
Supramolecular Complexes of Plant Neurotoxin Veratridine with Cyclodextrins and Their Antidote-like Effect on Neuro-2a Cell Viability

 , ,
, ,  and
and 
Abstract
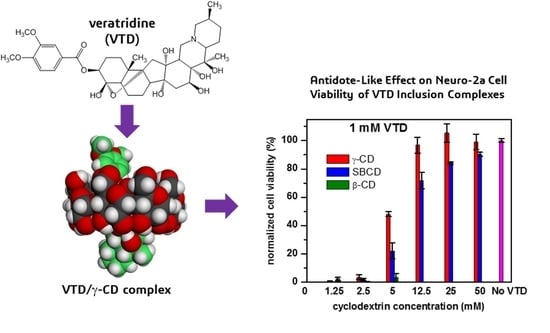
:
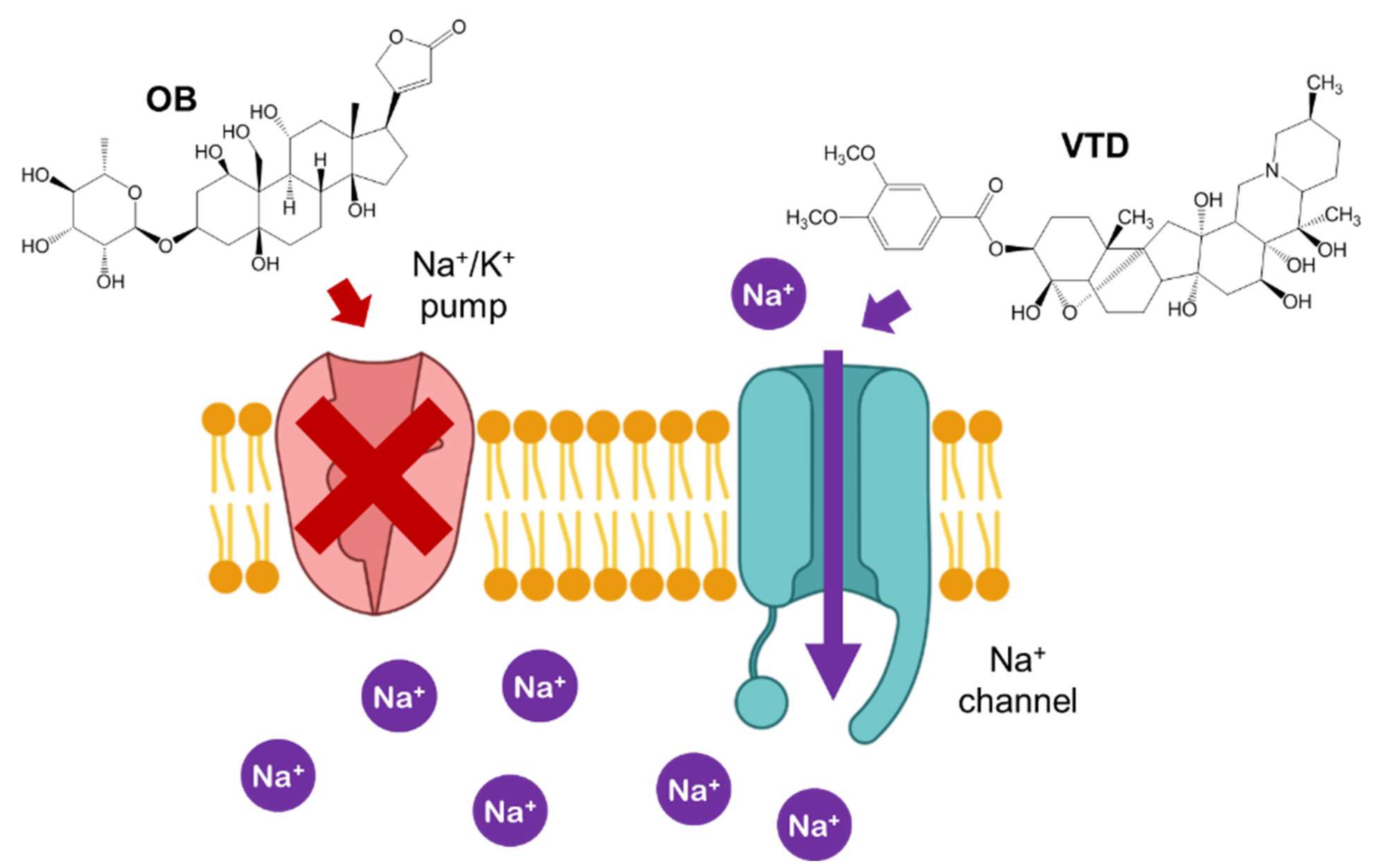
1. Introduction
2. Results and Discussion
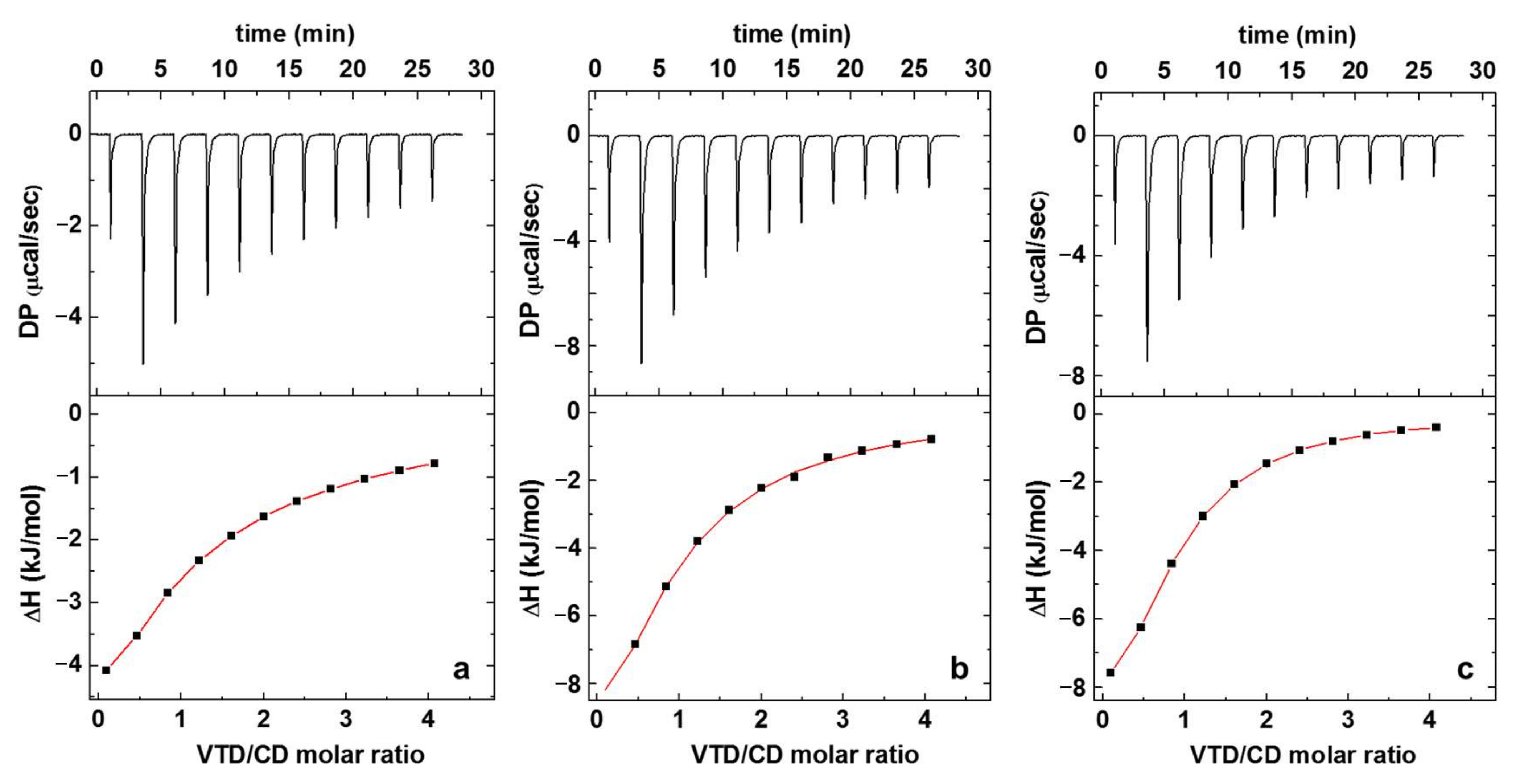
2.1. Isothermal Titration Calorimetry (ITC)
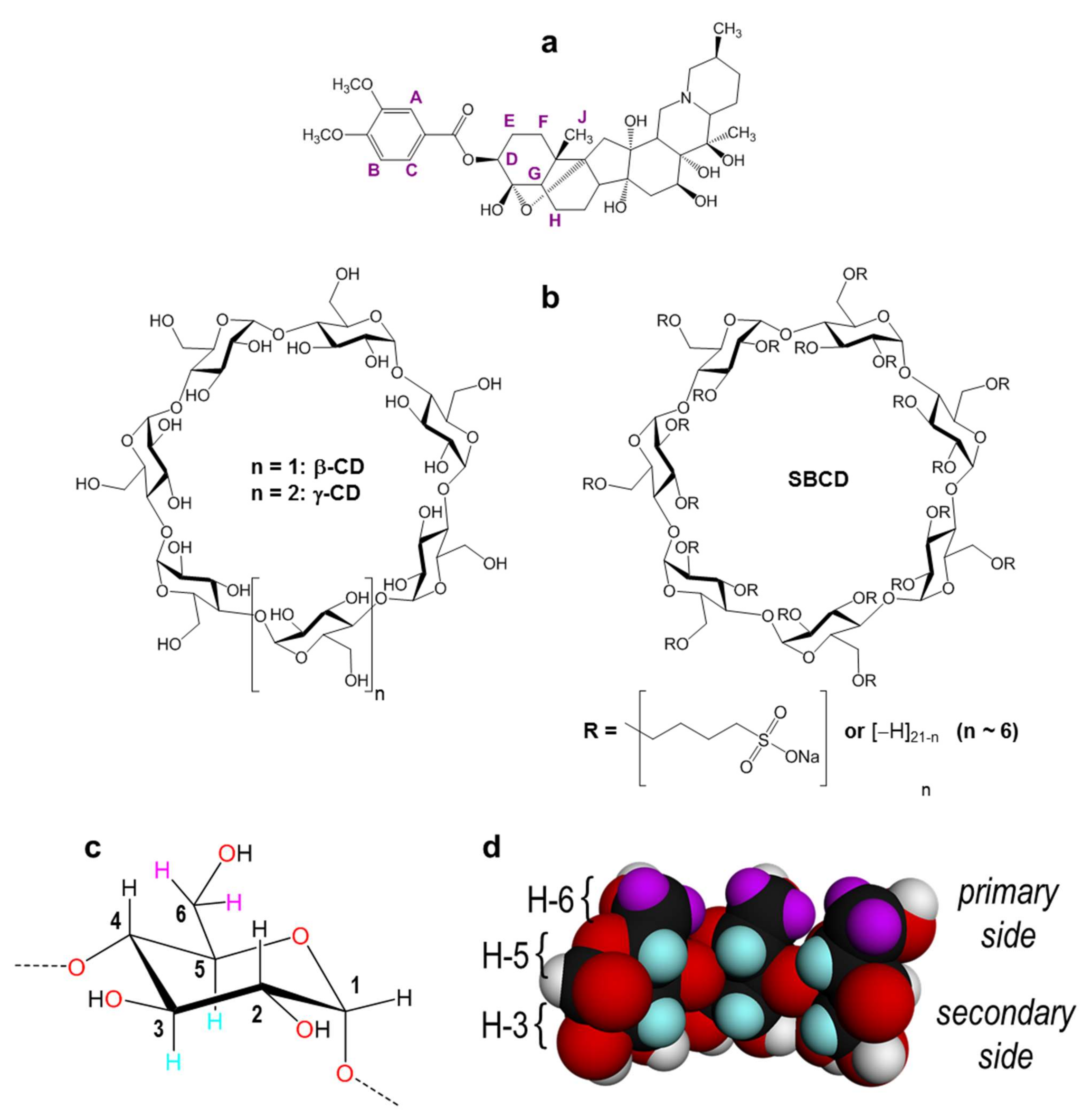
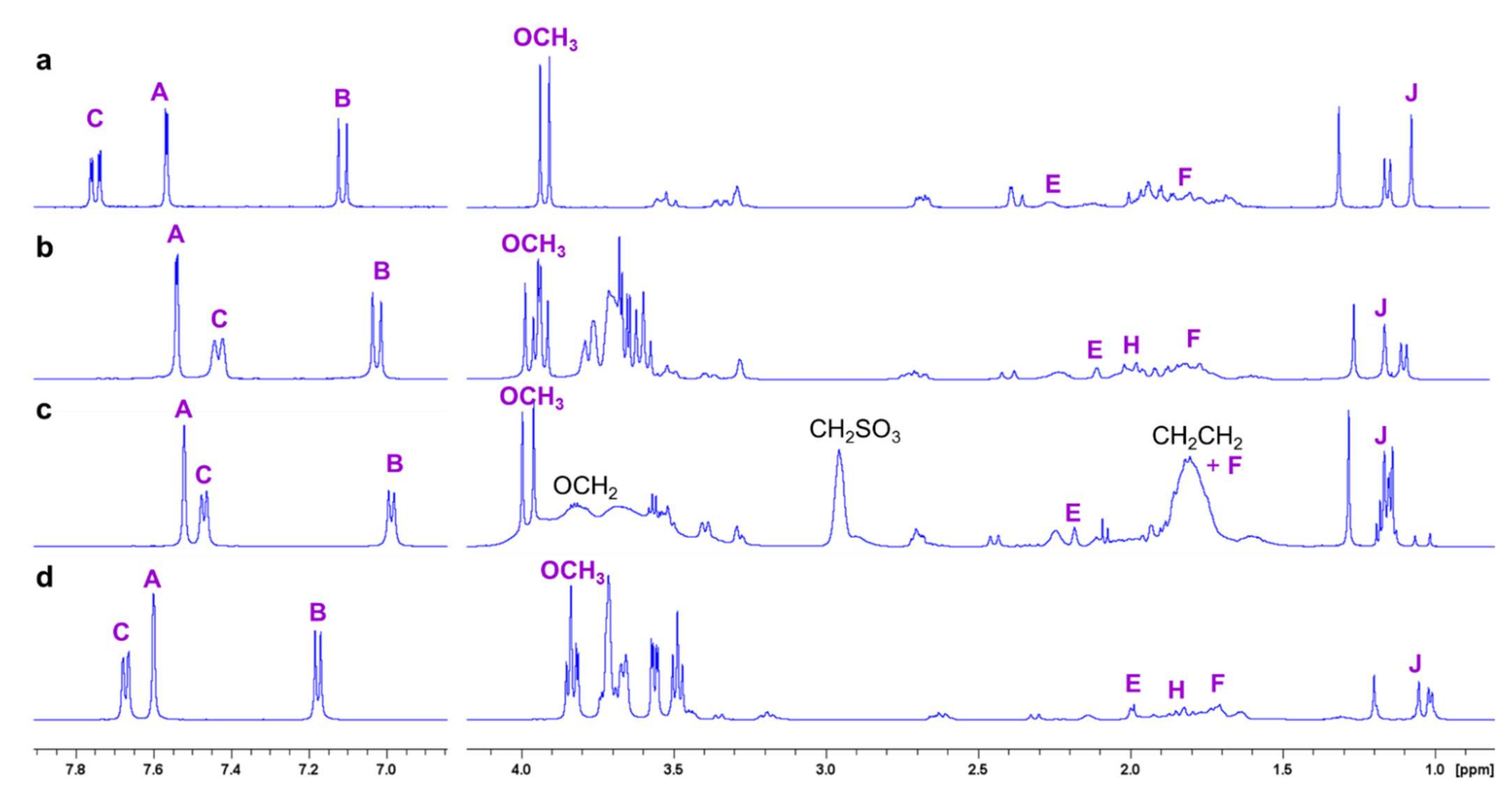
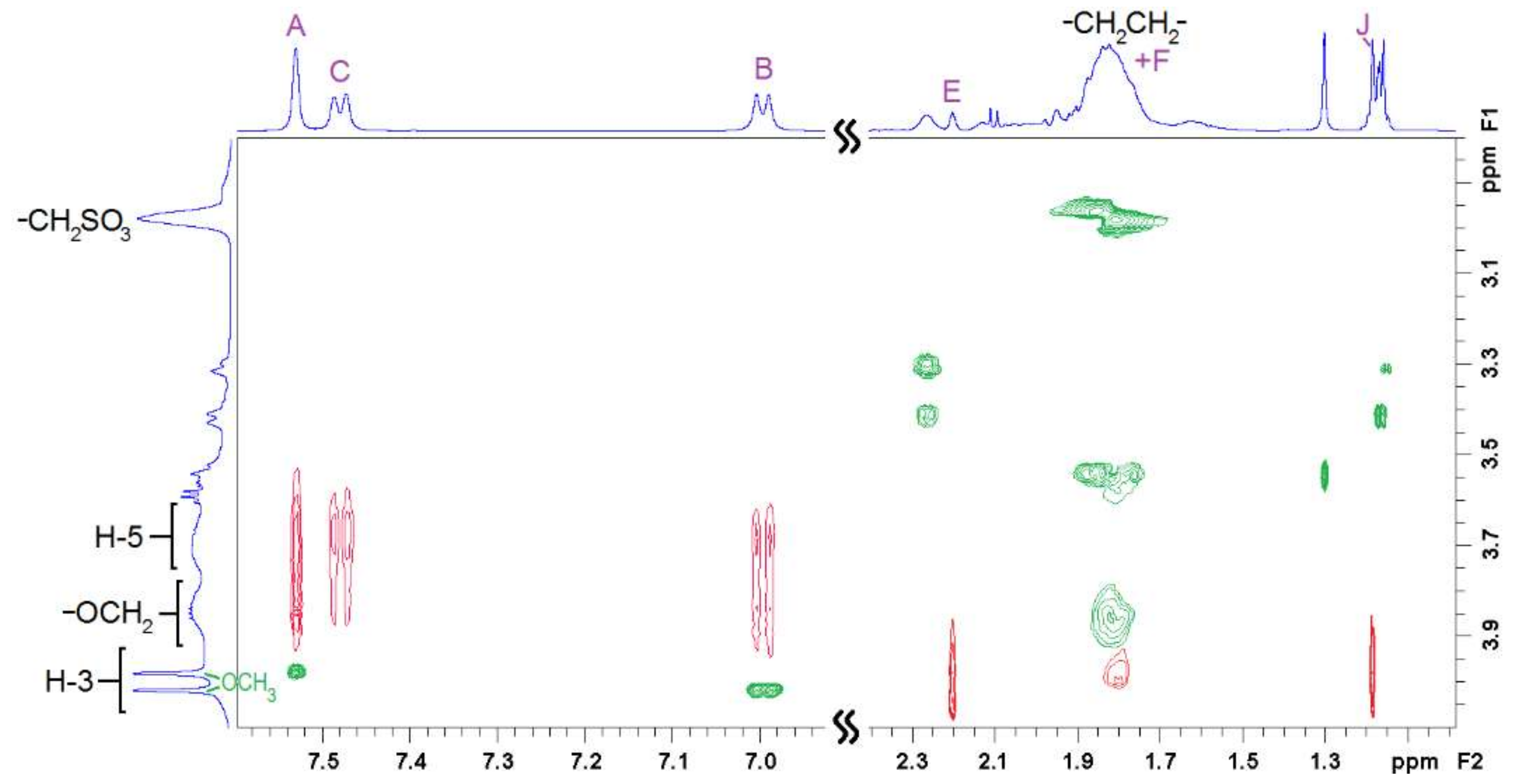
2.2. NMR Experiments
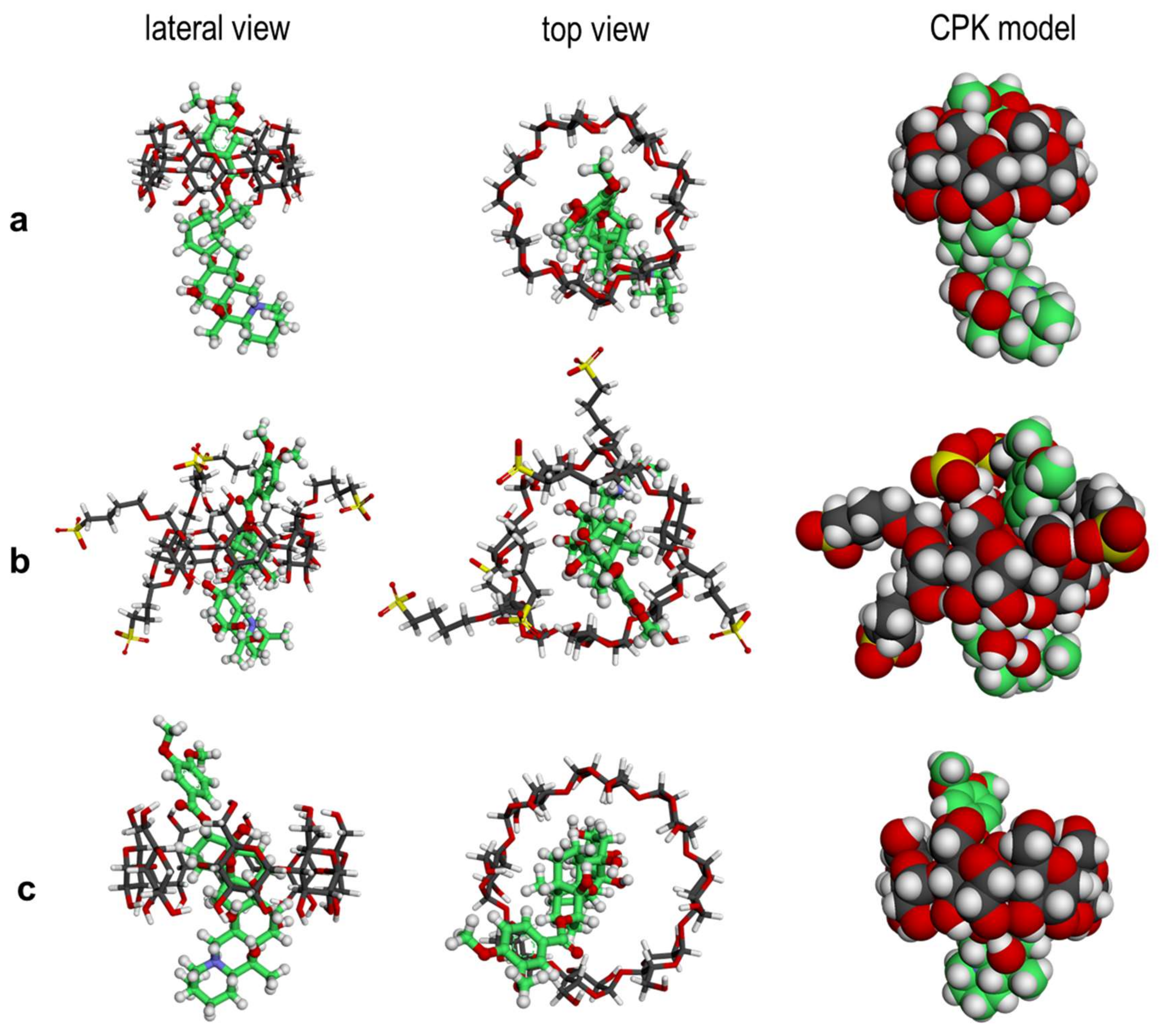
2.3. Molecular Simulations
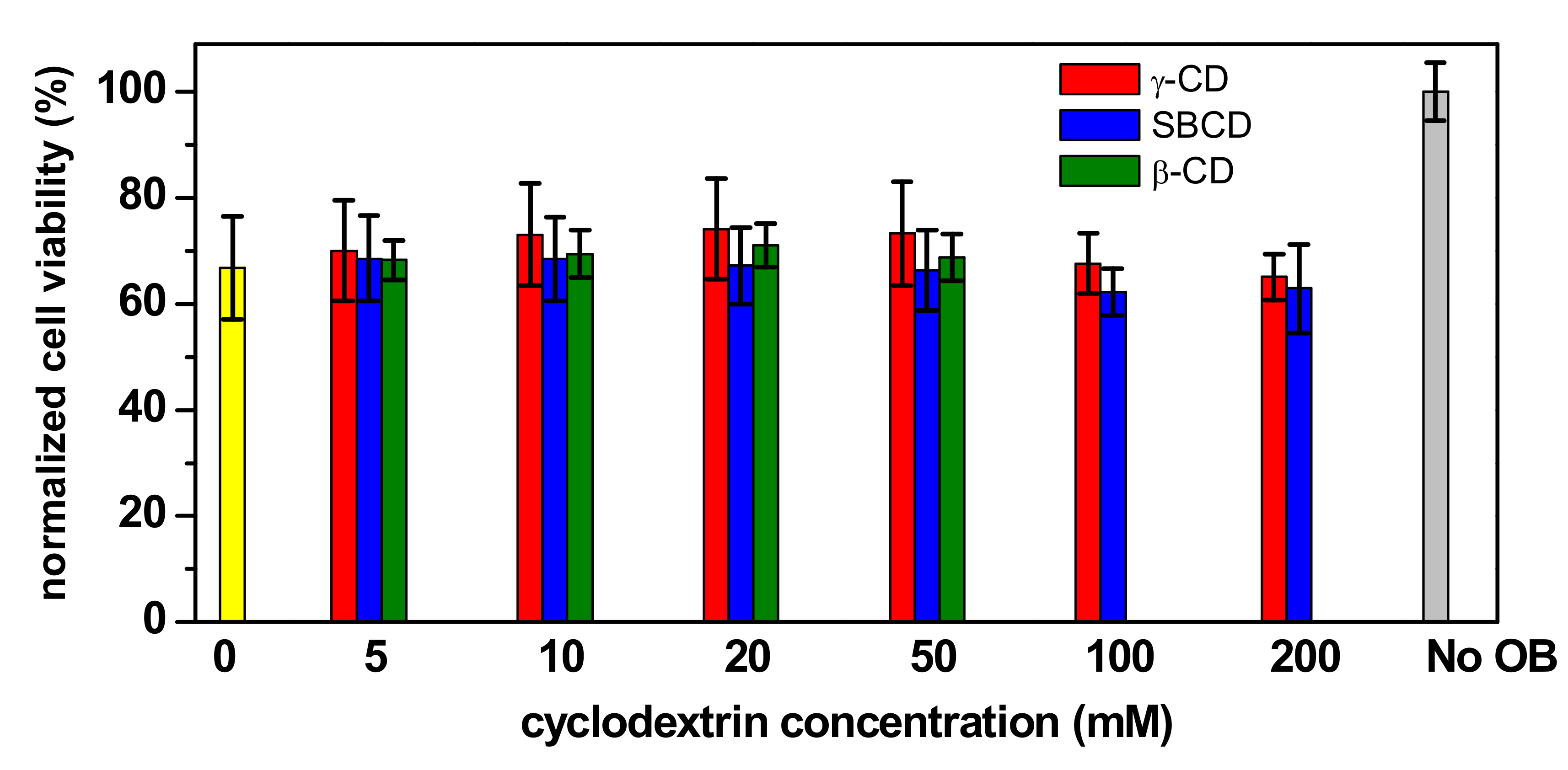
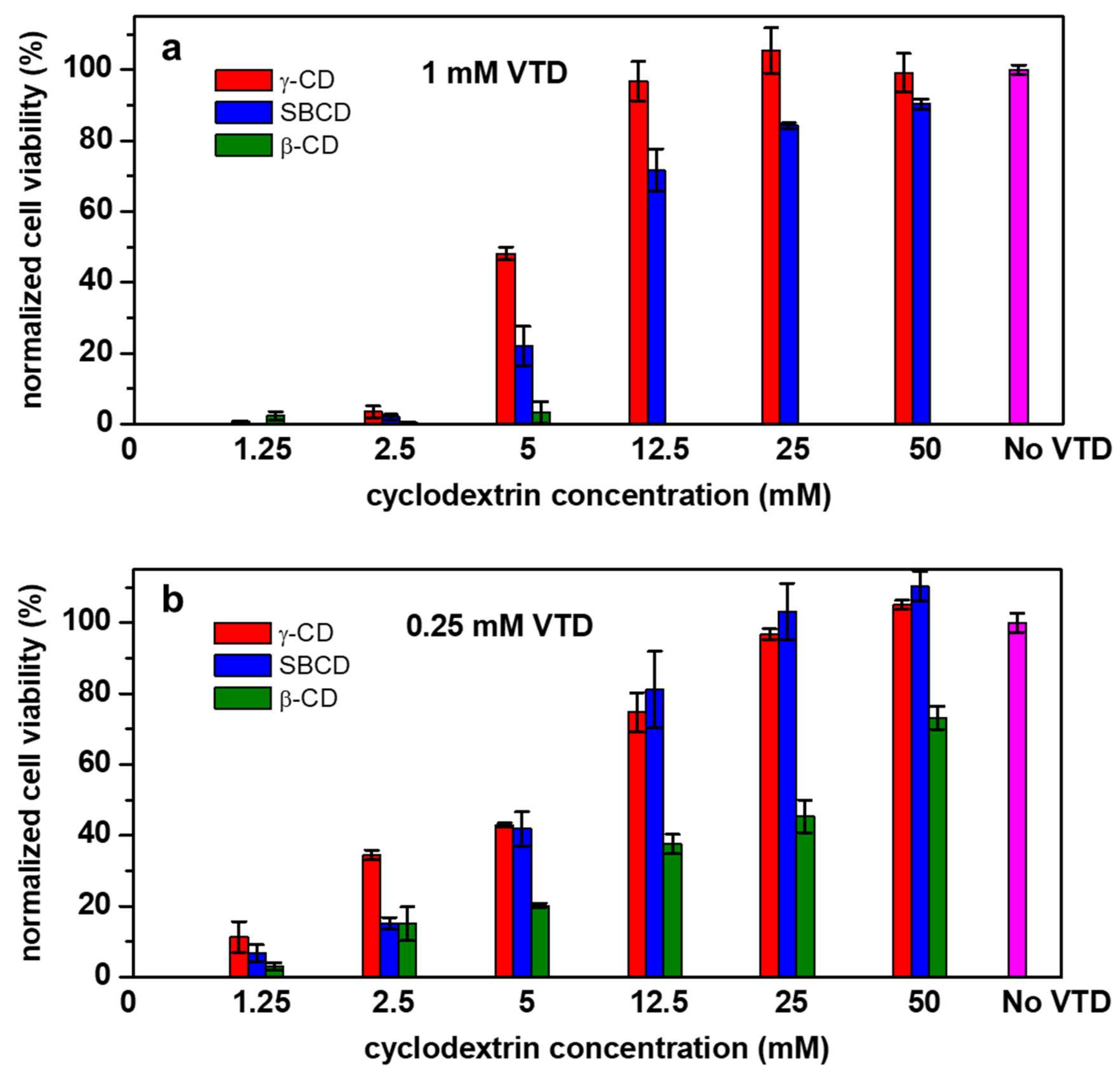
2.4. Cytotoxicity Evaluation
3. Materials and Methods
3.1. Isothermal Titration Calorimetry
3.2. NMR Spectroscopy
3.3. Molecular Simulation
3.3.1. Structure Preparation
3.3.2. Docking Studies
3.3.3. Molecular Dynamics Experiments
3.4. Neuro-2a Cell Viability Experiments
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Wang, S.-Y.; Wang, G.K. Voltage-gated sodium channels as primary targets of diverse lipid-soluble neurotoxins. Cell. Signal. 2003, 15, 151–159. [Google Scholar] [CrossRef]
- Caillaud, A.; Eixarch, H.; de la Iglesia, P.; Rodriguez, M.; Dominguez, L.; Andree, K.B.; Diogène, J. Towards the standardisation of the neuroblastoma (neuro-2a) cell-based assay for ciguatoxin-like toxicity detection in fish: Application to fish caught in the Canary Islands. Food Addit. Contam. Part A. 2012, 29, 1000–1010. [Google Scholar] [CrossRef] [PubMed]
- Mcdonough, J.H.; Jaax, N.K.; Crowley, R.A.; Mays, M.Z.; Modrow, H.E. Atropine and/or Diazepam Therapy Protects against Soman-Induced Neural and Cardiac Pathology. Fund. Appl. Toxicol. 1989, 13, 256–276. [Google Scholar] [CrossRef]
- Erecinfska, M.; Nelson, D.; Silver, I.A. Interactions of benztropine, atropine and ketamine with veratridine-activated sodium channels: Effects on membrane depolarization, K+-efflux and neutrotransmitter amino acid release. Br. J. Pharmacol. 1988, 94, 871–881. [Google Scholar] [CrossRef] [PubMed]
- Cataldi, M. Veratridine. Ref. Modul. Biomed. Sci. 2016, 1–9. [Google Scholar] [CrossRef]
- Del Valle, E.M.M. Cyclodextrins and their uses: A review. Process Biochem. 2004, 39, 1033–1046. [Google Scholar] [CrossRef]
- Szente, L.; Szejtli, J. Highly soluble cyclodextrin derivatives: Chemistry, properties, and trends in development. Adv. Drug Deliv. Rev. 1999, 36, 17–28. [Google Scholar] [CrossRef]
- Loftsson, T.; Fridhriksdottir, H.; Olafsdottir, B.J.; Gudhmundsson, O. Solubilization and stabilization of drugs through cyclodextrin complexation. Acta Pharm. Nord. 1991, 3, 215–217. [Google Scholar]
- Ho, S.; Thoo, Y.; Young, D.J.; Fong Siow, L. Cyclodextrin encapsulated catechin: Effect of pH, relative humidity and various food models on antioxidant stability. LWT 2017, 85, 232–239. [Google Scholar] [CrossRef]
- Mori, T.; Tsuchiya, R.; Doi, M.; Nagatani, N.; Tanaka, T. Solubilization of ultraviolet absorbers by cyclodextrin and their potential application in cosmetics. J. Incl. Phenom. Macrocycl. Chem. 2019, 93, 91–96. [Google Scholar] [CrossRef]
- Kfoury, M.; Landy, D.; Ruellan, S.; Auezova, L.; Greige-Gerges, H.; Fourmentin, S. Determination of formation constants and structural characterization of cyclodextrin inclusion complexes with two phenolic isomers: Carvacrol and thymol. Beilstein J. Org. Chem 2016, 12, 29–42. [Google Scholar] [CrossRef] [Green Version]
- Villalonga, R.; Cao, R.; Fragoso, A. Supramolecular chemistry of cyclodextrins in enzyme technology. Chem. Rev. 2007, 107, 3088–3116. [Google Scholar] [CrossRef] [PubMed]
- Landy, D.; Mallard, I.; Ponchel, A.; Monflier, E.; Fourmentin, S. Remediation technologies using cyclodextrins: An overview. Environ. Chem. Lett. 2012, 10, 225–237. [Google Scholar] [CrossRef]
- Loftsson, T.; Duchêne, D. Historical Perspectives Cyclodextrins and their pharmaceutical applications. Int. J. Pharm. 2007, 329, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Fliszár-Nyúl, E.; Lemli, B.; Kunsági-Máté, S.; Szente, L.; Poór, M. Interactions of Mycotoxin Alternariol with Cyclodextrins and Its Removal from Aqueous Solution by Beta-Cyclodextrin Bead Polymer. Biomolecules 2019, 9, 428. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ventrella, A.; Verrone, R.; Cosma, P.; Fini, P.; Longobardi, F.; Lippolis, V.; Pascale, M.; Catucci, L. Physico-Chemical Investigation on the Interaction between Ochratoxin A and Heptakis-2,6-di-O-Methyl-b-Cyclodextrin. J. Solut. Chem. 2014, 43, 1436–1447. [Google Scholar] [CrossRef]
- Amadasi, A.; Dall’Asta, C.; Ingletto, G.; Pela, R.; Marchelli, R.; Cozzini, P. Explaining cyclodextrin-mycotoxin interactions using a “natural” force field. Bioorganic Med. Chem. 2007, 15, 4585–4594. [Google Scholar] [CrossRef]
- Faisal, Z.; Kunsági-Máté, S.; Lemli, B.; Szente, L.; Bergmann, D.; Humpf, H.U.; Poór, M. Interaction of dihydrocitrinone with native and chemically modified cyclodextrins. Molecules 2019, 24, 1328. [Google Scholar] [CrossRef] [Green Version]
- Poór, M.; Matisz, G.; Kunsági-Máté, S.; Derdák, D.; Szente, L.; Lemli, B. Fluorescence spectroscopic investigation of the interaction of citrinin with native and chemically modified cyclodextrins. J. Lumin. 2015, 172, 23–28. [Google Scholar] [CrossRef]
- Poór, M.; Zand, A.; Szente, L.; Lemli, B.; Kunsági-Máté, S. Interaction of α-and β-zearalenols with β-cyclodextrins. Molecules 2017, 22, 1910. [Google Scholar] [CrossRef] [Green Version]
- Dernaika, H.; Chong, S.V.; Artur, C.G.; Tallon, J.L. Spectroscopic Identification of Neurotoxin Tetramethylenedisulfotetramine (TETS) Captured by Supramolecular Receptor beta-Cyclodextrin Immobilized on Nanostructured Gold Surfaces. J. Nanomater. 2014, 2014, 117. [Google Scholar] [CrossRef]
- Masurier, N.; Estour, F.; Froment, M.-T.; Lefèvre, B.; Debouzy, J.-C.; Brasme, B.; Masson, P.; Lafont, O. Synthesis of 2-substituted b-cyclodextrin derivatives with a hydrolytic activity against the organophosphorylester paraoxon. Eur. J. Med. Chem. 2005, 40, 615–623. [Google Scholar] [CrossRef] [PubMed]
- Campàs, M.; Rambla-Alegre, M.; Wirén, C.; Alcaraz, C.; Rey, M.; Safont, A.; Diogène, J.; Torréns, M.; Fragoso, A. Cyclodextrin polymers as passive sampling materials for lipophilic marine toxins in Prorocentrum lima cultures and a Dinophysis sacculus bloom in the NW Mediterranean Sea. Chemosphere 2021, 285, 131464. [Google Scholar] [CrossRef] [PubMed]
- Waldvogel, S. Book Review Cyclodextrins and Their Complexes: Chemistry, Analytical Methods, Applications; Dodziuk, H., Ed.; Wiley: Hoboken, NJ, USA, 2007; ISBN 9783527312801. [Google Scholar]
- Loftsson, T.; Saokham, P.; Sá Couto, A.R. Self-association of cyclodextrins and cyclodextrin complexes in aqueous solutions. Int. J. Pharm. 2019, 560, 228–234. [Google Scholar] [CrossRef] [PubMed]
- Do, T.T.; Van Hooghten, R.; Van den Mooter, G. A study of the aggregation of cyclodextrins: Determination of the critical aggregation concentration, size of aggregates and thermodynamics using isodesmic and K2–K models. Int. J. Pharm. 2017, 521, 318–326. [Google Scholar] [CrossRef] [PubMed]
- Mazzaferro, S.; Bouchemal, K.; Gallard, J.F.; Iorga, B.I.; Cheron, M.; Gueutin, C.; Steinmesse, C.; Ponchel, G. Bivalent sequential binding of docetaxel to methyl-β-cyclodextrin. Int. J. Pharm. 2011, 416, 171–180. [Google Scholar] [CrossRef] [PubMed]
- Bouchemal, K.; Mazzaferro, S. How to conduct and interpret ITC experiments accurately for cyclodextrin-guest interactions. Drug Discov. Today 2012, 17, 623–629. [Google Scholar] [CrossRef]
- Zheng, P.J.; Wang, C.; Hu, X.; Tam, K.C.; Li, L. Supramolecular complexes of azocellulose and α-cyclodextrin: Isothermal titration calorimetric and spectroscopic studies. Macromolecules 2005, 38, 2859–2864. [Google Scholar] [CrossRef]
- Sun, D.Z.; Li, L.; Qiu, X.M.; Liu, F.; Yin, B.L. Isothermal titration calorimetry and 1H NMR studies on host-guest interaction of paeonol and two of its isomers with β-cyclodextrin. Int. J. Pharm. 2006, 316, 7–13. [Google Scholar] [CrossRef]
- Forces, A. Solubility and pKa, of Veratridine. Anal. Biochem. 1986, 153, 33–38. [Google Scholar] [CrossRef]
- Veiga, F.J.B.; Fernandes, C.M.; Carvalho, R.A.; Geraldes, C.F.G.C. Molecular modelling and 1H-NMR: Ultimate tools for the investigation of tolbutamide: β-cyclodextrin and tolbutamide: Hydroxypropyl-β-cyclodextrin complexes. Chem. Pharm. Bull. 2001, 49, 1251–1256. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ganza-Gonzalez, A.; Vala-Jato, J.L.; Anguiano-Igea, S.; Otero-Espinar, F.; Blanco-Méndez, J. A proton nuclear magnetic resonance study of the inclusion complex of naproxen with beta-cyclodextrin. Int. J. Pharm. 1994, 106, 179–185. [Google Scholar] [CrossRef]
- Ma, D.Y.; Zhang, Y.M.; Xu, J.N. The synthesis and process optimization of sulfobutyl ether β-cyclodextrin derivatives. Tetrahedron 2016, 72, 3105–3112. [Google Scholar] [CrossRef]
- Li, T.; Guo, R.; Zong, Q.; Ling, G. Application of molecular docking in elaborating molecular mechanisms and interactions of supramolecular cyclodextrin. Carbohydr. Polym. 2022, 276, 118644. [Google Scholar] [CrossRef] [PubMed]
- Pantsar, T.; Poso, A. Binding affinity via docking: Fact and fiction. Molecules 2018, 23, 1899. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kuntz, I.D.; Blaney, J.M.; Oatley, S.J.; Langridge, R.; Ferrin, T.E. A Geometric Approach to Macromolecule-Ligand Interactions. J. Mol. Biol. 1982, 161, 269–288. [Google Scholar] [CrossRef]
- Genheden, S.; Ryde, U. The MM/PBSA and MM/GBSA methods to estimate ligand-binding affinities. Expert Opin. Drug Discov. 2015, 10, 449–461. [Google Scholar] [CrossRef]
- Manger, R.L.; Leja, L.S.; Lee, S.Y.; Hungerforg, J.M.; Wekell, M.M. Tetrazolium-based cell bioassay for neurotoxins active on voltage-sensitive sodium channels: Semiautomated assay for saxitoxins, brevetoxins, and ciguatoxins. Anal. Biochem. 1993, 214, 190–194. [Google Scholar] [CrossRef]
- Lingrel, J.B.; Kuntzweiler, T. Na+,K(+)-ATPase. J. Biol. Chem. 1994, 269, 19659–19662. [Google Scholar] [CrossRef]
- Caillaud, A.; Cañete, E.; de la Iglesia, P.; Giménez, G.; Diogène, J. Cell-based assay coupled with chromatographic fractioning: A strategy for marine toxins detection in natural samples. Toxicol. Vitr. 2009, 23, 1591–1596. [Google Scholar] [CrossRef]
- Cañete, E.; Diogène, J. Comparative study of the use of neuroblastoma cells (Neuro-2a) and neuroblastoma × glioma hybrid cells (NG108-15) for the toxic effect quantification of marine toxins. Toxicon 2008, 52, 541–550. [Google Scholar] [CrossRef] [PubMed]
- Ryzhakov, A.; Thi, T.D.; Stappaerts, J.; Bertoletti, L.; Kimpe, K.; Couto, A.R.S.; Saokham, P.; Mooter, G.V.D.; Augustijns, P.; Somsen, G.W.; et al. Self-Assembly of Cyclodextrins and Their Complexes in Aqueous Solutions. J. Pharm. Sci. 2016, 105, 2556–2569. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jain, A.S.; Date, A.A.; Pissurlenkar, R.R.S.; Coutinho, E.C.; Nagarsenker, M.S. Sulfobutyl Ether7 β-Cyclodextrin (SBE7 β-CD) Carbamazepine Complex: Preparation, Characterization, Molecular Modeling, and Evaluation of In Vivo Anti-epileptic Activity. AAPS PharmSciTech 2011, 12, 1163–1175. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shityakov, S.; Puskás, I.; Pápai, K.; Salvador, E.; Roewer, N.; Förster, C.; Broscheit, J.A. Sevoflurane-sulfobutylether-β-cyclodextrin complex: Preparation, characterization, cellular toxicity, molecular modeling and blood-brain barrier transport studies. Molecules 2015, 20, 10264–10279. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Harder, E.; Damm, W.; Maple, J.; Wu, C.; Reboul, M.; Xiang, J.Y.; Wang, L.; Lupyan, D.; Dahlgren, M.K.; Knight, J.L.; et al. OPLS3: A Force Field Providing Broad Coverage of Drug-like Small Molecules and Proteins. J. Chem. Theory Comput. 2015, 12, 281–296. [Google Scholar] [CrossRef] [PubMed]
- Codding, P.W. Structural Studies of Sodium Channel Neurotoxins. 2. Crystal Structure and Absolute Configuration of Veratridine Perchlorate. J. Am. Chem. Soc. 1983, 105, 3172–3176. [Google Scholar] [CrossRef]
- Friesner, R.A.; Banks, J.L.; Murphy, R.B.; Halgren, T.A.; Klicic, J.J.; Mainz, D.T.; Repasky, M.P.; Knoll, E.H.; Shelley, M.; Perry, J.K.; et al. Glide: A New Approach for Rapid, Accurate Docking and Scoring. 1. Method and Assessment of Docking Accuracy. J. Med. Chem. 2004, 47, 1739–1749. [Google Scholar] [CrossRef]
- Halgren, T.A.; Murphy, R.B.; Friesner, R.A.; Beard, H.S.; Frye, L.L.; Pollard, W.T.; Banks, J.L. Glide: A New Approach for Rapid, Accurate Docking and Scoring. 2. Enrichment Factors in Database Screening. J. Med. Chem. 2004, 47, 1750–1759. [Google Scholar] [CrossRef]
- Friesner, R.A.; Murphy, R.B.; Repasky, M.P.; Frye, L.L.; Greenwood, J.R.; Halgren, T.A.; Sanschagrin, P.C.; Mainz, D.T. Extra Precision Glide: Docking and Scoring Incorporating a Model of Hydrophobic Enclosure for Protein−Ligand Complexes. J. Med. Chem. 2006, 49, 6177–6196. [Google Scholar] [CrossRef] [Green Version]
- Weiner, P.K.; Kollman, P.A. AMBER: Assisted model building with energy refinement. A general program for modeling molecules and their interactions. J. Comput. Chem. 1981, 2, 287–303. [Google Scholar] [CrossRef]
- Wang, J.; Wolf, R.M.; Caldwell, J.W.; Kollman, P.A.; Case, D.A. Development and testing of a general Amber force field. J. Comput. Chem. 2004, 25, 1157–1174. [Google Scholar] [CrossRef] [PubMed]
- Allouche, A. Software News and Updates Gabedit—A Graphical User Interface for Computational Chemistry Softwares. J. Comput. Chem. 2012, 32, 174–182. [Google Scholar] [CrossRef] [PubMed]
- Jorgensen, W.L.; Chandrasekhar, J.; Madura, J.D.; Impey, R.W.; Klein, M.L. Comparison of simple potential functions for simulating liquid water. J. Chem. Phys. 1983, 79, 926–935. [Google Scholar] [CrossRef]
- Wu, X.; Brooks, B.R. Self-guided Langevin dynamics simulation method. Chem. Phys. Lett. 2003, 381, 512–518. [Google Scholar] [CrossRef]
- Berendsen, H.J.C.; Postma, J.P.M.; Van Gunsteren, W.F.; Dinola, A.; Haak, J.R. Molecular dynamics with coupling to an external bath. J. Chem. Phys. 1984, 81, 3684–3690. [Google Scholar] [CrossRef] [Green Version]
- Mosmann, T. Rapid colorimetric assay for cellular growth and survival: Application to proliferation and cytotoxicity assays. J. Immunol. Methods 1983, 65, 55–63. [Google Scholar] [CrossRef]











{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| n | Keq (M−1) | ∆H° (kJ/mol) | ∆G° (kJ/mol) | ∆S° (kJ/mol) | |
|---|---|---|---|---|---|
| β-CD | 0.7 ± 0.2 | 1500 ± 70 | n.d. | −18.1 ± 0.1 | n.d. |
| γ-CD | 0.95 ± 0.03 | 7200 ± 100 | −16.7 ± 0.2 | −22.1 ± 0.1 | 0.21 ± 0.01 |
| SBCD | 0.94 ± 0.02 | 8200 ± 60 | −14.5 ± 0.1 | −22.4 ± 0.1 | 0.31 ± 0.01 |
| Keq (M−1) | ∆G° (kJ/mol) ITC | ∆G° (kJ/mol) MD | Docking Score (kJ/mol) | |
|---|---|---|---|---|
| β-CD | 1500 ± 70 | −18.1 ± 0.1 | −1.6 ± 1.3 | −5.6 |
| SBCD | 8200 ± 60 | −22.4 ± 0.1 | n.d. | −25.9 |
| γ-CD | 7200 ± 100 | −22.1 ± 0.1 | −18 ± 3 | −26.7 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Uribe, L.A.; Leonardo, S.; Nielsen, T.T.; Steinmann, C.; Campàs, M.; Fragoso, A. Supramolecular Complexes of Plant Neurotoxin Veratridine with Cyclodextrins and Their Antidote-like Effect on Neuro-2a Cell Viability. Pharmaceutics 2022, 14, 598. https://doi.org/10.3390/pharmaceutics14030598
Uribe LA, Leonardo S, Nielsen TT, Steinmann C, Campàs M, Fragoso A. Supramolecular Complexes of Plant Neurotoxin Veratridine with Cyclodextrins and Their Antidote-like Effect on Neuro-2a Cell Viability. Pharmaceutics. 2022; 14(3):598. https://doi.org/10.3390/pharmaceutics14030598
Chicago/Turabian StyleUribe, Laura A., Sandra Leonardo, Thorbjørn Terndrup Nielsen, Casper Steinmann, Mònica Campàs, and Alex Fragoso. 2022. "Supramolecular Complexes of Plant Neurotoxin Veratridine with Cyclodextrins and Their Antidote-like Effect on Neuro-2a Cell Viability" Pharmaceutics 14, no. 3: 598. https://doi.org/10.3390/pharmaceutics14030598
APA StyleUribe, L. A., Leonardo, S., Nielsen, T. T., Steinmann, C., Campàs, M., & Fragoso, A. (2022). Supramolecular Complexes of Plant Neurotoxin Veratridine with Cyclodextrins and Their Antidote-like Effect on Neuro-2a Cell Viability. Pharmaceutics, 14(3), 598. https://doi.org/10.3390/pharmaceutics14030598