
Transcriptomic Analysis of LNCaP Tumor Xenograft to Elucidate the Components and Mechanisms Contributed by Tumor Environment as Targets for Dietary Prostate Cancer Prevention Studies


Abstract
1. Introduction
2. Materials and Methods
2.1. Chemicals and Reagents
2.2. Cell and Cell Culture
2.3. In Vivo Xenograft Bioassay
2.4. RNA Extraction and Sequencing Using RNA-Seq Technology
2.5. Data Analysis and Bioinformatics
2.5.1. Principal Component Analysis
2.5.2. Volcano Plot Analysis
2.5.3. Canonical Pathway Analysis of Data Sets
2.5.4. Network/Pathways Graphical Representation
2.6. In Vitro Modeling of Androgen Exposure, Cell–Matrix Interaction, Inflammation, and Hypoxia
2.6.1. Effects of Androgen on Gene Expressions
2.6.2. Effects of Hypoxia on Gene Expressions
2.6.3. Effects of Tumor Cell–Immune Cell Interaction on Gene Expressions
2.6.4. Effects of Subcellular Matrix on Gene Expressions
2.7. Real-Time PCR Analysis of Gene Expression
2.8. Statistical Analysis
3. Results
3.1. Global Transcriptomic Comparison of Parent LNCaP Cells and LNCaP Cell Tumor Xenograft
3.1.1. Principal Component Analysis of Cultured LNCaP Cells vs. LNCaP Xenograft Tumor
3.1.2. Volcano Plot of Cultured LNCaP Cells vs. LNCaP Xenograft Tumor
3.1.3. Comparison of Gene Expression in Cultured LNCaP Cells vs. LNCaP Xenograft Tumor Using IPA
3.2. Elucidating Pathway Differences between LNCaP Xenograft Tumor and Cultured LNCaP Cells
3.2.1. Top Canonical Pathways Differences Identified from IPA Analysis
3.2.2. Identification of Top Networks Differences
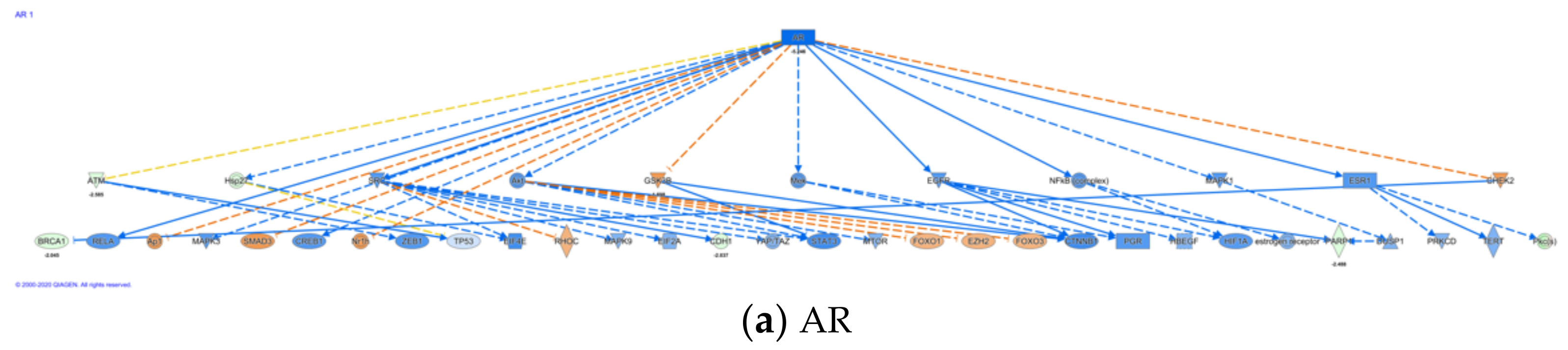
3.2.3. Identification of Top Upstream Regulators and Top Causal Networks
3.2.4. Alteration in Cytokine- and Chemokine-Related Networks
3.2.5. Genes/Pathways Alteration Associated with Cell–Cell Matrix/Adhesion
3.3. Validation of Gene Expressions in LNCaP Xenograft Tumors Compared to Cultured Cells
3.4. The Effects of In Vitro Environments on LNCaP Gene Expressions
3.4.1. Genes Responsive to Androgen
3.4.2. Genes Associated with Hypoxia
3.4.3. Genes Associated with Subcellular Matrix
3.4.4. Genes Associated with Tumor Cell–Immune Cell Interaction
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Wu, X.; Gong, S.; Roy-Burman, P.; Lee, P.; Culig, Z. Current mouse and cell models in prostate cancer research. Endocr. Relat. Cancer 2013, 20, R155–R170. [Google Scholar] [CrossRef]
- Parisotto, M.; Metzger, D. Genetically engineered mouse models of prostate cancer. Mol. Oncol. 2013, 7, 190–205. [Google Scholar] [CrossRef] [PubMed]
- Bosland, M.C. Use of animal models in defining efficacy of chemoprevention agents against prostate cancer. Eur. Urol. 1999, 35, 459–463. [Google Scholar] [CrossRef] [PubMed]
- Mavropoulos, J.C.; Buschemeyer, W.C., 3rd; Tewari, A.K.; Rokhfeld, D.; Pollak, M.; Zhao, Y.; Febbo, P.G.; Cohen, P.; Hwang, D.; Devi, G.; et al. The effects of varying dietary carbohydrate and fat content on survival in a murine LNCaP prostate cancer xenograft model. Cancer Prev. Res. 2009, 2, 557–565. [Google Scholar] [CrossRef] [PubMed]
- Aggarwal, M.; Saxena, R.; Asif, N.; Sinclair, E.; Tan, J.; Cruz, I.; Berry, D.; Kallakury, B.; Pham, Q.; Wang, T.T.Y.; et al. p53 mutant-type in human prostate cancer cells determines the sensitivity to phenethyl isothiocyanate induced growth inhibition. J. Exp. Clin. Cancer Res. 2019, 38, 1–17. [Google Scholar] [CrossRef] [PubMed]
- Wu, Y.; Li, R.W.; Huang, H.; Fletcher, A.; Yu, L.; Pham, Q.; Yu, L.; He, Q.; Wang, T.T.Y. Inhibition of Tumor Growth by Dietary Indole-3-Carbinol in a Prostate Cancer Xenograft Model May Be Associated with Disrupted Gut Microbial Interactions. Nutrients 2019, 11, 467. [Google Scholar] [CrossRef] [PubMed]
- Grabowska, M.M.; DeGraff, D.J.; Yu, X.; Jin, R.J.; Chen, Z.; Borowsky, A.D.; Matusik, R.J. Mouse models of prostate cancer: Picking the best model for the question. Cancer Metastasis Rev. 2014, 33, 377–397. [Google Scholar] [CrossRef]
- Greenberg, N.M.; DeMayo, F.; Finegold, M.J.; Medina, D.; Tilley, W.D.; Aspinall, J.O.; Cunha, G.R.; Donjacour, A.A.; Matusik, R.J.; Rosen, J.M. Prostate cancer in a transgenic mouse. Proc. Natl. Acad. Sci. USA 1995, 92, 3439–3443. [Google Scholar] [CrossRef] [PubMed]
- Tian, H.; Lyu, Y.; Yang, Y.-G.; Hu, Z. Humanized Rodent Models for Cancer Research. Front. Oncol. 2020, 10. [Google Scholar] [CrossRef]
- Ittmann, M.; Huang, J.; Radaelli, E.; Martin, P.; Signoretti, S.; Sullivan, R.; Simons, B.W.; Ward, J.M.; Robinson, B.D.; Chu, G.C.; et al. Animal Models of Human Prostate Cancer: The Consensus Report of the New York Meeting of the Mouse Models of Human Cancers Consortium Prostate Pathology Committee. Cancer Res. 2013, 73, 2718–2736. [Google Scholar] [CrossRef]
- Shultz, L.D.; Goodwin, N.; Ishikawa, F.; Hosur, V.; Lyons, B.L.; Greiner, D.L. Human Cancer Growth and Therapy in Immunodeficient Mouse Models. Cold Spring Harb. Protoc. 2014, 2014, 694–708. [Google Scholar] [CrossRef] [PubMed]
- Pandelouris, E. Absence of thymus in a mouse mutant. Nature 1968, 217, 370–371. [Google Scholar] [CrossRef] [PubMed]
- Fleshner, N.; Fair, W.; Huryk, R.; Heston, W. Vitamin E inhibits the high-fat diet promoted growth of established human prostate LNCaP tumors in nude mice. J. Urol. 1999, 161, 1651–1654. [Google Scholar] [CrossRef]
- Morton, C.L.; Houghton, P.J. Establishment of human tumor xenografts in immunodeficient mice. Nat. Protoc. 2007, 2, 247–250. [Google Scholar] [CrossRef]
- Van Weerden, W.M.; Bangma, C.; De Wit, R. Human xenograft models as useful tools to assess the potential of novel therapeutics in prostate cancer. Br. J. Cancer 2008, 100, 13–18. [Google Scholar] [CrossRef]
- Zhuang, L.; Kim, J.; Adam, R.M.; Solomon, K.R.; Freeman, M.R. Cholesterol targeting alters lipid raft composition and cell survival in prostate cancer cells and xenografts. J. Clin. Investig. 2005, 115, 959–968. [Google Scholar] [CrossRef]
- Namekawa, T.; Ikeda, K.; Horie-Inoue, K.; Inoue, S. Application of Prostate Cancer Models for Preclinical Study: Advantages and Limitations of Cell Lines, Patient-Derived Xenografts, and Three-Dimensional Culture of Patient-Derived Cells. Cells 2019, 8, 74. [Google Scholar] [CrossRef]
- Litvinov, I.V.; Antony, L.; Dalrymple, S.L.; Becker, R.; Cheng, L.; Isaacs, J.T. PC3, but not DU145, human prostate cancer cells retain the coregulators required for tumor suppressor ability of androgen receptor. Prostate 2006, 66, 1329–1338. [Google Scholar] [CrossRef]
- Horoszewicz, J.S.; Leong, S.S.; Kawinski, E.; Karr, J.P.; Rosenthal, H.; Chu, T.M.; Mirand, E.A.; Murphy, G.P. LNCaP model of human prostatic carcinoma. Cancer Res. 1983, 43, 1809–1818. [Google Scholar] [PubMed]
- Young, C.Y.; Montgomery, B.T.; Andrews, P.E.; Qui, S.D.; Bilhartz, D.L.; Tindall, D.J. Hormonal regulation of prostate-specific antigen messenger RNA in human prostatic adenocarcinoma cell line LNCaP. Cancer Res. 1991, 51, 3748–3752. [Google Scholar] [PubMed]
- Polyak, K.; Haviv, I.; Campbell, I.G. Co-evolution of tumor cells and their microenvironment. Trends Genet. 2009, 25, 30–38. [Google Scholar] [CrossRef]
- Grigoryev, D.N.; Long, B.J.; Njar, V.C.; Brodie, A.H. Pregnenolone stimulates LNCaP prostate cancer cell growth via the mutated androgen receptor. J. Steroid Biochem. Mol. Biol. 2000, 75, 1–10. [Google Scholar] [CrossRef]
- Vukanovic, J.; Isaacs, J.T. Human prostatic cancer cells are sensitive to programmed (apoptotic) death induced by the antiangiogenic agent linomide. Cancer Res. 1995, 55, 3517–3520. [Google Scholar] [PubMed]
- Papadopoulou, M.V.; Ji, M.; Ji, X.; Bloomer, W.D.; Hollingshead, M.G. Therapeutic advantage from combining paclitaxel with the hypoxia-selective cytotoxin NLCQ-1 in murine tumor- or human xenograft-bearing mice. Cancer Chemother Pharmacol. 2002, 50, 501–508. [Google Scholar] [CrossRef] [PubMed]
- Bucana, C.D.; Fabra, A.; Sanchez, R.; Fidler, I.J. Different patterns of macrophage infiltration into allogeneic-murine and xenogeneic-human neoplasms growing in nude mice. Am. J. Pathol. 1992, 141, 1225–1236. [Google Scholar] [PubMed]
- Niles, R.M.; Cook, C.P.; Meadows, G.G.; Fu, Y.M.; McLaughlin, J.L.; Rankin, G.O. Resveratrol is rapidly metabolized in athymic (nu/nu) mice and does not inhibit human melanoma xenograft tumor growth. J. Nutr. 2006, 136, 2542–2546. [Google Scholar] [CrossRef] [PubMed]
- Goodwin, S.; McPherson, J.; McCombie, W. Coming of age: Ten years of next-generation sequencing technologies. Nat. Rev. Genet. 2016, 17, 333–351. [Google Scholar] [CrossRef] [PubMed]
- Yu, L.; Pham, Q.; Yu, L.L.; Wang, T.T.Y. Modulation of CXC-motif chemokine receptor 7, but not 4, expression is related to migration of the human prostate cancer cell LNCaP: Regulation by androgen and inflammatory stimuli. Inflamm. Res. 2020, 69, 167–178. [Google Scholar] [CrossRef] [PubMed]
- Kao, F.T.; Hartz, J.; Horton, R.; Nemerson, Y.; Carson, S.D. Regional assignment of human tissue factor gene (F3) to chromosome 1p21-p22. Somat. Cell Mol. Genet. 1988, 14, 407–410. [Google Scholar] [CrossRef]
- Vellanki, R.N.; Zhang, L.; Volchuk, A. OASIS/CREB3L1 is induced by endoplasmic reticulum stress in human glioma cell lines and contributes to the unfolded protein response, extracellular matrix production and cell migration. PLoS ONE 2013, 8, e54060. [Google Scholar] [CrossRef]
- Astrup, L.B.; Skovgaard, K.; Rasmussen, R.S.; Iburg, T.M.; Agerholm, J.S.; Aalbæk, B.; Jensen, H.E.; Nielsen, O.L.; Johansen, F.F.; Heegaard, P.M.H.; et al. Staphylococcus aureus infected embolic stroke upregulates Orm1 and Cxcl2 in a rat model of septic stroke pathology. Neurol. Res. 2019, 41, 399–412. [Google Scholar] [CrossRef]
- Cao, X.; Qin, J.; Xie, Y.; Khan, O.; Dowd, F.; Scofield, M.; Lin, M.; Tu, Y. Regulator of G-protein signaling 2 (RGS2) inhibits androgen-independent activation of androgen receptor in prostate cancer cells. Oncogene 2006, 25, 3719–3734. [Google Scholar] [CrossRef]
- Eckfeld, C.; Häußler, D.; Schoeps, B.; Hermann, C.D.; Krüger, A. Functional disparities within the TIMP family in cancer: Hints from molecular divergence. Cancer Metastasis Rev. 2019, 38, 469–481. [Google Scholar] [CrossRef]
- Cui, N.; Hu, M.; Khalil, R.A. Biochemical and Biological Attributes of Matrix Metalloproteinases. Prog. Mol. Biol. Transl. Sci. 2017, 147, 1–73. [Google Scholar] [CrossRef] [PubMed]
- Harju, A.K.; Bootorabi, F.; Kuuslahti, M.; Supuran, C.T.; Parkkila, S. Carbonic anhydrase III: A neglected isozyme is stepping into the limelight. J. Enzyme Inhib. Med. Chem. 2013, 28, 231–239. [Google Scholar] [CrossRef]
- Chen, J.; Jaeger, K.; Den, Z.; Koch, P.J.; Sundberg, J.P.; Roop, D.R. Mice expressing a mutant Krt75 (K6hf) allele develop hair and nail defects resembling pachyonychia congenita. J. Invest. Dermatol. 2008, 128, 270–279. [Google Scholar] [CrossRef] [PubMed]
- Wie, J.; Bharthur, A.; Wolfgang, M.; Narayanan, V.; Ramsey, K.; Aranda, K.; Zhang, Q.; Zhou, Y.; Ren, D.; C4RCD Research Group. Intellectual disability-associated UNC80 mutations reveal inter-subunit interaction and dendritic function of the NALCN channel complex. Nat. Commun. 2020, 11, 3351. [Google Scholar] [CrossRef] [PubMed]
- Yoshizawa, Y.; Honda, S.; Shibuya, A. Involvement of Fcα/μR (CD351) in autoantibody production. Mol. Immunol. 2014, 57, 216–219. [Google Scholar] [CrossRef]
- Hu, D.G.; Mackenzie, P.I.; McKinnon, R.A.; Meech, R. Genetic polymorphisms of human UDP-glucuronosyltransferase (UGT) genes and cancer risk. Drug Metab. Rev. 2016, 48, 47–69. [Google Scholar] [CrossRef]
- Barbier, O.; Bélanger, A. Inactivation of androgens by UDP-glucuronosyltransferases in the human prostate. Best Pract. Res. Clin. Endocrinol. Metab. 2008, 22, 259–270. [Google Scholar] [CrossRef]
- Chouinard, S.; Barbier, O.; Bélanger, A. UDP-glucuronosyltransferase 2B15 (UGT2B15) and UGT2B17 enzymes are major determinants of the androgen response in prostate cancer LNCaP cells. J. Biol. Chem. 2007, 282, 33466–33474. [Google Scholar] [CrossRef]
- Saadi, H.; Seillier, M.; Carrier, A. The stress protein TP53INP1 plays a tumor suppressive role by regulating metabolic homeostasis. Biochimie 2015, 118, 44–50. [Google Scholar] [CrossRef]
- Avecilla, V.; Doke, M.; Felty, Q. Contribution of Inhibitor of DNA Binding/Differentiation-3 and Endocrine Disrupting Chemicals to Pathophysiological Aspects of Chronic Disease. Biomed. Res. Int. 2017, 2017, 6307109. [Google Scholar] [CrossRef]
- Garreis, F.; Jahn, J.; Wild, K.; Abrar, D.B.; Schicht, M.; Schröder, J.M.; Paulsen, F. Expression and Regulation of S100 Fused-Type Protein Hornerin at the Ocular Surface and Lacrimal Apparatus. Invest. Ophthalmol. Vis. Sci. 2017, 58, 5968–5977. [Google Scholar] [CrossRef]
- Allgöwer, C.; Kretz, A.L.; von Karstedt, S.; Wittau, M.; Henne-Bruns, D.; Lemke, J. Friend or Foe: S100 Proteins in Cancer. Cancers 2020, 12, 2037. [Google Scholar] [CrossRef]
- Chang, Y.T.; Lin, T.P.; Tang, J.T.; Campbell, M.; Luo, Y.L.; Lu, S.Y.; Yang, C.P.; Cheng, T.Y.; Chang, C.H.; Liu, T.T.; et al. HOTAIR is a REST-regulated lncRNA that promotes neuroendocrine differentiation in castration resistant prostate cancer. Cancer Lett. 2018, 433, 43–52. [Google Scholar] [CrossRef]
- Wang, Y.; Shi, J.; Chai, K.; Ying, X.; Zhou, B.P. The Role of Snail in EMT and Tumorigenesis. Curr. Cancer Drug Targets 2013, 13, 963–972. [Google Scholar] [CrossRef] [PubMed]
- Frambach, S.J.C.M.; de Haas, R.; Smeitink, J.A.M.; Rongen, G.A.; Russel, F.G.M.; Schirris, T.J.J. Brothers in Arms: ABCA1- and ABCG1-Mediated Cholesterol Efflux as Promising Targets in Cardiovascular Disease Treatment. Pharmacol. Rev. 2020, 72, 152–190. [Google Scholar] [CrossRef] [PubMed]
- Sharad, S.; Dobi, A.; Srivastava, S.; Srinivasan, A.; Li, H. PMEPA1 Gene Isoforms: A Potential Biomarker and Therapeutic Target in Prostate Cancer. Biomolecules 2020, 10, 1221. [Google Scholar] [CrossRef]
- Martin, S.K.; Kamelgarn, M.; Kyprianou, N. Cytoskeleton targeting value in prostate cancer treatment. Am. J. Clin. Exp. Urol. 2014, 2, 15–26. [Google Scholar] [PubMed]
- Jin, J.K.; Dayyani, F.; Gallick, G.E. Steps in prostate cancer progression that lead to bone metastasis. Int. J. Cancer. 2011, 128, 2545–2561. [Google Scholar] [CrossRef]
- Gong, Y.; Chippada-Venkata, U.D.; Oh, W.K. Roles of matrix metalloproteinases and their natural inhibitors in prostate cancer progression. Cancers 2014, 6, 1298–1327. [Google Scholar] [CrossRef] [PubMed]
- Green, S.M.; Mostaghel, E.A.; Nelson, P.S. Androgen action and metabolism in prostate cancer. Mol. Cell. Endocrinol. 2012, 360, 3–13. [Google Scholar] [CrossRef]
- Ruan, K.; Song, G.; Ouyang, G. Role of hypoxia in the hallmarks of human cancer. J. Cell Biochem. 2009, 107, 1053–1062. [Google Scholar] [CrossRef] [PubMed]
- Ciafrè, S.A.; Niola, F.; Giorda, E.; Farace, M.G.; Caporossi, D. CoCl(2)-simulated hypoxia in skeletal muscle cell lines: Role of free radicals in gene up-regulation and induction of apoptosis. Free Radic. Res. 2007, 41, 391–401. [Google Scholar] [CrossRef] [PubMed]
- Archer, M.; Dogra, N.; Kyprianou, N. Inflammation as a Driver of Prostate Cancer Metastasis and Therapeutic Resistance. Cancers 2020, 12, 2984. [Google Scholar] [CrossRef]
- Kuroda, E.; Yamashita, U. Mechanisms of enhanced macrophage-mediated prostaglandin E2 production and its suppressive role in Th1 activation in Th2-dominant BALB/c mice. J. Immunol. 2003, 170, 757–764. [Google Scholar] [CrossRef] [PubMed]
- Pirtskhalaishvili, G.; Nelson, J.B. Endothelium-derived factors as paracrine mediators of prostate cancer progression. Prostate 2000, 44, 77–87. [Google Scholar] [CrossRef]
- Voronov, E.; Shouval, D.S.; Krelin, Y.; Cagnano, E.; Benharroch, D.; Iwakura, Y.; Dinarello, C.A.; Apte, R.N. IL-1 is required for tumor invasiveness and angiogenesis. Proc. Natl. Acad. Sci. USA 2003, 100, 2645–2650. [Google Scholar] [CrossRef] [PubMed]
- Dinarello, C.A. Overview of the IL-1 family in innate inflammation and acquired immunity. Immunol. Rev. 2018, 281, 8–27. [Google Scholar] [CrossRef] [PubMed]
- Zhang, A.; Zhang, J.; Kaipainen, A.; Lucas, J.M.; Yang, H. Long non-coding RNA: A newly deciphered “code” in prostate cancer. Cancer Lett. 2016, 375, 323–330. [Google Scholar] [CrossRef] [PubMed]
- Bock, K.W. Aryl hydrocarbon receptor (AHR) functions: Balancing opposing processes including inflammatory reactions. Biochem. Pharmacol. 2020, 178, 114093. [Google Scholar] [CrossRef] [PubMed]
- Effner, R.; Hiller, J.; Eyerich, S.; Traidl-Hoffmann, C.; Brockow, K.; Triggiani, M.; Behrendt, H.; Schmidt-Weber, C.B.; Buters, J.T. Cytochrome P450s in human immune cells regulate IL-22 and c-Kit via an AHR feedback loop. Sci. Rep. 2017, 7, 44005. [Google Scholar] [CrossRef] [PubMed]
- Nagai, M.A. Pleckstrin homology-like domain, family A, member 1 (PHLDA1) and cancer. Biomed. Rep. 2016, 4, 275–281. [Google Scholar] [CrossRef] [PubMed]
- Rabinovitz, I.; Nagle, R.B.; Cress, A.E. Integrin alpha 6 expression in human prostate carcinoma cells is associated with a migratory and invasive phenotype in vitro and in vivo. Clin. Exp. Metastasis 1995, 13, 481–491. [Google Scholar] [CrossRef] [PubMed]
- Denard, B.; Seemann, J.; Chen, Q.; Gay, A.; Huang, H.; Chen, Y.; Ye, J. The membrane-bound transcription factor CREB3L1 is activated in response to virus infection to inhibit proliferation of virus-infected cells. Cell Host Microbe 2011, 10, 65–74. [Google Scholar] [CrossRef] [PubMed]
- Ligresti, G.; Aplin, A.C.; Dunn, B.E.; Morishita, A.; Nicosia, R.F. The acute phase reactant orosomucoid-1 is a bimodal regulator of angiogenesis with time- and context-dependent inhibitory and stimulatory properties. PLoS ONE 2012, 7, e41387. [Google Scholar] [CrossRef]
- Porez, G.; Gross, B.; Prawitt, J.; Gheeraert, C.; Berrabah, W.; Alexandre, J.; Staels, B.; Lefebvre, P. The hepatic orosomucoid/α1-acid glycoprotein gene cluster is regulated by the nuclear bile acid receptor FXR. Endocrinology 2013, 154, 3690–3701. [Google Scholar] [CrossRef] [PubMed]
- Yang, X.; Zhao, Q.; Zhu, L.; Zhang, W. The three complementarity-determining region-like loops in the second extracellular domain of human Fc alpha/mu receptor contribute to its binding of IgA and IgM. Immunobiology 2013, 218, 798–809. [Google Scholar] [CrossRef]
- Ouchida, R.; Mori, H.; Hase, K.; Takatsu, H.; Kurosaki, T.; Tokuhisa, T.; Ohno, H.; Wang, J.Y. Critical role of the IgM Fc receptor in IgM homeostasis, B-cell survival, and humoral immune responses. Proc. Natl. Acad. Sci. USA 2012, 109, E2699–E2706. [Google Scholar] [CrossRef] [PubMed]
- Dagar, M.; Singh, J.P.; Dagar, G.; Tyagi, R.K.; Bagchi, G. Phosphorylation of HSP90 by protein kinase A is essential for the nuclear translocation of androgen receptor. J. Biol. Chem. 2019, 294, 8699–8710. [Google Scholar] [CrossRef]
- Savoy, R.M.; Chen, L.; Siddiqui, S.; Melgoza, F.U.; Durbin-Johnson, B.; Drake, C.; Jathal, M.K.; Bose, S.; Steele, T.M.; Mooso, B.A.; et al. Transcription of Nrdp1 by the androgen receptor is regulated by nuclear filamin A in prostate cancer. Endocr. Relat. Cancer 2015, 22, 369–386. [Google Scholar] [CrossRef]
- Federer-Gsponer, J.R.; Quintavalle, C.; Müller, D.C.; Dietsche, T.; Perrina, V.; Lorber, T.; Juskevicius, D.; Lenkiewicz, E.; Zellweger, T.; Gasser, T.; et al. Delineation of human prostate cancer evolution identifies chromothripsis as a polyclonal event and FKBP4 as a potential driver of castration resistance. J. Pathol. 2018, 245, 74–84. [Google Scholar] [CrossRef]
- Zhong, W.; Zhu, H.; Sheng, F.; Tian, Y.; Zhou, J.; Chen, Y.; Li, S.; Lin, J. Activation of the MAPK11/12/13/14 (p38 MAPK) pathway regulates the transcription of autophagy genes in response to oxidative stress induced by a novel copper complex in HeLa cells. Autophagy 2014, 10, 1285–1300. [Google Scholar] [CrossRef] [PubMed]
- Van Beijnum, J.R.; Dings, R.P.; van der Linden, E.; Zwaans, B.M.; Ramaekers, F.C.; Mayo, K.H.; Griffioen, A.W. Gene expression of tumor angiogenesis dissected: Specific targeting of colon cancer angiogenic vasculature. Blood 2006, 108, 2339–2348. [Google Scholar] [CrossRef]
- Sánchez-Martín, L.; Estecha, A.; Samaniego, R.; Sánchez-Ramón, S.; Vega, M.Á.; Sánchez-Mateos, P. The chemokine CXCL12 regulates monocyte-macrophage differentiation and RUNX3 expression. Blood 2011, 117, 88–97. [Google Scholar] [CrossRef]
- Rigo, A.; Gottardi, M.; Zamò, A.; Mauri, P.; Bonifacio, M.; Krampera, M.; Damiani, E.; Pizzolo, G.; Vinante, F. Macrophages may promote cancer growth via a GM-CSF/HB-EGF paracrine loop that is enhanced by CXCL12. Mol. Cancer 2010, 9, 273. [Google Scholar] [CrossRef] [PubMed]
- Ray, P.; Stacer, A.C.; Fenner, J.; Cavnar, S.P.; Meguiar, K.; Brown, M.; Luker, K.E.; Luker, G.D. CXCL12-γ in primary tumors drives breast cancer metastasis. Oncogene 2015, 34, 2043–2051. [Google Scholar] [CrossRef]
- Sun, Y.X.; Wang, J.; Shelburne, C.E.; Lopatin, D.E.; Chinnaiyan, A.M.; Rubin, M.A.; Pienta, K.J.; Taichman, R.S. Expression of CXCR4 and CXCL12 (SDF-1) in human prostate cancers (PCa) in vivo. J. Cell Biochem. 2003, 89, 462–473. [Google Scholar] [CrossRef] [PubMed]
- Roy, I.; Zimmerman, N.P.; Mackinnon, A.C.; Tsai, S.; Evans, D.B.; Dwinell, M.B. CXCL12 chemokine expression suppresses human pancreatic cancer growth and metastasis. PLoS ONE 2014, 9, e90400. [Google Scholar] [CrossRef]
- Singh, S.; Singh, U.P.; Grizzle, W.E.; Lillard, J.W., Jr. CXCL12-CXCR4 interactions modulate prostate cancer cell migration, metalloproteinase expression and invasion. Lab. Invest. 2004, 84, 1666–1676. [Google Scholar] [CrossRef] [PubMed]
- Chinni, S.; Sivalogan, S.; Dong, Z.; Filho, J.; Deng, X.; Bonfil, R.; Cher, M. CXCL12/CXCR4 signaling activates Akt-1 and MMP-9 expression in prostate cancer cells: The role of bone microenvironment-associated CXCL12. Prostate 2006, 66, 32–48. [Google Scholar] [CrossRef] [PubMed]
- Brady, L.; Gil da Costa, R.M.; Coleman, I.M.; Matson, C.K.; Risk, M.C.; Coleman, R.T.; Nelson, P.S. A comparison of prostate cancer cell transcriptomes in 2D monoculture vs 3D xenografts identify consistent gene expression alterations associated with tumor microenvironments. Prostate 2020, 80, 491–499. [Google Scholar] [CrossRef] [PubMed]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| A. Upregulated genes | |||
| Symbol | Entrez Gene Name | Gene Symbol–Human (HUGO/HGNC/Entrez Gene) | Fold Change |
| F3 | coagulation factor III, tissue factor | F3 | 210.043 |
| CREB3L1 | cAMP responsive element binding protein 3 like 1 | CREB3L1 | 182.705 |
| ORM1 | orosomucoid 1 | ORM1 | 174.684 |
| RGS2 | regulator of G protein signaling 2 | RGS2 | 138.156 |
| TIMP1 | TIMP metallopeptidase inhibitor 1 | TIMP1 | 132.481 |
| COL12A1 | collagen type XII alpha 1 chain | COL12A1 | 110.37 |
| MAGEB17 | MAGE family member B17 | MAGEB17 | 77.306 |
| UNC80 | unc-80 homolog, NALCN channel complex subunit | C2orf21 | 75.717 |
| CA3 | carbonic anhydrase 3 | CA3 | 74.589 |
| KRT75 | keratin 75 | KRT75 | 74.395 |
| B. Downregulated genes | |||
| Symbol | Entrez Gene Name | Gene Symbol–Human (HUGO/HGNC/Entrez Gene) | Fold Change |
| S100A7 | S100 calcium binding protein A7 | S100A7 | −100.404 |
| FCAMR | Fc fragment of IgA and IgM receptor | FCAMR | −62.565 |
| UGT2B15 | UDP glucuronosyltransferase family 2 member B15 | UGT2B15 | −62.323 |
| UGT2B17 | UDP glucuronosyltransferase family 2 member B17 | UGT2B17 | −54.141 |
| GPX8 | glutathione peroxidase 8 (putative) | GPX8 | −35.5 |
| UGT2B10 | UDP glucuronosyltransferase family 2 member B10 | UGT2B10 | −35.444 |
| TP53INP1 | tumor protein p53 inducible nuclear protein 1 | TP53INP1 | −31.76 |
| ID3 | inhibitor of DNA binding 3, HLH protein | ID3 | −21.13 |
| UGT2B28 | UDP glucuronosyltransferase family 2 member B28 | UGT2B28 | −20.846 |
| HRNR | hornerin | HRNR | −20.03 |
| A. Top Pathways. | ||||
| Ingenuity Canonical Pathways | −log (p-value) | p-Value | Ratio | z-Score |
| Aryl Hydrocarbon Receptor Signaling | 4.79 | 0.0000162181 | 0.153 | −1.155 |
| AMPK Signaling | 3.28 | 0.000524807 | 0.114 | 1 |
| HOTAIR Regulatory Pathway | 2.67 | 0.002137962 | 0.115 | 1.414 |
| p53 Signaling | 2.6 | 0.002511886 | 0.133 | −0.577 |
| Sperm Motility | 2.57 | 0.002691535 | 0.104 | 0 |
| Sirtuin Signaling Pathway | 2.47 | 0.003388442 | 0.0954 | −0.853 |
| B. Top upregulated pathways | ||||
| Ingenuity Canonical Pathways | −log (p-Value) | p-Value | Ratio | z-Score |
| AMPK Signaling | 3.28 | 0.000524807 | 0.114 | 1 |
| PTEN Signaling | 1.7 | 0.019952623 | 0.103 | 1.387 |
| HOTAIR Regulatory Pathway | 2.67 | 0.002137962 | 0.115 | 1.414 |
| LXR/RXR Activation | 1.84 | 0.014454398 | 0.107 | 1.667 |
| Protein Kinase A Signaling | 1.74 | 0.018197009 | 0.0809 | 1.732 |
| C. Top downregulated pathways | ||||
| Ingenuity Canonical Pathways | −log (p-value) | p-Value | Ratio | z-Score |
| Cardiac Hypertrophy Signaling | 1.75 | 0.017782794 | 0.0894 | −2.683 |
| Glioblastoma Multiforme Signaling | 1.5 | 0.031622777 | 0.0926 | −2.138 |
| Acute Phase Response Signaling | 1.47 | 0.033884416 | 0.0899 | −2.53 |
| Actin Nucleation by ARP–WASP Complex | 1.44 | 0.036307805 | 0.114 | −2.646 |
| Cardiac Hypertrophy Signaling (Enhanced) | 1.36 | 0.043651583 | 0.0737 | −2.744 |
| GNRH Signaling | 1.35 | 0.044668359 | 0.0882 | −2.309 |
| Name | p-Value Range | Number of Molecules |
|---|---|---|
| Cellular Movement | 2.37 × 10−3–1.00 × 10−11 | 194 |
| Cellular Development | 1.87 × 10−3–1.10 × 10−9 | 188 |
| Cellular Growth and Proliferation | 2.96 × 10−3–1.10 × 10−9 | 174 |
| Cell Death and Survival | 2.84 × 10−3–1.67 × 10−7 | 180 |
| Cellular Assembly and Organization | 1.16 × 10−3–7.16 × 10−7 | 119 |
| Gene_ID | Symbol | Expr Fold Change | p-Value |
|---|---|---|---|
| ENSG00000107562 | CXCL12 | 26.703 | 9.61 × 10−36 |
| ENSG00000144476 | CXCR7 | −4.1208 | 1.25 × 10−6 |
| GENE_ID | Symbol | Entrez Gene Name | Fold Change | p-Value |
|---|---|---|---|---|
| ENSG00000164171 | ITGA2 | integrin subunit alpha 2 | −6.122550677 | 6.29843 × 10−20 |
| ENSG00000091409 | ITGA6 | integrin subunit alpha 6 | −2.749811242 | 4.22149 × 10−14 |
| ENSG00000150093 | ITGB1 | integrin subunit beta 1 | −1.669372972 | 1.92104 × 10−5 |
| ENSG00000090339 | ICAM1 | intercellular adhesion molecule 1 | 5.478055202 | 6.03337 × 10−7 |
| ENSG00000105376 | ICAM5 | intercellular adhesion molecule 5 | −5.772935224 | 7.54024 × 10−8 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yu, L.; Li, R.W.; Huang, H.; Pham, Q.; Yu, L.; Wang, T.T.Y. Transcriptomic Analysis of LNCaP Tumor Xenograft to Elucidate the Components and Mechanisms Contributed by Tumor Environment as Targets for Dietary Prostate Cancer Prevention Studies. Nutrients 2021, 13, 1000. https://doi.org/10.3390/nu13031000
Yu L, Li RW, Huang H, Pham Q, Yu L, Wang TTY. Transcriptomic Analysis of LNCaP Tumor Xenograft to Elucidate the Components and Mechanisms Contributed by Tumor Environment as Targets for Dietary Prostate Cancer Prevention Studies. Nutrients. 2021; 13(3):1000. https://doi.org/10.3390/nu13031000
Chicago/Turabian StyleYu, Lu, Robert W. Li, Haiqiu Huang, Quynhchi Pham, Liangli Yu, and Thomas T. Y. Wang. 2021. "Transcriptomic Analysis of LNCaP Tumor Xenograft to Elucidate the Components and Mechanisms Contributed by Tumor Environment as Targets for Dietary Prostate Cancer Prevention Studies" Nutrients 13, no. 3: 1000. https://doi.org/10.3390/nu13031000
APA StyleYu, L., Li, R. W., Huang, H., Pham, Q., Yu, L., & Wang, T. T. Y. (2021). Transcriptomic Analysis of LNCaP Tumor Xenograft to Elucidate the Components and Mechanisms Contributed by Tumor Environment as Targets for Dietary Prostate Cancer Prevention Studies. Nutrients, 13(3), 1000. https://doi.org/10.3390/nu13031000