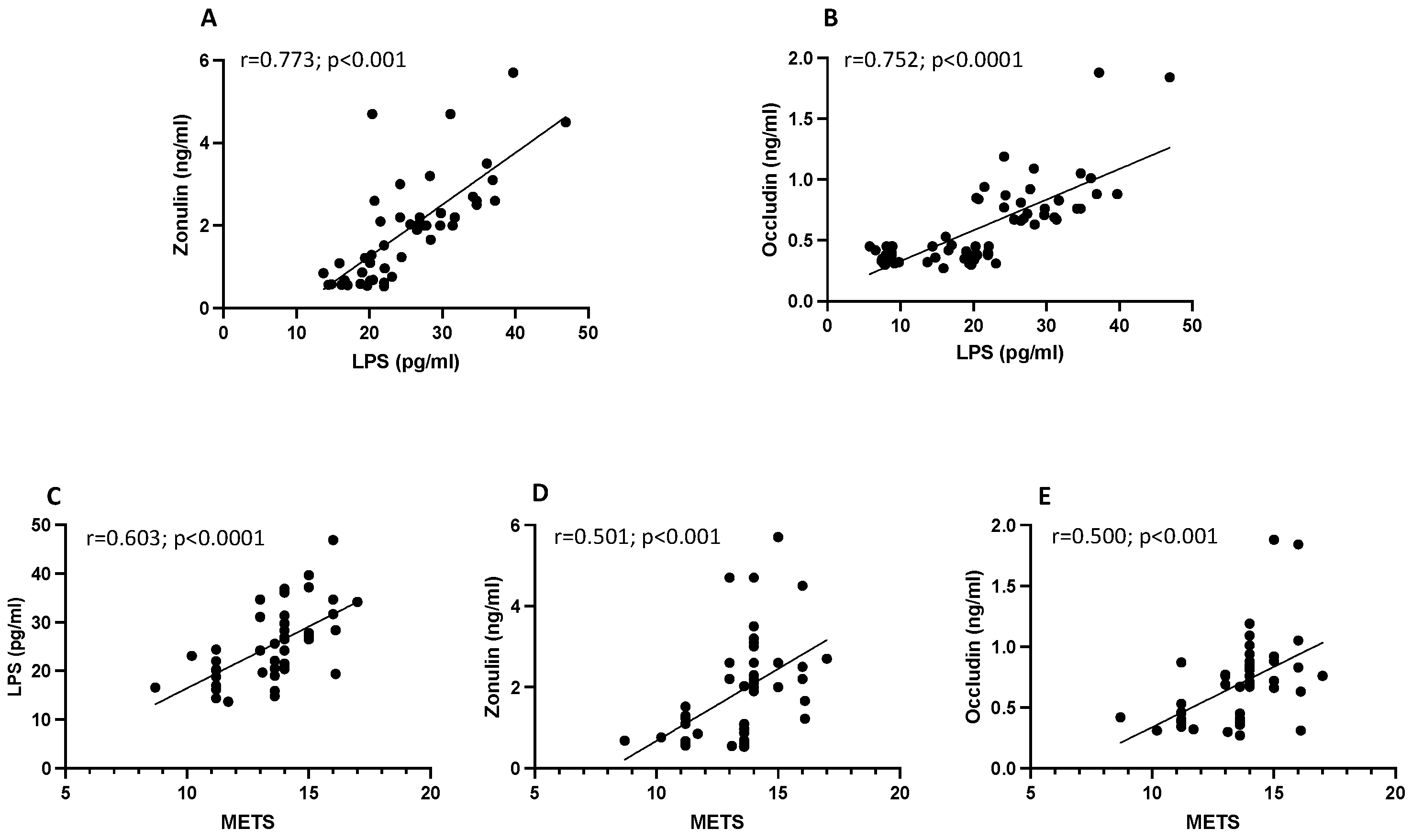
3.1. Observational Study
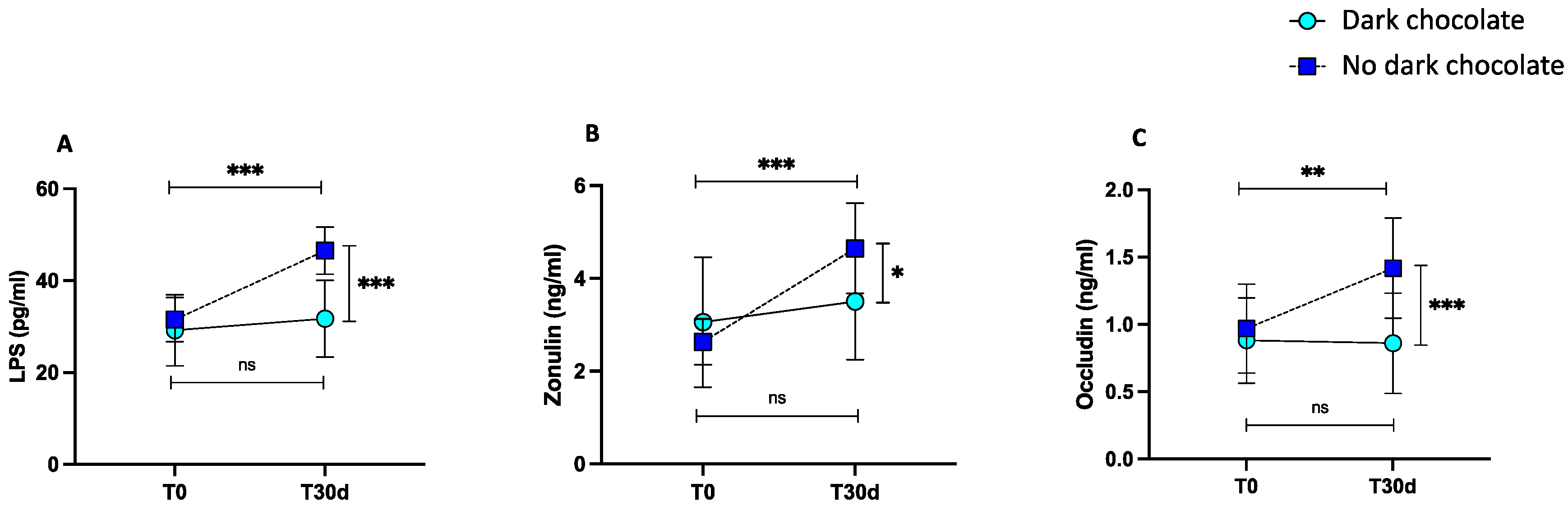
3.2. Intervention Study
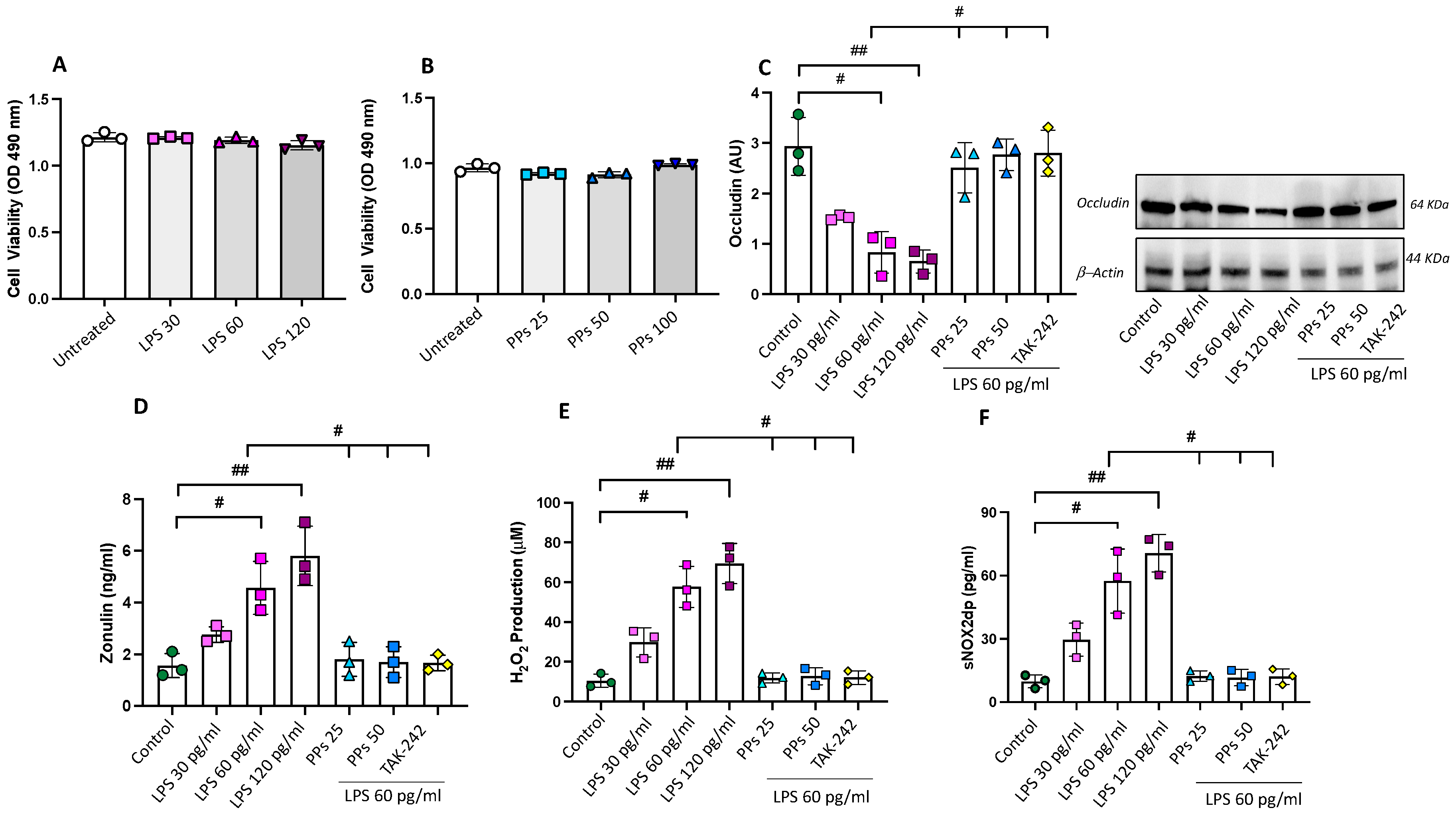
3.3. In Vitro Study
3.3.1. PPs Restore LPS-Induced Intestinal Epithelial Barrier Damage
3.3.2. PPs Reduce LPS-Induced Oxidative Stress
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Ghosh, S.S.; Wang, J.; Yannie, P.J.; Ghosh, S. Intestinal Barrier Dysfunction, LPS Translocation, and Disease Development. J. Endocr. Soc. 2020, 4, bvz039. [Google Scholar] [CrossRef] [PubMed]
- Ghosh, S.S.; Wang, J.; Yannie, P.J.; Cooper, R.C.; Sandhu, Y.K.; Kakiyama, G.; Korzun, W.J.; Ghosh, S. Over-Expression of Intestinal Alkaline Phosphatase Attenuates Atherosclerosis. Circ. Res. 2021, 128, 1646–1659. [Google Scholar] [CrossRef]
- Cataldi, S.; Poli, L.; Sahin, F.N.; Patti, A.; Santacroce, L.; Bianco, A.; Greco, G.; Ghinassi, B.; Di Baldassarre, A.; Fischetti, F. The Effects of Physical Activity on the Gut Microbiota and the Gut-Brain Axis in Preclinical and Human Models: A Narrative Review. Nutrients 2022, 14, 3293. [Google Scholar] [CrossRef] [PubMed]
- Oktedalen, O.; Lunde, O.C.; Opstad, P.K.; Aabakken, L.; Kvernebo, K. Changes in the gastrointestinal mucosa after long-distance running. Scand. J. Gastroenterol. 1992, 27, 270–274. [Google Scholar] [CrossRef] [PubMed]
- Ribeiro, F.M.; Petriz, B.; Marques, G.; Kamilla, L.H.; Franco, O.L. Is There an Exercise-Intensity Threshold Capable of Avoiding the Leaky Gut? Front. Nutr. 2021, 8, 627289. [Google Scholar] [CrossRef] [PubMed]
- March, D.S.; Marchbank, T.; Playford, R.J.; Jones, A.W.; Thatcher, R.; Davison, G. Intestinal fatty acid-binding protein and gut permeability responses to exercise. Eur. J. Appl. Physiol. 2017, 117, 931–941. [Google Scholar] [CrossRef]
- Mishra, A.; Makharia, G.K. Techniques of functional and motility test: How to perform and interpret intestinal permeability. J. Neurogastroenterol. Motil. 2012, 18, 443–447. [Google Scholar] [CrossRef]
- Hart, T.L.; Townsend, J.R.; Grady, N.J.; Johnson, K.D.; Littlefield, L.A.; Vergne, M.J.; Fundaro, G. Resistance Exercise Increases Gastrointestinal Symptoms, Markers of Gut Permeability, and Damage in Resistance-Trained Adults. Med. Sci. Sports Exerc. 2022, 54, 1761–1770. [Google Scholar] [CrossRef]
- Rodriguez-Daza, M.C.; Pulido-Mateos, E.C.; Lupien-Meilleur, J.; Guyonnet, D.; Desjardins, Y.; Roy, D. Polyphenol-Mediated Gut Microbiota Modulation: Toward Prebiotics and Further. Front. Nutr. 2021, 8, 689456. [Google Scholar] [CrossRef]
- Chantler, S.; Griffiths, A.; Matu, J.; Davison, G.; Holliday, A.; Jones, B. A systematic review: Role of dietary supplements on markers of exercise-associated gut damage and permeability. PLoS ONE 2022, 17, e0266379. [Google Scholar] [CrossRef]
- Buchman, A.L.; Killip, D.; Ou, C.N.; Rognerud, C.L.; Pownall, H.; Dennis, K.; Dunn, J.K. Short-term vitamin E supplementation before marathon running: A placebo-controlled trial. Nutrition 1999, 15, 278–283. [Google Scholar] [CrossRef]
- Szymanski, M.C.; Gillum, T.L.; Gould, L.M.; Morin, D.S.; Kuennen, M.R. Short-term dietary curcumin supplementation reduces gastrointestinal barrier damage and physiological strain responses during exertional heat stress. J. Appl. Physiol. 2018, 124, 330–340. [Google Scholar] [CrossRef] [PubMed]
- Khan, J.; Deb, P.K.; Priya, S.; Medina, K.D.; Devi, R.; Walode, S.G.; Rudrapal, M. Dietary Flavonoids: Cardioprotective Potential with Antioxidant Effects and Their Pharmacokinetic, Toxicological and Therapeutic Concerns. Molecules 2021, 26, 4021. [Google Scholar] [CrossRef]
- Rudrapal, M.; Khairnar, S.J.; Khan, J.; Dukhyil, A.B.; Ansari, M.A.; Alomary, M.N.; Alshabrmi, F.M.; Palai, S.; Deb, P.K.; Devi, R. Dietary Polyphenols and Their Role in Oxidative Stress-Induced Human Diseases: Insights Into Protective Effects, Antioxidant Potentials and Mechanism(s) of Action. Front. Pharmacol. 2022, 13, 806470. [Google Scholar] [CrossRef]
- Niwano, Y.; Kohzaki, H.; Shirato, M.; Shishido, S.; Nakamura, K. Putative Mechanisms Underlying the Beneficial Effects of Polyphenols in Murine Models of Metabolic Disorders in Relation to Gut Microbiota. Curr. Issues Mol. Biol. 2022, 44, 1353–1375. [Google Scholar] [CrossRef]
- Zare, R.; Kimble, R.; Ali Redha, A.; Cerullo, G.; Clifford, T. How can chokeberry (Aronia) (poly)phenol-rich supplementation help athletes? A systematic review of human clinical trials. Food Funct. 2023, 14, 5478–5491. [Google Scholar] [CrossRef] [PubMed]
- Abbott, W.; Hansell, E.J.; Brett, A.; Skarabot, J.; James, L.J.; Clifford, T. Curcumin Attenuates Delayed-Onset Muscle Soreness and Muscle Function Deficits Following a Soccer Match in Male Professional Soccer Players. Int. J. Sports Physiol. Perform. 2023, 18, 347–353. [Google Scholar] [CrossRef] [PubMed]
- Bowtell, J.; Kelly, V. Fruit-Derived Polyphenol Supplementation for Athlete Recovery and Performance. Sports Med. 2019, 49, 3–23. [Google Scholar] [CrossRef]
- Nieman, D.C.; Gillitt, N.D.; Knab, A.M.; Shanely, R.A.; Pappan, K.L.; Jin, F.; Lila, M.A. Influence of a polyphenol-enriched protein powder on exercise-induced inflammation and oxidative stress in athletes: A randomized trial using a metabolomics approach. PLoS ONE 2013, 8, e72215. [Google Scholar] [CrossRef]
- Xu, Z.; Sun, X.; Ding, B.; Zi, M.; Ma, Y. Resveratrol attenuated high intensity exercise training-induced inflammation and ferroptosis via Nrf2/FTH1/GPX4 pathway in intestine of mice. Turk. J. Med. Sci. 2023, 53, 446–454. [Google Scholar] [CrossRef]
- Cavarretta, E.; Peruzzi, M.; Del Vescovo, R.; Di Pilla, F.; Gobbi, G.; Serdoz, A.; Ferrara, R.; Schirone, L.; Sciarretta, S.; Nocella, C.; et al. Dark Chocolate Intake Positively Modulates Redox Status and Markers of Muscular Damage in Elite Football Athletes: A Randomized Controlled Study. Oxid. Med. Cell Longev. 2018, 2018, 4061901. [Google Scholar] [CrossRef] [PubMed]
- D’Amico, A.; Cavarretta, E.; Fossati, C.; Borrione, P.; Pigozzi, F.; Frati, G.; Sciarretta, S.; Costa, V.; De Grandis, F.; Nigro, A.; et al. Platelet Activation Favours NOX2-Mediated Muscle Damage in Elite Athletes: The Role of Cocoa-Derived Polyphenols. Nutrients 2022, 14, 1558. [Google Scholar] [CrossRef]
- Van der Slikke, R.M.A.; Sindall, P.; Goosey-Tolfrey, V.L.; Mason, B.S. Load and performance monitoring in wheelchair court sports: A narrative review of the use of technology and practical recommendations. Eur. J. Sport. Sci. 2023, 23, 189–200. [Google Scholar] [CrossRef]
- Pelliccia, A.; Caselli, S.; Sharma, S.; Basso, C.; Bax, J.J.; Corrado, D.; D’Andrea, A.; D’Ascenzi, F.; Di Paolo, F.M.; Edvardsen, T.; et al. European Association of Preventive Cardiology (EAPC) and European Association of Cardiovascular Imaging (EACVI) joint position statement: Recommendations for the indication and interpretation of cardiovascular imaging in the evaluation of the athlete’s heart. Eur. Heart J. 2018, 39, 1949–1969. [Google Scholar] [CrossRef] [PubMed]
- Perez-Diaz-Del-Campo, N.; Castelnuovo, G.; Ribaldone, D.G.; Caviglia, G.P. Fecal and Circulating Biomarkers for the Non-Invasive Assessment of Intestinal Permeability. Diagnostics 2023, 13, 1976. [Google Scholar] [CrossRef] [PubMed]
- Matsunaga, N.; Tsuchimori, N.; Matsumoto, T.; Ii, M. TAK-242 (resatorvid), a small-molecule inhibitor of Toll-like receptor (TLR) 4 signaling, binds selectively to TLR4 and interferes with interactions between TLR4 and its adaptor molecules. Mol. Pharmacol. 2011, 79, 34–41. [Google Scholar] [CrossRef] [PubMed]
- Carnevale, R.; Silvestri, R.; Loffredo, L.; Novo, M.; Cammisotto, V.; Castellani, V.; Bartimoccia, S.; Nocella, C.; Violi, F. Oleuropein, a component of extra virgin olive oil, lowers postprandial glycaemia in healthy subjects. Br. J. Clin. Pharmacol. 2018, 84, 1566–1574. [Google Scholar] [CrossRef]
- Lee, S.H. Intestinal permeability regulation by tight junction: Implication on inflammatory bowel diseases. Intest. Res. 2015, 13, 11–18. [Google Scholar] [CrossRef]
- Olsson, A.; Gustavsen, S.; Langkilde, A.R.; Hansen, T.H.; Sellebjerg, F.; Bach Sondergaard, H.; Oturai, A.B. Circulating levels of tight junction proteins in multiple sclerosis: Association with inflammation and disease activity before and after disease modifying therapy. Mult. Scler. Relat. Disord. 2021, 54, 103136. [Google Scholar] [CrossRef]
- Fasano, A. Intestinal permeability and its regulation by zonulin: Diagnostic and therapeutic implications. Clin. Gastroenterol. Hepatol. 2012, 10, 1096–1100. [Google Scholar] [CrossRef]
- Keirns, B.H.; Koemel, N.A.; Sciarrillo, C.M.; Anderson, K.L.; Emerson, S.R. Exercise and intestinal permeability: Another form of exercise-induced hormesis? Am. J. Physiol. Gastrointest. Liver Physiol. 2020, 319, G512–G518. [Google Scholar] [CrossRef]
- Chantler, S.; Griffiths, A.; Matu, J.; Davison, G.; Jones, B.; Deighton, K. The Effects of Exercise on Indirect Markers of Gut Damage and Permeability: A Systematic Review and Meta-analysis. Sports Med. 2021, 51, 113–124. [Google Scholar] [CrossRef]
- Gutekunst, K.; Kruger, K.; August, C.; Diener, M.; Mooren, F.C. Acute exercises induce disorders of the gastrointestinal integrity in a murine model. Eur. J. Appl. Physiol. 2014, 114, 609–617. [Google Scholar] [CrossRef] [PubMed]
- Bonomini-Gnutzmann, R.; Plaza-Diaz, J.; Jorquera-Aguilera, C.; Rodriguez-Rodriguez, A.; Rodriguez-Rodriguez, F. Effect of Intensity and Duration of Exercise on Gut Microbiota in Humans: A Systematic Review. Int. J. Environ. Res. Public Health 2022, 19, 9518. [Google Scholar] [CrossRef]
- Taranu, I.; Pistol, G.C.; Anghel, A.C.; Marin, D.; Bulgaru, C. Yeast-Fermented Rapeseed Meal Extract Is Able to Reduce Inflammation and Oxidative Stress Caused by Escherichia coli Lipopolysaccharides and to Replace ZnO in Caco-2/HTX29 Co-Culture Cells. Int. J. Mol. Sci. 2022, 23, 11640. [Google Scholar] [CrossRef]
- Carnevale, R.; Nocella, C.; Petrozza, V.; Cammisotto, V.; Pacini, L.; Sorrentino, V.; Martinelli, O.; Irace, L.; Sciarretta, S.; Frati, G.; et al. Localization of lipopolysaccharide from Escherichia Coli into human atherosclerotic plaque. Sci. Rep. 2018, 8, 3598. [Google Scholar] [CrossRef] [PubMed]
- Carnevale, R.; Sciarretta, S.; Valenti, V.; di Nonno, F.; Calvieri, C.; Nocella, C.; Frati, G.; Forte, M.; d’Amati, G.; Pignataro, M.G.; et al. Low-grade endotoxaemia enhances artery thrombus growth via Toll-like receptor 4: Implication for myocardial infarction. Eur. Heart J. 2020, 41, 3156–3165. [Google Scholar] [CrossRef] [PubMed]
- Violi, F.; Cammisotto, V.; Bartimoccia, S.; Pignatelli, P.; Carnevale, R.; Nocella, C. Gut-derived low-grade endotoxaemia, atherothrombosis and cardiovascular disease. Nat. Rev. Cardiol. 2023, 20, 24–37. [Google Scholar] [CrossRef]
- Bernardi, S.; Del Bo, C.; Marino, M.; Gargari, G.; Cherubini, A.; Andres-Lacueva, C.; Hidalgo-Liberona, N.; Peron, G.; Gonzalez-Dominguez, R.; Kroon, P.; et al. Polyphenols and Intestinal Permeability: Rationale and Future Perspectives. J. Agric. Food Chem. 2020, 68, 1816–1829. [Google Scholar] [CrossRef]
- Kaulmann, A.; Bohn, T. Bioactivity of Polyphenols: Preventive and Adjuvant Strategies toward Reducing Inflammatory Bowel Diseases-Promises, Perspectives, and Pitfalls. Oxid. Med. Cell Longev. 2016, 2016, 9346470. [Google Scholar] [CrossRef]
- Man, A.L.; Bertelli, E.; Rentini, S.; Regoli, M.; Briars, G.; Marini, M.; Watson, A.J.; Nicoletti, C. Age-associated modifications of intestinal permeability and innate immunity in human small intestine. Clin. Sci. 2015, 129, 515–527. [Google Scholar] [CrossRef] [PubMed]
- Wilms, E.; Troost, F.J.; Elizalde, M.; Winkens, B.; de Vos, P.; Mujagic, Z.; Jonkers, D.; Masclee, A.A.M. Intestinal barrier function is maintained with aging—A comprehensive study in healthy subjects and irritable bowel syndrome patients. Sci. Rep. 2020, 10, 475. [Google Scholar] [CrossRef] [PubMed]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Amateur Athletes (n = 23) | Elite Football Players (n = 24) | p | |
|---|---|---|---|
| Age (years) | 30.2 ± 4.6 | 17.2 ± 0.7 | <0.0001 |
| Gender (M/F) | 23/0 | 24/0 | - |
| WBCs (×103 μL) | 7.2 ± 2.2 | 5.6 ± 1.3 | <0.01 |
| PLTs (×103 μL) | 215.3 ± 40.3 | 228.0 ± 39.7 | 0.282 |
| RBCs (×106 μL) | 5.1 ± 0.4 | 5.8 ± 0.3 | <0.0001 |
| Cholesterol (mg/dL) | 185.1 ± 30.8 | 172.3 ± 29.4 | 0.151 |
| Glycemia (mg/dL) | 89.0 ± 28.8 | 83.5 ± 15.2 | 0.414 |
| BMI | 24.3 ± 1.9 | 22.5 ± 1.5 | <0.001 |
| Heart rate at rest (bpm) | 62.1 ± 10.4 | 56.3 ± 11.6 | 0.08 |
| Systolic blood pressure (mmHg) | 114.8 ± 11.2 | 111.7 ± 7.9 | 0.284 |
| Diastolic blood pressure (mmHg) | 74.1 ± 7.2 | 70.9 ± 6.8 | 0.128 |
| Training per week (h) | 5.1 ± 2.0 | 18.0 ± 2.0 | <0.0001 |
| Maximum workload (METs) | 12.3 ± 1.9 | 15.1 ± 1.4 | <0.001 |
| Peak heart rate (bpm) | 164.7 ± 6.9 | 169 ± 11.5 | 0.131 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Nocella, C.; Cavarretta, E.; Fossati, C.; Pigozzi, F.; Quaranta, F.; Peruzzi, M.; De Grandis, F.; Costa, V.; Sharp, C.; Manara, M.; et al. Dark Chocolate Intake Positively Modulates Gut Permeability in Elite Football Athletes: A Randomized Controlled Study. Nutrients 2023, 15, 4203. https://doi.org/10.3390/nu15194203
Nocella C, Cavarretta E, Fossati C, Pigozzi F, Quaranta F, Peruzzi M, De Grandis F, Costa V, Sharp C, Manara M, et al. Dark Chocolate Intake Positively Modulates Gut Permeability in Elite Football Athletes: A Randomized Controlled Study. Nutrients. 2023; 15(19):4203. https://doi.org/10.3390/nu15194203
Chicago/Turabian StyleNocella, Cristina, Elena Cavarretta, Chiara Fossati, Fabio Pigozzi, Federico Quaranta, Mariangela Peruzzi, Fabrizio De Grandis, Vincenzo Costa, Carwyn Sharp, Massimo Manara, and et al. 2023. "Dark Chocolate Intake Positively Modulates Gut Permeability in Elite Football Athletes: A Randomized Controlled Study" Nutrients 15, no. 19: 4203. https://doi.org/10.3390/nu15194203
APA StyleNocella, C., Cavarretta, E., Fossati, C., Pigozzi, F., Quaranta, F., Peruzzi, M., De Grandis, F., Costa, V., Sharp, C., Manara, M., Nigro, A., Cammisotto, V., Castellani, V., Picchio, V., Sciarretta, S., Frati, G., Bartimoccia, S., D’Amico, A., & Carnevale, R. (2023). Dark Chocolate Intake Positively Modulates Gut Permeability in Elite Football Athletes: A Randomized Controlled Study. Nutrients, 15(19), 4203. https://doi.org/10.3390/nu15194203