
Molecular Analysis of L-Asparaginases for Clarification of the Mechanism of Action and Optimization of Pharmacological Functions

 , ,
, ,
Abstract
1. Introduction
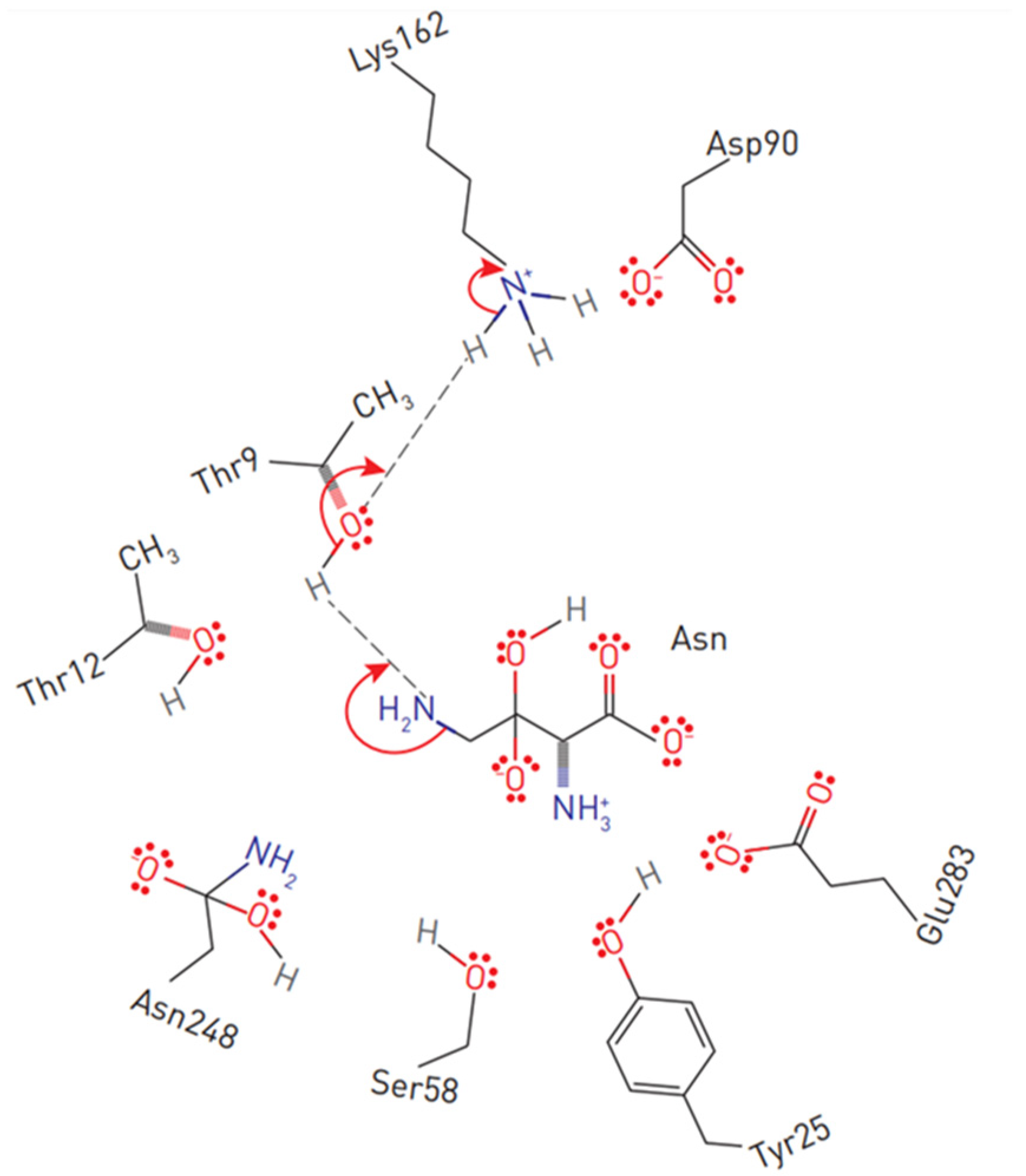
2. Structures of L-ASNases and the Mechanism of Action
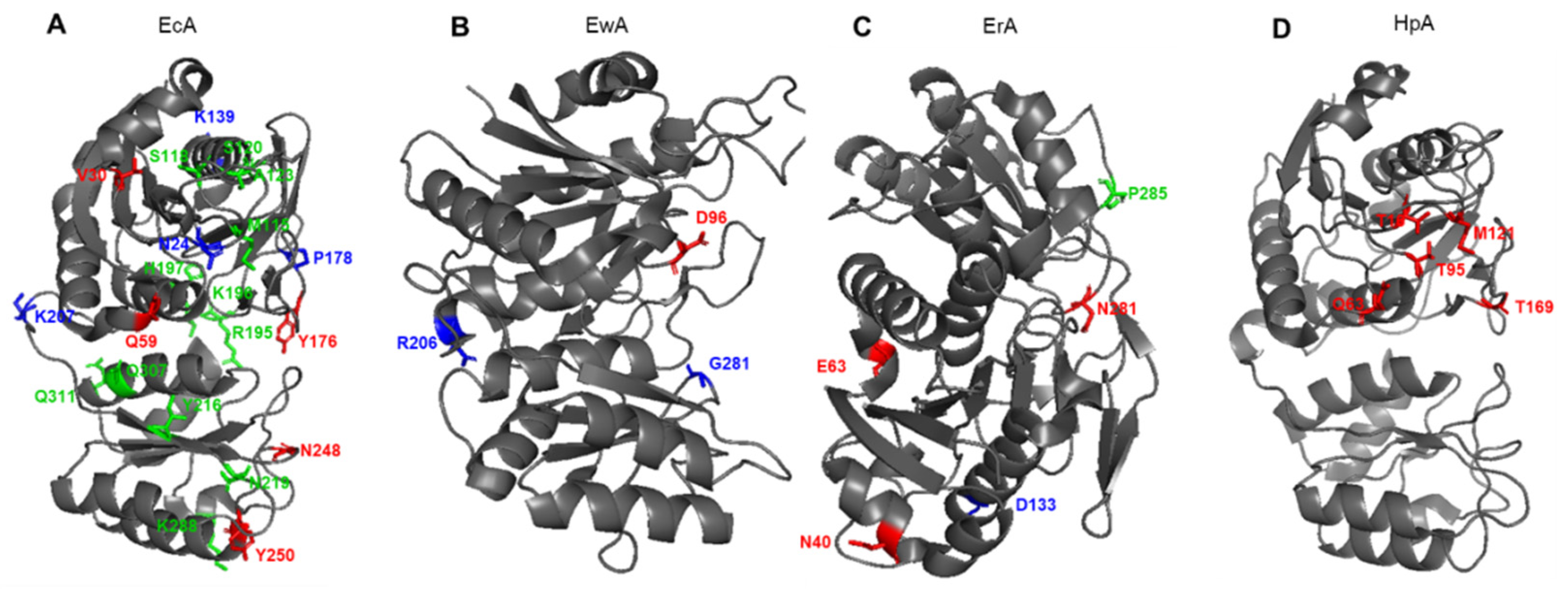
3. Stability and Activity of L-ASNases
4. Protease Resistance and Immunogenicity
5. Substrate Specificity
6. Alternative Approaches to the Development of Antitumor L-ASNases with Improved Functions
7. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Pokrovsky, V.S.; Chepikova, O.E.; Davydov, D.Z.; Zamyatnin, A.A., Jr.; Lukashev, A.N.; Lukasheva, E.V. Amino Acid Degrading Enzymes and their Application in Cancer Therapy. Curr. Med. Chem. 2019, 26, 446–464. [Google Scholar] [CrossRef] [PubMed]
- Beckett, A.; Gervais, D. What makes a good new therapeutic L-asparaginase? World J. Microbiol. Biotechnol. 2019, 35, 152. [Google Scholar] [CrossRef]
- Dinndorf, P.A.; Gootenberg, J.; Cohen, M.H.; Keegan, P.; Pazdur, R. FDA drug approval summary: Pegaspargase (oncaspar) for the first-line treatment of children with acute lymphoblastic leukemia (ALL). Oncologist 2007, 12, 991–998. [Google Scholar] [CrossRef] [PubMed]
- Jaccard, A.; Petit, B.; Girault, S.; Suarez, F.; Gressin, R.; Zini, J.-M.; Coiteux, V.; Larroche, C.; Devidas, A.; Thiéblemont, C.; et al. L-asparaginase-based treatment of 15 western patients with extranodal NK/T-cell lymphoma and leukemia and a review of the literature. Ann. Oncol. Off. J. Eur. Soc. Med. Oncol. 2009, 20, 110–116. [Google Scholar] [CrossRef] [PubMed]
- Völler, S.; Pichlmeier, U.; Zens, A.; Hempel, G. Pharmacokinetics of recombinant asparaginase in children with acute lymphoblastic leukemia. Cancer Chemother. Pharmacol. 2018, 81, 305–314. [Google Scholar] [CrossRef]
- Chien, W.-W.; Allas, S.; Rachinel, N.; Sahakian, P.; Julien, M.; Le Beux, C.; Lacroix, C.-E.; Abribat, T.; Salles, G. Pharmacology, immunogenicity, and efficacy of a novel pegylated recombinant Erwinia chrysanthemi-derived L-asparaginase. Investig. New Drugs 2014, 32, 795–805. [Google Scholar] [CrossRef]
- Sharma, D.; Singh, K.; Singh, K.; Mishra, A. Insights into the Microbial L-Asparaginases: From Production to Practical Applications. Curr. Protein Pept. Sci. 2018, 20, 452–464. [Google Scholar] [CrossRef]
- Lubkowski, J.; Vanegas, J.; Chan, W.-K.; Lorenzi, P.L.; Weinstein, J.N.; Sukharev, S.; Fushman, D.; Rempe, S.; Anishkin, A.; Wlodawer, A. Mechanism of Catalysis by l-Asparaginase. Biochemistry 2020, 59, 1927–1945. [Google Scholar] [CrossRef]
- Ghasemian, A.; Al-marzoqi, A.H.; Al-abodi, H.R.; Alghanimi, Y.K.; Kadhum, S.A.; Shokouhi Mostafavi, S.K.; Fattahi, A. Bacterial l-asparaginases for cancer therapy: Current knowledge and future perspectives. J. Cell. Physiol. 2019, 234, 19271–19279. [Google Scholar] [CrossRef]
- Batool, T.; Makky, E.A.; Jalal, M.; Yusoff, M.M. A Comprehensive Review on l-Asparaginase and Its Applications. Appl. Biochem. Biotechnol. 2016, 178, 900–923. [Google Scholar] [CrossRef]
- Michalska, K.; Jaskolski, M. Structural aspects of L-asparaginases, their friends and relations. Acta Biochim. Pol. 2006, 53, 627–640. [Google Scholar] [CrossRef] [PubMed]
- Niu, J.; Meng, F.; Zhou, Y.; Zhang, C.; Lu, Z.; Lu, F.; Chen, M. Non-classical secretion of a type I L-asparaginase in Bacillus subtilis. Int. J. Biol. Macromol. 2021, 180, 677–683. [Google Scholar] [CrossRef] [PubMed]
- Yao, M.; Yasutake, Y.; Morita, H.; Tanaka, I. Structure of the type I L-asparaginase from the hyperthermophilic archaeon Pyrococcus horikoshii at 2.16 angstroms resolution. Acta Crystallogr. D Biol. Crystallogr. 2005, 61, 294–301. [Google Scholar] [CrossRef] [PubMed]
- Jiao, L.; Chi, H.; Lu, Z.; Zhang, C.; Chia, S.R.; Show, P.L.; Tao, Y.; Lu, F. Characterization of a novel type I l-asparaginase from Acinetobacter soli and its ability to inhibit acrylamide formation in potato chips. J. Biosci. Bioeng. 2020, 129, 672–678. [Google Scholar] [CrossRef] [PubMed]
- Sharafi, Z.; Barati, M.; Khoshayand, M.R.; Adrangi, S. Screening for Type II L-Asparaginases: Lessons from the Genus Halomonas. Iran. J. Pharm. Res. IJPR 2017, 16, 1565–1573. [Google Scholar] [PubMed]
- Nowak-Göttl, U.; Wolff, J.E.A.; Kuhn, N.; Boos, J.; Kehrel, B.; Lilienweiss, V.; Schwabe, D.; Jürgens, H. Enhanced thrombin generation, P-von willebrand factor, P-fibrin D-dimer and P-plasminogen activator inhibitor 1: Predictive for venous thrombosis in asparaginase-treated children. Fibrinolysis 1994, 8, 63–65. [Google Scholar] [CrossRef]
- Leibundgut, K.; Hirt, A.; Zwicky, C.; Wuillemin, W.A. Cerebral sinovenous thrombosis during asparaginase treatment. Case 3. Hamostaseologie 2003, 23, 109–112. [Google Scholar]
- Fonseca, M.H.G.; Fiúza, T.D.S.; de Morais, S.B.; de Souza, T.A.C.B.; Trevizani, R. Circumventing the side effects of L-asparaginase. Biomed. Pharmacother. 2021, 139, 111616. [Google Scholar] [CrossRef]
- Da Silva Lacerda, G.R.; Cantalice, J.C.L.L.; de Souza Lima, G.M.; de Albuquerque, L.E.F.; da Silva, I.D.G.; de Melo, M.E.B.; Adam, M.L.; do Nascimento, S.C. Genotoxic activity of l-asparaginase produced by Streptomyces ansochromogenes UFPEDA 3420. World J. Microbiol. Biotechnol. 2019, 35, 41. [Google Scholar] [CrossRef]
- Aghaiypour, K.; Wlodawer, A.; Lubkowski, J. Structural basis for the activity and substrate specificity of Erwinia chrysanthemi L-asparaginase. Biochemistry 2001, 40, 5655–5664. [Google Scholar] [CrossRef]
- Kessel, D. Asparaginyl-transfer RNA. A substrate for l-asparaginase. BBA Sect. Nucleic Acids Protein Synth. 1971, 240, 554–557. [Google Scholar] [CrossRef]
- Bosmann, H.B.; Kessel, D. Inhibition of glycoprotein synthesis in L5178Y mouse lukaemic cells by L-asparaginase in vitro. Nature 1970, 226, 850–851. [Google Scholar] [CrossRef] [PubMed]
- Ankel, E.G.; Zirneski, J.; Ring, B.J.; Holcenberg, J.S. Effect of asparaginase on cell membranes of sensitive and resistants mouse lymphoma cells. In Vitro 1984, 20, 376–384. [Google Scholar] [CrossRef] [PubMed]
- Zhdanov, D.D.; Pokrovsky, V.S.; Pokrovskaya, M.V.; Alexandrova, S.S.; Eldarov, M.A.; Grishin, D.V.; Basharov, M.M.; Gladilina, Y.A.; Podobed, O.V.; Sokolov, N.N. Rhodospirillum rubrum L-asparaginase targets tumor growth by a dual mechanism involving telomerase inhibition. Biochem. Biophys. Res. Commun. 2017, 492, 282–288. [Google Scholar] [CrossRef] [PubMed]
- Zhdanov, D.D.; Pokrovsky, V.S.; Pokrovskaya, M.V.; Alexandrova, S.S.; Eldarov, M.A.; Grishin, D.V.; Basharov, M.M.; Gladilina, Y.A.; Podobed, O.V.; Sokolov, N.N. Inhibition of telomerase activity and induction of apoptosis by Rhodospirillum rubrum L-asparaginase in cancer Jurkat cell line and normal human CD4+T lymphocytes. Cancer Med. 2017, 6, 2697–2712. [Google Scholar] [CrossRef] [PubMed]
- Plyasova, A.A.; Pokrovskaya, M.V.; Lisitsyna, O.M.; Pokrovsky, V.S.; Alexandrova, S.S.; Hilal, A.; Sokolov, N.N.; Zhdanov, D.D. Penetration into cancer cells via clathrin-dependent mechanism allows l-asparaginase from Rhodospirillum rubrum to inhibit telomerase. Pharmaceuticals 2020, 13, 286. [Google Scholar] [CrossRef] [PubMed]
- Nomme, J.; Su, Y.; Konrad, M.; Lavie, A. Structures of apo and product-bound human L-asparaginase: Insights into the mechanism of autoproteolysis and substrate hydrolysis. Biochemistry 2012, 51, 6816–6826. [Google Scholar] [CrossRef] [PubMed]
- Schalk, A.M.; Lavie, A. Structural and kinetic characterization of guinea pig L-asparaginase type III. Biochemistry 2014, 53, 2318–2328. [Google Scholar] [CrossRef] [PubMed]
- Michalska, K.; Brzezinski, K.; Jaskolski, M. Crystal structure of isoaspartyl aminopeptidase in complex with L-aspartate. J. Biol. Chem. 2005, 280, 28484–28491. [Google Scholar] [CrossRef]
- Sokolov, N.N.; Eldarov, M.A.; Pokrovskaya, M.V.; Aleksandrova, S.S.; Abakumova, O.Y.; Podobed, O.V.; Melik-Nubarov, N.S.; Kudryashova, E.V.; Grishin, D.V.; Archakov, A.I. Bacterial recombinant L-asparaginases: Properties, structure and anti-proliferative activity. Biomed. Khim. 2015, 61, 312–324. [Google Scholar] [CrossRef][Green Version]
- Lubkowski, J.; Wlodawer, A. Structural and biochemical properties of L-asparaginase. FEBS J. 2021, 288, 4183–4209. [Google Scholar] [CrossRef] [PubMed]
- Loch, J.I.; Jaskolski, M. Structural and biophysical aspects of l-asparaginases: A growing family with amazing diversity. IUCrJ 2021, 8, 514–531. [Google Scholar] [CrossRef] [PubMed]
- Yun, M.-K.; Nourse, A.; White, S.W.; Rock, C.O.; Heath, R.J. Crystal structure and allosteric regulation of the cytoplasmic Escherichia coli L-asparaginase I. J. Mol. Biol. 2007, 369, 794–811. [Google Scholar] [CrossRef] [PubMed]
- Gesto, D.S.; Cerqueira, N.M.F.S.A.; Fernandes, P.A.; Ramos, M.J. Unraveling the enigmatic mechanism of L-asparaginase II with QM/QM calculations. J. Am. Chem. Soc. 2013, 135, 7146–7158. [Google Scholar] [CrossRef] [PubMed]
- Kozak, M.; Jurga, S. A comparison between the crystal and solution structures of Escherichia coli asparaginase II. Acta Biochim. Pol. 2002, 49, 509–513. [Google Scholar] [CrossRef]
- Swain, A.L.; Jaskólski, M.; Housset, D.; Rao, J.K.; Wlodawer, A. Crystal structure of Escherichia coli L-asparaginase, an enzyme used in cancer therapy. Proc. Natl. Acad. Sci. USA 1993, 90, 1474–1478. [Google Scholar] [CrossRef]
- Aghaiypour, K.; Wlodawer, A.; Lubkowski, J. Do bacterial L-asparaginases utilize a catalytic triad Thr-Tyr-Glu? Biochim. Biophys. Acta 2001, 1550, 117–128. [Google Scholar] [CrossRef]
- Simossis, V.A.; Heringa, J. PRALINE: A multiple sequence alignment toolbox that integrates homology-extended and secondary structure information. Nucleic Acids Res. 2005, 33, W289–W294. [Google Scholar] [CrossRef]
- Lubkowski, J.; Dauter, M.; Aghaiypour, K.; Wlodawer, A.; Dauter, Z. Atomic resolution structure of Erwinia chrysanthemi L-asparaginase. Acta Crystallogr. D Biol. Crystallogr. 2003, 59, 84–92. [Google Scholar] [CrossRef]
- Papageorgiou, A.C.; Posypanova, G.A.; Andersson, C.S.; Sokolov, N.N.; Krasotkina, J. Structural and functional insights into Erwinia carotovora L-asparaginase. FEBS J. 2008, 275, 4306–4316. [Google Scholar] [CrossRef]
- Lubkowski, J.; Wlodawer, A.; Ammon, H.L.; Copeland, T.D.; Swain, A.L. Structural characterization of Pseudomonas 7A glutaminase-asparaginase. Biochemistry 1994, 33, 10257–10265. [Google Scholar] [CrossRef] [PubMed]
- Lubkowski, J.; Wlodawer, A.; Housset, D.; Weber, I.T.; Ammon, H.L.; Murphy, K.C.; Swain, A.L. Refined crystal structure of Acinetobacter glutaminasificans glutaminase-asparaginase. Acta Crystallogr. D Biol. Crystallogr. 1994, 50, 826–832. [Google Scholar] [CrossRef] [PubMed]
- Röhm, K.H.; Van Etten, R.L. The 18O isotope effect in 13C nuclear magnetic resonance spectroscopy: Mechanistic studies on asparaginase from Escherichia coli. Arch. Biochem. Biophys. 1986, 244, 128–136. [Google Scholar] [CrossRef]
- Palm, G.J.; Lubkowski, J.; Derst, C.; Schleper, S.; Röhm, K.H.; Wlodawer, A. A covalently bound catalytic intermediate in Escherichia coli asparaginase: Crystal structure of a Thr-89-Val mutant. FEBS Lett. 1996, 390, 211–216. [Google Scholar] [CrossRef]
- Schalk, A.M.; Antansijevic, A.; Caffrey, M.; Lavie, A. Experimental Data in Support of a Direct Displacement Mechanism for Type I/II L-Asparaginases. J. Biol. Chem. 2016, 291, 5088–5100. [Google Scholar] [CrossRef]
- Strzelczyk, P.; Zhang, D.; Dyba, M.; Wlodawer, A.; Lubkowski, J. Generalized enzymatic mechanism of catalysis by tetrameric L-asparaginases from mesophilic bacteria. Sci. Rep. 2020, 10, 17516. [Google Scholar] [CrossRef]
- Harms, E.; Wehner, A.; Aung, H.P.; Röhm, K.H. A catalytic role for threonine-12 of E. coli asparaginase II as established by site-directed mutagenesis. FEBS Lett. 1991, 285, 55–58. [Google Scholar] [CrossRef]
- Sanches, M.; Krauchenco, S.; Polikarpov, I. Structure, substrate complexation and reaction mechanism of bacterial asparaginases. Curr. Chem. Biol. 2007, 1, 75–86. [Google Scholar]
- Dhavala, P.; Papageorgiou, A.C. Structure of Helicobacter pylori L-asparaginase at 1.4 A resolution. Acta Crystallogr. D Biol. Crystallogr. 2009, 65, 1253–1261. [Google Scholar] [CrossRef]
- Borek, D.; Kozak, M.; Pei, J.; Jaskolski, M. Crystal structure of active site mutant of antileukemic L-asparaginase reveals conserved zinc-binding site. FEBS J. 2014, 281, 4097–4111. [Google Scholar] [CrossRef]
- Derst, C.; Henseling, J.; Röhm, K.H. Probing the role of threonine and serine residues of E. coli asparaginase II by site-specific mutagenesis. Protein Eng. 1992, 5, 785–789. [Google Scholar] [CrossRef] [PubMed]
- Derst, C.; Wehner, A.; Specht, V.; Röhm, K.H. States and functions of tyrosine residues in Escherichia coli asparaginase II. Eur. J. Biochem. 1994, 224, 533–540. [Google Scholar] [CrossRef] [PubMed]
- Bagert, U.; Röhm, K.H. On the role of histidine and tyrosine residues in E. coli asparaginase. Chemical modification and 1H-nuclear magnetic resonance studies. Biochim. Biophys. Acta 1989, 999, 36–41. [Google Scholar] [CrossRef]
- Aung, H.P.; Bocola, M.; Schleper, S.; Röhm, K.H. Dynamics of a mobile loop at the active site of Escherichia coli asparaginase. Biochim. Biophys. Acta 2000, 1481, 349–359. [Google Scholar] [CrossRef]
- Ortlund, E.; Lacount, M.W.; Lewinski, K.; Lebioda, L. Reactions of Pseudomonas 7A glutaminase-asparaginase with diazo analogues of glutamine and asparagine result in unexpected covalent inhibitions and suggests an unusual catalytic triad Thr-Tyr-Glu. Biochemistry 2000, 39, 1199–1204. [Google Scholar] [CrossRef]
- Dodson, G.; Wlodawer, A. Catalytic triads and their relatives. Trends Biochem. Sci. 1998, 23, 347–352. [Google Scholar] [CrossRef]
- Derst, C.; Henseling, J.; Röhm, K.H. Engineering the substrate specificity of Escherichia coli asparaginase. II. Selective reduction of glutaminase activity by amino acid replacements at position 248. Protein Sci. 2000, 9, 2009–2017. [Google Scholar] [CrossRef]
- Maggi, M.; Chiarelli, L.R.; Valentini, G.; Scotti, C. Tackling Critical Catalytic Residues in Helicobacter pylori L-Asparaginase. Biomolecules 2015, 5, 306–317. [Google Scholar] [CrossRef]
- Jennings, M.P.; Beacham, I.R. Analysis of the Escherichia coli gene encoding L-asparaginase II, ansB, and its regulation by cyclic AMP receptor and FNR proteins. J. Bacteriol. 1990, 172, 1491–1498. [Google Scholar] [CrossRef]
- Karamitros, C.S.; Konrad, M. Fluorescence-Activated Cell Sorting of Human l-asparaginase Mutant Libraries for Detecting Enzyme Variants with Enhanced Activity. ACS Chem. Biol. 2016, 11, 2596–2607. [Google Scholar] [CrossRef]
- Dumina, M.V.; Zhgun, A.A.; Pokrovskay, M.V.; Aleksandrova, S.S.; Zhdanov, D.D.; Sokolov, N.N.; El’darov, M.A. Comparison of Enzymatic Activity of Novel Recombinant L-asparaginases of Extremophiles. Appl. Biochem. Microbiol. 2021, 57, 594–602. [Google Scholar] [CrossRef]
- Pritsa, A.A.; Kyriakidis, D.A. L-asparaginase of Thermus thermophilus: Purification, properties and identification of essential amino acids for its catalytic activity. Mol. Cell. Biochem. 2001, 216, 93–101. [Google Scholar] [CrossRef] [PubMed]
- Dumina, M.; Zhgun, A.; Pokrovskaya, M.; Aleksandrova, S.; Zhdanov, D.; Sokolov, N.; El’darov, M. A Novel L-Asparaginase from Hyperthermophilic Archaeon Thermococcus sibiricus: Heterologous Expression and Characterization for Biotechnology Application. Int. J. Mol. Sci. 2021, 22, 9894. [Google Scholar] [CrossRef] [PubMed]
- Dumina, M.; Zhgun, A.; Pokrovskaya, M.; Aleksandrova, S.; Zhdanov, D.; Sokolov, N.; El’darov, M. Highly Active Thermophilic L-Asparaginase from Melioribacter roseus Represents a Novel Large Group of Type II Bacterial L-Asparaginases from Chlorobi-Ignavibacteriae-Bacteroidetes Clade. Int. J. Mol. Sci. 2021, 22, 13632. [Google Scholar] [CrossRef] [PubMed]
- Fields, P.A. Review: Protein function at thermal extremes: Balancing stability and flexibility. Comp. Biochem. Physiol. A Mol. Integr. Physiol. 2001, 129, 417–431. [Google Scholar] [CrossRef]
- Feller, G. Protein stability and enzyme activity at extreme biological temperatures. J. Phys. Condens. Matter 2010, 22, 323101. [Google Scholar] [CrossRef]
- Miller, S.R. An appraisal of the enzyme stability-activity trade-off. Evolution 2017, 71, 1876–1887. [Google Scholar] [CrossRef]
- Offman, M.N.; Krol, M.; Patel, N.; Krishnan, S.; Liu, J.; Saha, V.; Bates, P.A. Rational engineering of L-asparaginase reveals importance of dual activity for cancer cell toxicity. Blood 2011, 117, 1614–1621. [Google Scholar] [CrossRef]
- Verma, S.; Mehta, R.K.; Maiti, P.; Röhm, K.-H.; Sonawane, A. Improvement of stability and enzymatic activity by site-directed mutagenesis of E. coli asparaginase II. Biochim. Biophys. Acta 2014, 1844, 1219–1230. [Google Scholar] [CrossRef]
- Long, S.; Zhang, X.; Rao, Z.; Chen, K.; Xu, M.; Yang, T.; Yang, S. Amino acid residues adjacent to the catalytic cavity of tetramer L-asparaginase II contribute significantly to its catalytic efficiency and thermostability. Enzym. Microb. Technol. 2016, 82, 15–22. [Google Scholar] [CrossRef]
- Bonthron, D.T.; Jaskólski, M. Why a “benign” mutation kills enzyme activity. Structure-based analysis of the A176V mutant of Saccharomyces cerevisiae L-asparaginase I. Acta Biochim. Pol. 1997, 44, 491–504. [Google Scholar] [CrossRef] [PubMed]
- Vidya, J.; Ushasree, M.V.; Pandey, A. Effect of surface charge alteration on stability of L-asparaginase II from Escherichia sp. Enzym. Microb. Technol. 2014, 56, 15–19. [Google Scholar] [CrossRef] [PubMed]
- Kotzia, G.A.; Labrou, N.E. Engineering thermal stability of L-asparaginase by in vitro directed evolution. FEBS J. 2009, 276, 1750–1761. [Google Scholar] [CrossRef] [PubMed]
- Maggi, M.; Mittelman, S.D.; Parmentier, J.H.; Colombo, G.; Meli, M.; Whitmire, J.M.; Merrell, D.S.; Whitelegge, J.; Scotti, C. A protease-resistant Escherichia coli asparaginase with outstanding stability and enhanced anti-leukaemic activity in vitro. Sci. Rep. 2017, 7, 14479. [Google Scholar] [CrossRef]
- Wehner, A.; Harms, E.; Jennings, M.P.; Beacham, I.R.; Derst, C.; Bast, P.; Röhm, K.H. Site-specific mutagenesis of Escherichia coli asparaginase II. None of the three histidine residues is required for catalysis. Eur. J. Biochem. 1992, 208, 475–480. [Google Scholar] [CrossRef]
- Kotzia, G.A.; Labrou, N.E. Structural and functional role of Gly281 in L-asparaginase from Erwinia carotovora. Protein Pept. Lett. 2013, 20, 1302–1307. [Google Scholar] [CrossRef]
- Li, X.; Zhang, X.; Xu, S.; Xu, M.; Yang, T.; Wang, L.; Zhang, H.; Fang, H.; Osire, T.; Rao, Z. Insight into the thermostability of thermophilic L-asparaginase and non-thermophilic L-asparaginase II through bioinformatics and structural analysis. Appl. Microbiol. Biotechnol. 2019, 103, 7055–7070. [Google Scholar] [CrossRef]
- Mahboobi, M.; Sedighian, H.; Hedayati, C.H.M.; Bambai, B.; Esmaeil Soofian, S.; Amani, J. Applying Bioinformatic Tools for Modeling and Modifying Type II E. coli l-Asparginase to Present a Better Therapeutic Agent/Drug for Acute Lymphoblastic Leukemia. Int. J. Cancer Manag. 2017, 10, e5785. [Google Scholar] [CrossRef]
- Sudhir, A.P.; Agarwaal, V.V.; Dave, B.R.; Patel, D.H.; Subramanian, R.B. Enhanced catalysis of L-asparaginase from Bacillus licheniformis by a rational redesign. Enzym. Microb. Technol. 2016, 86, 1–6. [Google Scholar] [CrossRef]
- Bansal, S.; Srivastava, A.; Mukherjee, G.; Pandey, R.; Verma, A.K.; Mishra, P.; Kundu, B. Hyperthermophilic asparaginase mutants with enhanced substrate affinity and antineoplastic activity: Structural insights on their mechanism of action. FASEB J. Off. Publ. Fed. Am. Soc. Exp. Biol. 2012, 26, 1161–1171. [Google Scholar] [CrossRef]
- Gervais, D.; Foote, N. Recombinant deamidated mutants of Erwinia chrysanthemi L-asparaginase have similar or increased activity compared to wild-type enzyme. Mol. Biotechnol. 2014, 56, 865–877. [Google Scholar] [CrossRef] [PubMed]
- Asselin, B.; Rizzari, C. Asparaginase pharmacokinetics and implications of therapeutic drug monitoring. Leuk. Lymphoma 2015, 56, 2273–2280. [Google Scholar] [CrossRef] [PubMed]
- Radadiya, A.; Zhu, W.; Coricello, A.; Alcaro, S.; Richards, N.G.J. Improving the Treatment of Acute Lymphoblastic Leukemia. Biochemistry 2020, 59, 3193–3200. [Google Scholar] [CrossRef] [PubMed]
- Patel, N.; Krishnan, S.; Offman, M.N.; Krol, M.; Moss, C.X.; Leighton, C.; van Delft, F.W.; Holland, M.; Liu, J.; Alexander, S.; et al. A dyad of lymphoblastic lysosomal cysteine proteases degrades the antileukemic drug L-asparaginase. J. Clin. Investig. 2009, 119, 1964–1973. [Google Scholar] [CrossRef] [PubMed]
- Koprivnikar, J.; McCloskey, J.; Faderl, S. Safety, efficacy, and clinical utility of asparaginase in the treatment of adult patients with acute lymphoblastic leukemia. OncoTargets. Ther. 2017, 10, 1413–1422. [Google Scholar] [CrossRef]
- Asselin, B.L.; Fisher, V. Impact of clinical and subclinical hypersensitivity to asparaginase in acute lymphoblastic leukemia. Clin. J. Oncol. Nurs. 2014, 18, E107–E112. [Google Scholar] [CrossRef]
- Rodrigues, M.A.D.; Pimenta, M.V.; Costa, I.M.; Zenatti, P.P.; Migita, N.A.; Yunes, J.A.; Rangel-Yagui, C.O.; de Sá, M.M.; Pessoa, A.; Costa-Silva, T.A.; et al. Influence of lysosomal protease sensitivity in the immunogenicity of the antitumor biopharmaceutical asparaginase. Biochem. Pharmacol. 2020, 182, 114230. [Google Scholar] [CrossRef]
- Peterson, R.G.; Handschumacher, R.E.; Mitchell, M.S. Immunological responses to L-asparaginase. J. Clin. Investig. 1971, 50, 1080–1090. [Google Scholar] [CrossRef]
- Heo, Y.-A.; Syed, Y.Y.; Keam, S.J. Pegaspargase: A Review in Acute Lymphoblastic Leukaemia. Drugs 2019, 79, 767–777. [Google Scholar] [CrossRef]
- Li, R.-J.; Jin, R.; Liu, C.; Cao, X.; Manning, M.L.; Di, X.M.; Przepiorka, D.; Namuswe, F.; Deisseroth, A.; Goldberg, K.B.; et al. FDA Approval Summary: Calaspargase Pegol-mknl For Treatment of Acute Lymphoblastic Leukemia in Children and Young Adults. Clin. Cancer Res. Off. J. Am. Assoc. Cancer Res. 2020, 26, 328–331. [Google Scholar] [CrossRef]
- Kotzia, G.A.; Lappa, K.; Labrou, N.E. Tailoring structure-function properties of L-asparaginase: Engineering resistance to trypsin cleavage. Biochem. J. 2007, 404, 337–343. [Google Scholar] [CrossRef]
- Newsted, W.J.; Ramjeesingh, M.; Zywulko, M.; Rothstein, S.J.; Shami, E.Y. Engineering resistance to trypsin inactivation into L-asparaginase through the production of a chimeric protein between the enzyme and a protective single-chain antibody. Enzym. Microb. Technol. 1995, 17, 757–764. [Google Scholar] [CrossRef]
- Zinsli, L.V.; Stierlin, N.; Loessner, M.J.; Schmelcher, M. Deimmunization of protein therapeutics—Recent advances in experimental and computational epitope prediction and deletion. Comput. Struct. Biotechnol. J. 2021, 19, 315–329. [Google Scholar] [CrossRef] [PubMed]
- Ramya, L.N.; Pulicherla, K.K. Studies on Deimmunization of Antileukaemic L-Asparaginase to have Reduced Clinical Immunogenicity—An in silico Approach. Pathol. Oncol. Res. 2015, 21, 909–920. [Google Scholar] [CrossRef] [PubMed]
- Cantor, J.R.; Panayiotou, V.; Agnello, G.; Georgiou, G.; Stone, E.M. Engineering reduced-immunogenicity enzymes for amino acid depletion therapy in cancer. Methods Enzymol. 2012, 502, 291–319. [Google Scholar] [CrossRef] [PubMed]
- González-Torres, I.; Perez-Rueda, E.; Evangelista-Martínez, Z.; Zárate-Romero, A.; Moreno-Enríquez, A.; Huerta-Saquero, A. Identification of L-asparaginases from Streptomyces strains with competitive activity and immunogenic profiles: A bioinformatic approach. PeerJ 2020, 8, e10276. [Google Scholar] [CrossRef]
- Jianhua, C.; Yujun, W.; Ruibo, J.; Min, W.; Wutong, W. Probing the antigenicity of E. coli L-asparaginase by mutational analysis. Mol. Biotechnol. 2006, 33, 57–65. [Google Scholar] [CrossRef]
- Mehta, R.K.; Verma, S.; Pati, R.; Sengupta, M.; Khatua, B.; Jena, R.K.; Sethy, S.; Kar, S.K.; Mandal, C.; Roehm, K.H.; et al. Mutations in subunit interface and B-cell epitopes improve antileukemic activities of Escherichia coli asparaginase-II: Evaluation of immunogenicity in mice. J. Biol. Chem. 2014, 289, 3555–3570. [Google Scholar] [CrossRef]
- Moola, Z.B.; Scawen, M.D.; Atkinson, T.; Nicholls, D.J. Erwinia chrysanthemi L-asparaginase: Epitope mapping and production of antigenically modified enzymes. Biochem. J. 1994, 302 Pt 3, 921–927. [Google Scholar] [CrossRef]
- Werner, A.; Röhm, K.-H.; Müller, H.-J. Mapping of B-cell epitopes in E. coli asparaginase II, an enzyme used in leukemia treatment. Biol. Chem. 2005, 386, 535–540. [Google Scholar] [CrossRef]
- Huerta-Saquero, A.; Evangelista-Martínez, Z.; Moreno-Enriquez, A.; Perez-Rueda, E. Rhizobium etli asparaginase II: An alternative for acute lymphoblastic leukemia (ALL) treatment. Bioengineered 2013, 4, 30–36. [Google Scholar] [CrossRef] [PubMed]
- Moreno-Enriquez, A.; Evangelista-Martinez, Z.; Gonzalez-Mondragon, E.G.; Calderon-Flores, A.; Arreguin, R.; Perez-Rueda, E.; Huerta-Saquero, A. Biochemical characterization of recombinant L-asparaginase (AnsA) from Rhizobium etli, a member of an increasing rhizobial-type family of L-asparaginases. J. Microbiol. Biotechnol. 2012, 22, 292–300. [Google Scholar] [CrossRef] [PubMed]
- Kringelum, J.V.; Lundegaard, C.; Lund, O.; Nielsen, M. Reliable B cell epitope predictions: Impacts of method development and improved benchmarking. PLoS Comput. Biol. 2012, 8, e1002829. [Google Scholar] [CrossRef] [PubMed]
- Ponomarenko, J.; Bui, H.-H.; Li, W.; Fusseder, N.; Bourne, P.E.; Sette, A.; Peters, B. ElliPro: A new structure-based tool for the prediction of antibody epitopes. BMC Bioinform. 2008, 9, 514. [Google Scholar] [CrossRef]
- Liang, S.; Zheng, D.; Standley, D.M.; Yao, B.; Zacharias, M.; Zhang, C. EPSVR and EPMeta: Prediction of antigenic epitopes using support vector regression and multiple server results. BMC Bioinform. 2010, 11, 381. [Google Scholar] [CrossRef]
- Pokrovsky, V.S.; Kazanov, M.D.; Dyakov, I.N.; Pokrovskaya, M.V.; Aleksandrova, S.S. Comparative immunogenicity and structural analysis of epitopes of different bacterial L-asparaginases. BMC Cancer 2016, 16, 89. [Google Scholar] [CrossRef]
- Sidoruk, K.V.; Pokrovsky, V.S.; Borisova, A.A.; Omeljanuk, N.M.; Aleksandrova, S.S.; Pokrovskaya, M.V.; Gladilina, J.A.; Bogush, V.G.; Sokolov, N.N. Creation of a producent, optimization of expression, and purification of recombinant Yersinia pseudotuberculosis L-asparaginase. Bull. Exp. Biol. Med. 2011, 152, 219–223. [Google Scholar] [CrossRef]
- Shibayama, K.; Takeuchi, H.; Wachino, J.-I.; Mori, S.; Arakawa, Y. Biochemical and pathophysiological characterization of Helicobacter pylori asparaginase. Microbiol. Immunol. 2011, 55, 408–417. [Google Scholar] [CrossRef]
- Lubkowski, J.; Palm, G.J.; Gilliland, G.L.; Derst, C.; Röhm, K.H.; Wlodawer, A. Crystal structure and amino acid sequence of Wolinella succinogenes L-asparaginase. Eur. J. Biochem. 1996, 241, 201–207. [Google Scholar] [CrossRef]
- Pokrovskaya, M.V.; Pokrovskiy, V.S.; Aleksandrova, S.S.; Anisimova, N.Y.; Andrianov, R.M.; Treschalina, E.M.; Ponomarev, G.V.; Sokolov, N.N. Recombinant intracellular Rhodospirillum rubrum L-asparaginase with low L-glutaminase activity and antiproliferative effect. Biochem. Suppl. Ser. B Biomed. Chem. 2012, 6, 123–131. [Google Scholar] [CrossRef]
- Yeung, V.P.; Chang, J.; Miller, J.; Barnett, C.; Stickler, M.; Harding, F.A. Elimination of an immunodominant CD4+ T cell epitope in human IFN-beta does not result in an in vivo response directed at the subdominant epitope. J. Immunol. 2004, 172, 6658–6665. [Google Scholar] [CrossRef] [PubMed]
- Harding, F.A.; Liu, A.D.; Stickler, M.; Razo, O.J.; Chin, R.; Faravashi, N.; Viola, W.; Graycar, T.; Yeung, V.P.; Aehle, W.; et al. A beta-lactamase with reduced immunogenicity for the targeted delivery of chemotherapeutics using antibody-directed enzyme prodrug therapy. Mol. Cancer Ther. 2005, 4, 1791–1800. [Google Scholar] [CrossRef] [PubMed]
- Fernandez, C.A.; Smith, C.; Yang, W.; Daté, M.; Bashford, D.; Larsen, E.; Bowman, W.P.; Liu, C.; Ramsey, L.B.; Chang, T.; et al. HLA-DRB1*07:01 is associated with a higher risk of asparaginase allergies. Blood 2014, 124, 1266–1276. [Google Scholar] [CrossRef] [PubMed]
- Cantor, J.R.; Yoo, T.H.; Dixit, A.; Iverson, B.L.; Forsthuber, T.G.; Georgiou, G. Therapeutic enzyme deimmunization by combinatorial T-cell epitope removal using neutral drift. Proc. Natl. Acad. Sci. USA 2011, 108, 1272–1277. [Google Scholar] [CrossRef]
- Nomme, J.; Su, Y.; Lavie, A. Elucidation of the specific function of the conserved threonine triad responsible for human L-asparaginase autocleavage and substrate hydrolysis. J. Mol. Biol. 2014, 426, 2471–2485. [Google Scholar] [CrossRef]
- Belviso, S.; Iuliano, R.; Amato, R.; Perrotti, N.; Menniti, M. The human asparaginase enzyme (ASPG) inhibits growth in leukemic cells. PLoS ONE 2017, 12, e0178174. [Google Scholar] [CrossRef]
- Krishnapura, P.R.; Belur, P.D.; Subramanya, S. A critical review on properties and applications of microbial l-asparaginases. Crit. Rev. Microbiol. 2016, 42, 720–737. [Google Scholar] [CrossRef]
- Schalk, A.M.; Nguyen, H.-A.; Rigouin, C.; Lavie, A. Identification and structural analysis of an L-asparaginase enzyme from guinea pig with putative tumor cell killing properties. J. Biol. Chem. 2014, 289, 33175–33186. [Google Scholar] [CrossRef]
- Cantor, J.R.; Stone, E.M.; Chantranupong, L.; Georgiou, G. The human asparaginase-like protein 1 hASRGL1 is an Ntn hydrolase with beta-aspartyl peptidase activity. Biochemistry 2009, 48, 11026–11031. [Google Scholar] [CrossRef]
- Cerofolini, L.; Giuntini, S.; Carlon, A.; Ravera, E.; Calderone, V.; Fragai, M.; Parigi, G.; Luchinat, C. Characterization of PEGylated Asparaginase: New Opportunities from NMR Analysis of Large PEGylated Therapeutics. Chemistry 2019, 25, 1984–1991. [Google Scholar] [CrossRef]
- Brumano, L.P.; da Silva, F.V.S.; Costa-Silva, T.A.; Apolinário, A.C.; Santos, J.H.P.M.; Kleingesinds, E.K.; Monteiro, G.; Rangel-Yagui, C.D.O.; Benyahia, B.; Junior, A.P. Development of L-Asparaginase Biobetters: Current Research Status and Review of the Desirable Quality Profiles. Front. Bioeng. Biotechnol. 2018, 6, 212. [Google Scholar] [CrossRef] [PubMed]
- Meneguetti, G.P.; Santos, J.H.P.M.; Obreque, K.M.T.; Barbosa, C.M.V.; Monteiro, G.; Farsky, S.H.P.; de Oliveira, A.M.; Angeli, C.B.; Palmisano, G.; Ventura, S.P.M.; et al. Correction: Novel site-specific PEGylated L-asparaginase. PLoS ONE 2019, 14, e0224592. [Google Scholar] [CrossRef] [PubMed]
- Roberts, J. Purification and properties of a highly potent antitumor glutaminase-asparaginase from Pseudomonas 7Z. J. Biol. Chem. 1976, 251, 2119–2123. [Google Scholar] [CrossRef]
- Roberts, J.; Holcenberg, J.S.; Dolowy, W.C. Isolation, crystallization, and properties of Achromobacteraceae glutaminase-asparaginase with antitumor activity. J. Biol. Chem. 1972, 247, 84–90. [Google Scholar] [CrossRef]
- Parmentier, J.H.; Maggi, M.; Tarasco, E.; Scotti, C.; Avramis, V.I.; Mittelman, S.D. Glutaminase activity determines cytotoxicity of l-asparaginases on most leukemia cell lines. Leuk. Res. 2015, 39, 757–762. [Google Scholar] [CrossRef]
- Cappelletti, D.; Chiarelli, L.R.; Pasquetto, M.V.; Stivala, S.; Valentini, G.; Scotti, C. Helicobacter pyloril-asparaginase: A promising chemotherapeutic agent. Biochem. Biophys. Res. Commun. 2008, 377, 1222–1226. [Google Scholar] [CrossRef]
- Nguyen, H.A.; Su, Y.; Lavie, A. Structural Insight into Substrate Selectivity of Erwinia chrysanthemi L-asparaginase. Biochemistry 2016, 55, 1246–1253. [Google Scholar] [CrossRef]
- Nguyen, H.A.; Durden, D.L.; Lavie, A. The differential ability of asparagine and glutamine in promoting the closed/active enzyme conformation rationalizes the Wolinella succinogenes L-asparaginase substrate specificity. Sci. Rep. 2017, 7, 41643. [Google Scholar] [CrossRef]
- Timofeev, V.I.; Zhukhlistova, N.E.; Kuranova, I.P. Features of the Three Dimensional Structure of the Mutant Form of Wolinella succinogenes L-Asparaginase in Complexes with L-Aspartic and L-Glutamic Acids. Russ. J. Bioorg. Chem. 2020, 46, 171–180. [Google Scholar] [CrossRef]
- Aghaeepoor, M.; Akbarzadeh, A.; Mirzaie, S.; Hadian, A.; Jamshidi Aval, S.; Dehnavi, E. Selective reduction in glutaminase activity of l-Asparaginase by asparagine 248 to serine mutation: A combined computational and experimental effort in blood cancer treatment. Int. J. Biol. Macromol. 2018, 120, 2448–2457. [Google Scholar] [CrossRef]
- Ardalan, N.; Mirzaie, S.; Sepahi, A.A.; Khavari-Nejad, R.A. Novel mutant of Escherichia coli asparaginase II to reduction of the glutaminase activity in treatment of acute lymphocytic leukemia by molecular dynamics simulations and QM-MM studies. Med. Hypotheses 2018, 112, 7–17. [Google Scholar] [CrossRef] [PubMed]
- Ln, R.; Doble, M.; Rekha, V.P.B.; Pulicherla, K.K. In silico engineering of L-asparaginase to have reduced glutaminase side activity for effective treatment of acute lymphoblastic leukemia. J. Pediatr. Hematol. Oncol. 2011, 33, 617–621. [Google Scholar] [CrossRef] [PubMed]
- Maggi, M.; Chiarelli, L.R.; Valentini, G.; Scotti, C. Engineering of Helicobacter pylori L-asparaginase: Characterization of two functionally distinct groups of mutants. PLoS ONE 2015, 10, e0117025. [Google Scholar] [CrossRef] [PubMed]
- Chan, W.K.; Lorenzi, P.L.; Anishkin, A.; Purwaha, P.; Rogers, D.M.; Sukharev, S.; Rempe, S.B.; Weinstein, J.N. The glutaminase activity of L-asparaginase is not required for anticancer activity against ASNS-negative cells. Blood 2014, 123, 3596–3606. [Google Scholar] [CrossRef]
- Covini, D.; Tardito, S.; Bussolati, O.; Chiarelli, L.R.; Pasquetto, M.V.; Digilio, R.; Valentini, G.; Scotti, C. Expanding targets for a metabolic therapy of cancer: L-asparaginase. Recent Pat. Anticancer. Drug Discov. 2012, 7, 4–13. [Google Scholar] [CrossRef]
- Lomelino, C.L.; Andring, J.T.; McKenna, R.; Kilberg, M.S. Asparagine synthetase: Function, structure, and role in disease. J. Biol. Chem. 2017, 292, 19952–19958. [Google Scholar] [CrossRef]
- Ali, U.; Naveed, M.; Ullah, A.; Ali, K.; Shah, S.A.; Fahad, S.; Mumtaz, A.S. L-asparaginase as a critical component to combat Acute Lymphoblastic Leukaemia (ALL): A novel approach to target ALL. Eur. J. Pharmacol. 2016, 771, 199–210. [Google Scholar] [CrossRef]
- Loureiro, C.B.; Borges, K.S.; Andrade, A.F.; Tone, L.G.; Said, S. Purification and Biochemical Characterization of Native and Pegylated Form of L-Asparaginase from Aspergillus terreus and Evaluation of Its Antiproliferative Activity. Ai Mag. 2012, 2, 138–145. [Google Scholar]
- Meghavarnam, A.K.; Salah, M.; Sreepriya, M.; Janakiraman, S. Growth inhibitory and proapoptotic effects of l-asparaginase from Fusarium culmorum ASP-87 on human leukemia cells (Jurkat). Fundam. Clin. Pharmacol. 2017, 31, 292–300. [Google Scholar] [CrossRef]
- Einsfeldt, K.; Baptista, I.C.; Pereira, J.C.C.V.; Costa-Amaral, I.C.; da Costa, E.S.; Ribeiro, M.C.M.; Land, M.G.P.; Alves, T.L.M.; Larentis, A.L.; Almeida, R.V. Recombinant L-Asparaginase from Zymomonas mobilis: A Potential New Antileukemic Agent Produced in Escherichia coli. PLoS ONE 2016, 11, e0156692. [Google Scholar] [CrossRef]
- Facchinetti de Castro Girão, L.; Gonçalves da Rocha, S.L.; Sobral, R.S.; Dinis Ano Bom, A.P.; Franco Sampaio, A.L.; Godinho da Silva, J.J.; Ferrara, M.A.; Pinto da Silva Bon, E.; Perales, J. Saccharomyces cerevisiae asparaginase II, a potential antileukemic drug: Purification and characterization of the enzyme expressed in Pichia pastoris. Protein Expr. Purif. 2016, 120, 118–125. [Google Scholar] [CrossRef] [PubMed]
- Golbabaie, A.; Nouri, H.; Moghimi, H.; Khaleghian, A. l-asparaginase production and enhancement by Sarocladium strictum: In vitro evaluation of anti-cancerous properties. J. Appl. Microbiol. 2020, 129, 356–366. [Google Scholar] [CrossRef] [PubMed]
- Nguyen, H.A.; Su, Y.; Zhang, J.Y.; Antanasijevic, A.; Caffrey, M.; Schalk, A.M.; Liu, L.; Rondelli, D.; Oh, A.; Mahmud, D.L.; et al. A Novel l-Asparaginase with low l-Glutaminase Coactivity Is Highly Efficacious against Both T- and B-cell Acute Lymphoblastic Leukemias In Vivo. Cancer Res. 2018, 78, 1549–1560. [Google Scholar] [CrossRef] [PubMed]
- Mahajan, R.V.; Kumar, V.; Rajendran, V.; Saran, S.; Ghosh, P.C.; Saxena, R.K. Purification and characterization of a novel and robust L-asparaginase having low-glutaminase activity from Bacillus licheniformis: In vitro evaluation of anti-cancerous properties. PLoS ONE 2014, 9, e99037. [Google Scholar] [CrossRef] [PubMed]
- Saeed, H.; Hemida, A.; El-Nikhely, N.; Abdel-Fattah, M.; Shalaby, M.; Hussein, A.; Eldoksh, A.; Ataya, F.; Aly, N.; Labrou, N.; et al. Highly efficient Pyrococcus furiosus recombinant L-asparaginase with no glutaminase activity: Expression, purification, functional characterization, and cytotoxicity on THP-1, A549 and Caco-2 cell lines. Int. J. Biol. Macromol. 2020, 156, 812–828. [Google Scholar] [CrossRef]
- El-Naggar, N.E.-A.; Deraz, S.F.; El-Ewasy, S.M.; Suddek, G.M. Purification, characterization and immunogenicity assessment of glutaminase free L-asparaginase from Streptomyces brollosae NEAE-115. BMC Pharmacol. Toxicol. 2018, 19, 51. [Google Scholar] [CrossRef]
- Tomar, R.; Sharma, P.; Srivastava, A.; Bansal, S.; Kundu, B. Structural and functional insights into an archaeal L-asparaginase obtained through the linker-less assembly of constituent domains. Acta Crystallogr. D Biol. Crystallogr. 2014, 70, 3187–3197. [Google Scholar] [CrossRef]
- Pokrovskaia, M.V.; Pokrovskiĭ, V.S.; Aleksandrova, S.S.; Anisimova, N.I.; Adrianov, R.M.; Treshchalina, E.M.; Ponomarev, G.V.; Sokolov, N.N. Recombinant intracellular Rhodospirillum rubrum L-asparaginase with low L-glutaminase activity and antiproliferative effect. Biomed. Khim. 2012, 59, 192–208. [Google Scholar] [CrossRef][Green Version]
- Moguel, I.S.; Yamakawa, C.K.; Pessoa, A.J.; Mussatto, S.I. L-asparaginase Production by Leucosporidium scottii in a Bench-Scale Bioreactor with Co-production of Lipids. Front. Bioeng. Biotechnol. 2020, 8, 576511. [Google Scholar] [CrossRef]
- Prakash, P.; Singh, H.R.; Jha, S.K. Production, purification and kinetic characterization of glutaminase free anti-leukemic L-asparaginase with low endotoxin level from novel soil isolate. Prep. Biochem. Biotechnol. 2020, 50, 260–271. [Google Scholar] [CrossRef]
- Maqsood, B.; Basit, A.; Khurshid, M.; Bashir, Q. Characterization of a thermostable, allosteric L-asparaginase from Anoxybacillus flavithermus. Int. J. Biol. Macromol. 2020, 152, 584–592. [Google Scholar] [CrossRef] [PubMed]
- Zuo, S.; Zhang, T.; Jiang, B.; Mu, W. Reduction of acrylamide level through blanching with treatment by an extremely thermostable L-asparaginase during French fries processing. Extremophiles 2015, 19, 841–851. [Google Scholar] [CrossRef] [PubMed]
- El-Nagga, N.E.-A.; El-Ewasy, S.M.; El-Shweihy, N.M. Microbial L-asparaginase as a Potential Therapeutic Agent for the Treatment of Acute Lymphoblastic Leukemia: The Pros and Cons. Int. J. Pharmacol. 2014, 10, 182–199. [Google Scholar] [CrossRef]
- El-Naggar, N.E.-A.; El-Shweihy, N.M. Bioprocess development for L-asparaginase production by Streptomyces rochei, purification and in-vitro efficacy against various human carcinoma cell lines. Sci. Rep. 2020, 10, 7942. [Google Scholar] [CrossRef] [PubMed]
- Ashok, A.; Doriya, K.; Rao, J.V.; Qureshi, A.; Tiwari, A.K.; Kumar, D.S. Microbes Producing L-Asparaginase free of Glutaminase and Urease isolated from Extreme Locations of Antarctic Soil and Moss. Sci. Rep. 2019, 9, 1423. [Google Scholar] [CrossRef] [PubMed]
- Mohamed, S.A.; Elshal, M.F.; Kumosani, T.A.; Aldahlawi, A.M.; Basbrain, T.A.; Alshehri, F.A.; Choudhry, H. L-Asparaginase Isolated from Phaseolus vulgaris Seeds Exhibited Potent Anti-Acute Lymphoblastic Leukemia Effects In-Vitro and Low Immunogenic Properties In-Vivo. Int. J. Environ. Res. Public Health 2016, 13, 1008. [Google Scholar] [CrossRef]
- Saeed, H.; Soudan, H.; El-Sharkawy, A.; Farag, A.; Embaby, A.; Ataya, F. Expression and Functional Characterization of Pseudomonas aeruginosa Recombinant L. Asparaginase. Protein J. 2018, 37, 461–471. [Google Scholar] [CrossRef]
- Souza, P.M.; de Freitas, M.M.; Cardoso, S.L.; Pessoa, A.; Guerra, E.N.S.; Magalhães, P.O. Optimization and purification of l-asparaginase from fungi: A systematic review. Crit. Rev. Oncol. Hematol. 2017, 120, 194–202. [Google Scholar] [CrossRef]
- Wade, H.E. Asparaginase: Molecular and enzymic requirements for use as an antileukaemia agent. J. Gen. Microbiol. 1971, 65, x–xi. [Google Scholar]
- Prihanto, A.A.; Wakayama, M. Marine Microorganism: An Underexplored Source of l-Asparaginase. Adv. Food Nutr. Res. 2016, 79, 1–25. [Google Scholar] [CrossRef]
- Lee, S.-J.; Lee, Y.; Park, G.-H.; Umasuthan, N.; Heo, S.-J.; De Zoysa, M.; Jung, W.-K.; Lee, D.-W.; Kim, H.; Kang, D.-H.; et al. A Newly Identified Glutaminase-Free ʟ-Asparaginase (ʟ-ASPG86) from the Marine Bacterium Mesoflavibacter zeaxanthinifaciens. J. Microbiol. Biotechnol. 2016, 26, 1115–1123. [Google Scholar] [CrossRef] [PubMed]
- Alrumman, S.A.; Mostafa, Y.S.; Al-Izran, K.A.; Alfaifi, M.Y.; Taha, T.H.; Elbehairi, S.E. Production and Anticancer Activity of an L-Asparaginase from Bacillus licheniformis Isolated from the Red Sea, Saudi Arabia. Sci. Rep. 2019, 9, 3756. [Google Scholar] [CrossRef] [PubMed]
- Pokrovskaya, M.V.; Zhdanov, D.D.; Eldarov, M.A.; Aleksandrova, S.S.; Veselovsky, A.V.; Pokrovskiy, V.S.; Grishin, D.V.; Gladilina, J.A.; Sokolov, N.N. Suppression of telomerase activity in leukemic cells by mutant forms of Rhodospirillum rubrum L-asparaginase. Biochem. Suppl. Ser. B Biomed. Chem. 2017, 11, 219–233. [Google Scholar] [CrossRef]
- Borek, D.; Jaskólski, M. Sequence analysis of enzymes with asparaginase activity. Acta Biochim. Pol. 2001, 48, 893–902. [Google Scholar] [CrossRef]
- Graham, M.L. Pegaspargase: A review of clinical studies. Adv. Drug Deliv. Rev. 2003, 55, 1293–1302. [Google Scholar] [CrossRef]
- Avramis, V.I.; Tiwari, P.N. Asparaginase (native ASNase or pegylated ASNase) in the treatment of acute lymphoblastic leukemia. Int. J. Nanomedicine 2006, 1, 241–254. [Google Scholar]
- Rytting, M. Peg-asparaginase for acute lymphoblastic leukemia. Expert Opin. Biol. Ther. 2010, 10, 833–839. [Google Scholar] [CrossRef]
- Sukhoverkov, K.V.; Kudryashova, E. V PEG-chitosan and glycol-chitosan for improvement of biopharmaceutical properties of recombinant L-asparaginase from Erwinia carotovora. Biochemistry 2015, 80, 113–119. [Google Scholar] [CrossRef]
- Malakhova, M.A.; Pokrovskaya, M.V.; Alexandrova, S.S.; Sokolov, N.N.; Kudryashova, E. V Regulation of Catalytic Activity of Recombinant L-Asparaginase from Rhodospirillum rubrum by Conjugation with a PEG-Chitosan Copolymer. Mosc. Univ. Chem. Bull. 2018, 73, 185–191. [Google Scholar] [CrossRef]
- Sannikova, E.P.; Bulushova, N.V.; Cheperegin, S.E.; Gubaydullin, I.I.; Chestukhina, G.G.; Ryabichenko, V.V.; Zalunin, I.A.; Kotlova, E.K.; Konstantinova, G.E.; Kubasova, T.S.; et al. The Modified Heparin-Binding L-Asparaginase of Wolinella succinogenes. Mol. Biotechnol. 2016, 58, 528–539. [Google Scholar] [CrossRef]
- Domenech, C.; Thomas, X.; Chabaud, S.; Baruchel, A.; Gueyffier, F.; Mazingue, F.; Auvrignon, A.; Corm, S.; Dombret, H.; Chevallier, P.; et al. l-asparaginase loaded red blood cells in refractory or relapsing acute lymphoblastic leukaemia in children and adults: Results of the GRASPALL 2005-01 randomized trial. Br. J. Haematol. 2011, 153, 58–65. [Google Scholar] [CrossRef] [PubMed]
- Jorge, J.C.; Perez-Soler, R.; Morais, J.G.; Cruz, M.E. Liposomal palmitoyl-L-asparaginase: Characterization and biological activity. Cancer Chemother. Pharmacol. 1994, 34, 230–234. [Google Scholar] [CrossRef] [PubMed]
- Li, H.; Yang, Y.; Hong, W.; Huang, M.; Wu, M.; Zhao, X. Applications of genome editing technology in the targeted therapy of human diseases: Mechanisms, advances and prospects. Signal Transduct. Target. Ther. 2020, 5, 1. [Google Scholar] [CrossRef] [PubMed]
- Anishchenko, I.; Pellock, S.J.; Chidyausiku, T.M.; Ramelot, T.A.; Ovchinnikov, S.; Hao, J.; Bafna, K.; Norn, C.; Kang, A.; Bera, A.K.; et al. De novo protein design by deep network hallucination. Nature 2021, 600, 547–552. [Google Scholar] [CrossRef]
- Evans, R.; O’tNeill, M.; Pritzel, A.; Antropova, N.; Senior, A.; Green, T.; Židek, A.; Bates, R.; Blackwell, S.; Yim, J.; et al. Protein complex prediction with AlphaFold-Multimer. bioRxiv 2021, 3034. [Google Scholar] [CrossRef]
- Li, T.; Wang, N.; Li, S.; Zhao, Q.; Guo, M.; Zhang, C. Optimization of covalent immobilization of pectinase on sodium alginate support. Biotechnol. Lett. 2007, 29, 1413–1416. [Google Scholar] [CrossRef]
- Nguyen, H.A.; Su, Y.; Lavie, A. Design and Characterization of Erwinia Chrysanthemi l-Asparaginase Variants with Diminished l-Glutaminase Activity. J. Biol. Chem. 2016, 291, 17664–17676. [Google Scholar] [CrossRef]
- Pokrovskaya, M.V.; Aleksandrova, S.S.; Pokrovsky, V.S.; Veselovsky, A.V.; Grishin, D.V.; Abakumova, O.Y.; Podobed, O.V.; Mishin, A.A.; Zhdanov, D.D.; Sokolov, N.N. Identification of functional regions in the Rhodospirillum rubrum L-asparaginase by site-directed mutagenesis. Mol. Biotechnol. 2015, 57, 251–264. [Google Scholar] [CrossRef]
- Costa, I.M.; Schultz, L.; de Araujo Bianchi Pedra, B.; Leite, M.S.M.; Farsky, S.H.P.; de Oliveira, M.A.; Pessoa, A.; Monteiro, G. Recombinant L-asparaginase 1 from Saccharomyces cerevisiae: An allosteric enzyme with antineoplastic activity. Sci. Rep. 2016, 6, 36239. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Source | Mutations | Results Achieved |
|---|---|---|
| Bacillus licheniformis | D103V | Higher thermal stability and increased L-asparaginase activity [79] |
| Bacillus subtilis | G107D | Increased stability and asparaginase activity [70] |
| Escherichia coli II | L23G, K129L, S263C, R291F | Nontoxic, increased stability and longer half-life [78] |
| Escherichia coli II | N24T, N24A | Increased asparaginase activity and protease resistance [68] |
| Escherichia coli II | N24G | Asparagine endopeptidase-resistant; retains 45% of L-asparaginase activity [84] |
| Escherichia coli II | N24A, R195S | 50% glutaminase activity and ~100% L-asparaginase activity [68] |
| Escherichia coli II | N24S | Improved thermal stability and protease resistance [74] |
| Escherichia coli II | V27T | Glutaminase activity reduction and more stable [131] |
| Escherichia coli II | N24A/Y250L | ~0% glutaminase activity and ~72% L-asparaginase activity [68] |
| Escherichia coli II | Q59L | 0% glutaminase activity and ~80% L-asparaginase activity [134] |
| Escherichia coli II | W66Y | Significantly more apoptosis in lymphocytes from acute lymphoblastic leukemia patients [69,98] |
| Escherichia coli II | Y176F | Glutaminase activity reduction and ~100% L-asparaginase activity [69,98] Increase in Vmax and Km for L-asparagine and beta-hydroxamate [69] |
| Escherichia coli II | Y176S | Increase in Vmax and Km for L-asparagine and beta-hydroxamate Glutaminase activity reduction and ~100% L-asparaginase activity [69,98] |
| Escherichia coli II | P178N | Retention of 90% L-asparaginase activity at 50 °C [176] |
| Escherichia coli II | R195A, K196A, H197A | Less immunogenic [97] |
| Escherichia coli II | N248A | Glutaminase activity reduction [75] |
| Escherichia coli II | N248S | Glutaminase activity reduction [130] |
| Escherichia coli II | K288S, Y176F | 10-fold less immunogenic Reduction of glutaminase activity Intact asparaginase activity [69,98] |
| Escherichia coli II | K139D/K207D, K139A, K207A | Increased stability [72] |
| Escherichia coli II | M115V, S118P, S120R, A123P, I215V, N219G, Q307T, Q312N | Substantially reduced immunogenicity [114] |
| Erwinia carotovora (Pectobacterium carotovorum) | D96A | Decreased glutaminase activity and increased asparaginase activity [132] |
| Erwinia carotovora (Pectobacterium carotovorum) | R206H | Resistance to trypsin degradation and higher thermal stability [91] |
| Erwinia carotovora (Pectobacterium carotovorum) | G281S | Increased half-inactivation temperature; decreased catalytic activity for L-asparagine [76] |
| Dickeya dadantii (Erwinia chrysanthemi) | E63Q | Decreased L-glutaminase activity [177] |
| Dickeya dadantii (Erwinia chrysanthemi) | D133V, D133L | Higher thermal stability [73] |
| Dickeya dadantii (Erwinia chrysanthemi) | P285T | Eight-fold reduced immunogenicity [99] |
| Dickeya dadantii (Erwinia chrysanthemi) | N41D, N281D | Increased asparaginase activity and decreased L-glutaminase activity [81] |
| Helicobacter pylori | T16D | Reduction of L-asparaginase and L-glutaminase activities [74] |
| Helicobacter pylori | Q63E | Decreased glutaminase activity [74] |
| Helicobacter pylori | T95E | Reduction of L-asparaginase and L-glutaminase activities [74] |
| Helicobacter pylori | M121C/T169M | Undetectable L-glutaminase activity [133] |
| Pyrococcus furiosus | K274E | Increased L-asparaginase activity, resistance to proteolytic digestion and no displayed glutaminase activity [80] |
| Rhodospirillum rubrum | D60K, F61L | Improvement of kinetic parameters and enzyme stabilization [178] |
| Rhodospirillum rubrum | E149R, V150P, F151T | Able to reduce the expression hTERT subunit of telomerase and suppress telomerase activity [24,25,163] |
| Saccharomyces cerevisiae | T64A, Y78A, T141A, K215A | 99.9% loss of activity [179] |
| Wolinella succinogenes | V23Q, K24T | Resistance to trypsin degradation, decreased glutaminase activity and reduced immunogenicity [170] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Pokrovskaya, M.V.; Pokrovsky, V.S.; Aleksandrova, S.S.; Sokolov, N.N.; Zhdanov, D.D. Molecular Analysis of L-Asparaginases for Clarification of the Mechanism of Action and Optimization of Pharmacological Functions. Pharmaceutics 2022, 14, 599. https://doi.org/10.3390/pharmaceutics14030599
Pokrovskaya MV, Pokrovsky VS, Aleksandrova SS, Sokolov NN, Zhdanov DD. Molecular Analysis of L-Asparaginases for Clarification of the Mechanism of Action and Optimization of Pharmacological Functions. Pharmaceutics. 2022; 14(3):599. https://doi.org/10.3390/pharmaceutics14030599
Chicago/Turabian StylePokrovskaya, Marina V., Vadim S. Pokrovsky, Svetlana S. Aleksandrova, Nikolay N. Sokolov, and Dmitry D. Zhdanov. 2022. "Molecular Analysis of L-Asparaginases for Clarification of the Mechanism of Action and Optimization of Pharmacological Functions" Pharmaceutics 14, no. 3: 599. https://doi.org/10.3390/pharmaceutics14030599
APA StylePokrovskaya, M. V., Pokrovsky, V. S., Aleksandrova, S. S., Sokolov, N. N., & Zhdanov, D. D. (2022). Molecular Analysis of L-Asparaginases for Clarification of the Mechanism of Action and Optimization of Pharmacological Functions. Pharmaceutics, 14(3), 599. https://doi.org/10.3390/pharmaceutics14030599