The Melanocortin System behind the Dysfunctional Eating Behaviors

 ,
,  , ,
, ,  ,
,  and
and
Abstract
1. Introduction
2. An Overview of the Melanocortin Receptors in the Control of Food Intake
3. Melanocortin Receptors in Feeding
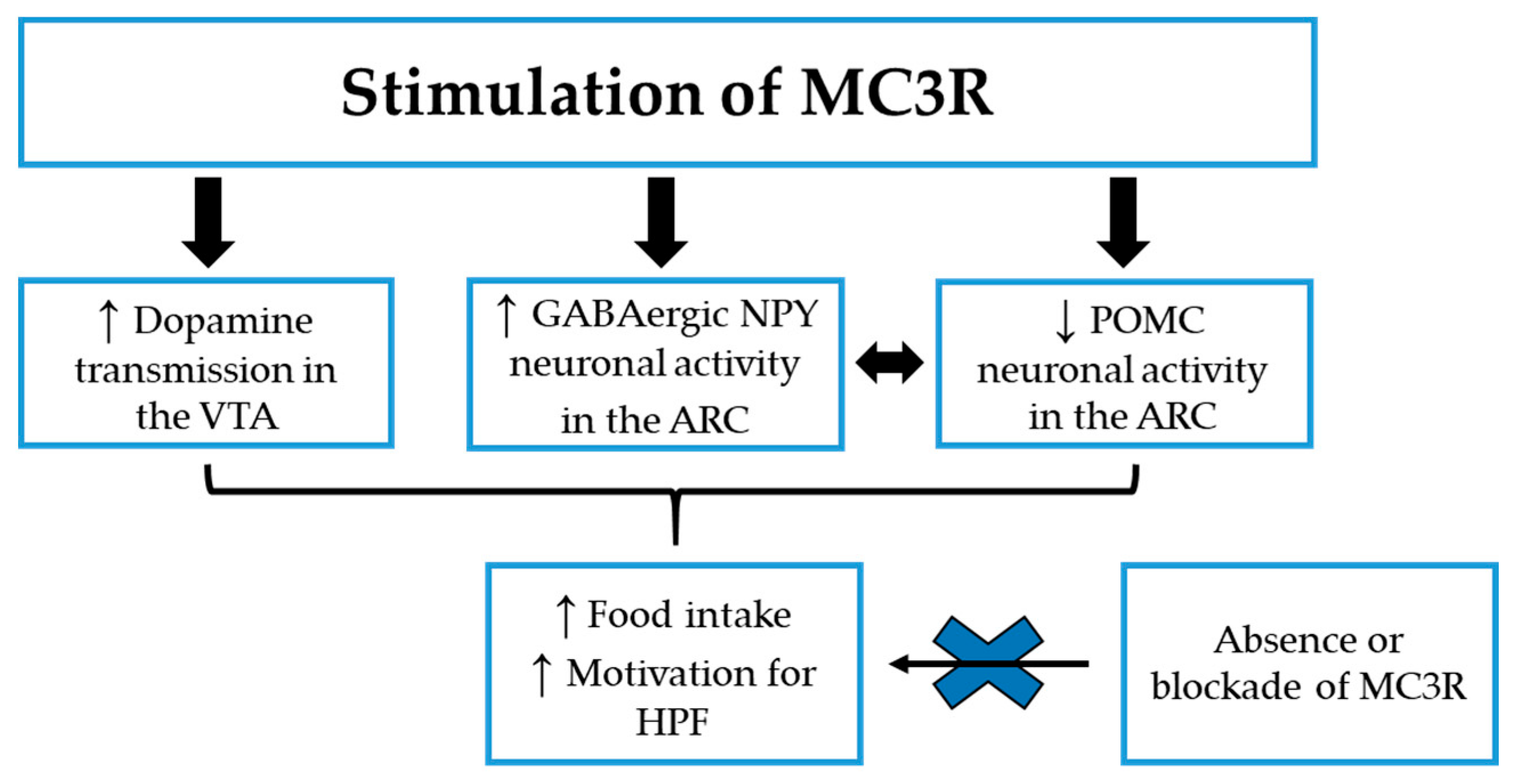
3.1. MC3R
3.1.1. MC3R: Preclinical Studies on Eating Behavior
3.1.2. MC3R: Preclinical Studies in Food Reward
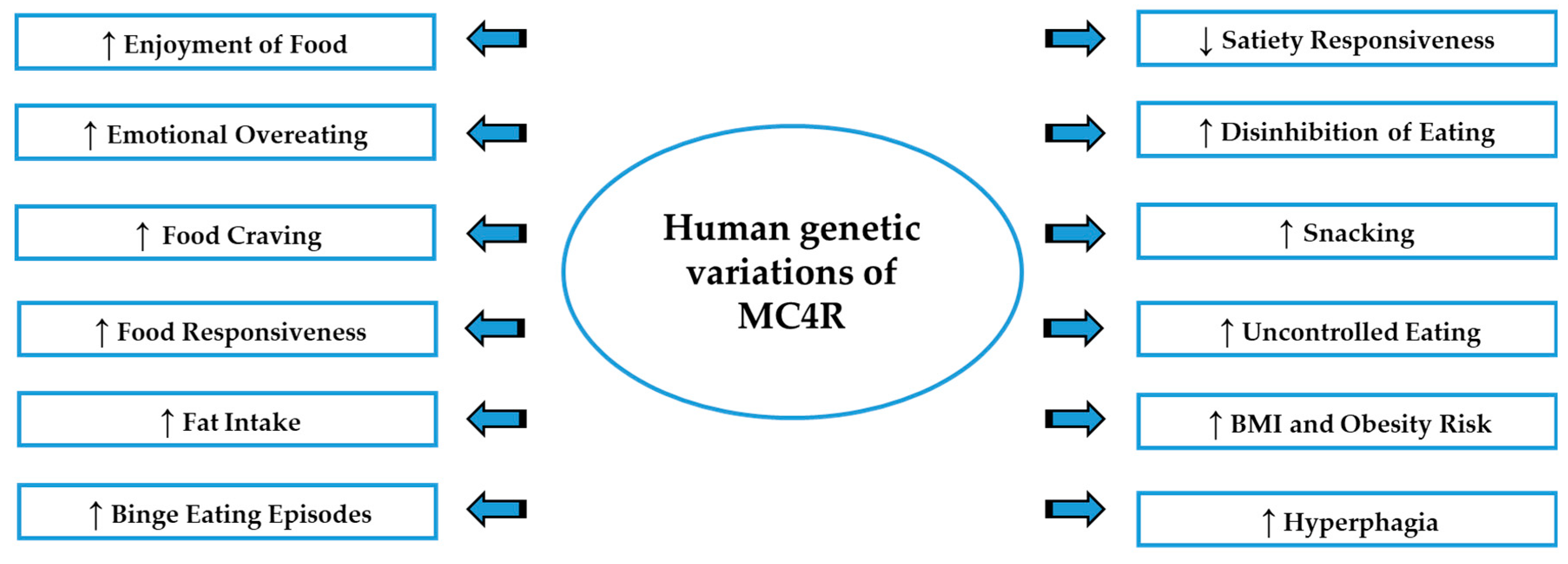
3.2. MC4R
3.2.1. MC4R: Preclinical Studies on Food Preference and Motivation
3.2.2. Clinical Studies on MC4R Mutations
3.2.3. The Polymorphism rs17782313 Nearby MC4R Gene and Eating Behavior
4. Melanocortin System and Stress Responses
5. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Cordain, L.; Eaton, S.B.; Sebastian, A.; Mann, N.; Lindeberg, S.; Watkins, B.A.; O’Keefe, J.H.; Brand-Miller, J. Origins and evolution of the Western diet: Health implications for the 21st century. Am. J. Clin. Nutr. 2005, 81, 341–354. [Google Scholar] [CrossRef] [PubMed]
- Giudetti, A.M.; Micioni Di Bonaventura, M.V.; Ferramosca, A.; Longo, S.; Micioni Di Bonaventura, E.; Friuli, M.; Romano, A.; Gaetani, S.; Cifani, C. Brief daily access to cafeteria-style diet impairs hepatic metabolism even in the absence of excessive body weight gain in rats. FASEB J. 2020, 34, 9358–9371. [Google Scholar] [CrossRef] [PubMed]
- Cone, R.D. The Central Melanocortin System and Energy Homeostasis. Trends Endocrinol. Metab. 1999, 10, 211–216. [Google Scholar] [CrossRef]
- Cone, R.D. Studies on the physiological functions of the melanocortin system. Endocr. Rev. 2006, 27, 736–749. [Google Scholar] [CrossRef] [PubMed]
- Girardet, C.; Butler, A.A. Neural melanocortin receptors in obesity and related metabolic disorders. Biochim. Biophys. Acta 2014, 1842, 482–494. [Google Scholar] [CrossRef] [PubMed]
- Butler, A.A. The melanocortin system and energy balance. Peptides 2006, 27, 281–290. [Google Scholar] [CrossRef]
- Fan, W.; Boston, B.A.; Kesterson, R.A.; Hruby, V.J.; Cone, R.D. Role of melanocortinergic neurons in feeding and the agouti obesity syndrome. Nature 1997, 385, 165–168. [Google Scholar] [CrossRef]
- Beckers, S.; Zegers, D.; de Freitas, F.; Mertens, I.L.; Van Gaal, L.F.; Van Hul, W. Association study of MC4R with complex obesity and replication of the rs17782313 association signal. Mol. Genet. Metab. 2011, 103, 71–75. [Google Scholar] [CrossRef]
- Benoit, S.; Schwartz, M.; Baskin, D.; Woods, S.C.; Seeley, R.J. CNS melanocortin system involvement in the regulation of food intake. Horm. Behav. 2000, 37, 299–305. [Google Scholar] [CrossRef]
- Hinney, A.; Volckmar, A.L.; Knoll, N. Melanocortin-4 receptor in energy homeostasis and obesity pathogenesis. Prog. Mol. Biol. Transl. Sci. 2013, 114, 147–191. [Google Scholar] [CrossRef]
- Huszar, D.; Lynch, C.A.; Fairchild-Huntress, V.; Dunmore, J.H.; Fang, Q.; Berkemeier, L.R.; Gu, W.; Kesterson, R.A.; Boston, B.A.; Cone, R.D.; et al. Targeted disruption of the melanocortin-4 receptor results in obesity in mice. Cell 1997, 88, 131–141. [Google Scholar] [CrossRef]
- Kleinendorst, L.; van Haelst, M.M.; van den Akker, E.L.T. Genetics of Obesity. Exp. Suppl. 2019, 111, 419–441. [Google Scholar] [CrossRef] [PubMed]
- Vaisse, C.; Clement, K.; Guy-Grand, B.; Froguel, P. A frameshift mutation in human MC4R is associated with a dominant form of obesity. Nat. Genet. 1998, 20, 113–114. [Google Scholar] [CrossRef] [PubMed]
- Yeo, G.S.; Farooqi, I.S.; Aminian, S.; Halsall, D.J.; Stanhope, R.G.; O’Rahilly, S. A frameshift mutation in MC4R associated with dominantly inherited human obesity. Nat. Genet. 1998, 20, 111–112. [Google Scholar] [CrossRef]
- Branson, R.; Potoczna, N.; Kral, J.G.; Lentes, K.U.; Hoehe, M.R.; Horber, F.F. Binge eating as a major phenotype of melanocortin 4 receptor gene mutations. N. Engl. J. Med. 2003, 348, 1096–1103. [Google Scholar] [CrossRef]
- Butler, A.A.; Marks, D.L.; Fan, W.; Kuhn, C.M.; Bartolome, M.; Cone, R.D. Melanocortin-4 receptor is required for acute homeostatic responses to increased dietary fat. Nat. Neurosci. 2001, 4, 605–611. [Google Scholar] [CrossRef]
- Ho-Urriola, J.; Guzman-Guzman, I.P.; Smalley, S.V.; Gonzalez, A.; Weisstaub, G.; Dominguez-Vasquez, P.; Valladares, M.; Amador, P.; Hodgson, M.I.; Obregon, A.M.; et al. Melanocortin-4 receptor polymorphism rs17782313: Association with obesity and eating in the absence of hunger in Chilean children. Nutrition 2014, 30, 145–149. [Google Scholar] [CrossRef]
- Liu, J.; Garza, J.C.; Li, W.; Lu, X.Y. Melanocortin-4 receptor in the medial amygdala regulates emotional stress-induced anxiety-like behaviour, anorexia and corticosterone secretion. Int. J. Neuropsychopharmacol. 2013, 16, 105–120. [Google Scholar] [CrossRef]
- Santos, J.L.; Amador, P.; Valladares, M.; Albala, C.; Martinez, J.A.; Marti, A. Obesity and eating behaviour in a three-generation Chilean family with carriers of the Thrl50Ile mutation in the melanocortin-4 receptor gene. J. Physiol. Biochem. 2008, 64, 205–210. [Google Scholar] [CrossRef]
- Vega, J.A.; Salazar, G.; Hodgson, M.I.; Cataldo, L.R.; Valladares, M.; Obregon, A.M.; Santos, J.L. Melanocortin-4 Receptor Gene Variation Is Associated with Eating Behavior in Chilean Adults. Ann. Nutr. Metab. 2016, 68, 35–41. [Google Scholar] [CrossRef]
- Mavrikaki, M.; Girardet, C.; Kern, A.; Faruzzi Brantley, A.; Miller, C.A.; Macarthur, H.; Marks, D.L.; Butler, A.A. Melanocortin-3 receptors in the limbic system mediate feeding-related motivational responses during weight loss. Mol. Metab. 2016, 5, 566–579. [Google Scholar] [CrossRef] [PubMed]
- Pandit, R.; Omrani, A.; Luijendijk, M.C.; de Vrind, V.A.; Van Rozen, A.J.; Ophuis, R.J.; Garner, K.; Kallo, I.; Ghanem, A.; Liposits, Z.; et al. Melanocortin 3 Receptor Signaling in Midbrain Dopamine Neurons Increases the Motivation for Food Reward. Neuropsychopharmacology 2016, 41, 2241–2251. [Google Scholar] [CrossRef] [PubMed]
- Pandit, R.; van der Zwaal, E.M.; Luijendijk, M.C.; Brans, M.A.; van Rozen, A.J.; Oude Ophuis, R.J.; Vanderschuren, L.J.; Adan, R.A.; la Fleur, S.E. Central melanocortins regulate the motivation for sucrose reward. PLoS ONE 2015, 10, e0121768. [Google Scholar] [CrossRef]
- De Franca, G.V.; Gigante, D.P.; Olinto, M.T. Binge eating in adults: Prevalence and association with obesity, poor self-rated health status and body dissatisfaction. Public Health Nutr. 2014, 17, 932–938. [Google Scholar] [CrossRef]
- Hudson, J.I.; Hiripi, E.; Pope, H.G., Jr.; Kessler, R.C. The prevalence and correlates of eating disorders in the National Comorbidity Survey Replication. Biol. Psychiatry 2007, 61, 348–358. [Google Scholar] [CrossRef] [PubMed]
- Kessler, R.C.; Berglund, P.A.; Chiu, W.T.; Deitz, A.C.; Hudson, J.I.; Shahly, V.; Aguilar-Gaxiola, S.; Alonso, J.; Angermeyer, M.C.; Benjet, C.; et al. The prevalence and correlates of binge eating disorder in the World Health Organization World Mental Health Surveys. Biol. Psychiatry 2013, 73, 904–914. [Google Scholar] [CrossRef]
- Kishi, T.; Aschkenasi, C.J.; Lee, C.E.; Mountjoy, K.G.; Saper, C.B.; Elmquist, J.K. Expression of melanocortin 4 receptor mRNA in the central nervous system of the rat. J. Comp. Neurol. 2003, 457, 213–235. [Google Scholar] [CrossRef]
- Yoon, Y.R.; Baik, J.H. Melanocortin 4 Receptor and Dopamine D2 Receptor Expression in Brain Areas Involved in Food Intake. Endocrinol. Metab. 2015, 30, 576–583. [Google Scholar] [CrossRef] [PubMed]
- American Psychiatric Association. Diagnostic and Statistical Manual of Mental Disorders: DSM-5; American Psychiatric Association: Arlington, VA, USA, 2013. [Google Scholar]
- Guerdjikova, A.I.; Mori, N.; Casuto, L.S.; McElroy, S.L. Update on Binge Eating Disorder. Med. Clin. N. Am. 2019, 103, 669–680. [Google Scholar] [CrossRef]
- Ulfvebrand, S.; Birgegard, A.; Norring, C.; Hogdahl, L.; von Hausswolff-Juhlin, Y. Psychiatric comorbidity in women and men with eating disorders results from a large clinical database. Psychiatry Res. 2015, 230, 294–299. [Google Scholar] [CrossRef] [PubMed]
- Yanovski, S.Z. Binge eating disorder: Current knowledge and future directions. Obes. Res. 1993, 1, 306–324. [Google Scholar] [CrossRef] [PubMed]
- Schag, K.; Schonleber, J.; Teufel, M.; Zipfel, S.; Giel, K.E. Food-related impulsivity in obesity and binge eating disorder—A systematic review. Obes. Rev. 2013, 14, 477–495. [Google Scholar] [CrossRef] [PubMed]
- Manwaring, J.L.; Green, L.; Myerson, J.; Strube, M.J.; Wilfley, D.E. Discounting of Various types of rewards by women with and without binge eating Disorder: Evidence for general rather than specific Differences. Psychol. Rec. 2011, 61, 561–582. [Google Scholar] [CrossRef] [PubMed]
- Moore, C.F.; Sabino, V.; Koob, G.F.; Cottone, P. Neuroscience of Compulsive Eating Behavior. Front. Neurosci. 2017, 11, 469. [Google Scholar] [CrossRef]
- Zeeck, A.; Stelzer, N.; Linster, H.W.; Joos, A.; Hartmann, A. Emotion and eating in binge eating disorder and obesity. Eur. Eat. Disord. Rev. 2011, 19, 426–437. [Google Scholar] [CrossRef]
- Adam, T.C.; Epel, E.S. Stress, eating and the reward system. Physiol. Behav. 2007, 91, 449–458. [Google Scholar] [CrossRef]
- Micioni Di Bonaventura, M.V.; Micioni Di Bonaventura, E.; Polidori, C.; Cifani, C. Preclinical Models of Stress and Environmental Influences on Binge Eating. In Binge Eating: A Transdiagnostic Psychopathology; Frank, G.K.W., Berner, L.A., Eds.; Springer International Publishing: Cham, Switzerland, 2020; pp. 85–101. [Google Scholar] [CrossRef]
- Rosenbaum, D.L.; White, K.S. The relation of anxiety, depression, and stress to binge eating behavior. J. Health Psychol. 2015, 20, 887–898. [Google Scholar] [CrossRef]
- He, J.; Cai, Z.; Fan, X. Prevalence of binge and loss of control eating among children and adolescents with overweight and obesity: An exploratory meta-analysis. Int. J. Eat. Disord. 2017, 50, 91–103. [Google Scholar] [CrossRef]
- Bagnol, D.; Lu, X.Y.; Kaelin, C.B.; Day, H.E.; Ollmann, M.; Gantz, I.; Akil, H.; Barsh, G.S.; Watson, S.J. Anatomy of an endogenous antagonist: Relationship between Agouti-related protein and proopiomelanocortin in brain. J. Neurosci. 1999, 19, RC26. [Google Scholar] [CrossRef]
- Ollmann, M.M.; Wilson, B.D.; Yang, Y.K.; Kerns, J.A.; Chen, Y.; Gantz, I.; Barsh, G.S. Antagonism of central melanocortin receptors in vitro and in vivo by agouti-related protein. Science 1997, 278, 135–138. [Google Scholar] [CrossRef]
- Gantz, I.; Konda, Y.; Tashiro, T.; Shimoto, Y.; Miwa, H.; Munzert, G.; Watson, S.J.; DelValle, J.; Yamada, T. Molecular cloning of a novel melanocortin receptor. J. Biol. Chem. 1993, 268, 8246–8250. [Google Scholar] [PubMed]
- Gantz, I.; Miwa, H.; Konda, Y.; Shimoto, Y.; Tashiro, T.; Watson, S.J.; DelValle, J.; Yamada, T. Molecular cloning, expression, and gene localization of a fourth melanocortin receptor. J. Biol. Chem. 1993, 268, 15174–15179. [Google Scholar] [PubMed]
- Mountjoy, K.G.; Robbins, L.S.; Mortrud, M.T.; Cone, R.D. The cloning of a family of genes that encode the melanocortin receptors. Science 1992, 257, 1248–1251. [Google Scholar] [CrossRef] [PubMed]
- Roselli-Rehfuss, L.; Mountjoy, K.G.; Robbins, L.S.; Mortrud, M.T.; Low, M.J.; Tatro, J.B.; Entwistle, M.L.; Simerly, R.B.; Cone, R.D. Identification of a receptor for gamma melanotropin and other proopiomelanocortin peptides in the hypothalamus and limbic system. Proc. Natl. Acad. Sci. USA 1993, 90, 8856–8860. [Google Scholar] [CrossRef]
- Adan, R.A.; Cone, R.D.; Burbach, J.P.; Gispen, W.H. Differential effects of melanocortin peptides on neural melanocortin receptors. Mol. Pharmacol. 1994, 46, 1182–1190. [Google Scholar]
- Mountjoy, K.G. Distribution and function of melanocortin receptors within the brain. Adv. Exp. Med. Biol. 2010, 681, 29–48. [Google Scholar] [CrossRef]
- Jegou, S.; Boutelet, I.; Vaudry, H. Melanocortin-3 receptor mRNA expression in pro-opiomelanocortin neurones of the rat arcuate nucleus. J. Neuroendocrinol. 2000, 12, 501–505. [Google Scholar] [CrossRef]
- Mountjoy, K.G.; Mortrud, M.T.; Low, M.J.; Simerly, R.B.; Cone, R.D. Localization of the melanocortin-4 receptor (MC4-R) in neuroendocrine and autonomic control circuits in the brain. Mol. Endocrinol. 1994, 8, 1298–1308. [Google Scholar] [CrossRef]
- Mountjoy, K.G.; Wild, J.M. Melanocortin-4 receptor mRNA expression in the developing autonomic and central nervous systems. Dev. Brain Res. 1998, 107, 309–314. [Google Scholar] [CrossRef]
- Small, C.J.; Kim, M.S.; Stanley, S.A.; Mitchell, J.R.; Murphy, K.; Morgan, D.G.; Ghatei, M.A.; Bloom, S.R. Effects of chronic central nervous system administration of agouti-related protein in pair-fed animals. Diabetes 2001, 50, 248–254. [Google Scholar] [CrossRef]
- Beckers, S.; Zegers, D.; Van Gaal, L.F.; Van Hul, W. The role of the leptin-melanocortin signalling pathway in the control of food intake. Crit. Rev. Eukaryot. Gene Expr. 2009, 19, 267–287. [Google Scholar] [CrossRef] [PubMed]
- Fan, W.; Dinulescu, D.M.; Butler, A.A.; Zhou, J.; Marks, D.L.; Cone, R.D. The central melanocortin system can directly regulate serum insulin levels. Endocrinology 2000, 141, 3072–3079. [Google Scholar] [CrossRef] [PubMed]
- Williams, K.W.; Scott, M.M.; Elmquist, J.K. Modulation of the central melanocortin system by leptin, insulin, and serotonin: Co-ordinated actions in a dispersed neuronal network. Eur. J. Pharmacol. 2011, 660, 2–12. [Google Scholar] [CrossRef] [PubMed]
- Micioni Di Bonaventura, M.V.; Micioni Di Bonaventura, E.; Cifani, C.; Polidori, C. N/OFQ-NOP System in Food Intake. Handb. Exp. Pharmacol. 2019, 254, 279–295. [Google Scholar] [CrossRef] [PubMed]
- Cifani, C.; Guerrini, R.; Massi, M.; Polidori, C. Chronic intracerebroventricular infusion of nociceptin/orphanin FQ increases food and ethanol intake in alcohol-preferring rats. Peptides 2006, 27, 2803–2810. [Google Scholar] [CrossRef] [PubMed]
- Bomberg, E.M.; Grace, M.K.; Levine, A.S.; Olszewski, P.K. Functional interaction between nociceptin/orphanin FQ and alpha-melanocyte-stimulating hormone in the regulation of feeding. Peptides 2006, 27, 1827–1834. [Google Scholar] [CrossRef] [PubMed]
- Filaferro, M.; Ruggieri, V.; Novi, C.; Calo, G.; Cifani, C.; Micioni Di Bonaventura, M.V.; Sandrini, M.; Vitale, G. Functional antagonism between nociceptin/orphanin FQ and corticotropin-releasing factor in rat anxiety-related behaviors: Involvement of the serotonergic system. Neuropeptides 2014, 48, 189–197. [Google Scholar] [CrossRef]
- Vitale, G.; Filaferro, M.; Micioni Di Bonaventura, M.V.; Ruggieri, V.; Cifani, C.; Guerrini, R.; Simonato, M.; Zucchini, S. Effects of [Nphe(1), Arg(14), Lys(15)] N/OFQ-NH2 (UFP-101), a potent NOP receptor antagonist, on molecular, cellular and behavioural alterations associated with chronic mild stress. J. Psychopharmacol. 2017, 31, 691–703. [Google Scholar] [CrossRef]
- Hardaway, J.A.; Jensen, J.; Kim, M.; Mazzone, C.M.; Sugam, J.A.; Diberto, J.F.; Lowery-Gionta, E.G.; Hwa, L.S.; Pleil, K.E.; Bulik, C.M.; et al. Nociceptin receptor antagonist SB 612111 decreases high fat diet binge eating. Behav. Brain Res. 2016, 307, 25–34. [Google Scholar] [CrossRef]
- Micioni Di Bonaventura, M.V.; Ubaldi, M.; Liberati, S.; Ciccocioppo, R.; Massi, M.; Cifani, C. Caloric restriction increases the sensitivity to the hyperphagic effect of nociceptin/orphanin FQ limiting its ability to reduce binge eating in female rats. Psychopharmacology 2013, 228, 53–63. [Google Scholar] [CrossRef]
- Pucci, M.; Micioni Di Bonaventura, M.V.; Giusepponi, M.E.; Romano, A.; Filaferro, M.; Maccarrone, M.; Ciccocioppo, R.; Cifani, C.; D’Addario, C. Epigenetic regulation of nociceptin/orphanin FQ and corticotropin-releasing factor system genes in frustration stress-induced binge-like palatable food consumption. Addict. Biol. 2016, 21, 1168–1185. [Google Scholar] [CrossRef]
- Statnick, M.A.; Chen, Y.; Ansonoff, M.; Witkin, J.M.; Rorick-Kehn, L.; Suter, T.M.; Song, M.; Hu, C.; Lafuente, C.; Jimenez, A.; et al. A Novel Nociceptin Receptor Antagonist LY2940094 Inhibits Excessive Feeding Behavior in Rodents: A Possible Mechanism for the Treatment of Binge Eating Disorder. J. Pharmacol. Exp. Ther. 2016, 356, 493–502. [Google Scholar] [CrossRef] [PubMed]
- Butler, A.A.; Kesterson, R.A.; Khong, K.; Cullen, M.J.; Pelleymounter, M.A.; Dekoning, J.; Baetscher, M.; Cone, R.D. A unique metabolic syndrome causes obesity in the melanocortin-3 receptor-deficient mouse. Endocrinology 2000, 141, 3518–3521. [Google Scholar] [CrossRef] [PubMed]
- Chen, A.S.; Marsh, D.J.; Trumbauer, M.E.; Frazier, E.G.; Guan, X.M.; Yu, H.; Rosenblum, C.I.; Vongs, A.; Feng, Y.; Cao, L.; et al. Inactivation of the mouse melanocortin-3 receptor results in increased fat mass and reduced lean body mass. Nat. Genet. 2000, 26, 97–102. [Google Scholar] [CrossRef] [PubMed]
- Ste Marie, L.; Miura, G.I.; Marsh, D.J.; Yagaloff, K.; Palmiter, R.D. A metabolic defect promotes obesity in mice lacking melanocortin-4 receptors. Proc. Natl. Acad. Sci. USA 2000, 97, 12339–12344. [Google Scholar] [CrossRef] [PubMed]
- Sutton, G.M.; Trevaskis, J.L.; Hulver, M.W.; McMillan, R.P.; Markward, N.J.; Babin, M.J.; Meyer, E.A.; Butler, A.A. Diet-genotype interactions in the development of the obese, insulin-resistant phenotype of C57BL/6J mice lacking melanocortin-3 or -4 receptors. Endocrinology 2006, 147, 2183–2196. [Google Scholar] [CrossRef] [PubMed]
- Begriche, K.; Girardet, C.; McDonald, P.; Butler, A.A. Melanocortin-3 receptors and metabolic homeostasis. Prog. Mol. Biol. Transl. Sci. 2013, 114, 109–146. [Google Scholar] [CrossRef] [PubMed]
- Farooqi, I.S.; Keogh, J.M.; Yeo, G.S.; Lank, E.J.; Cheetham, T.; O’Rahilly, S. Clinical spectrum of obesity and mutations in the melanocortin 4 receptor gene. N. Engl. J. Med. 2003, 348, 1085–1095. [Google Scholar] [CrossRef]
- Hinney, A.; Schmidt, A.; Nottebom, K.; Heibult, O.; Becker, I.; Ziegler, A.; Gerber, G.; Sina, M.; Gorg, T.; Mayer, H.; et al. Several mutations in the melanocortin-4 receptor gene including a nonsense and a frameshift mutation associated with dominantly inherited obesity in humans. J. Clin. Endocrinol. Metab. 1999, 84, 1483–1486. [Google Scholar] [CrossRef]
- Schalin-Jantti, C.; Valli-Jaakola, K.; Oksanen, L.; Martelin, E.; Laitinen, K.; Krusius, T.; Mustajoki, P.; Heikinheimo, M.; Kontula, K. Melanocortin-3-receptor gene variants in morbid obesity. Int. J. Obes. Relat. Metab. Disord. 2003, 27, 70–74. [Google Scholar] [CrossRef]
- Begriche, K.; Sutton, G.M.; Butler, A.A. Homeostastic and non-homeostatic functions of melanocortin-3 receptors in the control of energy balance and metabolism. Physiol. Behav. 2011, 104, 546–554. [Google Scholar] [CrossRef] [PubMed]
- Lippert, R.N.; Ellacott, K.L.; Cone, R.D. Gender-specific roles for the melanocortin-3 receptor in the regulation of the mesolimbic dopamine system in mice. Endocrinology 2014, 155, 1718–1727. [Google Scholar] [CrossRef] [PubMed]
- Duncan, A.; Heyer, M.P.; Ishikawa, M.; Caligiuri, S.P.B.; Liu, X.A.; Chen, Z.; Micioni Di Bonaventura, M.V.; Elayouby, K.S.; Ables, J.L.; Howe, W.M.; et al. Habenular TCF7L2 links nicotine addiction to diabetes. Nature 2019, 574, 372–377. [Google Scholar] [CrossRef]
- Zhang, Y.; Kilroy, G.E.; Henagan, T.M.; Prpic-Uhing, V.; Richards, W.G.; Bannon, A.W.; Mynatt, R.L.; Gettys, T.W. Targeted deletion of melanocortin receptor subtypes 3 and 4, but not CART, alters nutrient partitioning and compromises behavioral and metabolic responses to leptin. FASEB J. 2005, 19, 1482–1491. [Google Scholar] [CrossRef] [PubMed]
- Lee, B.; Koo, J.; Yun Jun, J.; Gavrilova, O.; Lee, Y.; Seo, A.Y.; Taylor-Douglas, D.C.; Adler-Wailes, D.C.; Chen, F.; Gardner, R.; et al. A mouse model for a partially inactive obesity-associated human MC3R variant. Nat. Commun. 2016, 7, 10522. [Google Scholar] [CrossRef] [PubMed]
- Feng, N.; Young, S.F.; Aguilera, G.; Puricelli, E.; Adler-Wailes, D.C.; Sebring, N.G.; Yanovski, J.A. Co-occurrence of two partially inactivating polymorphisms of MC3R is associated with pediatric-onset obesity. Diabetes 2005, 54, 2663–2667. [Google Scholar] [CrossRef]
- Lee, Y.S.; Poh, L.K.; Kek, B.L.; Loke, K.Y. The role of melanocortin 3 receptor gene in childhood obesity. Diabetes 2007, 56, 2622–2630. [Google Scholar] [CrossRef]
- Savastano, D.M.; Tanofsky-Kraff, M.; Han, J.C.; Ning, C.; Sorg, R.A.; Roza, C.A.; Wolkoff, L.E.; Anandalingam, K.; Jefferson-George, K.S.; Figueroa, R.E.; et al. Energy intake and energy expenditure among children with polymorphisms of the melanocortin-3 receptor. Am. J. Clin. Nutr. 2009, 90, 912–920. [Google Scholar] [CrossRef]
- Butler, A.A.; Girardet, C.; Mavrikaki, M.; Trevaskis, J.L.; Macarthur, H.; Marks, D.L.; Farr, S.A. A Life without Hunger: The Ups (and Downs) to Modulating Melanocortin-3 Receptor Signaling. Front. Neurosci. 2017, 11, 128. [Google Scholar] [CrossRef]
- Sutton, G.M.; Perez-Tilve, D.; Nogueiras, R.; Fang, J.; Kim, J.K.; Cone, R.D.; Gimble, J.M.; Tschop, M.H.; Butler, A.A. The melanocortin-3 receptor is required for entrainment to meal intake. J. Neurosci. 2008, 28, 12946–12955. [Google Scholar] [CrossRef]
- Begriche, K.; Marston, O.J.; Rossi, J.; Burke, L.K.; McDonald, P.; Heisler, L.K.; Butler, A.A. Melanocortin-3 receptors are involved in adaptation to restricted feeding. Genes Brain Behav. 2012, 11, 291–302. [Google Scholar] [CrossRef] [PubMed]
- Girardet, C.; Mavrikaki, M.M.; Stevens, J.R.; Miller, C.A.; Marks, D.L.; Butler, A.A. Melanocortin-3 receptors expressed in Nkx2.1(+ve) neurons are sufficient for controlling appetitive responses to hypocaloric conditioning. Sci. Rep. 2017, 7, 44444. [Google Scholar] [CrossRef] [PubMed]
- Renquist, B.J.; Murphy, J.G.; Larson, E.A.; Olsen, D.; Klein, R.F.; Ellacott, K.L.; Cone, R.D. Melanocortin-3 receptor regulates the normal fasting response. Proc. Natl. Acad. Sci. USA 2012, 109, E1489. [Google Scholar] [CrossRef]
- Cifani, C.; Polidori, C.; Melotto, S.; Ciccocioppo, R.; Massi, M. A preclinical model of binge eating elicited by yo-yo dieting and stressful exposure to food: Effect of sibutramine, fluoxetine, topiramate, and midazolam. Psychopharmacology 2009, 204, 113–125. [Google Scholar] [CrossRef] [PubMed]
- Hagan, M.M.; Wauford, P.K.; Chandler, P.C.; Jarrett, L.A.; Rybak, R.J.; Blackburn, K. A new animal model of binge eating: Key synergistic role of past caloric restriction and stress. Physiol. Behav. 2002, 77, 45–54. [Google Scholar] [CrossRef]
- Cowley, M.A.; Cone, R.; Enriori, P.; Louiselle, I.; Williams, S.M.; Evans, A.E. Electrophysiological actions of peripheral hormones on melanocortin neurons. Ann. N. Y. Acad. Sci. 2003, 994, 175–186. [Google Scholar] [CrossRef]
- Cowley, M.A.; Smart, J.L.; Rubinstein, M.; Cerdan, M.G.; Diano, S.; Horvath, T.L.; Cone, R.D.; Low, M.J. Leptin activates anorexigenic POMC neurons through a neural network in the arcuate nucleus. Nature 2001, 411, 480–484. [Google Scholar] [CrossRef]
- Marks, D.L.; Hruby, V.; Brookhart, G.; Cone, R.D. The regulation of food intake by selective stimulation of the type 3 melanocortin receptor (MC3R). Peptides 2006, 27, 259–264. [Google Scholar] [CrossRef]
- Gehlert, D.R. Role of hypothalamic neuropeptide Y in feeding and obesity. Neuropeptides 1999, 33, 329–338. [Google Scholar] [CrossRef]
- Lee, M.; Kim, A.; Conwell, I.M.; Hruby, V.; Mayorov, A.; Cai, M.; Wardlaw, S.L. Effects of selective modulation of the central melanocortin-3-receptor on food intake and hypothalamic POMC expression. Peptides 2008, 29, 440–447. [Google Scholar] [CrossRef]
- West, K.S.; Lu, C.; Olson, D.P.; Roseberry, A.G. Alpha-melanocyte stimulating hormone increases the activity of melanocortin-3 receptor-expressing neurons in the ventral tegmental area. J. Physiol. 2019, 597, 3217–3232. [Google Scholar] [CrossRef] [PubMed]
- Kenny, P.J. Reward mechanisms in obesity: New insights and future directions. Neuron 2011, 69, 664–679. [Google Scholar] [CrossRef] [PubMed]
- Volkow, N.D.; Wang, G.J.; Baler, R.D. Reward, dopamine and the control of food intake: Implications for obesity. Trends Cogn. Sci. 2011, 15, 37–46. [Google Scholar] [CrossRef] [PubMed]
- Nair, S.G.; Navarre, B.M.; Cifani, C.; Pickens, C.L.; Bossert, J.M.; Shaham, Y. Role of dorsal medial prefrontal cortex dopamine D1-family receptors in relapse to high-fat food seeking induced by the anxiogenic drug yohimbine. Neuropsychopharmacology 2011, 36, 497–510. [Google Scholar] [CrossRef]
- Wise, R.A. Roles for nigrostriatal—Not just mesocorticolimbic—Dopamine in reward and addiction. Trends Neurosci. 2009, 32, 517–524. [Google Scholar] [CrossRef]
- Lindblom, J.; Opmane, B.; Mutulis, F.; Mutule, I.; Petrovska, R.; Klusa, V.; Bergstrom, L.; Wikberg, J.E. The MC4 receptor mediates alpha-MSH induced release of nucleus accumbens dopamine. Neuroreport 2001, 12, 2155–2158. [Google Scholar] [CrossRef]
- Sanchez, M.S.; Barontini, M.; Armando, I.; Celis, M.E. Correlation of increased grooming behavior and motor activity with alterations in nigrostriatal and mesolimbic catecholamines after alpha-melanotropin and neuropeptide glutamine-isoleucine injection in the rat ventral tegmental area. Cell. Mol. Neurobiol. 2001, 21, 523–533. [Google Scholar] [CrossRef]
- Torre, E.; Celis, M.E. Alpha-MSH injected into the substantia nigra or intraventricularly alters behavior and the striatal dopaminergic activity. Neurochem. Int. 1986, 9, 85–89. [Google Scholar] [CrossRef]
- Torre, E.; Celis, M.E. Cholinergic mediation in the ventral tegmental area of alpha-melanotropin induced excessive grooming: Changes of the dopamine activity in the nucleus accumbens and caudate putamen. Life Sci. 1988, 42, 1651–1657. [Google Scholar] [CrossRef]
- Dietrich, M.O.; Bober, J.; Ferreira, J.G.; Tellez, L.A.; Mineur, Y.S.; Souza, D.O.; Gao, X.B.; Picciotto, M.R.; Araujo, I.; Liu, Z.W.; et al. AgRP neurons regulate development of dopamine neuronal plasticity and nonfood-associated behaviors. Nat. Neurosci. 2012, 15, 1108–1110. [Google Scholar] [CrossRef]
- King, C.M.; Hentges, S.T. Relative number and distribution of murine hypothalamic proopiomelanocortin neurons innervating distinct target sites. PLoS ONE 2011, 6, e25864. [Google Scholar] [CrossRef] [PubMed]
- Liu, H.; Kishi, T.; Roseberry, A.G.; Cai, X.; Lee, C.E.; Montez, J.M.; Friedman, J.M.; Elmquist, J.K. Transgenic mice expressing green fluorescent protein under the control of the melanocortin-4 receptor promoter. J. Neurosci. 2003, 23, 7143–7154. [Google Scholar] [CrossRef] [PubMed]
- Martinez-Hernandez, J.; Lanuza, E.; Martinez-Garcia, F. Selective dopaminergic lesions of the ventral tegmental area impair preference for sucrose but not for male sexual pheromones in female mice. Eur. J. Neurosci. 2006, 24, 885–893. [Google Scholar] [CrossRef] [PubMed]
- Shibata, R.; Kameishi, M.; Kondoh, T.; Torii, K. Bilateral dopaminergic lesions in the ventral tegmental area of rats influence sucrose intake, but not umami and amino acid intake. Physiol. Behav. 2009, 96, 667–674. [Google Scholar] [CrossRef] [PubMed]
- Culbert, K.M.; Racine, S.E.; Klump, K.L. Hormonal Factors and Disturbances in Eating Disorders. Curr. Psychiatry Rep. 2016, 18, 65. [Google Scholar] [CrossRef] [PubMed]
- Edler, C.; Lipson, S.F.; Keel, P.K. Ovarian hormones and binge eating in bulimia nervosa. Psychol. Med. 2007, 37, 131–141. [Google Scholar] [CrossRef]
- Micioni Di Bonaventura, M.V.; Lutz, T.A.; Romano, A.; Pucci, M.; Geary, N.; Asarian, L.; Cifani, C. Estrogenic suppression of binge-like eating elicited by cyclic food restriction and frustrative-nonreward stress in female rats. Int. J. Eat. Disord. 2017, 50, 624–635. [Google Scholar] [CrossRef]
- Alboni, S.; Micioni Di Bonaventura, M.V.; Benatti, C.; Giusepponi, M.E.; Brunello, N.; Cifani, C. Hypothalamic expression of inflammatory mediators in an animal model of binge eating. Behav. Brain Res. 2017, 320, 420–430. [Google Scholar] [CrossRef]
- Davis, J.F.; Choi, D.L.; Shurdak, J.D.; Krause, E.G.; Fitzgerald, M.F.; Lipton, J.W.; Sakai, R.R.; Benoit, S.C. Central melanocortins modulate mesocorticolimbic activity and food seeking behavior in the rat. Physiol. Behav. 2011, 102, 491–495. [Google Scholar] [CrossRef]
- Hagan, M.M.; Rushing, P.A.; Benoit, S.C.; Woods, S.C.; Seeley, R.J. Opioid receptor involvement in the effect of AgRP- (83-132) on food intake and food selection. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2001, 280, R814–R821. [Google Scholar] [CrossRef]
- Tracy, A.L.; Clegg, D.J.; Johnson, J.D.; Davidson, T.L.; Benoit, S.C. The melanocortin antagonist AgRP (83-132) increases appetitive responding for a fat, but not a carbohydrate, reinforcer. Pharmacol. Biochem. Behav. 2008, 89, 263–271. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Samama, P.; Rumennik, L.; Grippo, J.F. The melanocortin receptor MCR4 controls fat consumption. Regul. Pept. 2003, 113, 85–88. [Google Scholar] [CrossRef]
- Boghossian, S.; Park, M.; York, D.A. Melanocortin activity in the amygdala controls appetite for dietary fat. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2010, 298, R385–R393. [Google Scholar] [CrossRef] [PubMed]
- Bassareo, V.; Di Chiara, G. Differential influence of associative and nonassociative learning mechanisms on the responsiveness of prefrontal and accumbal dopamine transmission to food stimuli in rats fed ad libitum. J. Neurosci. 1997, 17, 851–861. [Google Scholar] [CrossRef]
- Botticelli, L.; Micioni Di Bonaventura, E.; Del Bello, F.; Giorgioni, G.; Piergentili, A.; Romano, A.; Quaglia, W.; Cifani, C.; Micioni Di Bonaventura, M.V. Underlying Susceptibility to Eating Disorders and Drug Abuse: Genetic and Pharmacological Aspects of Dopamine D4 Receptors. Nutrients 2020, 12, 2288. [Google Scholar] [CrossRef]
- Cifani, C.; Koya, E.; Navarre, B.M.; Calu, D.J.; Baumann, M.H.; Marchant, N.J.; Liu, Q.R.; Khuc, T.; Pickel, J.; Lupica, C.R.; et al. Medial prefrontal cortex neuronal activation and synaptic alterations after stress-induced reinstatement of palatable food seeking: A study using c-fos-GFP transgenic female rats. J. Neurosci. 2012, 32, 8480–8490. [Google Scholar] [CrossRef]
- Zheng, H.; Corkern, M.M.; Crousillac, S.M.; Patterson, L.M.; Phifer, C.B.; Berthoud, H.R. Neurochemical phenotype of hypothalamic neurons showing Fos expression 23 h after intracranial AgRP. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2002, 282, R1773–R1781. [Google Scholar] [CrossRef][Green Version]
- Stuber, G.D.; Wise, R.A. Lateral hypothalamic circuits for feeding and reward. Nat. Neurosci. 2016, 19, 198–205. [Google Scholar] [CrossRef]
- Piccoli, L.; Micioni Di Bonaventura, M.V.; Cifani, C.; Costantini, V.J.; Massagrande, M.; Montanari, D.; Martinelli, P.; Antolini, M.; Ciccocioppo, R.; Massi, M.; et al. Role of orexin-1 receptor mechanisms on compulsive food consumption in a model of binge eating in female rats. Neuropsychopharmacology 2012, 37, 1999–2011. [Google Scholar] [CrossRef]
- Roseberry, A.G. Altered feeding and body weight following melanocortin administration to the ventral tegmental area in adult rats. Psychopharmacology 2013, 226, 25–34. [Google Scholar] [CrossRef]
- Yen, H.H.; Roseberry, A.G. Decreased consumption of rewarding sucrose solutions after injection of melanocortins into the ventral tegmental area of rats. Psychopharmacology 2015, 232, 285–294. [Google Scholar] [CrossRef] [PubMed]
- Shanmugarajah, L.; Dunigan, A.I.; Frantz, K.J.; Roseberry, A.G. Altered sucrose self-administration following injection of melanocortin receptor agonists and antagonists into the ventral tegmental area. Psychopharmacology 2017, 234, 1683–1692. [Google Scholar] [CrossRef] [PubMed]
- Potoczna, N.; Branson, R.; Kral, J.G.; Piec, G.; Steffen, R.; Ricklin, T.; Hoehe, M.R.; Lentes, K.U.; Horber, F.F. Gene variants and binge eating as predictors of comorbidity and outcome of treatment in severe obesity. J. Gastrointest. Surg. 2004, 8, 971–981; discussion 972–981. [Google Scholar] [CrossRef] [PubMed]
- Sina, M.; Hinney, A.; Ziegler, A.; Neupert, T.; Mayer, H.; Siegfried, W.; Blum, W.F.; Remschmidt, H.; Hebebrand, J. Phenotypes in three pedigrees with autosomal dominant obesity caused by haploinsufficiency mutations in the melanocortin-4 receptor gene. Am. J. Hum. Genet. 1999, 65, 1501–1507. [Google Scholar] [CrossRef]
- Spitzer, R.L.; Yanovski, S.; Wadden, T.; Wing, R.; Marcus, M.D.; Stunkard, A.; Devlin, M.; Mitchell, J.; Hasin, D.; Horne, R.L. Binge eating disorder: Its further validation in a multisite study. Int. J. Eat. Disord. 1993, 13, 137–153. [Google Scholar]
- Gotoda, T. Binge eating as a phenotype of melanocortin 4 receptor gene mutations. N. Engl. J. Med. 2003, 349, 606–609. [Google Scholar] [CrossRef]
- Hebebrand, J.; Geller, F.; Dempfle, A.; Heinzel-Gutenbrunner, M.; Raab, M.; Gerber, G.; Wermter, A.K.; Horro, F.F.; Blundell, J.; Schafer, H.; et al. Binge-eating episodes are not characteristic of carriers of melanocortin-4 receptor gene mutations. Mol. Psychiatry 2004, 9, 796–800. [Google Scholar] [CrossRef][Green Version]
- Lubrano-Berthelier, C.; Dubern, B.; Lacorte, J.M.; Picard, F.; Shapiro, A.; Zhang, S.; Bertrais, S.; Hercberg, S.; Basdevant, A.; Clement, K.; et al. Melanocortin 4 receptor mutations in a large cohort of severely obese adults: Prevalence, functional classification, genotype-phenotype relationship, and lack of association with binge eating. J. Clin. Endocrinol. Metab. 2006, 91, 1811–1818. [Google Scholar] [CrossRef]
- Tao, Y.X.; Segaloff, D.L. Functional analyses of melanocortin-4 receptor mutations identified from patients with binge eating disorder and nonobese or obese subjects. J. Clin. Endocrinol. Metab. 2005, 90, 5632–5638. [Google Scholar] [CrossRef]
- Gamero-Villarroel, C.; Rodriguez-Lopez, R.; Jimenez, M.; Carrillo, J.A.; Garcia-Herraiz, A.; Albuquerque, D.; Flores, I.; Gervasini, G. Melanocortin-4 receptor gene variants are not associated with binge-eating behavior in nonobese patients with eating disorders. Psychiatr. Genet. 2015, 25, 35–38. [Google Scholar] [CrossRef]
- Valette, M.; Poitou, C.; Le Beyec, J.; Bouillot, J.L.; Clement, K.; Czernichow, S. Melanocortin-4 receptor mutations and polymorphisms do not affect weight loss after bariatric surgery. PLoS ONE 2012, 7, e48221. [Google Scholar] [CrossRef] [PubMed]
- Bonnefond, A.; Keller, R.; Meyre, D.; Stutzmann, F.; Thuillier, D.; Stefanov, D.G.; Froguel, P.; Horber, F.F.; Kral, J.G. Eating Behavior, Low-Frequency Functional Mutations in the Melanocortin-4 Receptor (MC4R) Gene, and Outcomes of Bariatric Operations: A 6-Year Prospective Study. Diabetes Care 2016, 39, 1384–1392. [Google Scholar] [CrossRef] [PubMed]
- Valette, M.; Poitou, C.; Kesse-Guyot, E.; Bellisle, F.; Carette, C.; Le Beyec, J.; Hercberg, S.; Clement, K.; Czernichow, S. Association between melanocortin-4 receptor mutations and eating behaviors in obese patients: A case-control study. Int. J. Obes. 2014, 38, 883–885. [Google Scholar] [CrossRef]
- Van der Klaauw, A.A.; von dem Hagen, E.A.; Keogh, J.M.; Henning, E.; O’Rahilly, S.; Lawrence, A.D.; Calder, A.J.; Farooqi, I.S. Obesity-associated melanocortin-4 receptor mutations are associated with changes in the brain response to food cues. J. Clin. Endocrinol. Metab. 2014, 99, E2101–E2106. [Google Scholar] [CrossRef] [PubMed]
- Furlong, T.M.; Jayaweera, H.K.; Balleine, B.W.; Corbit, L.H. Binge-like consumption of a palatable food accelerates habitual control of behavior and is dependent on activation of the dorsolateral striatum. J. Neurosci. 2014, 34, 5012–5022. [Google Scholar] [CrossRef]
- Wang, G.J.; Geliebter, A.; Volkow, N.D.; Telang, F.W.; Logan, J.; Jayne, M.C.; Galanti, K.; Selig, P.A.; Han, H.; Zhu, W.; et al. Enhanced striatal dopamine release during food stimulation in binge eating disorder. Obesity 2011, 19, 1601–1608. [Google Scholar] [CrossRef]
- Thorleifsson, G.; Walters, G.B.; Gudbjartsson, D.F.; Steinthorsdottir, V.; Sulem, P.; Helgadottir, A.; Styrkarsdottir, U.; Gretarsdottir, S.; Thorlacius, S.; Jonsdottir, I.; et al. Genome-wide association yields new sequence variants at seven loci that associate with measures of obesity. Nat. Genet. 2009, 41, 18–24. [Google Scholar] [CrossRef]
- Loos, R.J.; Lindgren, C.M.; Li, S.; Wheeler, E.; Zhao, J.H.; Prokopenko, I.; Inouye, M.; Freathy, R.M.; Attwood, A.P.; Beckmann, J.S.; et al. Common variants near MC4R are associated with fat mass, weight and risk of obesity. Nat. Genet. 2008, 40, 768–775. [Google Scholar] [CrossRef]
- Horstmann, A.; Kovacs, P.; Kabisch, S.; Boettcher, Y.; Schloegl, H.; Tonjes, A.; Stumvoll, M.; Pleger, B.; Villringer, A. Common genetic variation near MC4R has a sex-specific impact on human brain structure and eating behavior. PLoS ONE 2013, 8, e74362. [Google Scholar] [CrossRef]
- Qi, L.; Kraft, P.; Hunter, D.J.; Hu, F.B. The common obesity variant near MC4R gene is associated with higher intakes of total energy and dietary fat, weight change and diabetes risk in women. Hum. Mol. Genet. 2008, 17, 3502–3508. [Google Scholar] [CrossRef]
- Stutzmann, F.; Cauchi, S.; Durand, E.; Calvacanti-Proenca, C.; Pigeyre, M.; Hartikainen, A.L.; Sovio, U.; Tichet, J.; Marre, M.; Weill, J.; et al. Common genetic variation near MC4R is associated with eating behaviour patterns in European populations. Int. J. Obes. 2009, 33, 373–378. [Google Scholar] [CrossRef] [PubMed]
- Yilmaz, Z.; Davis, C.; Loxton, N.J.; Kaplan, A.S.; Levitan, R.D.; Carter, J.C.; Kennedy, J.L. Association between MC4R rs17782313 polymorphism and overeating behaviors. Int. J. Obes. 2015, 39, 114–120. [Google Scholar] [CrossRef] [PubMed]
- Blaine, R.E.; Kachurak, A.; Davison, K.K.; Klabunde, R.; Fisher, J.O. Food parenting and child snacking: A systematic review. Int. J. Behav. Nutr. Phys. Act. 2017, 14, 146. [Google Scholar] [CrossRef] [PubMed]
- Potter, M.; Vlassopoulos, A.; Lehmann, U. Snacking Recommendations Worldwide: A Scoping Review. Adv. Nutr. 2018, 9, 86–98. [Google Scholar] [CrossRef] [PubMed]
- Acosta, A.; Camilleri, M.; Shin, A.; Carlson, P.; Burton, D.; O’Neill, J.; Eckert, D.; Zinsmeister, A.R. Association of melanocortin 4 receptor gene variation with satiation and gastric emptying in overweight and obese adults. Genes Nutr. 2014, 9, 384. [Google Scholar] [CrossRef]
- Valladares, M.; Dominguez-Vasquez, P.; Obregon, A.M.; Weisstaub, G.; Burrows, R.; Maiz, A.; Santos, J.L. Melanocortin-4 receptor gene variants in Chilean families: Association with childhood obesity and eating behavior. Nutr. Neurosci. 2010, 13, 71–78. [Google Scholar] [CrossRef] [PubMed]
- Wardle, J.; Guthrie, C.A.; Sanderson, S.; Rapoport, L. Development of the Children’s Eating Behaviour Questionnaire. J. Child Psychol. Psychiatry Allied Discip. 2001, 42, 963–970. [Google Scholar] [CrossRef]
- De Lauzon, B.; Romon, M.; Deschamps, V.; Lafay, L.; Borys, J.M.; Karlsson, J.; Ducimetiere, P.; Charles, M.A.; Fleurbaix Laventie Ville Sante Study, G. The Three-Factor Eating Questionnaire-R18 is able to distinguish among different eating patterns in a general population. J. Nutr. 2004, 134, 2372–2380. [Google Scholar] [CrossRef]
- Turton, R.; Chami, R.; Treasure, J. Emotional Eating, Binge Eating and Animal Models of Binge-Type Eating Disorders. Curr. Obes. Rep. 2017, 6, 217–228. [Google Scholar] [CrossRef]
- Van Strien, T.; Cebolla, A.; Etchemendy, E.; Gutierrez-Maldonado, J.; Ferrer-Garcia, M.; Botella, C.; Banos, R. Emotional eating and food intake after sadness and joy. Appetite 2013, 66, 20–25. [Google Scholar] [CrossRef]
- Obregon, A.M.; Oyarce, K.; Santos, J.L.; Valladares, M.; Goldfield, G. Association of the melanocortin 4 receptor gene rs17782313 polymorphism with rewarding value of food and eating behavior in Chilean children. J. Physiol. Biochem. 2017, 73, 29–35. [Google Scholar] [CrossRef] [PubMed]
- Yilmaz, Z.; Kaplan, A.S.; Tiwari, A.K.; Levitan, R.D.; Piran, S.; Bergen, A.W.; Kaye, W.H.; Hakonarson, H.; Wang, K.; Berrettini, W.H.; et al. The role of leptin, melanocortin, and neurotrophin system genes on body weight in anorexia nervosa and bulimia nervosa. J. Psychiatr. Res. 2014, 55, 77–86. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Abadi, A.; Peralta-Romero, J.; Suarez, F.; Gomez-Zamudio, J.; Burguete-Garcia, A.I.; Cruz, M.; Meyre, D. Assessing the effects of 35 European-derived BMI-associated SNPs in Mexican children. Obesity 2016, 24, 1989–1995. [Google Scholar] [CrossRef] [PubMed]
- Cheung, C.Y.; Tso, A.W.; Cheung, B.M.; Xu, A.; Ong, K.L.; Fong, C.H.; Wat, N.M.; Janus, E.D.; Sham, P.C.; Lam, K.S. Obesity susceptibility genetic variants identified from recent genome-wide association studies: Implications in a Chinese population. J. Clin. Endocrinol. Metab. 2010, 95, 1395–1403. [Google Scholar] [CrossRef] [PubMed]
- El Hajj Chehadeh, S.; Osman, W.; Nazar, S.; Jerman, L.; Alghafri, A.; Sajwani, A.; Alawlaqi, M.; AlObeidli, M.; Jelinek, H.F.; AlAnouti, F.; et al. Implication of genetic variants in overweight and obesity susceptibility among the young Arab population of the United Arab Emirates. Gene 2020, 739, 144509. [Google Scholar] [CrossRef]
- Khalilitehrani, A.; Qorbani, M.; Hosseini, S.; Pishva, H. The association of MC4R rs17782313 polymorphism with dietary intake in Iranian adults. Gene 2015, 563, 125–129. [Google Scholar] [CrossRef]
- Lv, D.; Zhang, D.D.; Wang, H.; Zhang, Y.; Liang, L.; Fu, J.F.; Xiong, F.; Liu, G.L.; Gong, C.X.; Luo, F.H.; et al. Genetic variations in SEC16B, MC4R, MAP2K5 and KCTD15 were associated with childhood obesity and interacted with dietary behaviors in Chinese school-age population. Gene 2015, 560, 149–155. [Google Scholar] [CrossRef]
- Rana, S.; Rahmani, S.; Mirza, S. MC4R variant rs17782313 and manifestation of obese phenotype in Pakistani females. RSC Adv. 2018, 8, 16957–16972. [Google Scholar] [CrossRef]
- Shi, J.; Long, J.; Gao, Y.T.; Lu, W.; Cai, Q.; Wen, W.; Zheng, Y.; Yu, K.; Xiang, Y.B.; Hu, F.B.; et al. Evaluation of genetic susceptibility loci for obesity in Chinese women. Am. J. Epidemiol. 2010, 172, 244–254. [Google Scholar] [CrossRef]
- Zobel, D.P.; Andreasen, C.H.; Grarup, N.; Eiberg, H.; Sorensen, T.I.; Sandbaek, A.; Lauritzen, T.; Borch-Johnsen, K.; Jorgensen, T.; Pedersen, O.; et al. Variants near MC4R are associated with obesity and influence obesity-related quantitative traits in a population of middle-aged people: Studies of 14,940 Danes. Diabetes 2009, 58, 757–764. [Google Scholar] [CrossRef]
- Orkunoglu-Suer, F.E.; Harmon, B.T.; Gordish-Dressman, H.; Clarkson, P.M.; Thompson, P.D.; Angelopoulos, T.J.; Gordon, P.M.; Hubal, M.J.; Moyna, N.M.; Pescatello, L.S.; et al. MC4R variant is associated with BMI but not response to resistance training in young females. Obesity 2011, 19, 662–666. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Stevenson, R.J.; Francis, H.M. The hippocampus and the regulation of human food intake. Psychol. Bull. 2017, 143, 1011–1032. [Google Scholar] [CrossRef] [PubMed]
- Uher, R.; Murphy, T.; Brammer, M.J.; Dalgleish, T.; Phillips, M.L.; Ng, V.W.; Andrew, C.M.; Williams, S.C.; Campbell, I.C.; Treasure, J. Medial prefrontal cortex activity associated with symptom provocation in eating disorders. Am. J. Psychiatry 2004, 161, 1238–1246. [Google Scholar] [CrossRef] [PubMed]
- Freeman, L.M.; Gil, K.M. Daily stress, coping, and dietary restraint in binge eating. Int. J. Eat. Disord. 2004, 36, 204–212. [Google Scholar] [CrossRef]
- Cifani, C.; Di Bonaventura, M.V.M.; Ciccocioppo, R.; Massi, M. Binge eating in female rats induced by yo-yo dieting and stress. In Animal Models of Eating Disorders; Springer: Totowa, NJ, USA, 2013; pp. 27–49. [Google Scholar]
- Polivy, J.; Zeitlin, S.B.; Herman, C.P.; Beal, A.L. Food restriction and binge eating: A study of former prisoners of war. J. Abnorm. Psychol. 1994, 103, 409–411. [Google Scholar] [CrossRef]
- Woods, A.M.; Racine, S.E.; Klump, K.L. Examining the relationship between dietary restraint and binge eating: Differential effects of major and minor stressors. Eat. Behav. 2010, 11, 276–280. [Google Scholar] [CrossRef]
- Gluck, M.E.; Geliebter, A.; Hung, J.; Yahav, E. Cortisol, hunger, and desire to binge eat following a cold stress test in obese women with binge eating disorder. Psychosom. Med. 2004, 66, 876–881. [Google Scholar] [CrossRef]
- Gluck, M.E.; Geliebter, A.; Lorence, M. Cortisol stress response is positively correlated with central obesity in obese women with binge eating disorder (BED) before and after cognitive-behavioral treatment. Ann. N. Y. Acad. Sci. 2004, 1032, 202–207. [Google Scholar] [CrossRef]
- Cifani, C.; Micioni Di, B.M.; Vitale, G.; Ruggieri, V.; Ciccocioppo, R.; Massi, M. Effect of salidroside, active principle of Rhodiola rosea extract, on binge eating. Physiol. Behav. 2010, 101, 555–562. [Google Scholar] [CrossRef]
- Micioni Di Bonaventura, M.V.; Vitale, G.; Massi, M.; Cifani, C. Effect of Hypericum perforatum Extract in an Experimental Model of Binge Eating in Female Rats. J. Obes. 2012, 2012, 956137. [Google Scholar] [CrossRef]
- Epel, E.; Lapidus, R.; McEwen, B.; Brownell, K. Stress may add bite to appetite in women: A laboratory study of stress-induced cortisol and eating behavior. Psychoneuroendocrinology 2001, 26, 37–49. [Google Scholar] [CrossRef]
- Coutinho, W.F.; Moreira, R.O.; Spagnol, C.; Appolinario, J.C. Does binge eating disorder alter cortisol secretion in obese women? Eat. Behav. 2007, 8, 59–64. [Google Scholar] [CrossRef] [PubMed]
- Chaki, S.; Okubo, T. Melanocortin-4 receptor antagonists for the treatment of depression and anxiety disorders. Curr. Top. Med. Chem. 2007, 7, 1145–1151. [Google Scholar] [CrossRef] [PubMed]
- Chaki, S.; Okuyama, S. Involvement of melanocortin-4 receptor in anxiety and depression. Peptides 2005, 26, 1952–1964. [Google Scholar] [CrossRef] [PubMed]
- Adan, R.A.; Szklarczyk, A.W.; Oosterom, J.; Brakkee, J.H.; Nijenhuis, W.A.; Schaaper, W.M.; Meloen, R.H.; Gispen, W.H. Characterization of melanocortin receptor ligands on cloned brain melanocortin receptors and on grooming behavior in the rat. Eur. J. Pharmacol. 1999, 378, 249–258. [Google Scholar] [CrossRef]
- De Barioglio, S.R.; Lezcano, N.; Celis, M.E. Alpha MSH-induced excessive grooming behavior involves a GABAergic mechanism. Peptides 1991, 12, 203–205. [Google Scholar] [CrossRef]
- Spruijt, B.M.; van Hooff, J.A.; Gispen, W.H. Ethology and neurobiology of grooming behavior. Physiol. Rev. 1992, 72, 825–852. [Google Scholar] [CrossRef]
- Mul, J.D.; van Boxtel, R.; Bergen, D.J.; Brans, M.A.; Brakkee, J.H.; Toonen, P.W.; Garner, K.M.; Adan, R.A.; Cuppen, E. Melanocortin receptor 4 deficiency affects body weight regulation, grooming behavior, and substrate preference in the rat. Obesity 2012, 20, 612–621. [Google Scholar] [CrossRef]
- Yamano, Y.; Yoshioka, M.; Toda, Y.; Oshida, Y.; Chaki, S.; Hamamoto, K.; Morishima, I. Regulation of CRF, POMC and MC4R gene expression after electrical foot shock stress in the rat amygdala and hypothalamus. J. Vet. Med Sci. 2004, 66, 1323–1327. [Google Scholar] [CrossRef]
- Davis, M.; Shi, C. The extended amygdala: Are the central nucleus of the amygdala and the bed nucleus of the stria terminalis differentially involved in fear versus anxiety? Ann. N. Y. Acad. Sci. 1999, 877, 281–291. [Google Scholar] [CrossRef]
- Blasio, A.; Iemolo, A.; Sabino, V.; Petrosino, S.; Steardo, L.; Rice, K.C.; Orlando, P.; Iannotti, F.A.; Di Marzo, V.; Zorrilla, E.P.; et al. Rimonabant precipitates anxiety in rats withdrawn from palatable food: Role of the central amygdala. Neuropsychopharmacology 2013, 38, 2498–2507. [Google Scholar] [CrossRef] [PubMed]
- Micioni Di Bonaventura, M.V.; Pucci, M.; Giusepponi, M.E.; Romano, A.; Lambertucci, C.; Volpini, R.; Micioni Di Bonaventura, E.; Gaetani, S.; Maccarrone, M.; D’Addario, C.; et al. Regulation of adenosine A2A receptor gene expression in a model of binge eating in the amygdaloid complex of female rats. J. Psychopharmacol. 2019. [Google Scholar] [CrossRef] [PubMed]
- Karami Kheirabad, M.; Namavar Jahromi, B.; Tamadon, A.; Ramezani, A.; Ahmadloo, S.; Sabet Sarvestan, F.; Koohi-Hosseinabadi, O. Expression of Melanocortin-4 Receptor mRNA in Male Rat Hypothalamus During Chronic Stress. Int. J. Mol. Cell. Med. 2015, 4, 182–187. [Google Scholar] [PubMed]
- Chagra, S.L.; Zavala, J.K.; Hall, M.V.; Gosselink, K.L. Acute and repeated restraint differentially activate orexigenic pathways in the rat hypothalamus. Regul. Pept. 2011, 167, 70–78. [Google Scholar] [CrossRef]
- Von Frijtag, J.C.; Croiset, G.; Gispen, W.H.; Adan, R.A.; Wiegant, V.M. The role of central melanocortin receptors in the activation of the hypothalamus-pituitary-adrenal-axis and the induction of excessive grooming. Br. J. Pharmacol. 1998, 123, 1503–1508. [Google Scholar] [CrossRef][Green Version]
- Lu, X.Y.; Barsh, G.S.; Akil, H.; Watson, S.J. Interaction between alpha-melanocyte-stimulating hormone and corticotropin-releasing hormone in the regulation of feeding and hypothalamo-pituitary-adrenal responses. J. Neurosci. 2003, 23, 7863–7872. [Google Scholar] [CrossRef]
- Hwang, B.H.; Guntz, J.M. Downregulation of corticotropin-releasing factor mRNA, but not vasopressin mRNA, in the paraventricular hypothalamic nucleus of rats following nutritional stress. Brain Res. Bull. 1997, 43, 509–514. [Google Scholar] [CrossRef]
- Liposits, Z.; Sievers, L.; Paull, W.K. Neuropeptide-Y and ACTH-immunoreactive innervation of corticotropin releasing factor (CRF)-synthesizing neurons in the hypothalamus of the rat. An immunocytochemical analysis at the light and electron microscopic levels. Histochemistry 1988, 88, 227–234. [Google Scholar] [CrossRef]
- Sarkar, S.; Legradi, G.; Lechan, R.M. Intracerebroventricular administration of alpha-melanocyte stimulating hormone increases phosphorylation of CREB in TRH- and CRH-producing neurons of the hypothalamic paraventricular nucleus. Brain Res. 2002, 945, 50–59. [Google Scholar] [CrossRef]
- Liu, J.; Garza, J.C.; Truong, H.V.; Henschel, J.; Zhang, W.; Lu, X.Y. The melanocortinergic pathway is rapidly recruited by emotional stress and contributes to stress-induced anorexia and anxiety-like behavior. Endocrinology 2007, 148, 5531–5540. [Google Scholar] [CrossRef]
- Ulrich-Lai, Y.M.; Herman, J.P. Neural regulation of endocrine and autonomic stress responses. Nat. Rev. Neurosci. 2009, 10, 397–409. [Google Scholar] [CrossRef] [PubMed]
- Bedse, G.; Romano, A.; Tempesta, B.; Lavecchia, M.A.; Pace, L.; Bellomo, A.; Duranti, A.; Micioni Di Bonaventura, M.V.; Cifani, C.; Cassano, T.; et al. Inhibition of anandamide hydrolysis enhances noradrenergic and GABAergic transmission in the prefrontal cortex and basolateral amygdala of rats subjected to acute swim stress. J. Neurosci. Res. 2015, 93, 777–787. [Google Scholar] [CrossRef] [PubMed]
- Dayas, C.V.; Buller, K.M.; Day, T.A. Neuroendocrine responses to an emotional stressor: Evidence for involvement of the medial but not the central amygdala. Eur. J. Neurosci. 1999, 11, 2312–2322. [Google Scholar] [CrossRef]
- Roozendaal, B.; McEwen, B.S.; Chattarji, S. Stress, memory and the amygdala. Nat. Rev. Neurosci. 2009, 10, 423–433. [Google Scholar] [CrossRef] [PubMed]
- Choi, D.C.; Furay, A.R.; Evanson, N.K.; Ostrander, M.M.; Ulrich-Lai, Y.M.; Herman, J.P. Bed nucleus of the stria terminalis subregions differentially regulate hypothalamic-pituitary-adrenal axis activity: Implications for the integration of limbic inputs. J. Neurosci. 2007, 27, 2025–2034. [Google Scholar] [CrossRef]
- Forray, M.I.; Gysling, K. Role of noradrenergic projections to the bed nucleus of the stria terminalis in the regulation of the hypothalamic-pituitary-adrenal axis. Brain Res. Brain Res. Rev. 2004, 47, 145–160. [Google Scholar] [CrossRef] [PubMed]
- Micioni Di Bonaventura, M.V.; Ciccocioppo, R.; Romano, A.; Bossert, J.M.; Rice, K.C.; Ubaldi, M.; St Laurent, R.; Gaetani, S.; Massi, M.; Shaham, Y.; et al. Role of bed nucleus of the stria terminalis corticotrophin-releasing factor receptors in frustration stress-induced binge-like palatable food consumption in female rats with a history of food restriction. J. Neurosci. 2014, 34, 11316–11324. [Google Scholar] [CrossRef] [PubMed]
- Micioni Di Bonaventura, M.V.; Ubaldi, M.; Giusepponi, M.E.; Rice, K.C.; Massi, M.; Ciccocioppo, R.; Cifani, C. Hypothalamic CRF1 receptor mechanisms are not sufficient to account for binge-like palatable food consumption in female rats. Int. J. Eat. Disord. 2017, 50, 1194–1204. [Google Scholar] [CrossRef]
- Chaffin, A.T.; Fang, Y.; Larson, K.R.; Mul, J.D.; Ryan, K.K. Sex-dependent effects of MC4R genotype on HPA axis tone: Implications for stress-associated cardiometabolic disease. Stress 2019, 22, 571–580. [Google Scholar] [CrossRef]
- Ryan, K.K.; Mul, J.D.; Clemmensen, C.; Egan, A.E.; Begg, D.P.; Halcomb, K.; Seeley, R.J.; Herman, J.P.; Ulrich-Lai, Y.M. Loss of melanocortin-4 receptor function attenuates HPA responses to psychological stress. Psychoneuroendocrinology 2014, 42, 98–105. [Google Scholar] [CrossRef]
- Cottone, P.; Sabino, V.; Roberto, M.; Bajo, M.; Pockros, L.; Frihauf, J.B.; Fekete, E.M.; Steardo, L.; Rice, K.C.; Grigoriadis, D.E.; et al. CRF system recruitment mediates dark side of compulsive eating. Proc. Natl. Acad. Sci. USA 2009, 106, 20016–20020. [Google Scholar] [CrossRef] [PubMed]
- Iemolo, A.; Blasio, A.; St Cyr, S.A.; Jiang, F.; Rice, K.C.; Sabino, V.; Cottone, P. CRF-CRF1 receptor system in the central and basolateral nuclei of the amygdala differentially mediates excessive eating of palatable food. Neuropsychopharmacology 2013, 38, 2456–2466. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Romano, A.; Micioni Di Bonaventura, M.V.; Gallelli, C.A.; Koczwara, J.B.; Smeets, D.; Giusepponi, M.E.; De Ceglia, M.; Friuli, M.; Micioni Di Bonaventura, E.; Scuderi, C.; et al. Oleoylethanolamide decreases frustration stress-induced binge-like eating in female rats: A novel potential treatment for binge eating disorder. Neuropsychopharmacology 2020, 45, 1931–1941. [Google Scholar] [CrossRef] [PubMed]
- Cifani, C.; Micioni Di Bonaventura, E.; Botticelli, L.; Del Bello, F.; Giorgioni, G.; Pavletic, P.; Piergentili, A.; Quaglia, W.; Bonifazi, A.; Schepmann, D.; et al. Novel Highly Potent and Selective Sigma1 Receptor Antagonists Effectively Block the Binge Eating Episode in Female Rats. ACS Chem. Neurosci. 2020, 11, 3107–3116. [Google Scholar] [CrossRef] [PubMed]
- Pucci, M.; Micioni Di Bonaventura, M.V.; Zaplatic, E.; Bellia, F.; Maccarrone, M.; Cifani, C.; D’Addario, C. Transcriptional regulation of the endocannabinoid system in a rat model of binge-eating behavior reveals a selective modulation of the hypothalamic fatty acid amide hydrolase gene. Int. J. Eat. Disord. 2018, 52, 51–60. [Google Scholar] [CrossRef] [PubMed]
- Micioni Di Bonaventura, M.V.; Cifani, C.; Lambertucci, C.; Volpini, R.; Cristalli, G.; Massi, M. A2A adenosine receptor agonists reduce both high-palatability and low-palatability food intake in female rats. Behav. Pharmacol. 2012, 23, 567–574. [Google Scholar] [CrossRef] [PubMed]



{kind=link}
{kind=link}
| Species | Experiment | Result | Ref. |
|---|---|---|---|
| MC3R KO vs. WT mice | Sucrose preference test | ↓ sucrose intake and preference in female MC3R KO mice | [74] |
| MC3Rtm1Butl (MC3RTB/TB) vs. WT mice | Food self-administration under fixed and progressive ratio protocols | ↓ self-administration of a food reward in MC3RTB/TB mice exposed to caloric restriction | [21] |
| Rats | Food self-administration under fixed and progressive ratio protocols | ↑ operant response, but not free access to sucrose after injection of the MC3R agonist γ-MSH | [22] |
| Rats | Consumption of a high-fat vs. low-fat diet | ↑ intake of a high-fat diet vs. a low-fat diet after i.c.v. injection of AgRP | [112] |
| MC4R +/+ vs. MC4R −/− mice | Consumption of a three-choice diet (fat, protein, carbohydrate) | ↓ fat intake in MC4R +/+, but not in MC4R −/− mice after injections of the MC3R/MC4R agonist MTII and the MC4R agonist (pentacyclo(D-K)-Asp-cis Apc-(D)Phe-Arg-Trp-Lys-NH2) | [114] |
| Rats | High-fat vs. low-fat diet paradigm | ↓ the high-fat diet intake after injection of MTII in the CeA; ↑ high-fat diet consumption after the injection MCRs antagonists SHU-9119 and AgRP | [115] |
| Rats | Fat and sugar consumption under an operant conditioning paradigm | ↑ active response to earn a peanut oil emulsion (100% fat) reinforcer, but not a sucrose (100% carbohydrate) reinforcer after i.c.v. injection of AgRP | [113] |
| Rats | Conditioned place preference for high-fat diet and sucrose pellets | AgRP supports conditioned place preference for a high-fat diet, while blocks the acquisition of place preference for sucrose pellets | [111] |
| Rats | Consumption of standard chow | ↓ intake of standard chow after intra-VTA injection of MTII; ↑ 24-h food intake with SHU-9119 | [122] |
| Rats | Two-bottle choice paradigm for a sucrose solution | ↓ consumption of a 1 and 2% sucrose solutions with intra-VTA injections of MTII; ↓ intake of the more appetizing 10% sucrose solution only at the highest dose of MTII | [39] |
| Rats | Food self-administration under fixed and progressive ratio protocols | ↓ operant response with α-MSH and ↑ operant response with AgRP injected in the NAc shell; no influence on free consumption of sucrose pellets | [23] |
| Rats | Food self-administration under fixed and progressive ratio protocols | ↓ sucrose self-administration on both fixed and progressive ratio schedules with intra-VTA injections of MTII; ↑ self-administration, only under fixed ratio protocols with SHU-9119 | [124] |
| Subjects with the MC4R Variant rs17782313 | Result | Ref. |
|---|---|---|
| Normal weight vs. obese children of both sexes | Obese children present high scores of Enjoyment of Food, Emotional Overeating, Food Responsiveness and lower Satiety Responsiveness | [17] |
| Adult participants | In both genders high scores of Emotional Eating associated with BMI were found, while only in women the Uncontrolled Eating scores were associated with BMI. | [20] |
| Healthy adult volunteers | Only women, especially homozygous carriers of MC4R variant rs17782313, demonstrated Emotional Eating and Disinhibition of Eating. | [141] |
| Adult women | Women had significantly higher intake of energy from fat, compared to carbohydrate. | [142] |
| Children, teenagers and adults | Children and teenagers presented snacking and eating large amounts of food during meals. Adults presented a greater hunger score | [143] |
| Adults between the ages of 24 and 50 years | Overeating behaviors, Emotional Eating and Food Cravings. | [144] |
| Overweight or obese participants | Less postprandial satiation symptoms after a fully caloric meal | [147] |
| Obese children | Low Satiety Responsiveness scores and high scores for the Enjoyment of Food | [148] |
| Obese, overweight and normal weight children | In obese girls were found significant lower scores of the Satiety Responsiveness and higher scores of the Uncontrolled Eating | [153] |
| Women | Association with increased BMI and obesity | [160,161,163] |
| Lean, overweight, and obese children | Association with increased BMI and obesity | [155,159] |
| Normal weight vs. obese adults | Association with increased BMI and obesity | [156,162] |
| Normal weight vs. obese adults | Association with increased BMI and obesity and a significant higher intake of energy from fat compared to carbohydrate. | [158] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Micioni Di Bonaventura, E.; Botticelli, L.; Tomassoni, D.; Tayebati, S.K.; Micioni Di Bonaventura, M.V.; Cifani, C. The Melanocortin System behind the Dysfunctional Eating Behaviors. Nutrients 2020, 12, 3502. https://doi.org/10.3390/nu12113502
Micioni Di Bonaventura E, Botticelli L, Tomassoni D, Tayebati SK, Micioni Di Bonaventura MV, Cifani C. The Melanocortin System behind the Dysfunctional Eating Behaviors. Nutrients. 2020; 12(11):3502. https://doi.org/10.3390/nu12113502
Chicago/Turabian StyleMicioni Di Bonaventura, Emanuela, Luca Botticelli, Daniele Tomassoni, Seyed Khosrow Tayebati, Maria Vittoria Micioni Di Bonaventura, and Carlo Cifani. 2020. "The Melanocortin System behind the Dysfunctional Eating Behaviors" Nutrients 12, no. 11: 3502. https://doi.org/10.3390/nu12113502
APA StyleMicioni Di Bonaventura, E., Botticelli, L., Tomassoni, D., Tayebati, S. K., Micioni Di Bonaventura, M. V., & Cifani, C. (2020). The Melanocortin System behind the Dysfunctional Eating Behaviors. Nutrients, 12(11), 3502. https://doi.org/10.3390/nu12113502