

Organic Acids in Food Preservation: Exploring Synergies, Molecular Insights, and Sustainable Applications



Abstract
1. Introduction
2. General Characteristics of the Organic Acids
3. Mode of Action of the Organic Acids on Cells
4. Antimicrobial Activities of Organic Acids and Their Interactions
4.1. Acetic Acid
4.2. Adipic Acid
4.3. Butyric Acid
4.4. Caprylic Acid
4.5. Citric Acid
4.6. Fumaric Acid
4.7. Lactic Acid
4.8. Malic Acid
4.9. Phenyllactic Acid
4.10. Propionic Acid
4.11. Gallic Acid
4.12. Succinic Acid
4.13. Tartaric Acid
5. Application of Organic Acids in Combination with Other Bioactive Compounds in Food Preservation
6. Sustainability Considerations in the Use of Organic Acids as Food Preservatives
7. Conclusions and Future Perspectives
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
| GRAS | Generally recognized as safe |
| FDA | Food and Drug Administration |
| TLA | Acid constant |
| LD | Adenosine Triphosphate |
| ZnO | Zinc Oxide |
| MIC | Minimum inhibitory concentration |
| MBC | Minimum bactericidal concentration |
| MFC | Minimum fungicidal concentration |
| PBAT | Poly (butylene adipate-co-terephthalate) |
| CNPs | Porous Carbon Nanoparticles |
| AAME | Adipic Acid Monoethyl Ester |
| ClO2 | Chlorine dioxide |
| UV-A | Ultraviolet-A |
| AgNPs | Nano-sized silver particles |
| ACP | Atmospheric Cold Plasma |
| TVC | Total viable count |
| EPS | Extracellular polymeric substances |
| OEO | Oregano essential oil |
| GSE | Grape seed extract |
References
- Aaliya, B.; Sunooj, K.V.; Navaf, M.; Akhila, P.P.; Sudheesh, C.; Mir, S.A.; Sabu, S.; Sasidharan, A.; Theingi Hlaing, M.; George, J. Recent trends in bacterial decontamination of food products by hurdle technology: A synergistic approach using thermal and non-thermal processing techniques. Food Res. Int. 2021, 147, 110514. [Google Scholar] [CrossRef] [PubMed]
- Novais, C.; Molina, A.K.; Abreu, R.M.V.; Santo-Buelga, C.; Ferreira, I.C.F.R.; Pereira, C.; Barros, L. Natural Food Colorants and Preservatives: A Review, a Demand, and a Challenge. J. Agric. Food Chem. 2022, 70, 2789–2805. [Google Scholar] [CrossRef] [PubMed]
- Coban, H.B. Organic acids as antimicrobial food agents: Applications and microbial productions. Bioprocess Biosyst. Eng. 2020, 43, 569–591. [Google Scholar] [CrossRef] [PubMed]
- Punia Bangar, S.; Suri, S.; Trif, M.; Ozogul, F. Organic Acids Production from Lactic Acid Bacteria: A Preservation Approach. Food Biosci. 2022, 46, 101615. [Google Scholar] [CrossRef]
- Ali, H.; Zulkali, M. Utilization of agro-residual ligno-cellulosic substances by using solid state fermentation: A Review. Hrvat. Časopis Prehrambenu Tehnol. Biotehnol. Nutr. 2011, 6, 5–12. [Google Scholar]
- Theron, M.M.; Lues, J.F. Organic acids and meat preservation: A review. Food Rev. Int. 2007, 23, 141–158. [Google Scholar] [CrossRef]
- Pellissery, A.J.; Vinayamohan, P.G.; Amalaradjou, M.A.R.; Venkitanarayanan, K. Spoilage bacteria and meat quality. In Meat Quality Analysis; Academic Press: Cambridge, MA, USA, 2020; pp. 307–334. [Google Scholar]
- Bonetti, A.; Tugnoli, B.; Rossi, B.; Giovagnoni, G.; Piva, A.; Grilli, E. Nature-identical compounds and organic acids reduce E. coli K88 growth and virulence gene expression in vitro. Toxins 2020, 12, 468. [Google Scholar] [CrossRef]
- Anyasi, T.; Jideani, A.; Edokpayi, J.; Anokwuru, C. Application of organic acids in food preservation. In Organic Acids, Characteristics, Properties and Synthesis; Vargas, C., Ed.; Nova Science Publishers: Hauppauge, NY, USA, 2017; pp. 1–47. [Google Scholar]
- Chahardoli, A.; Jalilian, F.; Farzaei, H.; Shokoohinia, Y. Chapter 26-Analysis of organic acids. In Recent Advances in Natural Products Analysis; Elsevier: Amsterdam, The Netherlands, 2020; pp. 767–823. [Google Scholar]
- Davidson, P.M.; Sofos, J.N.; Branen, A.L. Antimicrobials in Food; CRC Press: Boca Raton, FL, USA, 2005. [Google Scholar]
- Khan, S.H.; Iqbal, J. Recent advances in the role of organic acids in poultry nutrition. J. Appl. Anim. Res. 2016, 44, 359–369. [Google Scholar] [CrossRef]
- Ben Braïek, O.; Smaoui, S. Chemistry, safety, and challenges of the use of organic acids and their derivative salts in meat preservation. J. Food Qual. 2021, 2021, 6653190. [Google Scholar] [CrossRef]
- Noorzai, S.; Verbeek, C.J.R. Collagen: From Waste to Gold. In Biotechnological Applications of Biomass; Chapter 12; Intech Open: London, UK, 2020; pp. 1–36. [Google Scholar]
- Deng, Y.; Ma, L.; Mao, Y. Biological production of adipic acid from renewable substrates: Current and future methods. Biochem. Eng. J. 2016, 105, 16–26. [Google Scholar] [CrossRef]
- Makowski, Z.; Lipiński, K.; Mazur-Kuśnirek, M. The Effects of sodium butyrate, coated sodium butyrate, and butyric acid glycerides on nutrient digestibility, gastrointestinal function, and fecal microbiota in turkeys. Animals 2022, 12, 1836. [Google Scholar] [CrossRef] [PubMed]
- Behera, B.C.; Mishra, R.; Mohapatra, S. Microbial citric acid: Production, properties, application, and future perspectives. Food Front. 2021, 2, 62–76. [Google Scholar] [CrossRef]
- Guo, F.; Wu, M.; Dai, Z.; Zhang, S.; Zhang, W.; Dong, W.; Zhou, J.; Jiang, M.; Xin, F. Current advances on biological production of fumaric acid. Biochem. Eng. J. 2020, 153, 107397. [Google Scholar] [CrossRef]
- Stanojevic-Nikolic, S.; Dimic, G.; Mojovic, L.; Pejin, J.; Djukic-Vukovic, A.; Kocic-Tanackov, S. Antimicrobial activity of lactic acid against pathogen and spoilage microorganisms. J. Food Process. Preserv. 2015, 40, 990–998. [Google Scholar] [CrossRef]
- Chi, Z.; Wang, Z.P.; Wang, G.Y.; Khan, I.; Chi, Z.M. Microbial biosynthesis and secretion of l-malic acid and its applications. Crit. Rev. Biotechnol. 2016, 36, 99–107. [Google Scholar] [CrossRef] [PubMed]
- Ning, Y.; Yan, A.; Yang, K.; Wang, Z.; Li, X.; Jia, Y. Antibacterial activity of phenyllactic acid against Listeria monocytogenes and Escherichia coli by dual mechanisms. Food Chem. 2017, 228, 533–540. [Google Scholar] [CrossRef]
- Eş, I.; Khaneghah, A.M.; Hashemi, S.M.; Koubaa, M. Current advances in biological production of propionic acid. Biotechnol. Lett. 2017, 39, 635–645. [Google Scholar] [CrossRef]
- Xiang, Z.; Guan, H.; Zhao, X.; Xie, Q.; Xie, Z.; Cai, F.; Dang, R.; Li, M.; Wang, C. Dietary gallic acid as an antioxidant: A review of its food industry applications, health benefits, bioavailability, nano-delivery systems, and drug interactions. Food Res. Int. 2024, 180, 114068. [Google Scholar] [CrossRef] [PubMed]
- Nghiem, N.P.; Kleff, S.; Schwegmann, S. Succinic acid: Technology development and commercialization. Fermentation 2017, 3, 26. [Google Scholar] [CrossRef]
- Mani-López, E.; García, H.S.; López-Malo, A. Organic acids as antimicrobials to control Salmonella in meat and poultry products. Food Res. Int. 2012, 45, 713–721. [Google Scholar] [CrossRef]
- Lianou, A.; Koutsoumanis, K.P.; Sofos, J.N. Organic acids and other chemical treatments for microbial decontamination of food. In Microbial Decontamination in the Food Industry: Novel Methods and Applications; Demirci, A., Ngadi, M.O., Eds.; Woodhead Publishing Inc.: Oxford, UK, 2012; pp. 592–664. ISBN 978-0-85709-575-6. [Google Scholar]
- Abdelhamid, A.G.; El-Dougdoug, N.K. Controlling foodborne pathogens with natural antimicrobials by biological control and antivirulence strategies. Heliyon 2020, 6, e05020. [Google Scholar] [CrossRef] [PubMed]
- Dagnas, S.; Gauvry, E.; Onno, B.; Membré, J.M. Quantifying effect of lactic, acetic, and propionic acids on growth of molds isolated from spoiled bakery products. J. Food Prot. 2015, 78, 1689–1698. [Google Scholar] [CrossRef] [PubMed]
- Liu, F.; Wang, F.; Du, L.; Zhao, T.; Doyle, M.P.; Wang, D.; Zhang, X.; Sun, Z.; Xu, W. Antibacterial and antibiofilm activity of phenyllactic acid against Enterobacter cloacae. Food Control 2018, 84, 442–448. [Google Scholar] [CrossRef]
- Ng, W.K.; Koh, C.B. The utilization and mode of action of organic acids in the feeds of cultured aquatic animals. Rev. Aquac. 2017, 9, 342–368. [Google Scholar] [CrossRef]
- Mirhosseini, M.; Arjmand, V. Reducing pathogens by using zinc oxide nanoparticles and acetic acid in sheep meat. J. Food Prot. 2014, 77, 1599–1604. [Google Scholar] [CrossRef]
- Weissinger, W.R.; Mcwatters, K.H.; Beuchat, L.R. Evaluation of volatile chemical treatments for lethality to Salmonella on alfalfa seeds and sprouts. J. Food Prot. 2001, 64, 442–450. [Google Scholar] [CrossRef]
- Pangprasit, N.; Srithanasuwan, A.; Suriyasathaporn, W.; Pikulkaew, S.; Bernard, J.K.; Chaisri, W. Antibacterial activities of acetic acid against major and minor pathogens isolated from mastitis in dairy cows. Pathogens 2020, 9, 961. [Google Scholar] [CrossRef]
- Divya, K.; Vijayan, S.; George, T.K.; Jisha, M.S. Antimicrobial properties of chitosan nanoparticles: Mode of action and factors affecting activity. Fibers Polym. 2017, 18, 221–230. [Google Scholar] [CrossRef]
- Lastauskienė, E.; Zinkevičienė, A.; Girkontaitė, I.; Kaunietis, A.; Kvedarienė, V. Formic acid and acetic acid induce a programmed cell death in pathogenic Candida species. Curr. Microbiol. 2014, 69, 303–310. [Google Scholar] [CrossRef]
- Ivanišová, E.; Meňhartová, K.; Terentjeva, M.; Harangozo, Ľ.; Kántor, A.; Kačániová, M. The evaluation of chemical, antioxidant, antimicrobial and sensory properties of kombucha tea beverage. J. Food Sci. Technol. 2020, 57, 1840–1846. [Google Scholar] [CrossRef]
- Kang, H.; Park, Y.; Go, S. Growth inhibition of a phytopathogenic fungus, Colletotrichum species by acetic acid. Microbiol. Res. 2003, 158, 321–326. [Google Scholar] [CrossRef]
- Hassan, R.A.; Sand, M.I.; El-Kadi, S.M. Effect of some organic acids on fungal growth and their toxins production. J. Agric. Chem. Biotechnol. 2012, 3, 391–397. [Google Scholar] [CrossRef]
- Quattrini, M.; Liang, N.; Fortina, M.G.; Xiang, S.; Curtis, J.M.; Gänzle, M. Exploiting synergies of sourdough and antifungal organic acids to delay fungal spoilage of bread. Int. J. Food Microbiol. 2019, 302, 8–14. [Google Scholar] [CrossRef] [PubMed]
- Moon, Y.S.; Kim, H.M.; Chun, H.S.; Lee, S.E. Organic acids suppress aflatoxin production via lowering expression of aflatoxin biosynthesis-related genes in Aspergillus flavus. Food Control 2018, 88, 207–216. [Google Scholar] [CrossRef]
- Adili, L.; Roufegarinejad, L.; Tabibiazar, M.; Hamishehkar, H.; Alizadeh, A. Development and characterization of reinforced ethyl cellulose based oleogel with adipic acid: Its application in cake and beef burger. LWT Food Sci. Technol. 2020, 126, 109277. [Google Scholar] [CrossRef]
- Zehetmeyer, G.; Meira, S.M.M.M.; Scheibel, J.M.; de Oliveira, R.V.B.; Brandelli, A.; Soares, R.M.D. Influence of melt processing on biodegradable nisin-PBAT films intended for active food packaging applications. J. Appl. Polym. Sci. 2016, 133, 1–10. [Google Scholar] [CrossRef]
- Venkatesan, R.; Rajeswari, N. ZnO/PBAT nanocomposite films: Investigation on the mechanical and biological activity for food packaging. Polym. Adv. Technol. 2017, 28, 20–27. [Google Scholar] [CrossRef]
- Moraes Filho, L.E.P.T.; Andrade, M.F.; Freitas, L.F.; Palha, M.D.L.A.P.F.; Vinhas, G.M. Development and Characterization of Poly(Butylene Adipate-Co-terephthalate) (PBAT) Antimicrobial Films with Clove and Cinnamon Essential Oils. Food Process. Preserv. 2022, 46, e16489. [Google Scholar] [CrossRef]
- Venkatesan, R.; Sivaprakash, P.; Kim, I.; Eldesoky, G.; Kim, S. Tannic acid as a crosslinking agent in poly (butylene adipate-co-terephthalate) composite films enhanced with carbon nanoparticles: Processing, characterization, and antimicrobial activities for food packaging. J. Environ. Chem. Eng. 2023, 11, 110194. [Google Scholar] [CrossRef]
- Vicedo, B.; de la O Leyva, M.; Flors, V.; Finiti, I.; Del Amo, G.; Walters, D.; Real, M.D.; García-Agustín, P.; González-Bosch, C. Control of the phytopathogen Botrytis cinerea using adipic acid monoethyl ester. Arch. Microbiol. 2006, 184, 316–326. [Google Scholar] [CrossRef]
- Flors, V.; Miralles, C.; González-Bosch, C.; Carda, M.; García-Agustín, P. Three novel synthetic amides of adipic acid protect Capsicum anuum plants against the necrotrophic pathogen Alternaria solani. Physiol. Mol. Plant Pathol. 2003, 63, 151–158. [Google Scholar] [CrossRef]
- Klinmalai, P.; Srisa, A.; Laorenza, Y.; Katekhong, W.; Harnkarnsujarit, N. Antifungal and plasticization effects of carvacrol in biodegradable poly (lactic acid) and poly (butylene adipate terephthalate) blend films for bakery packaging. LWT 2021, 152, 112356. [Google Scholar] [CrossRef]
- Agnihotri, S.; Yin, D.-M.; Mahboubi, A.; Sapmaz, T.; Varjani, S.; Qiao, W.; Koseoglu-Imer, D.Y.; Taherzadeh, M.J. A glimpse of the world of volatile fatty acids production and application: A review. Bioengineered 2022, 13, 1249–1275. [Google Scholar] [CrossRef]
- Sun, C.Q.; O’Connor, C.J.; Turner, S.J.; Lewis, G.D.; Stanley, R.A.; Roberton, A.M. The effect of pH on the inhibition of bacterial growth by physiological concentrations of butyric acid: Implications for neonates fed on suckled milk. Chem. Interact. 1998, 113, 117–131. [Google Scholar] [CrossRef]
- Namkung, H.; Yu, H.; Gong, J.; Leeson, S. Antimicrobial activity of butyrate glycerides toward Salmonella typhimurium and Clostridium perfringens. Poult. Sci. 2011, 90, 2217–2222. [Google Scholar] [CrossRef] [PubMed]
- Liu, S.; Ruan, W.; Li, J.; Xu, H.; Wang, J.; Gao, Y.; Wang, J. Biological control of phytopathogenic fungi by fatty acids. Mycopathologia 2008, 166, 93–102. [Google Scholar] [CrossRef]
- Ruiz-Rico, M.; Fuentes, C.; Pérez-Esteve, É.; Jiménez-Belenguer, A.I.; Quiles, A.; Marcos, M.D.; Martínez-Máñez, R.; Barat, J.M. Bactericidal activity of caprylic acid entrapped in mesoporous silica nanoparticles. Food Control 2015, 56, 77–85. [Google Scholar] [CrossRef]
- Nair, M.K.M.; Vasudevan, P.; Hoagland, T.; Venkitanarayanan, K. Inactivation of Escherichia coli O157:H7 and Listeria monocytogenes in milk by caprylic acid and monocaprylin. Food Microbiol. 2004, 21, 611–616. [Google Scholar] [CrossRef]
- Wang, J.; Ma, M.; Yang, J.; Chen, L.; Yu, P.; Wang, J. In vitro antibacterial activity and mechanism of monocaprylin against Escherichia coli and Staphylococcus aureus. J. Food Prot. 2018, 81, 1988–1996. [Google Scholar] [CrossRef]
- Wang, W.; Wang, R.; Zhang, G.; Chen, F.; Xu, B. In vitro antibacterial activities and mechanisms of action of fatty acid monoglycerides against four foodborne bacteria. J. Food Prot. 2020, 83, 331–337. [Google Scholar] [CrossRef]
- Jadhav, A.; Mortale, S.; Halbandge, S.; Jangid, P.; Patil, R.; Gade, W.; Kharat, K.; Karuppayil, S.M. The dietary food components capric acid and caprylic acid inhibit virulence factors in Candida albicans through multitargeting. J. Med. Food 2017, 20, 1083–1090. [Google Scholar] [CrossRef] [PubMed]
- Rosenblatt, J.; Reitzel, R.A.; Vargas-Cruz, N.; Chaftari, A.M.; Hachem, R.; Raad, I. Caprylic and polygalacturonic acid combinations for eradication of microbial organisms embedded in biofilm. Front. Microbiol. 2017, 8, 1999. [Google Scholar] [CrossRef] [PubMed]
- Akula, S.T.; Nagaraja, A.; Ravikanth, M.; Kumar, N.G.R.; Kalyan, Y.; Divya, D. Antifungal efficacy of LA and caprylic acid—Derivatives of virgin coconut oil against Candida albicans. Biomed. Biotechnol. Res. J. 2021, 5, 229–234. [Google Scholar] [CrossRef]
- López-Velázquez, J.G.; Ayón-Reyna, L.E.; Vega-García, M.O.; López-Angulo, G.; López-López, M.E.; López-Zazueta, B.A.; Delgado-Vargas, F. Caprylic acid in Vitex mollis fruit and its inhibitory activity against a thiabendazole-resistant Colletotrichum gloeosporioides strain. Pest. Manag. Sci. 2022, 78, 5271–5280. [Google Scholar] [CrossRef]
- Sorrells, K.M.; Enigl, D.C.; Hatfield, J.R. Effect of pH, acidulant, time, and temperature on the growth and survival of Listeria monocytogenes. J. Food Prot. 1989, 52, 571–573. [Google Scholar] [CrossRef]
- Brackett, R.E. Effects of various acids on growth and survival of Yersinia Enterocolitica. J. Food Prot. 1987, 50, 598–602. [Google Scholar] [CrossRef]
- In, Y.W.; Kim, J.J.; Kim, H.J.; Oh, S.W. Antimicrobial activities of acetic acid, citric acid and lactic acid against Shigella species. J. Food Saf. 2013, 33, 79–85. [Google Scholar] [CrossRef]
- Seo, S.; Jung, D.; Wang, X.; Seo, D.J.; Lee, M.H.; Lee, B.-H.; Choi, C. Combined effect of lactic acid bacteria and citric acid on Escherichia coli O157:H7 and Salmonella typhimurium. Food Sci. Biotechnol. 2013, 22, 1171–1174. [Google Scholar] [CrossRef]
- Al-Rousan, W.M.; Olaimat, A.N.; Osaili, T.M.; Al-Nabulsi, A.A.; Ajo, R.Y.; Holley, R.A. Use of acetic and citric acids to inhibit Escherichia coli O157:H7, Salmonella typhimurium and Staphylococcus aureus in tabbouleh salad. Food Microb. 2018, 73, 61–66. [Google Scholar] [CrossRef]
- Zhao, X.; Zhen, Z.; Wang, X.; Guo, N. Synergy of a combination of nisin and citric acid against Staphylococcus aureus and Listeria monocytogenes. Food Addit. Contam. Part A Chem. Anal. Control. Expo Risk Assess 2017, 34, 2058–2068. [Google Scholar] [CrossRef]
- Shokri, H. Evaluation of inhibitory effects of citric and tartaric acids and their combination on the growth of Trichophyton mentagrophytes, Aspergillus fumigatus, Candida albicans, and Malassezia furfur. Comp. Clin. Path. 2011, 20, 543–545. [Google Scholar] [CrossRef]
- Nielsen, M.K.; Arneborg, N. The effect of citric acid and pH on growth and metabolism of anaerobic Saccharomyces cerevisiae and Zygosaccharomyces bailii cultures. Food Microbiol. 2007, 24, 101–105. [Google Scholar] [CrossRef]
- Kim, Y.J.; Kim, M.H.; Song, K.B. Efficacy of aqueous chlorine dioxide and fumaric acid for inactivating pre-existing microorganisms and Escherichia coli O157:H7, Salmonella typhimurium, and Listeria monocytogenes on broccoli sprouts. Food Control 2009, 20, 1002–1005. [Google Scholar] [CrossRef]
- Tango, C.N.; Mansur, A.R.; Oh, D.-H. Fumaric Acid and slightly acidic electrolyzed water inactivate gram-positive and gram-negative foodborne pathogens. Microorganisms 2015, 3, 34–46. [Google Scholar] [CrossRef]
- Chen, X.; Tango, C.N.; Daliri, E.B.; Oh, S.Y.; Oh, D.H. Disinfection efficacy of slightly acidic electrolyzed water combined with chemical treatments on fresh fruits at the industrial scale. Foods 2019, 8, 497. [Google Scholar] [CrossRef] [PubMed]
- Fernández, M.; Rodriguez, A.; Fulco, M.; Soteras, T.; Mozgovoj, M.; Cap, M. Effects of lactic, malic and fumaric acids on Salmonella spp. counts and on chicken meat quality and sensory characteristics. J. Food Sci. Technol. 2021, 58, 3817–3824. [Google Scholar] [CrossRef]
- Jeon, M.-J.; Ha, J.-W. Inactivating Foodborne Pathogens in Apple Juice by Combined Treatment with Fumaric Acid and Ultraviolet-A Light, and Mechanisms of Their Synergistic Bactericidal Action. Food Microbiol. 2020, 87, 103387. [Google Scholar] [CrossRef]
- Khan, I.; Tango, C.N.; Miskeen, S.; Oh, D.-H. Evaluation of Nisin-Loaded Chitosan-Monomethyl Fumaric Acid Nanoparticles as a Direct Food Additive. Carbohydr. Polym 2018, 184, 100–107. [Google Scholar] [CrossRef]
- Beuchat, L.R. Influence of Organic Acids on Heat Resistance Characteristics of Talaromyces flavus Ascospores. Int. J. Food Microbiol. 1988, 6, 97–105. [Google Scholar] [CrossRef]
- Kotula, K.L.; Thelappurate, R. Microbiological and Sensory Attributes of Retail Cuts of Beef Treated with Acetic and Lactic Acid Solutions. JFP 1994, 57, 665–670. [Google Scholar] [CrossRef]
- Dan, S.D.; Mihaiu, M.; Reget, O.; Tăbăran, A. Residual Antimicrobial Effect of Weak Organic Acids on Spoilage Psychrotrophs at Pig Carcasses. Lucr. Științifice USAMV Iași Ser. Med. Vet. 2017, 60, 258–264. [Google Scholar]
- Laury, A.M.; Alvarado, M.V.; Nace, G.; Alvarado, C.Z.; Brooks, J.C.; Echeverry, A.; Brashears, M.M. Validation of a Lactic Acid– and Citric Acid–Based Antimicrobial Product for the Reduction of Escherichia coli O157:H7 and Salmonella on Beef Tips and Whole Chicken Carcasses. JFP 2009, 72, 2208–2211. [Google Scholar] [CrossRef]
- Zhang, S. The Effects of Various Disinfectants against Listeria monocytogenes Fresh-Cut Vegetables. Food Microbiol. 1996, 13, 311–321. [Google Scholar] [CrossRef]
- Amrutha, B.; Sundar, K.; Shetty, P.H. Effect of Organic Acids on Biofilm Formation and Quorum Signaling of Pathogens from Fresh Fruits and Vegetables. Microb. Pathog. 2017, 111, 156–162. [Google Scholar] [CrossRef] [PubMed]
- Kavčič, S.; Knez, Ž.; Leitgeb, M. Antimicrobial Activity of N-Butyl Lactate Obtained via Enzymatic Esterification of Lactic Acid with n-Butanol in Supercritical Trifluoromethane. J. Supercrit. Fluids 2014, 85, 143–150. [Google Scholar] [CrossRef]
- Lind, H.; Jonsson, H.; Schnürer, J. Antifungal effect of dairy propionibacteria–contribution of organic acids. Int. J. Food Microbiol. 2003, 98, 157–165. [Google Scholar] [CrossRef]
- Peláez, A.L.; Cataño, C.S.; Yepes, E.Q.; Villarroel, R.G.; de Antoni, G.; Giannuzzi, L. Inhibitory activity of lactic and acetic acid on Aspergillus Flavus growth for food preservation. Food Control 2012, 24, 177–183. [Google Scholar] [CrossRef]
- Kövilein, A.; Kubisch, C.; Cai, L.; Ochsenreither, K. Malic acid production from renewables: A review. J. Chem. Technol. Biotechnol. 2020, 95, 513–526. [Google Scholar] [CrossRef]
- Raybaudi-Massilia, R.M.; Mosqueda-Melgar, J.; Martín-Belloso, O. Antimicrobial activity of malic acid against Listeria monocytogenes, Salmonella enteritidis and Escherichia coli O157:H7 in apple, pear and melon juices. Food Control 2009, 2, 105–112. [Google Scholar] [CrossRef]
- Eswaranandam, S.; Hettiarachchy, N.S.; Johnson, M.G. Antimicrobial activity of citric, lactic, malic, or tartaric acids and nisin-incorporated soy protein film against Listeria monocytogenes, Escherichia coli O157:H7, and Salmonella gaminara. J. Food Sci. 2006, 69, FMS79–FMS84. [Google Scholar] [CrossRef]
- Olaimat, A.N.; Al-Holy, M.A.; Ghoush, M.H.A.; Al-Nabulsi, A.A.; Qatatsheh, A.A.; Shahbaz, H.M.; Osaili, T.M.; Holley, R.A. The use of malic and acetic acids in washing solution to control Salmonella spp. on chicken breast. J. Food Sci. 2018, 83, 2197–2203. [Google Scholar] [CrossRef] [PubMed]
- Hashemi, S.M.B.; Jafarpour, D. Ultrasound and malic acid treatment of sweet lemon juice: Microbial inactivation and quality changes. J. Food Process. Preserv. 2020, 44, 14866. [Google Scholar] [CrossRef]
- Borah, H.J.; Borah, A.; Yadav, A.; Hazarika, S. Extraction of malic acid from Dillenia Indica in organic solvents and its antimicrobial activity. Sep. Sci. Technol. 2022, 58, 314–325. [Google Scholar] [CrossRef]
- Pintado, C.M.B.S.; Ferreira, M.A.S.S.; Sousa, I. Control of pathogenic and spoilage microorganisms from cheese surface by whey protein films containing malic acid, nisin and natamycin. Food Control 2010, 21, 240–246. [Google Scholar] [CrossRef]
- Dilshad, E.; Bibi, M.; Sheikh, N.A.; Tamrin, K.F.; Mansoor, Q.; Maqbool, Q.; Nawaz, M. Synthesis of functional silver nanoparticles and microparticles with modifiers and evaluation of their antimicrobial, anticancer, and antioxidant activity. J. Funct. Biomater. 2020, 11, 76. [Google Scholar] [CrossRef]
- Zheng, R.; Zhao, T.; Hung, Y.C.; Adhikari, K. Evaluation of bactericidal effects of phenyllactic acid on Escherichia coli O157:H7 and Salmonella typhimurium on beef meat. J. Food Prot. 2019, 82, 2016–2022. [Google Scholar] [CrossRef]
- Jiang, Y.-H.; Yang, L.-Y.; Xin, W.-G.; Zhang, Q.-L. Combined antibacterial and antibiofilm activity of phenyllactic acid and bacteriocin XJS01 against Shigella flexneri. Food Biosci. 2022, 45, 101512. [Google Scholar] [CrossRef]
- Kim, Y.; Cho, J.-Y.; Kuk, J.-H.; Moon, J.-H.; Cho, J.-I.; Kim, Y.-C.; Park, K.-H. Identification and antimicrobial activity of phenylacetic acid produced by Bacillus licheniformis isolated from fermented soybean, chungkook-jang. Curr. Microbiol. 2004, 48, 312–317. [Google Scholar] [CrossRef]
- Lavermicocca, P.; Valerio, F.; Visconti, A. Antifungal activity of phenyllactic acid against molds isolated from bakery products. Appl. Environ. Microbiol. 2003, 69, 634–640. [Google Scholar] [CrossRef]
- Gerez, C.L.; Carbajo, M.S.; Rollan, G.; Torres Leal, G.; Font de Valdez, G. Inhibition of citrus fungal pathogens by using lactic acid bacteria. J. Food Sci. 2010, 75, M354–M359. [Google Scholar] [CrossRef]
- Svanström, Å.; Boveri, S.; Boström, E.; Melin, P. The lactic acid bacteria metabolite phenyllactic acid inhibits both radial growth and sporulation of filamentous fungi. BMC Res. Notes 2013, 6, 464. [Google Scholar] [CrossRef]
- Fan, W.; Li, B.; Du, N.; Hui, T.; Cao, Y.; Li, X.; Ren, H. Energy metabolism as the target of 3-phenyllactic acid against Rhizopus oryzae. Int. J. Food Microbiol. 2022, 369, 109606. [Google Scholar] [CrossRef]
- Al-Lahham, S.H.; Peppelenbosch, M.P.; Roelofsen, H.; Vonk, R.J.; Venema, K. Biological effects of propionic acid in humans; metabolism, potential applications and underlying mechanisms. Biochim. Biophys. Acta 2010, 1801, 1175–1183. [Google Scholar] [CrossRef] [PubMed]
- Pilevar, Z.; Mousavi Khaneghah, A.; Hosseini, H.; Ranaei, V. Propionic Acid: Method of production, current state and perspectives. Food Technol. Biotechnol. 2020, 58, 115–127. [Google Scholar]
- Haque, M.N.; Chowdhury, R.; Islam, K.; Akbar, M.A. Propionic acid is an alternative to antibiotics in poultry diet. Bangladesh J. Anim. Sci. 1970, 38, 115–122. [Google Scholar] [CrossRef]
- Seo, Y.; Sung, M.; Hwang, J.; Yoon, Y. Minimum Inhibitory Concentration (MIC) of Propionic Acid, Sorbic Acid, and Benzoic Acid against Food Spoilage Microorganisms in Animal Products to Use MIC as Threshold for Natural Preservative Production. Food Sci. Anim. Resour. 2023, 43, 319–330. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Dai, A.; Huang, S.; Kuo, S.; Shu, M.; Tapia, C.P.; Yu, J.; Two, A.; Zhang, H.; Gallo, R.L.; et al. Propionic acid and its esterified derivative suppress the growth of methicillin-resistant Staphylococcus aureus USA300. Benef. Microbes 2014, 5, 161–168. [Google Scholar] [CrossRef] [PubMed]
- Antone, U.; Ciprovica, I.; Zolovs, M.; Scerbaka, R.; Liepins, J. Propionic acid fermentation—Study of substrates, strains, and antimicrobial properties. Fermentation 2022, 9, 26. [Google Scholar] [CrossRef]
- Dijksterhuis, J.; Meijer, M.; van Doorn, T.; Houbraken, J.; Bruinenberg, P. The preservative propionic acid differentially affects survival of conidia and germ tubes of feed spoilage fungi. Int. J. Food Microbiol. 2019, 306, 108258. [Google Scholar] [CrossRef]
- Sorrentino, E.; Succi, M.; Tipaldi, L.; Pannella, G.; Maiuro, L.; Sturchio, M.; Coppola, R.; Tremonte, P. Antimicrobial activity of gallic acid against food-related Pseudomonas strains and its use as biocontrol tool to improve the shelf life of fresh black truffles. Int. J. Food Microbiol. 2018, 266, 183–189. [Google Scholar] [CrossRef]
- Alkan, D.; Aydemir, L.Y.; Arcan, I.; Yavuzdurmaz, H.; Atabay, H.I.; Ceylan, C.; Yemenicioglu, A. Development of flexible antimicrobial packaging materials against Campylobacter jejuni by incorporation of gallic acid into zein-based films. J. Agric. Food. Chem. 2011, 59, 11003–11010. [Google Scholar] [CrossRef] [PubMed]
- Borges, A.; Ferreira, C.; Saavedra, M.J.; Simões, M. Antibacterial activity and mode of action of ferulic and gallic acids against pathogenic bacteria. Microb. Drug Resist. 2013, 19, 256–265. [Google Scholar] [CrossRef]
- Shao, D.; Li, J.; Li, J.; Tang, R.; Liu, L.; Shi, J.; Huang, Q.; Yang, H. Inhibition of gallic acid on the growth and biofilm formation of Escherichia coli and Streptococcus mutans. J. Food Sci. 2015, 80, 1299–1305. [Google Scholar] [CrossRef] [PubMed]
- Alves, C.T.; Ferreira, I.C.; Barros, L.; Silva, S.; Azeredo, J.; Henriques, M. Antifungal activity of phenolic compounds identified in flowers from Northeastern Portugal against Candida species. Future Microbiol. 2014, 9, 139–146. [Google Scholar] [CrossRef]
- Fujita, K.; Kubo, I. Antifungal activity of octyl gallate. Int. J. Food Microbiol. 2002, 79, 193–201. [Google Scholar] [CrossRef]
- Karpova, N.; Shagdarova, B.; Lunkov, A.; Ilina, A.; Varlamov, V. Antifungal action of chitosan in combination with fungicides in vitro and chitosan conjugate with gallic acid on tomatoes against Botrytis cinerea. Biotechnol. Lett. 2021, 43, 1565–1574. [Google Scholar] [CrossRef] [PubMed]
- Cox, N.A.; Mercuri, A.J.; Juven, B.J.; Thomson, J.E.; Chew, V. Evaluation of succinic acid and heat to improve the microbiological quality of poultry meat. J. Food Sci. 1974, 39, 985–987. [Google Scholar] [CrossRef]
- Gao, Z.; Shao, J.; Sun, H.; Zhong, W.; Zhuang, W.; Zhang, Z. Evaluation of different kinds of organic acids and their antibacterial activity in Japanese Apricot fruits. Afr. J. Agric. Res. 2012, 7, 4911–4918. [Google Scholar] [CrossRef]
- Khalil, A.A.; Deraz, S.F.; Elrahman, S.A.; El-Fawal, G. Enhancement of mechanical properties, microstructure, and antimicrobial activities of zein films cross-linked using succinic anhydride, eugenol, and citric Acid. Prep. Biochem. Biotechnol. 2015, 45, 551–567. [Google Scholar] [CrossRef]
- Radkowski, M.; Zdrodowska, B.; Gomółka-Pawlicka, M. Effect of succinic acid on elimination of Salmonella in chicken meat. J. Food Prot. 2018, 81, 1491–1495. [Google Scholar] [CrossRef]
- Mikaelyan, A.R.; Babayan, B.G.; Grigogerez, A.L.; Grigoryan, A.M.; Asatryan, N.L.; Melkumyan, M.A. Tartaric acid new derivatives as prospective and safe alternative to antimicrobials for food products Packing. FFHD 2024, 14, 33. [Google Scholar] [CrossRef]
- Wu, W.; Zhang, J.; Li, T.; Hu, Z.; Huang, S.; Lin, M.; Xie, Y.; Yu, Z. Antimicrobial Effect of Plasma-Activated Water Combined with Tartaric Acid against Staphylococcus Aureus and Its Application on Fresh-Cut Asparagus Lettuce. Food Biosci. 2024, 60, 104293. [Google Scholar] [CrossRef]
- Tavsanli, H.; Irkin, R.; Kisadere, I. Effects of Addition of Different Organic Acids to White Cheese Brine on Microflora and Pathogen Listeria Monocytogenes. Kafkas Univ. Vet. Fak. Derg. 2018, 25, 201–207. [Google Scholar]
- Karpiński, T.M.; Ożarowski, M. Plant Organic Acids as Natural Inhibitors of Foodborne Pathogens. Appl. Sci. 2024, 14, 6340. [Google Scholar] [CrossRef]
- Guo, Y.; Chen, X.; Gong, P.; Wang, R.; Han, A.; Deng, Z.; Qi, Z.; Long, H.; Wang, J.; Yao, W.; et al. Advances in the Role and Mechanisms of Essential Oils and Plant Extracts as Natural Preservatives to Extend the Postharvest Shelf Life of Edible Mushrooms. Foods 2023, 12, 801. [Google Scholar] [CrossRef]
- Chowdhury, M.A.K.; Song, H.; Liu, Y.; Bunod, J.-D.; Dong, X.-H. Effects of Microencapsulated Organic Acid and Their Salts on Growth Performance, Immunity, and Disease Resistance of Pacific White Shrimp Litopenaeus Vannamei. Sustainability 2021, 13, 7791. [Google Scholar] [CrossRef]
- Kim, T.-K.; Hwang, K.-E.; Lee, M.-A.; Paik, H.-D.; Kim, Y.-B.; Choi, Y.-S. Quality Characteristics of Pork Loin Cured with Green Nitrite Source and Some Organic Acids. Meat Sci. 2019, 152, 141–145. [Google Scholar] [CrossRef]
- Mohan, A.; Purohit, A.S. Anti-Salmonella Activity of Pyruvic and Succinic Acid in Combination with Oregano Essential Oil. Food Control 2020, 110, 106960. [Google Scholar] [CrossRef]
- Dogruyol, H.; Mol, S.; Cosansu, S. Increased Thermal Sensitivity of Listeria monocytogenes in Sous-Vide Salmon by Oregano Essential Oil and Citric Acid. Food Microbiol. 2020, 90, 103496. [Google Scholar] [CrossRef]
- Lin, Y.T.; Labbe, R.G.; Shetty, K. Inhibition of Vibrio parahaemolyticus in Seafood Systems Using Oregano and Cranberry Phytochemical Synergies and Lactic Acid. IFSET 2005, 6, 453–458. [Google Scholar] [CrossRef]
- Kim, J.-H.; Kwon, K.-H.; Oh, S.-W. Effects of Malic Acid or/and Grapefruit Seed Extract for the Inactivation of Common Food Pathogens on Fresh-Cut Lettuce. Food Sci. Biotechnol. 2016, 25, 1801–1804. [Google Scholar] [CrossRef] [PubMed]
- Satterlee, T.; McDonough, C.M.; Gold, S.E.; Chen, C.; Glenn, A.E.; Pokoo-Aikins, A. Synergistic Effects of Essential Oils and Organic Acids against Aspergillus flavus Contamination in Poultry Feed. Toxins 2023, 15, 635. [Google Scholar] [CrossRef] [PubMed]
- Popescu, P.-A.; Palade, L.M.; Nicolae, I.-C.; Popa, E.E.; Miteluț, A.C.; Drăghici, M.C.; Matei, F.; Popa, M.E. Chitosan-Based Edible Coatings Containing Essential Oils to Preserve the Shelf Life and Postharvest Quality Parameters of Organic Strawberries and Apples during Cold Storage. Foods 2022, 11, 3317. [Google Scholar] [CrossRef] [PubMed]
- Jin, T.Z.; Fan, X.; Mukhopadhyay, S. Antimicrobial Coating with Organic Acids and Essential Oil for the Enhancement of Safety and Shelf Life of Grape Tomatoes. Int. J. Food Microbiol. 2022, 378, 109827. [Google Scholar] [CrossRef]
- Khaledian, Y.; Pajohi-Alamoti, M.; Bazargani-Gilani, B. Development of Cellulose Nanofibers Coating Incorporated with Ginger Essential Oil and Citric Acid to Extend the Shelf Life of Ready-to-cook Barbecue Chicken. J. Food Process. Preserv. 2019, 43, e14114. [Google Scholar] [CrossRef]
- Wang, J.; Cui, Z.; Li, Y.; Cao, L.; Lu, Z. Techno-Economic Analysis and Environmental Impact Assessment of Citric Acid Production through Different Recovery Methods. J. Clean. Prod. 2020, 249, 119315. [Google Scholar] [CrossRef]
- Becker, M.; Kohlheb, N.; Hunger, S.; Eschrich, S.; Müller, R.; Aurich, A. Early-stage Sustainability Assessment of Biotechnological Processes: A Case Study of Citric Acid Production. ELS 2020, 20, 90–103. [Google Scholar] [CrossRef]
- Budsberg, E.; Morales-Vera, R.; Crawford, J.T.; Bura, R.; Gustafson, R. Production Routes to Bio-Acetic Acid: Life Cycle Assessment. Biotechnol. Biofuels 2020, 13, 154. [Google Scholar] [CrossRef]
- Li, Y.; Bhagwat, S.S.; Cortés-Peña, Y.R.; Ki, D.; Rao, C.V.; Jin, Y.-S.; Guest, J.S. Sustainable Lactic Acid Production from Lignocellulosic Biomass. ACS Sustain. Chem. Eng. 2021, 9, 1341–1351. [Google Scholar] [CrossRef]
- Kung, L.; Smith, M.L.; Benjamim Da Silva, E.; Windle, M.C.; Da Silva, T.C.; Polukis, S.A. An Evaluation of the Effectiveness of a Chemical Additive Based on Sodium Benzoate, Potassium Sorbate, and Sodium Nitrite on the Fermentation and Aerobic Stability of Corn Silage. J. Dairy Sci. 2018, 101, 5949–5960. [Google Scholar] [CrossRef]


{kind=link}
| Organic Acid | Chemical Formula | Chemical Structure | Molecular Weight (g/mol) | pKa | Solubility (in Water) (g/L) | GRAS Status | References |
|---|---|---|---|---|---|---|---|
| Acetic acid | C2H4O2 | 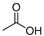 | 60.05 | 4.75 | Miscible | Approved | [14] |
| Adipic acid | C6H10O4 |  | 146.14 | 4.43, 5.41 | 24 g/L | Approved | [15] |
| Butyric acid | C4H8O2 | 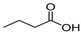 | 88.11 | 4.82 | Miscible | Approved | [16] |
| Caprylic acid | C8H16O2 |  | 144.21 | 4.89 | 0.68 | Approved | [4] |
| Citric acid | C6H8O7 |  | 192.12 | 3.12, 4.76, 6.39 | 590 g/L | Approved | [17] |
| Fumaric acid | C4H4O4 |  | 116.07 | 3.86, 15.01 | Miscible | Approved | [18] |
| Lactic acid | C3H6O3 |  | 90.07 | 3.03, 4.44 | 6.3 g/L | Approved | [19] |
| Malic acid | C4H6O5 | 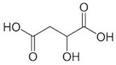 | 134.08 | 3.40, 5.20, 14.50 | 558 g/L | Approved | [20] |
| Phenyllactic acid | C8H8O2 |  | 166.17 | 4.31 | 15 g/L | Not Approved | [21] |
| Propionic acid | C3H6O2 |  | 74.00 | 4.87 | Miscible | Approved | [22] |
| Gallic acid | C7H6O5 | 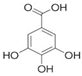 | 170.12 | 4.40 | 11.9 g/L | Approved | [23] |
| Succinic acid | C4H6O4 | 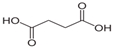 | 118.88 | 4.61, 5.61 | 80 g/L | Approved | [24] |
| Tartaric acid | C4H6O6 | 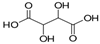 | 158.08 | 2.89, 4.40 | Miscible | Approved | [10] |
| Food Product | Preservation Method | Target Microorganism | Effects | References |
|---|---|---|---|---|
| Meat | Fermented spinach (0.08%) with organic acid (ascorbic acid, malic acid, citric acid, and tartaric acid (0.06%)) | - | Enhanced redness, lowered the residual nitrite level, prevented oxidation | [123] |
| Chicken | Pyruvic acid (PA) (6%) and succinic acid (SA) (1%) + oregano essential oil (OEO) (0.08%) | S. typhimurium | PA + EO: 1.4 log10 CFU/g reduction, SA + EO: 1.0 log10 CFU/g reduction | [124] |
| Salmon | Citric acid (0.5%), OEO (1%), citric acid + OEO (combination), cooked at 55–62.5 °C | L. monocytogenes (ATCC 7644) | Combination of citric acid and oregano oil reduced D-values significantly, with z-values ranging from 5.50 °C (control) to 6.92 °C (combination), reducing inactivation time | [125] |
| Sea food | Oregano extract + Cranberry extract (0.1 mg/mL phenolic compound) + lactic acid (pH 6) | Vibrio parahaemolyticus | Complete inhibition of V. parahaemolyticus after 8 days at 4 °C | [126] |
| Lettuce | Malic Acid + grapefruit seed extract (1% MA + 0.5% GSE) at 5 °C for 14 days | E. coli O157:H7 (ATCC 43895, ATCC 35150, and ATCC 43894), S. typhimurium (ATCC 19585, ATCC 6994, and ATCC 14028), L. monocytogenes (ATCC 7644, ATCC 19111, and ATCC 19115) | Reduction (log CFU/g): E. coli: 4.96; Salmonella: 4.80; L. monocytogenes: 3.95 | [127] |
| Corn Kernels | Combination of organic acids (acetic acid 1500 mg/kg, butyric acid 1760 mg/kg, propionic acid 2222 mg/kg) + cinnamon oil (8000 µL/L air) | A. flavus (NRRL3357) | Complete inhibition: 1/2 MIC (750 mg/kg AA, 880 mg/kg BA, 1111 mg/kg PA) with 1/4 MIC (2000 µL/L air) of cinnamon oil | [128] |
| Strawberries | 2% chitosan, (1% w/v), acetic acid, sea buckthorn essentials oil (7.5%) dipping for 20 s and store (4 °C and 8 °C) | - | Provided the lowest count of molds and yeasts until day 7 of analysis; preserved the ascorbic acid, total polyphenol content, and antioxidant activity compared to control | [129] |
| Grape Tomatoes | Chitosan (1%), acetic, lactic, levulinic acids, 2% allyl isothiocyanate (AIT) | Salmonella enterica (S. Panama 19,454, S. Poona 953, and S. Stanley H0558), L. monocytogenes | Chitosan reduced S. enterica to 1.28 log CFU/tomato (Day 1); Chitosan + AIT reduced it to <0.70 log CFU/tomato (Day 1); both treatments made S. enterica undetectable from day 2 to day 21. L. monocytogenes became undetectable by Day 6. | [130] |
| Ready-to-cook barbecue chicken | Edible cellulose nanofiber coating incorporated with 2% ginger essential oil (GEO) and 1% citric acid | Enterobacteriaceae spp., Pseudomonas spp., yeasts and molds, (total viable count) TVC | Reduction in microbial counts: TVC: 4.87 log CFU/g; Enterobacteriaceae spp.: 4.75 log CFU/g; Pseudomonas spp.: 4.91 log CFU/g; and PTC: 4.93 log CFU/g. Other treatments significantly reduced the yeast and mold count at p < 0.05 and extended shelf life by up to 6 days. | [131] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sorathiya, K.B.; Melo, A.; Hogg, M.C.; Pintado, M. Organic Acids in Food Preservation: Exploring Synergies, Molecular Insights, and Sustainable Applications. Sustainability 2025, 17, 3434. https://doi.org/10.3390/su17083434
Sorathiya KB, Melo A, Hogg MC, Pintado M. Organic Acids in Food Preservation: Exploring Synergies, Molecular Insights, and Sustainable Applications. Sustainability. 2025; 17(8):3434. https://doi.org/10.3390/su17083434
Chicago/Turabian StyleSorathiya, Kavita Bhavin, Adma Melo, Maria Conceição Hogg, and Manuela Pintado. 2025. "Organic Acids in Food Preservation: Exploring Synergies, Molecular Insights, and Sustainable Applications" Sustainability 17, no. 8: 3434. https://doi.org/10.3390/su17083434
APA StyleSorathiya, K. B., Melo, A., Hogg, M. C., & Pintado, M. (2025). Organic Acids in Food Preservation: Exploring Synergies, Molecular Insights, and Sustainable Applications. Sustainability, 17(8), 3434. https://doi.org/10.3390/su17083434