Abstract
Aim: This study explored the differences in functional traits and soil physical and chemical properties of coastal plant communities under different disturbance intensities. It investigated the correlations between them to gain a deeper understanding of how plant communities adjust their functional traits in response to habitat changes. However, the mechanisms by which human disturbance influences plant functional traits remain unclear. This research endeavors to reveal the adaptive mechanisms and ecological strategies employed by coastal plant communities under different levels of anthropogenic disturbance. Methods: The study examined plant communities in three levels of disturbance (severe, moderate, and mild) in the coastal areas of Pingtan Island. Nine soil physicochemical property indicators and 16 plant functional trait indicators were collected to analyze the correlation between coastal green space plant functional traits and soil physicochemical properties. Results: Soil physicochemical properties (ST, SS, pH) of coastal plants varied under different disturbance intensities. Concurrently, plant functional traits (SLA, LDMC, LTD, LNC, LCC, LPC, LSC, RTD, RPC) also exhibited significant differences. Notably, the interactions among plant functional traits also varied under different disturbance intensities. Furthermore, plant functional traits exhibited distinct response mechanisms to changes in soil physicochemical properties. The plant community adjusts its resource allocation strategy to adapt to environmental changes, which is specifically manifested in the coordination of SRL, SRA, SLA, RPC, RNC, RCC, RSC, LPC, LNC, LT, LTD, and LDMC. Conclusions: Under severe disturbance, plant communities tend to adopt short-term rapid investment-return strategies to cope with harsh environmental conditions; moderate disturbance prompts slow investment-return strategies for long-term stable growth; mild disturbance triggers rapid investment-return strategies to enhance environmental adaptability. The research results indicate that by selecting appropriate plant resources based on different habitat characteristics, it is beneficial for the survival and reproduction of the plant community.
1. Introduction
Coastal green spaces, as transitional zones between land and sea, possess important ecological functions such as windbreak and sand fixation, as well as flood prevention and disaster mitigation. However, due to its unique habitat, coastal soils were subject to erosion from tidal forces and salt spray, typically exhibiting characteristics of infertility, poor nutrient retention capacity, and elevated soil salinity levels []. Currently, research on the relationship between plant functional traits and environmental physicochemical properties has primarily focused on forest, desert, and freshwater ecosystems [,,]. This has resulted in relatively poor restoration outcomes for coastal ecosystems. Therefore, research on the responses of plant communities to soil physical and chemical properties under different disturbance intensities can provide a scientific basis for understanding the habitat changes caused by human activities’ interference with coastal green spaces.
The physical and chemical properties of soil are closely related to the expression of plant functional traits. These physical and chemical properties have a direct impact on the functional traits of both the aboveground and underground parts of plants []. Plants in coastal habitats have exhibited remarkable salt tolerance, drought resistance and wind resistance capabilities. Through the synergy between vegetation and soil, they formed an ecosystem with functions of wind prevention, sand fixation and coastal protection, effectively alleviated natural disasters in the coastal zone [,,]. However, human interference has significantly impacted plant community structure and soil ecosystems, leading to ecosystem degradation and exacerbating soil salinization []. In recent years, Pingtan’s economy has experienced rapid growth driven by the development of the international tourism island. However, human activities such as real estate development have exerted persistent pressure on the island’s ecological environment, leading to increasingly prominent issues including increased soil compaction, accelerated soil salinization, and reduced water-holding capacity [,,]. Pingtan is China’s fifth-largest island and the largest island in Fujian Province []. Balancing its economic development with ecological conservation has become an urgent challenge that must be addressed. Against this backdrop, achieving coordinated development between tourism development and ecological restoration has become a core issue in advancing ecological sustainability.
The growth strategies of plant functional traits are influenced by both the plants themselves and external environmental factors. During the process of plants’ adaptation to environmental heterogeneity, the plasticity of different tissues and organs plays a crucial role. As an organ that is extremely sensitive to environmental changes and has high plasticity, the morphological characteristics of leaves profoundly reflect the trade-off relationship between the environment and plants []. The root system, as the main organ for obtaining water and nutrients, had a close relationship with the carbon and nitrogen cycles in the ecosystem during its respiration, decomposition and turnover processes []. During the growth process of plants, in response to environmental changes, they adapted by altering the morphological structures of leaves and fine roots []. For instance, increasing the thickness of leaves to enhance the utilization efficiency of light resources, and simultaneously increasing the root length to improve the ability to acquire nutrients, in order to responded to environmental changes [,,]. Through the study of plant functional traits, one can gain a deeper understanding of the internal mechanisms by which plants adapt to different environments, providing strategic basis for ecological protection and sustainable development.
Against this backdrop, this study conducted a survey of plant communities in Pingtan’s coastal green spaces, focusing on three different disturbance intensity types. The objective of this study is to address the following questions by examining plant functional traits and their responses to soil conditions across different disturbance levels in coastal areas: (1) How do soil physicochemical properties (including soil temperature (ST), soil moisture content (SMC), soil bulk density (SBD), soil salinity (SS), pH, soil organic carbon content (SNC), soil carbon content (SCC), soil sulfur content (SSC), and soil phosphorus content (SPC)) and plant functional traits (including plant height (H), leaf thickness (LT), specific leaf area (SLA), leaf dry matter content (LDMC), leaf thickness-to-density ratio (LTD), leaf nitrogen content (LNC), leaf carbon content (LCC), leaf phosphorus content (LPC), leaf sulfur content (LSC), root length (SRL), root area (SRA), root diameter (RTD), root nitrogen content (RNC), root carbon content (RCC), root phosphorus content (RPC), root sulfur content (RSC)). (2) What is the relationship between the survival strategies adopted by plants and the physical and chemical properties of the soil under different levels of disturbance? (3) How do human-induced disturbances (severe disturbance, moderate disturbance, and mild disturbance, etc.), soil physical and chemical properties (soil organic matter, soil structure, and soil nutrient content, etc.), as well as plant characteristics (plant species, life form, and functional traits, etc.) among both human and natural factors affect the growth strategies of plants on the eastern coast of Pingtan? In-depth discussions on these issues will help us reveal the adaptive mechanisms and ecological strategies adopted by coastal plant communities under different levels of human disturbance and provide a scientific basis for the protection and rational utilization of coastal plants.
2. Materials and Methods
2.1. Overview of the Study Area
The southeastern coast of Pingtan is located in Pingtan County, Fujian Province, in the southeastern coastal region of China, as shown in Figure 1, with geographical coordinates of 119°75′–119°80′ E and 25°45′–25°50′ N. The region is subject to the influence of monsoons and typhoons throughout the year, with an average annual wind speed of approximately 9 m/s, an average annual precipitation of 1180 mm, and an average annual temperature of 19.6 °C, characteristic of a typical South subtropical marine monsoon climate []. The study area is located along the southeast coast, forming a narrow strip with a total length of 28.1 km. It covers approximately 13.7 km2 of coastal green space. This coastal zone features rich vegetation with high coverage but also exhibits relatively high soil salinity. The main tree species include the Casuarina equisetifolia, Ficus altissima, and Araucaria heterophylla, while the main shrub species include the Murraya exotica, Heptapleurum arboricola, Mirabilis jalapa, and Ligustrum sinense. The herbaceous plants include the Bidens pilosa, Agave sisalana Perr, Oxalis corymbosa, Dianella ensifolia, Ruellia simplex, and Youngia japonica.
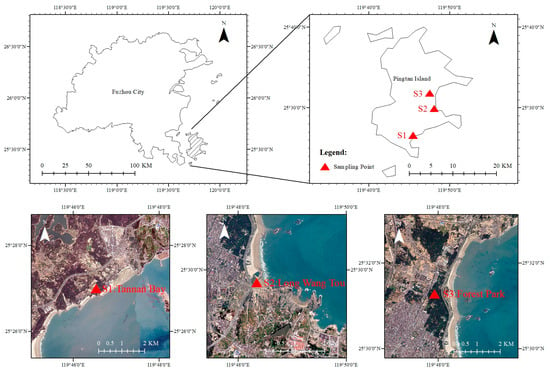
Figure 1.
Map of sampling locations in Pingtan County, Fujian Province, China.
2.2. Classification
Through on-site visits and investigations, this study, referring to the concept of “human disturbance degree” [], regarded fertilization, irrigation frequency, pruning frequency, and tourism as important indicators for measuring the intensity of human interference. The plant communities were classified into the following three types: (1) Severe disturbance type I: near the popular scenic spots of Longwangtou and Tananwan, artificially planted and managed, with high ornamental and economic value. It is fertilized at least 3 times a year and irrigated regularly (once a week); the number of visitors per day is ≥200; (2) Moderate disturbance type II: Pingtan Island Forest Park, including communities formed after artificial planting and natural regeneration and succession, or communities mildly disturbed by some human activities; the disturbance intensity is 1–2 fertilizations per year, mainly relying on natural precipitation; the number of visitors per day is ≥100; (3) Mild disturbance type III: Unexploited areas, natural formations, or communities artificially planted with almost no intervention, no fertilization, or irrigation; the number of visitors per day is <100. The specific classification is shown in Table 1.

Table 1.
Basic Information on Survey Locations.
2.3. Plot Setup
Using the typical plot method [], five survey plots were established for each of the three different disturbance types between September and November 2024, with each plot spaced 50 m apart. In each sample plot, based on the plant community conditions within the plot, large tree sample plots (with an area of 20 m × 20 m), shrub sample plots (with an area of 5 m × 5 m, and 4 shrub sample plots were set at the four corners of the plot where shrubs were present), and herb sample plots (with an area of 1 m × 1 m, and 5 herb sample plots were set at the four corners and the center position line of the shrub sample plot) were set up. A total of 150 sample plots were set up. Within each plot, the following data were recorded: tree species name, number of trees, height, breast height diameter, branch height, crown spread, whether the crown is asymmetrical, and canopy cover; shrub species name, number of shrub clumps, shrub height, direction of leaning, and crown spread; and herb species name, cover, and degree of leaning. In addition, the height of the plants is measured using a height gauge, the trunk diameter is measured with a tree girth ruler, and the coverage is determined by visual estimation. Simultaneously, a portable Global Positioning System (GPS) device was used to record the geographic location information (latitude and longitude) of each plot, along with environmental information (distance from the sea, type of green space), descriptions of human activities, and disturbance conditions.
2.4. Sample Collection
2.4.1. Soil Sample Collection
After determining the sample points, the soil was sampled using the five-point method. At each sample point, four corners and the center position were selected. After removing the impurities on the soil surface, a 100 cm3 ring knife was pressed vertically into the soil. Then, the ring knife was removed using a shovel, and 5 soil samples from the same sample point were mixed and placed in a sealed bag, which was taken back to the laboratory for soil indicator determination. At the same time, soil samples from 0–20 cm depth were also collected at the center position of each sample plot using the ring knife method and brought back to the laboratory to measure soil moisture content (MC) and soil bulk density (BD).
2.4.2. Sampling of Plant Specimens
Select plants that are growing well, and take samples from the four corners and the center of each sample plot. Select 3–5 individuals of each tree species, 5–10 individuals of each shrub species, and 10 or more individuals of herbaceous plants as samples. Twenty to twenty-five leaves with consistent growth conditions, good health, no pests or diseases, and sufficient moisture should be collected. At the same time, samples of the plant’s root system should also be taken. After collecting the leaves, dig out a 20 × 20 × 20 cm soil block 0.5 m away from the base of the standard plant stake. Remove any excess extraneous root systems, clean the root systems of impurities, and pack them into labeled self-sealing bags for separate storage, and then bring them back to the laboratory for cleaning and measurement. After collection, trim the leaf petioles, place the leaves between two sheets of semi-moist absorbent paper or filter paper, and store them in a sealed bag. Label and number the bags according to the quadrat location and species name, then refrigerate them in a foam box with ice packs for freshness preservation. Send them to the laboratory for storage in a refrigerator.
2.5. Indicator Measurement
2.5.1. Soil Indicator Measurement
To clarify the influence of soil physical and chemical properties on plant functional traits, nine indicators were measured. Soil temperature (ST) was measured and recorded on-site using a soil thermometer. Soil moisture content (SMC) was determined using the oven-drying method: soil samples were placed in an oven at 105 °C for 24 h, then cooled and weighed to calculate the soil moisture content (BD). Use a conductivity meter to measure soil electrical conductivity (EC) and convert it to soil salinity (SS); use the potential measurement method to measure pH; use an elemental analyzer to measure total nitrogen (SNC), total carbon (SCC), and total sulfur (SSC); and use the molybdenum blue colorimetric method to measure total phosphorus (SPC).
2.5.2. Plant Trait Measurement
According to the “New Manual for Standardized Measurement of Global Plant Functional Traits [],” 16 functional traits of two plant organs (leaves and roots) were measured, including plant height (H), leaf thickness (LT), specific leaf area (SLA), leaf tissue density (LTD), leaf dry matter content (LDMC), leaf nitrogen content (LNC), leaf carbon content (LCC), leaf sulfur content (LSC), specific root length (SRL), specific root area (SRA), root tissue density (RTD), root nitrogen content (RNC), root carbon content (RCC), and root sulfur content (RSC). Additionally, the molybdenum blue colorimetric method was used to determine leaf phosphorus content (LPC) and root phosphorus content (RPC).
2.6. Data Calculation and Statistical Analysis
Community-weighted mean values were calculated for plant functional trait indices, using species importance values within plots as weights. Community functional traits were represented by the community-weighted mean (CWM) of species traits.
(1) Importance Value Calculation
The formula for calculating the importance value of trees is as follows:
The formula for calculating the importance value of shrubs and herbs is as follows:
(2) Weighted average calculation:
In the formula, s is the importance value greater than the number of plant species within the quadrat; Pi is the importance value of the species in the community; ti is the functional trait value of the species in the community.
In this study, we employed Excel 2010 for preliminary statistical analysis. SPSS 26.0 was utilized to conduct a non-parametric (Kruskal–Wallis H) test to examine the differences in plant community functional traits and soil physical and chemical properties among different community types. The significance level was set at p < 0.05. Please refer to Tables S1 and S2 in the Supplementary Materials. The Pearson correlation coefficient was used to elucidate the correlation between plant functional traits and soil physical and chemical properties, and the data were reported in the form of standard deviation ± mean. The software Canoco 5.0 and Origin 2021 were employed for plotting, and a Spearman correlation analysis was performed on plant functional traits and environmental factors. To further investigate the influence of plant functional traits on soil physical and chemical properties, principal component analysis (PCA) was employed to analyze the key indicators affecting the functional traits of plant communities. Additionally, redundant analysis (RDA) was conducted on the soil physical and chemical properties and plant functional traits of different community types using the Canoco 5.0 multivariate statistical analysis software.
3. Results
3.1. Differential Soil Physicochemical Properties Among Plant Communities Under Different Disturbance Types
As shown in Figure 2, SS and pH were highest in the severely disturbed type and significantly greater than in other disturbance types (p < 0.05). ST was significantly higher in the moderate disturbance type than in the severe and mild disturbance types (p < 0.05). In the moderate disturbance type, the contents of SNC, SCC, SSC, and SPC showed an increasing trend compared to the severe and mild disturbance types. Additionally, mild interference-type ST and SS were significantly lower than other interference types, while SMC was relatively higher.
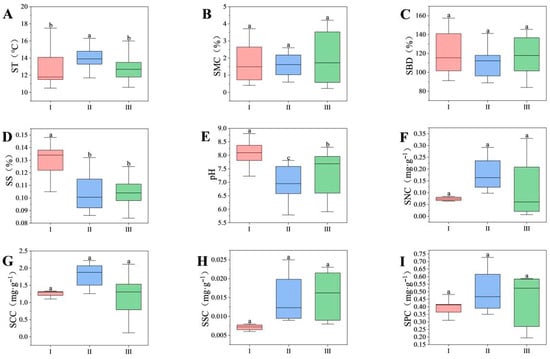
Figure 2.
Soil physical and chemical properties under different disturbance types. Different lowercase letters indicate significant differences among regions (p < 0.05). (I) Severe disturbance type; (II) Moderate disturbance type; (III) Mild disturbance type. The variation characteristics of soil physical and chemical properties under different levels of human disturbance are as follows: ST (A), SMC (B), SBD (C), SS (D), pH (E), SNC (F), SCC (G), SSC (H), and SPC (I).
A Pearson correlation heatmap analysis of the physicochemical properties of Pingtan coastal soils (Figure 3) revealed the following: SPC and SSC showed a significant positive correlation (p < 0.05). SSC exhibited a highly significant positive correlation with both SCC and SNC (p < 0.001). SNC also demonstrated a highly significant positive correlation with SCC (p < 0.05). Soil pH exhibited a highly significant positive correlation with SS (p < 0.01).
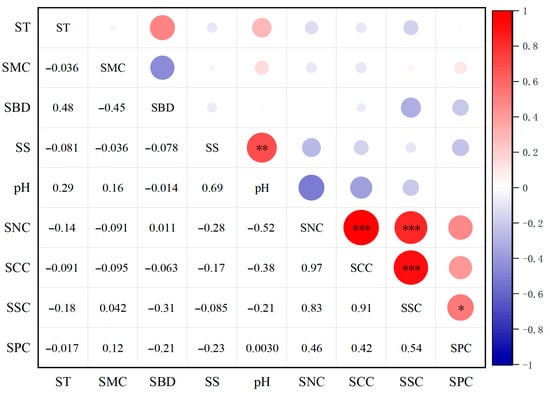
Figure 3.
Heatmap of Correlations in Soil Physicochemical Properties Among Plant Communities Under Different Disturbance Types. Note: *. Indicates significant correlation at the 0.05 level; **. indicates significant correlation at the 0.01 level; ***. indicates significant correlation at the 0.01 level.
3.2. Differences in Functional Traits of Plant Communities Under Different Disturbance Types
As shown in Figure 4, LPC and RPC were significantly higher in heavily disturbed communities than in moderately and mildly disturbed communities (p < 0.05). The SLA was significantly lower in the highly disturbed communities, and it decreased as the degree of human disturbance increased. Moderately disturbed communities exhibited the highest LDMC values, showing significant differences compared to other types (p < 0.05). Moderately disturbed communities exhibited the highest RTD values, showing significant differences compared to other types (p < 0.05). In mildly disturbed communities, SLA, LTD, LNC, LCC, and LSC were significantly higher than in other types (p < 0.05). Additionally, LT, SRL, SRA, and RPC in severely disturbed plant communities increased significantly with increasing human disturbance. RNC, RCC, and RSC in moderately disturbed plant communities showed an increasing trend relative to severely and mildly disturbed plant communities.
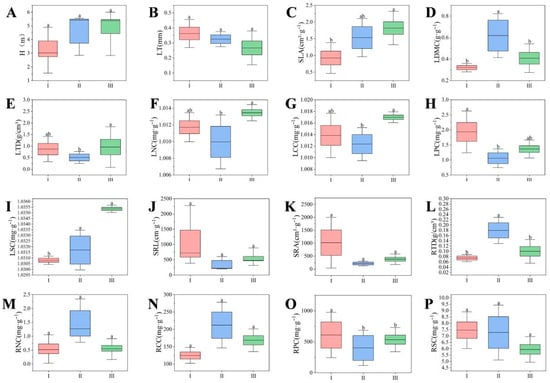
Figure 4.
Functional trait differences in plant communities under various disturbance types. Different lowercase letters indicate significant differences among regions (p < 0.05). (I) Severe disturbance type; (II) Moderate disturbance type; (III) Mild disturbance type. The variation characteristics of functional traits in plant communities under different levels of human disturbance are as follows: Plant Height (H) (A), Leaf Thickness (LT) (B), Specific Leaf Area (SLA) (C), Leaf Dry Matter Content (LDMC) (D), Leaf Thickness to Diameter Ratio (LTD) (E), Leaf Nitrogen Content (LNC) (F), Leaf Carbon Content (LCC) (G), Leaf Phosphorus Content (LPC) (H), Leaf Sulfur Content (LSC) (I), Shoot Root Length (SRL) (J), Shoot Root Area (SRA) (K), Root Tissue Density (RTD) (L), Root Nitrogen Content (RNC) (M), Root Carbon Content (RCC) (N), Root Phosphorus Content (RPC) (O), and Root Sulfur Content (RSC) (P).
A Pearson correlation heatmap analysis of functional traits in plant communities revealed (Figure 5): Leaf traits exhibited strong correlations, with LNC and LSC showing significant positive correlations (p < 0.05), and LNC exhibiting an extremely significant positive correlation with LCC (p < 0.001). LSC showed a significant negative correlation with SLA (p < 0.05). Root traits exhibited high correlations: RNC was highly significantly positively correlated with both RCC and RSC (p < 0.001). RSC and RCC were highly significantly positively correlated (p < 0.001). SRL and SRA also demonstrated a highly significant positive correlation (p < 0.001). Significant relationships also existed between traits of different organs: RPC and LPC showed a significant correlation (p < 0.05). H exhibited significant positive correlations with RSC, RCC, and RNC (p < 0.05), but significant negative correlations with LNC and LCC (p < 0.05). SLA showed significant positive correlations with RCC and RTD (p < 0.05), while exhibiting a significant negative correlation with LSC (p < 0.05).
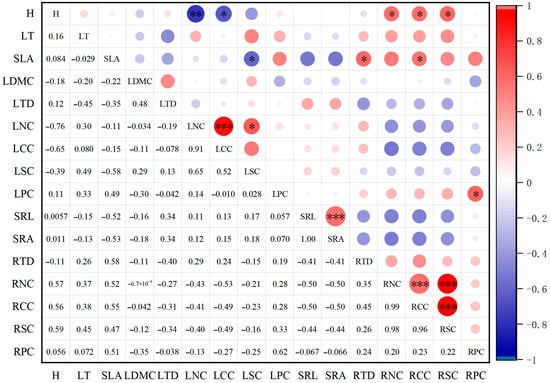
Figure 5.
Correlation Heatmap of Plant Community Functional Shapes Under Different Disturbance Types. Note: *. indicates significant correlation at the 0.05 level, **. indicates significant correlation at the 0.01 level, ***. indicates significant correlation at the 0.01 level.
As shown in Table 2, the principal component analysis indicates that the cumulative contribution rate of the first four PCA axes is 83.411%, and all eigenvalues are greater than 1, indicating that the trait indicators are significantly important in the first four axes. The eigenvalues of the four principal components are 3.981, 3.548, 3.253, and 2.563, respectively, with contribution rates of 24.884%, 22.173%, 20.334%, and 16.020%, respectively. The first principal component (PC1) is highly correlated with indicators such as SRL, SRA, SLA, and RPC, representing root trait indicators. The second principal component (PC2) is closely correlated with indicators such as RNC, RCC, and RSC, representing root system trait indicators. The third principal component (PC3) is closely associated with LPC and LNC, representing leaf functional trait indicators. The fourth principal component, LT, LTD, and LDMC, represents leaf functional traits (PC4).
Table 2.
CoInitial factor rotation component matrix and principal component contribution rate.
3.3. Relationship Between Functional Traits of Plant Communities and Soil Physical and Chemical Properties Under Different Disturbance Types
As shown in Figure 6, the cumulative explanatory power of the first two axes in the severely disturbed type (I) was 73.62%. SBD, SS, SCC, and SPC were the main factors influencing plant functional traits, and as shown in Figure 7I, the explanatory power was SS > SPC > SCC > SBD. The key soil indicator SBD is positively correlated with LDMC and SLA. Among these, SS is significantly positively correlated with SRA, SRL, LTD, RPC, RSC, RNC, and RCC, and negatively correlated with other soil indicators. SPC is significantly positively correlated with SCC and significantly positively correlated with LPC, LT, and LNC, while being significantly negatively correlated with SBD and SS.
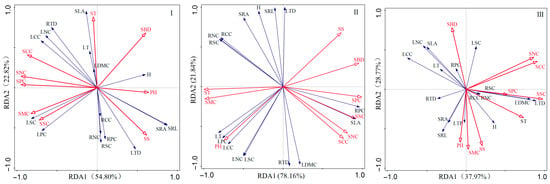
Figure 6.
Redundancy analysis (RDA) ordination diagram of plant community functional traits and soil physicochemical properties under different disturbance types in Pingtan. (I) severe disturbance type redundancy analysis (RDA); (II) moderate disturbance type redundancy analysis (RDA); (III) mild disturbance type redundancy analysis (RDA).

Figure 7.
Analysis of the Variance Explained by Soil Physicochemical Properties. (I) Severe disturbance; (II) Moderate disturbance; (III) Slight disturbance.
In the moderately disturbed type (II), the cumulative explanatory power of the first and second axes was 86.64%, with SCC and SNC being the primary influencing factors for the functional traits of this plant community. As shown in Figure 7II, the explanatory power of SCC was greater than that of SNC. Among these, SCC and SNC exhibit a significant positive correlation with the primary factors influencing soil physicochemical properties and also show significant positive correlations with SLA, RPC, and LDMC, while showing a negative correlation with RNC, RSC, RCC, SRA, LDMC, SRL, and LTD.
In the mildly disturbed type (III), the cumulative explanatory power of the first and second axes was 66.74%, with SMC, SPC, SSC, ST, and SS being the key factors influencing plant community functional traits. As shown in Figure 7III, the explanatory power ranked as ST > SPC > SMC > SS > SSC. The key influencing factors SPC, SSC, ST, SS, and SMC exhibit significant positive correlations with SRL, RPC, and SRA, and negative correlations with all other plant trait indicators. The key indicators RNC, RSC, and RCC show a significant positive correlation with SS and a positive correlation with LDMC and LTD, while exhibiting a negative correlation with all other plant indicators. Among these, ST and SSC show a positive correlation with LPC and a significant negative correlation with SLA, LT, and LNC.
4. Discussion
4.1. Soil Physical and Chemical Properties of Plant Communities with Different Types of Disturbance
Soil is the foundation for plant survival. However, human activities have disrupted the soil’s structure, texture, and other physicochemical properties such as fertility []. This study investigated three habitats with different degrees of disturbance and found that the habitats of plant communities also changed accordingly in response to the varying levels of disturbance. Soil organic matter, total nitrogen, total carbon, total phosphorus, total sulfur, and pH exert significant influence on plant communities under varying degrees of disturbance, holding important practical implications for ecological restoration, land management, and biodiversity conservation.
In the severely disturbed type, SBD was relatively high, which may be related to the trampling of tourists during tourism activities. With the increase in human activities, the degree of compaction in the severely disturbed type increases, the soil becomes more compact, and its bulk density increases accordingly, which was consistent with the research of Zhang Xibiao et al. []. However, as a result of trampling and other human activities, the loss of nutrients has been exacerbated, resulting in relatively low contents of SCC, SNC, SSC, and SPC. This was consistent with the research results of Zhu Fang et al. []. Furthermore, the relatively high SS content might be attributed to the unreasonable irrigation by humans, which has led to the destruction of soil structure, reduced soil permeability, and hindered the downward leaching of salts, thereby intensifying the accumulation of salts in the soil surface. This finding was consistent with the research results of Niu et al. [].
In moderately disturbed soils, the content of SNC, SCC, SPC, and ST was higher than in severely disturbed and mildly disturbed soils, and the soil pH value was significantly lower than in severely disturbed and mildly disturbed soils. According to research by Ma Luwen, the levels of ST and pH value are related to soil microorganisms []. The research results show that the soil with moderate disturbance has a lower moisture content, and low soil moisture content was closely related to soil temperature []. That is, an increase in soil temperature will accelerate the rate of microbial decomposition of organic matter, leading to an increase in soil organic matter and higher soil nutrients. Plants absorb more nutrients to promote the secretion of more organic acids by their root systems, and as a result, the soil pH value was reduced []. Furthermore, the higher contents of SNC, SCC, and SPC in the soil may also be related to human interference (sweeping). Due to the failure to timely sweep the surface litter, nutrients are continuously released through the decomposition activities of microorganisms and contribute to the self-nutrient return to the soil []. In summary, moderate human disturbance can be favorable for improving soil structure to some extent, increasing soil fertility, lowering soil pH, and promoting plant growth.
In mildly disturbed communities, ST and SS were significantly lower than in heavily and moderately disturbed communities, while SMC was higher than in both types. This may be closely related to the fact that this community was subject to minimal artificial management, with litter not being promptly cleared and instead accumulating on the ground surface, extensively covering exposed soil, effectively retaining precipitation, and reducing surface water evaporation [], thereby increasing soil moisture content. Simultaneously, litter cover reduces soil moisture evaporation, inhibiting the migration of salts to the surface through evaporation, thereby lowering soil salinity. This mechanism aligns with previous research findings [,], which indicate that litter cover plays a crucial role in maintaining soil moisture and salinity balance. In summary, in mildly disturbed communities, there was less artificial management, more litter on the ground, and the decomposition of litter lowers the pH value and inhibits the upward movement of soil salts by reducing evaporation, thereby reducing SS.
4.2. Functional Trait Characteristics of Plant Communities Under Different Disturbance Types
This study indicates that there are no significant differences in the functional traits of plant communities, such as H, LT, LSC, SRL, SRA, RNC, RCC, and RSC, under different disturbance intensities (p > 0.05). This indicates that under coastal environmental conditions, drought and saline-alkali stress drive plant communities to coexist by selecting species with similar functional traits, thereby promoting the convergence of plant functional structures []. The key indicators SLA, LDMC, and LTD all showed significant responses under three different disturbance levels, indicating that plant communities adapt to habitat changes by altering their functional traits through growth strategy trade-offs. Specifically, plants adapt to their environment through “short-term rapid investment-return strategies” (rapid growth and reproduction), “slow investment-return strategies” (slow growth and resource storage), and “rapid investment-return strategies” (rapid growth and reproduction).
The trade-off strategies of traits in different types of plant communities vary with the degree of human disturbance []. In the severely disturbed type, LPC and RPC are significantly high, while LDMC and SLA are significantly low, reflecting that the plant community adopts a “short-term rapid investment and return strategy.” Usually, plant communities preserve water or nutrients by reducing SLA. However, for plants with a lower SLA, most substances in the leaves are used to build protective structures and increase the density of mesophyll cells []. Therefore, plants with thicker leaves usually lack the extra energy to increase leaf area, and SLA decreases accordingly. In the severe disturbance type, intermittent and short-term concentrated human disturbances (irrigation, fertilization) lead to the enrichment of phosphorus content, driving the plant community to adopt a “short-term rapid investment and return strategy” to cope with the changes in the habitat, which was different from the traditional ecological strategies []. However, under the influence of human disturbances (irrigation, fertilization, and trampling), the effect of ecological strategies was weakened by their adaptive traits [].
Moderately disturbed LDMC was significantly high, while LNC, LCC, and LPC were significantly low, indicating that plants adapt to the environment by balancing different leaf functional traits. Current research indicates that the low nutrient content in leaves while the high content of dry matter in leaves, was a manifestation of the plants in this type of community adapting to an environment with abundant nutrients. This suggests that plants tend to adopt a “slow investment-return strategy,” extending leaf lifespan to enhance nutrient retention capacity and improve resource utilization efficiency []. Additionally, in moderately disturbed communities, plant communities exhibit higher RTD and RNC, RCC, and RSC levels, suggesting they may be directly influenced by soil nutrients. Moderately disturbed plant communities simultaneously possess characteristics of artificial management (fertilization, irrigation), thereby resulting in higher nutrient content in the root system. It indicates that moderate artificial management can provide more nutrient sources for plants, which is beneficial for their healthy growth.
In mildly disturbed leaves, SLA, LNC, and LCC were significantly higher, indicating that plants enhance light resource acquisition by increasing SLA. Meanwhile, the leaves have a relatively high nutrient content. The plants increase LNC and LCC to improve their ability to store more nutrients, such as N and C, in order to adapt to relatively extreme environments (such as seawater erosion). Furthermore, in this study, both LNC and SLA were significantly higher. A larger SLA facilitates the rate of light energy utilization [], while plants enhance the regulation of key photosynthetic enzyme activity by increasing LNC, thereby influencing photosynthesis []. In conclusion, under mild disturbances, plants can effectively regulate their leaf functions. They increase SLA and LNC to promote photosynthesis and adapt to the influence of the environment.
4.3. Response of Plant Community Functional Traits to Soil Physical and Chemical Properties
This study shows that in different disturbance types, the soil physical and chemical properties that affect plant functional traits vary. In the severe disturbance type, soil bulk density (SBD), soil salt content (SS), soil carbon (SCC), and phosphorus (SPC) content jointly regulate plant functional traits. Under low-nutrient conditions, specific leaf area (SLA) was significantly lower across all three disturbance types. Plants adapt to nutrient deficiency by reducing SLA []. In the severely disturbed type, plant communities adopt a “short-term high-yield investment strategy” to adapt to habitat changes. Soil salinity (SS) was significantly positively correlated with specific root length (SRL), which reflects plant nutrient utilization efficiency. This aligns with the findings of Wang Jiwei et al.: in high-salinity environments, plant communities increase SRL to enhance nutrient absorption from the soil, thereby resisting salt stress []. In summary, through these adaptive adjustments, plants can maintain growth and photosynthetic efficiency in high-salinity environments.
In moderately disturbed communities, plant functional traits are jointly influenced by soil carbon content (SCC) and soil nitrogen content (SNC). This plant community adopts a “slow investment-return strategy.” By increasing the leaf dry matter content (LDMC), it extends the lifespan of the plant leaves, enabling them to carry out photosynthesis for a longer period of time. This strategy contributes to the long-term stable growth and reproduction of the plant community. This is consistent with the findings of Huo Jiaxuan [] in desert plant research. Meanwhile, in a stable and nutrient-rich environment, the root growth of plants under moderate disturbance is promoted, and a higher RTD helps plants to more effectively utilize the abundant nutrient resources, thereby enhancing their growth rate and stress resistance in this environment []. Therefore, in moderate disturbances, under the influence of artificial management, the plant communities have higher RTD and LDMC, which is more conducive to the adaptation of the plant communities to environmental changes.
The functional traits of mildly disturbed plant communities are jointly influenced by soil moisture content (SMC), soil temperature (ST), soil phosphorus content (SPC), and soil sulfur content (SSC). In mildly disturbed communities, SMC was relatively high. However, excessively high soil moisture content restricts root growth [], resulting in significantly lower specific root length (SRL) and specific root area (SRA) in this type of community, thereby affecting the plant roots’ ability to absorb nutrients such as phosphorus and sulfur from the soil. Therefore, the plant community increases SLA to enhance the interception area of light, thereby strengthening photosynthesis and compensating for the insufficient nutrient absorption caused by restricted root systems []. This phenomenon indicates that plant communities adopt a “rapid investment-return strategy” by investing in aboveground parts to enhance photosynthesis, thereby adapting to extreme environments []. Thus, plant communities optimize resource acquisition strategies by increasing SLA, enabling them to survive in limited environments.
5. Conclusions
The results show that under different levels of interference, there are significant differences in plant functional traits and soil physical and chemical properties. Plant communities under severe disturbance adapt to habitat changes by adopting a “short-term rapid investment-return strategy” through reduced SLA and LDMC. Mildly disturbed types adapt to extreme environments by adopting a “rapid investment-return strategy” through increasing leaf nitrogen content (LNC) and specific leaf area (SLA). Compared with the severely disturbed type and the mildly disturbed type, the moderately disturbed communities in the coastal areas maximize resource utilization by enhancing RTD and LDMC and adopting the “slow investment and high return strategy.” Based on this, it is recommended to select and configure plant species with suitable characteristics according to different habitat features. This provides an important scientific basis for the conservation of plant resources in Pingtan’s coastal green spaces and the restoration of island coastal ecosystems, contributing to the sustainable development of ecosystems.
Supplementary Materials
The following supporting information can be downloaded at https://www.mdpi.com/article/10.3390/su172210337/s1: Table S1. Non-parametric Test (Kruskal-Wallis H) of Soil Physicochemical Properties; Table S2. Non-parametric Test (Kruskal-Wallis H) of Plant Functional Traits.
Author Contributions
Conceptualization, Y.Y. and H.Y.; methodology, Y.C. and H.Y.; software, Y.Y. and K.C.; validation, Y.Y. and P.L.; formal analysis, Y.Y.; investigation, P.L. and K.C.; resources, H.Y. and Y.C.; data curation, Y.Y. and P.L.; writing—original draft preparation, Y.Y.; writing—review and editing, H.Y.; visualization, P.L.; supervision, Y.C.; project administration, H.Y.; funding acquisition, H.Y. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by the Huadong Engineering (Fujian) Corporation Ltd. (Grant No. ZKY2021-FJ-02-05).
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
No new data were created or analyzed in this study. Data sharing is not applicable to this article.
Acknowledgments
The authors would like to thank the editors and reviewers for their insights and comments that have substantially improved this paper.
Conflicts of Interest
Authors Yanling Yang and Hongli Yu were employed by the company Huadong Engineering (Fujian) Corporation Ltd. The remaining authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
References
- Wang, X.; Liu, J.K.; Luo, S.X.; Zhao, K.K.; Li, X.J. Divergence of leaf functional traits of coastal sand plants and their relationship with soil physicochemical properties in Hainan Island. J. Trop. Subtrop. Bot. 2022, 30, 708–717. [Google Scholar]
- Hu, Y.S.; Yao, X.Y.; Liu, Y.H. Functional traits of forest plants and their relationships with topographic factors at different successional stages in Changbai Mountain. Acta Ecol. Sin. 2014, 34, 5915–5924. [Google Scholar]
- Song, D.H.; Zhang, X.N.; Yang, J.F.; Tian, J.Y. Functional trait characteristics of different desert plant functional groups and their relationships with soil environment. Acta Ecol. Sin. 2023, 43, 7403–7411. [Google Scholar]
- Luo, L.K.; Zhang, W.; Yuan, M.; Cheng, Y.; Yang, Y.F.; Zhao, K.; Wu, J.H.; Xiao, J.J. Functional traits of dominant plants in the tidal wetlands of the lower Yangtze River and their relationships with soil physicochemical properties. Acta Ecol. Sin. 2025, 45, 13–24. [Google Scholar]
- Chen, X.; Chen, D.Q.; Liu, H.K.; Zhao, C.Z.; Zhao, W.T.; Dong, Z.; Zhang, Y.T.; Wang, Y.P. Responses of fine root anatomical traits of 11 afforestation tree species to soil conditions in the coastal saline-alkali land of the Yellow River Delta. Acta Ecol. Sin. 2021, 41, 4150–4159. [Google Scholar]
- Du, Y.L.; Chen, Y.Z.; Qian, A.G.; Li, J.; Xu, J.; Fu, J.; Cai, L.; Sun, Y.; Liu, B.; Zhang, F.; et al. Salt tolerance of four typical coastal plant species. J. Appl. Oceanogr. 2022, 41, 15–24. [Google Scholar]
- Chen, H.Y.; Tang, K.X.; Sun, Y.M.; Cai, L.C.; Song, H.; Tu, W.-L. Screening of drought-resistant plants and research on drought-resistant techniques in island vegetation restoration. J. Appl. Oceanogr. 2016, 35, 223–228. [Google Scholar]
- Zhou, R.L.; Pang, J.Q.; Song, Y. Physiological responses of the coastal wind-resistant plant Pinus thunbergii to clean wind and aeolian sand flow. Acta Ecol. Sin. 2021, 41, 2033–2044. [Google Scholar]
- You, Y.J.; Wang, Y.X.; Zhang, H.F.; Qiu, W.T.; Wu, M.J. Effects of different human disturbance measures on soil fertility and water retention capacity of natural secondary shrublands. Acta Ecol. Sin. 2018, 38, 1097–1105. [Google Scholar]
- Ye, Z.Y. Floristic Geography of Seed Plants on Pingtan Island, Fujian and the Impact of Alien Plants. Guihaia 2017, 37, 280–293. [Google Scholar]
- Lin, X.P. The State Council’s approval of the construction plan for Pingtan International Tourism Island. Fujian Light Ind. 2016, 8, 32–33. [Google Scholar]
- Huang, L.J.; Zhang, Z.K.; Zheng, J.M.; Huang, S.J.; Lin, L.L.; Wang, Q.; Deng, C.Y.; Liu, X.Z. Functional trait characteristics and evolutionary trends of wild plants on continental islands: A case study of Pingtan Island. J. Trop. Subtrop. Bot. 2017, 25, 429–437. [Google Scholar]
- Yin, F.J.; Chen, H.X.; Huang, F.M.; Chen, Q.M.; Yang, S.Z. Identification and optimization of land-sea connected ecological corridors based on coastal zone ecological function assessment. J. Appl. Oceanogr. 2025, 44, 84–98. [Google Scholar]
- Xiang, S.; Chen, Y.; Zheng, B.S.; Yan, D.L. Research progress on plant leaf morphological characteristics, functional traits and their interactions in environmental adaptation. Anhui For. Sci. Technol. 2024, 50, 11–18+51. [Google Scholar]
- Bakerbrosh, K.F.; Peet, R.K. The ecological significance of lobed and toothed leaves in temperate forest trees. Ecology 1997, 78, 1250–1255. [Google Scholar] [CrossRef]
- Miao, Y.J.; Zong, N.; Li, Z.W. Research progress on the trade-off strategies of functional traits in alpine plants. Chin. J. Appl. Environ. Biol. 2025, 31, 1157–1172. [Google Scholar]
- Liu, W.; Zheng, L.; Qi, D. Variation in leaf traits at different altitudes reflects the adaptive strategy of plants to environmental changes. Ecol. Evol. 2020, 10, 8166–8175. [Google Scholar] [CrossRef]
- Huang, W.J.; Li, Z.J.; Yang, Z.P.; Bai, G.Z. Structural traits and their interrelationships of Populus euphratica heteromorphic leaves. Acta Ecol. Sin. 2010, 30, 4636–4642. [Google Scholar]
- Wu, F.; Wu, C.; Zhang, Y.H.; Yu, H.; Wei, Z.H.; Zheng, W.; Liu, X.F.; Chen, S.D.; Yang, Z.J.; Xiong, D.C. Effects of warming on the growth, morphology, and physiological metabolism of fine roots in a mature Masson pine plantation across different seasons. Chin. J. Plant Ecol. 2023, 47, 856–866. [Google Scholar] [CrossRef]
- Zheng, J.; Arif, M.; Zhang, S.; Yuan, Z.; Zhang, L.; Li, J.; Ding, D.; Li, C. Dam inundation simplifies the plant community composition. Sci. Total Environ. 2021, 801, 149827. [Google Scholar] [CrossRef]
- Peng, T.; Chen, Y.L.; Wang, F.; Xiong, K.H.; Ma, X.D.; Deng, C.Y. Survey of wild liana resources and analysis of their landscape applications in Pingtan. For. Resour. Manag. 2021, 28, 121–129. [Google Scholar]
- Chen, L.D.; Fu, B.J. Types, characteristics, and ecological significance of disturbances. Acta Ecol. Sin. 2000, 20, 581–586. [Google Scholar]
- Pérez-Harguindeguy, N.; Díaz, S.; Garnier, E.; Lavorel, S.; Poorter, H.; Jaureguiberry, P.; Bret-Harte, M.S.; Cornwell, W.K.; Craine, J.M.; Gurvich, D.E.; et al. Corrigendum to: New handbook for standardised measurement of plant functional traits worldwide. Aust. J. Bot. 2016, 64, 715–716. [Google Scholar] [CrossRef]
- Matziris, E.; Stefanou, S.; Papazafeiriou, A.; Seilopoulos, D.; Papaioannou, A. Impacts of human activities on soil physical properties of urban green areas: A case study in Thessaloniki city, Greece. Carpathian J. Earth. Environ. Sci. 2016, 11, 381–394. [Google Scholar]
- Zhang, X.B.; Shen, Z.P. Impact of human disturbance on soil physical properties of Pinus tabuliformis artificial forests in Ziwuling, Loess Plateau. Acta Ecol. Sin. 2006, 26, 3685–3695. [Google Scholar]
- Zhu, F.; Bai, Z.L. Effects of tourism disturbance on vegetation and soil properties in Poyang Lake National Wetland Park. Res. Soil Water Conserv. 2015, 22, 33–39. [Google Scholar]
- Niu, Z.L.; Song, C.L.; Wang, L.; Qi, T.; Chen, M.; Jiang, S.; Zhang, L.; Xu, L.; Liu, J. Spatial variability characteristics of soil salinization in the northern plain irrigation district of Yinchuan, Ningxia. J. Resour. Ecol. 2025, 16, 148–158. (In English) [Google Scholar]
- Ma, L.; Liu, L.; Lu, Y.; Chen, L.; Zhang, Z.; Zhang, H.; Wang, X.; Shu, L.; Yang, Q.; Song, Q.; et al. When microclimates meet soil microbes: Temperature controls soil microbial diversity along an elevational gradient in subtropical forests. Soil Biol. Biochem. 2022, 166, 108566. [Google Scholar] [CrossRef]
- Chen, L.J.; Zhang, X.M.; Wang, X.J.; Cheng, Z.Y.; Shan, Y.Y. Effects of different soil moisture treatments on soil temperature of spring wheat under film irrigation. Trans. Chin. Soc. Agric. Eng. 2008, 24, 9–13. [Google Scholar]
- Kuang, Y.W.; Wen, D.Z.; Zhong, C.W.; Zhou, G.Y. Root exudates and their roles in phytoremediation. Chin. J. Plant Ecol. 2003, 27, 709–717. [Google Scholar]
- He, W.Q.; Chen, L.; Pang, D.B.; Cao, M.H.; Zhang, Y.Q.; Li, X.B. Effects of litter input changes on soil physicochemical properties in forest ecosystems. Acta Ecol. Sin. 2024, 44, 1755–1763. [Google Scholar]
- Zhang, H.; Mu, X.M.; Gao, P. Accumulation and Water-Holding Characteristics of Litter under Different Site Conditions on the Loess Plateau. Res. Soil Water Conserv. 2021, 28, 45–52. [Google Scholar]
- Zhu, Y.; Shi, R.K.; Li, D.H.; Xu, R.Q.; Wu, D.F.; Miao, R.H. Effects of litter input on the chemical stoichiometric characteristics of forest soil. J. Shandong Agric. Univ. (Nat. Sci. Ed.) 2024, 55, 70–75. [Google Scholar]
- He, J.; Ma, L.; Sun, X.; Zhang, J.G. Water retention characteristics of litter and soil and their influencing factors under single rainfall conditions. Chin. J. Appl. Ecol. 2023, 34, 3169–3176. [Google Scholar]
- Meng, T.T.; Ni, J.; Wang, G.H. Plant functional traits and environment and ecosystem functions. Chin. J. Plant Ecol. 2007, 31, 150–165. [Google Scholar]
- Liu, Z.D.; Zhou, X.L.; Tian, J.J.; Yang, L.; Wang, Y.H.; Shen, S.K. Mechanism of Terrestrial Plant Community Assembly under Different Intensities of Anthropogenic Disturbance in Dianchi Lakeside. Forests 2023, 14, 670. [Google Scholar] [CrossRef]
- Huang, W.J.; Li, Z.J.; Yang, Z.P.; Liang, J.Y.; Bai, G.Z. Heteromorphic leaf structural characteristics and their correlations with diameter at breast height of Populus euphratica. Chin. J. Ecol. 2010, 29, 2347–2352. [Google Scholar]
- Grime, J.P. Evidence for the existence of three primary strategies in plants and its relevance to ecological and evolutionary theory. Am. Nat. 1977, 111, 1169–1194. [Google Scholar] [CrossRef]
- Korablev, A.P.; Sandalova, E.V.; Arapov, K.A.; Zaripova, K.M. Biomorphological traits and leaf dry matter content are important to plant persistence in a highly unstable volcanic ground. Nat. Conserv. Res. 2024, 9, 73–89. [Google Scholar] [CrossRef]
- Liu, W.Q.; Li, J.X.; Gong, J.W.; Zhao, L.J.; Xiang, W.H.; Cheng, X.; Wang, W.J.; Zhang, Y.L. Leaf trait variation and adaptive strategies of dominant tree species in evergreen broad-leaved forests of Castanopsis sclerophylla. Acta Ecol. Sin. 2022, 42, 7256–7265. [Google Scholar]
- Li, Q.; Zhao, C.Z.; Zhao, L.C.; Wang, J.W.; Wen, J. Correlation analysis between specific leaf area and photosynthetic efficiency of Phragmites australis in the Qiwangchuan salt marsh wetland. Acta Ecol. Sin. 2019, 39, 7124–7133. [Google Scholar]
- Wang, Z.Q.; Zha, T.S.; Jia, X.; Wu, Y.J.; Zhang, M.Y.; Mu, J.W. Seasonal dynamics of photosynthetic parameters of Artemisia ordosica and their relationships with leaf nitrogen content and specific leaf area. Chin. J. Ecol. 2017, 36, 916–924. [Google Scholar]
- Poorter, H.; Niinemets, Ü.; Poorter, L.; Wright, I.J.; Villar, R. Causes and consequences of variation in leaf mass per area (LMA): A meta-analysis. New Phytol. 2009, 182, 565–588. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.W.; Zhao, C.Z.; Zhao, L.C.; Wang, X.P.; Li, Q. Response of root morphology and biomass allocation of Phragmites australis in inland salt marshes to soil salinity factors. Acta Ecol. Sin. 2018, 38, 4843–4851. [Google Scholar]
- Huo, J.X.; Ren, L.; Pan, Y.P.; Zhao, J.; Xiang, X.; Yu, C.; Meng, D.H.; Wang, Y.Y.; Lu, R.J.; Huang, Y.M. Functional traits of desert plants in the Qaidam Basin and their responses to environmental factors. Acta Ecol. Sin. 2022, 42, 4494–4503. [Google Scholar]
- Zhang, L.; Wang, B. Intraspecific interactions shift from competitive to facilitative across a low to high disturbance gradient in a salt marsh. Plant Ecol. 2016, 217, 959–967. [Google Scholar] [CrossRef]
- Feng, G.L.; Liu, C.M.; Wang, L. Regulatory effects of soil moisture on crop root growth and distribution. Res. Eco-Agric. 1996, 4, 7–11. [Google Scholar]
- Zhou, R.L.; Wen, Z.M.; Liu, Y.Y.; Cui, M.Y.; Yang, Y.T.; Lu, J.X.; Zheng, C.; Lin, Z.Q.; Yuan, L.H. Variations in plant functional traits and their responses to environmental factors across different vegetation zones in the Yan River Basin. Acta Ecol. Sin. 2023, 43, 6045–6057. [Google Scholar]
- Li, Z.C.; Xuan, J.; Wang, Q.X.; Peng, S.T.; Sun, G.M.; Huang, L.Q. Differences in leaf functional traits of plants in different habitats on the Minjiang River island and their responses to soil properties. Acta Ecol. Sin. 2024, 44, 6301–6316. [Google Scholar]
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).