
Single-Dose Immunogenic DNA Vaccines Coding for Live-Attenuated Alpha- and Flaviviruses


Abstract
1. Introduction: Advantages of iDNA as DNA Vaccines
2. Advantages of iDNA as Live Vaccines
3. Potential Role of iDNA Vaccines in Epidemic Preparedness
4. Immunogenic iDNA Vaccines for Alphaviruses
4.1. CHIKV iDNA Vaccines
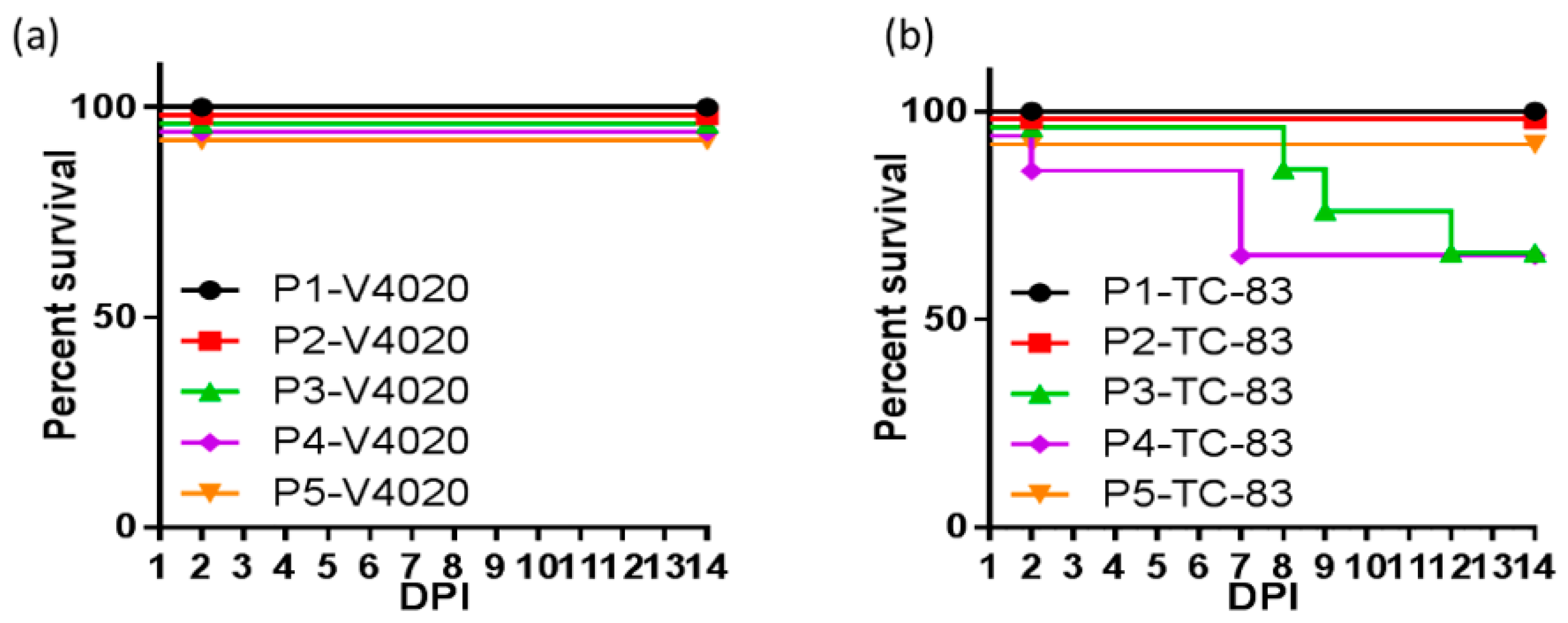
4.2. VEEV iDNA Vaccines
5. Immunogenic iDNA Vaccines for Flaviviruses
5.1. WNV Vaccines
5.2. iDNA Vaccine for JEV
5.3. ZIKV Vaccine
5.4. YFV iDNA Vaccine
6. Challenges and Perspectives for iDNA Vaccines
7. Discussion and Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Chen, K.; Zhang, L.; Fang, Z.; Li, J.; Li, C.; Song, W.; Huang, Z.; Chen, R.; Zhang, Y.; Li, J. Analysis of the protective efficacy of approved COVID-19 vaccines against Omicron variants and the prospects for universal vaccines. Front. Immunol. 2023, 14, 1294288. [Google Scholar] [CrossRef]
- Ferraro, B.; Morrow, M.P.; Hutnick, N.A.; Shin, T.H.; Lucke, C.E.; Weiner, D.B. Clinical applications of DNA vaccines: Current progress. Clin. Infect. Dis. 2011, 53, 296–302. [Google Scholar] [CrossRef]
- Kutzler, M.A.; Weiner, D.B. DNA vaccines: Ready for prime time? Nat. Rev. Genet. 2008, 9, 776–788. [Google Scholar] [CrossRef]
- Liu, M.A. DNA vaccines: An historical perspective and view to the future. Immunol. Rev. 2011, 239, 62–84. [Google Scholar] [CrossRef] [PubMed]
- Pagliari, S.; Dema, B.; Sanchez-Martinez, A.; Montalvo Zurbia-Flores, G.; Rollier, C.S. DNA Vaccines: History, Molecular Mechanisms and Future Perspectives. J. Mol. Biol. 2023, 435, 168297. [Google Scholar] [CrossRef]
- Sallberg, M.; Frelin, L.; Ahlen, G.; Sallberg-Chen, M. Electroporation for therapeutic DNA vaccination in patients. Med. Microbiol. Immunol. 2015, 204, 131–135. [Google Scholar] [CrossRef] [PubMed]
- Dupuy, L.C.; Schmaljohn, C.S. DNA vaccines for biodefense. Expert Rev. Vaccines 2009, 8, 1739–1754. [Google Scholar] [CrossRef] [PubMed]
- Schiller, J.T.; Lowy, D.R. Raising expectations for subunit vaccine. J. Infect. Dis. 2015, 211, 1373–1375. [Google Scholar] [CrossRef] [PubMed]
- Pushko, P.; Pujanauski, L.M.; Sun, X.; Pearce, M.; Hidajat, R.; Kort, T.; Schwartzman, L.M.; Tretyakova, I.; Chunqing, L.; Taubenberger, J.K.; et al. Recombinant H7 hemagglutinin forms subviral particles that protect mice and ferrets from challenge with H7N9 influenza virus. Vaccine 2015, 33, 4975–4982. [Google Scholar] [CrossRef]
- Pushko, P.; Pumpens, P.; Grens, E. Development of virus-like particle technology from small highly symmetric to large complex virus-like particle structures. Intervirology 2013, 56, 141–165. [Google Scholar] [CrossRef]
- Holtz, K.M.; Robinson, P.S.; Matthews, E.E.; Hashimoto, Y.; McPherson, C.E.; Khramtsov, N.; Reifler, M.J.; Meghrous, J.; Rhodes, D.G.; Cox, M.M.; et al. Modifications of cysteine residues in the transmembrane and cytoplasmic domains of a recombinant hemagglutinin protein prevents cross-linked multimer formation and potency loss. BMC Biotechnol. 2014, 14, 111. [Google Scholar] [CrossRef]
- Ramshaw, I.A.; Ramsay, A.J. The prime-boost strategy: Exciting prospects for improved vaccination. Immunol. Today 2000, 21, 163–165. [Google Scholar] [CrossRef]
- Villarreal, D.O.; Talbott, K.T.; Choo, D.K.; Shedlock, D.J.; Weiner, D.B. Synthetic DNA vaccine strategies against persistent viral infections. Expert Rev. Vaccines 2013, 12, 537–554. [Google Scholar] [CrossRef] [PubMed]
- Paulsen, G.C.; Frenck, R.; Tomashek, K.M.; Alarcon, R.M.; Hensel, E.; Lowe, A.; Brocato, R.L.; Kwilas, S.A.; Josleyn, M.D.; Hooper, J.W. Safety and Immunogenicity of an Andes Virus DNA Vaccine by Needle-Free Injection: A Randomized, Controlled Phase 1 Study. J. Infect. Dis. 2023, 229, 30–38. [Google Scholar] [CrossRef] [PubMed]
- Broderick, K.E.; Humeau, L.M. Electroporation-enhanced delivery of nucleic acid vaccines. Expert Rev. Vaccines 2015, 14, 195–204. [Google Scholar] [CrossRef] [PubMed]
- Lambricht, L.; Lopes, A.; Kos, S.; Sersa, G.; Preat, V.; Vandermeulen, G. Clinical potential of electroporation for gene therapy and DNA vaccine delivery. Expert Opin. Drug Deliv. 2015, 13, 295–310. [Google Scholar] [CrossRef] [PubMed]
- Dupuy, L.C.; Richards, M.J.; Ellefsen, B.; Chau, L.; Luxembourg, A.; Hannaman, D.; Livingston, B.D.; Schmaljohn, C.S. A DNA vaccine for venezuelan equine encephalitis virus delivered by intramuscular electroporation elicits high levels of neutralizing antibodies in multiple animal models and provides protective immunity to mice and nonhuman primates. Clin. Vaccine Immunol. 2011, 18, 707–716. [Google Scholar] [CrossRef] [PubMed]
- Kisakov, D.N.; Belyakov, I.M.; Kisakova, L.A.; Yakovlev, V.A.; Tigeeva, E.V.; Karpenko, L.I. The use of electroporation to deliver DNA-based vaccines. Expert Rev. Vaccines 2024, 23, 102–123. [Google Scholar] [CrossRef] [PubMed]
- Dupuy, L.C.; Richards, M.J.; Reed, D.S.; Schmaljohn, C.S. Immunogenicity and protective efficacy of a DNA vaccine against Venezuelan equine encephalitis virus aerosol challenge in nonhuman primates. Vaccine 2010, 28, 7345–7350. [Google Scholar] [CrossRef]
- Luke, J.M.; Simon, G.G.; Soderholm, J.; Errett, J.S.; August, J.T.; Gale, M., Jr.; Hodgson, C.P.; Williams, J.A. Coexpressed RIG-I agonist enhances humoral immune response to influenza virus DNA vaccine. J. Virol. 2011, 85, 1370–1383. [Google Scholar] [CrossRef]
- Williams, J.A. Improving DNA vaccine performance through vector design. Curr. Gene Ther. 2014, 14, 170–189. [Google Scholar] [CrossRef] [PubMed]
- Mairhofer, J.; Lara, A.R. Advances in host and vector development for the production of plasmid DNA vaccines. Methods Mol. Biol. 2014, 1139, 505–541. [Google Scholar] [CrossRef]
- Xenopoulos, A.; Pattnaik, P. Production and purification of plasmid DNA vaccines: Is there scope for further innovation? Expert Rev. Vaccines 2014, 13, 1537–1551. [Google Scholar] [CrossRef]
- Pushko, P.; Lukashevich, I.S.; Weaver, S.C.; Tretyakova, I. DNA-launched live-attenuated vaccines for biodefense applications. Expert Rev. Vaccines 2016, 15, 1223–1234. [Google Scholar] [CrossRef]
- Tretyakova, I.; Hearn, J.; Wang, E.; Weaver, S.; Pushko, P. DNA vaccine initiates replication of live attenuated chikungunya virus in vitro and elicits protective immune response in mice. J. Infect. Dis. 2014, 209, 1882–1890. [Google Scholar] [CrossRef]
- Tretyakova, I.; Lukashevich, I.S.; Glass, P.; Wang, E.; Weaver, S.; Pushko, P. Novel vaccine against Venezuelan equine encephalitis combines advantages of DNA immunization and a live attenuated vaccine. Vaccine 2013, 31, 1019–1025. [Google Scholar] [CrossRef] [PubMed]
- Tretyakova, I.; Nickols, B.; Hidajat, R.; Jokinen, J.; Lukashevich, I.S.; Pushko, P. Plasmid DNA initiates replication of yellow fever vaccine in vitro and elicits virus-specific immune response in mice. Virology 2014, 468–470, 28–35. [Google Scholar] [CrossRef] [PubMed]
- Hall, R.A.; Nisbet, D.J.; Pham, K.B.; Pyke, A.T.; Smith, G.A.; Khromykh, A.A. DNA vaccine coding for the full-length infectious Kunjin virus RNA protects mice against the New York strain of West Nile virus. Proc. Natl. Acad. Sci. USA 2003, 100, 10460–10464. [Google Scholar] [CrossRef]
- Mishin, V.P.; Cominelli, F.; Yamshchikov, V.F. A ‘minimal’ approach in design of flavivirus infectious DNA. Virus Res. 2001, 81, 113–123. [Google Scholar] [CrossRef]
- Yamshchikov, V. Development of a human live attenuated West Nile infectious DNA vaccine: Conceptual design of the vaccine candidate. Virology 2015, 484, 59–68. [Google Scholar] [CrossRef]
- Pushko, P.; Bredenbeek, P.P.; Lukashevich, I.S. Experimental DNA-launched live-attenuated vaccines against infections caused by Flavi-and alphaviruses. In Novel Technologies for Vaccine Development; Lukashevich, I., Shirwan, H., Eds.; Springer: Vienna, Austria, 2014. [Google Scholar]
- WHO. A Brief History of Vaccination. Available online: https://www.who.int/news-room/spotlight/history-of-vaccination/a-brief-history-of-vaccination (accessed on 1 February 2024).
- Minor, P.D. Live attenuated vaccines: Historical successes and current challenges. Virology 2015, 479–480, 379–392. [Google Scholar] [CrossRef]
- Coffey, L.L.; Beeharry, Y.; Borderia, A.V.; Blanc, H.; Vignuzzi, M. Arbovirus high fidelity variant loses fitness in mosquitoes and mice. Proc. Natl. Acad. Sci. USA 2011, 108, 16038–16043. [Google Scholar] [CrossRef]
- Weaver, S.C.; Bellew, L.A.; Gousset, L.; Repik, P.M.; Scott, T.W.; Holland, J.J. Diversity within natural populations of eastern equine encephalomyelitis virus. Virology 1993, 195, 700–709. [Google Scholar] [CrossRef]
- Plotkin, S.A. Vaccines: Past, present and future. Nat. Med. 2005, 11, S5–S11. [Google Scholar] [CrossRef]
- Hoke, C.H., Jr.; Pace-Templeton, J.; Pittman, P.; Malinoski, F.J.; Gibbs, P.; Ulderich, T.; Mathers, M.; Fogtman, B.; Glass, P.; Vaughn, D.W. US Military contributions to the global response to pandemic chikungunya. Vaccine 2012, 30, 6713–6720. [Google Scholar] [CrossRef]
- Kenney, J.L.; Volk, S.M.; Pandya, J.; Wang, E.; Liang, X.; Weaver, S.C. Stability of RNA virus attenuation approaches. Vaccine 2011, 29, 2230–2234. [Google Scholar] [CrossRef][Green Version]
- Ludwig, G.V.; Turell, M.J.; Vogel, P.; Kondig, J.P.; Kell, W.K.; Smith, J.F.; Pratt, W.D. Comparative neurovirulence of attenuated and non-attenuated strains of Venezuelan equine encephalitis virus in mice. Am. J. Trop. Med. Hyg. 2001, 64, 49–55. [Google Scholar] [CrossRef]
- Schneider, M.; Narciso-Abraham, M.; Hadl, S.; McMahon, R.; Toepfer, S.; Fuchs, U.; Hochreiter, R.; Bitzer, A.; Kosulin, K.; Larcher-Senn, J.; et al. Safety and immunogenicity of a single-shot live-attenuated chikungunya vaccine: A double-blind, multicentre, randomised, placebo-controlled, phase 3 trial. Lancet 2023, 401, 2138–2147. [Google Scholar] [CrossRef] [PubMed]
- Davis, E.H.; Thompson, J.K.; Widen, S.G.; Barrett, A.D.T. Genome Characterization of Yellow Fever Virus Wild-Type Strain Asibi, Parent to Live-Attenuated 17D Vaccine, from Three Different Sources. Viruses 2021, 13, 1383. [Google Scholar] [CrossRef] [PubMed]
- Salmona, M.; Gazaignes, S.; Mercier-Delarue, S.; Garnier, F.; Korimbocus, J.; Colin de Verdiere, N.; LeGoff, J.; Roques, P.; Simon, F. Molecular characterization of the 17D-204 yellow fever vaccine. Vaccine 2015, 33, 5432–5436. [Google Scholar] [CrossRef] [PubMed]
- WHO. Standard Operating Procedure. Mutant Analysis by PCR and Restriction Enzyme Cleavage (MAPREC) for Oral Poliovirus (Sabin) Vaccine Types 1, 2 or 3. Available online: http://www.who.int/biologicals/vaccines/MAPREC_SOP_Final_09112012.pdf (accessed on 27 May 2015).
- Gout, J.F.; Li, W.; Fritsch, C.; Li, A.; Haroon, S.; Singh, L.; Hua, D.; Fazelinia, H.; Smith, Z.; Seeholzer, S.; et al. The landscape of transcription errors in eukaryotic cells. Sci. Adv. 2017, 3, e1701484. [Google Scholar] [CrossRef]
- Jiang, X.; Dalebout, T.J.; Lukashevich, I.S.; Bredenbeek, P.J.; Franco, D. Molecular and immunological characterization of a DNA-launched yellow fever virus 17D infectious clone. J. Gen. Virol. 2015, 96, 804–814. [Google Scholar] [CrossRef]
- Hidajat, R.; Nickols, B.; Forrester, N.; Tretyakova, I.; Weaver, S.; Pushko, P. Next generation sequencing of DNA-launched Chikungunya vaccine virus. Virology 2016, 490, 83–90. [Google Scholar] [CrossRef]
- Barrett, A.D.; Beasley, D.W. Development pathway for biodefense vaccines. Vaccine 2009, 27 (Suppl. S4), D2–D7. [Google Scholar] [CrossRef]
- Han, J.J.; Song, H.A.; Pierson, S.L.; Shen-Gunther, J.; Xia, Q. Emerging Infectious Diseases Are Virulent Viruses-Are We Prepared? An Overview. Microorganisms 2023, 11, 2618. [Google Scholar] [CrossRef] [PubMed]
- Weaver, S.C.; Barrett, A.D. Transmission cycles, host range, evolution and emergence of arboviral disease. Nat. Rev. Microbiol. 2004, 2, 789–801. [Google Scholar] [CrossRef] [PubMed]
- Wolf, J.; Jannat, R.; Dubey, S.; Troth, S.; Onorato, M.T.; Coller, B.A.; Hanson, M.E.; Simon, J.K. Development of Pandemic Vaccines: ERVEBO Case Study. Vaccines 2021, 9, 190. [Google Scholar] [CrossRef] [PubMed]
- Walsh, S.R.; Dolin, R. Vaccinia viruses: Vaccines against smallpox and vectors against infectious diseases and tumors. Expert Rev. Vaccines 2011, 10, 1221–1240. [Google Scholar] [CrossRef] [PubMed]
- Clegg, C.H.; Rininger, J.A.; Baldwin, S.L. Clinical vaccine development for H5N1 influenza. Expert Rev. Vaccines 2013, 12, 767–777. [Google Scholar] [CrossRef] [PubMed]
- Mansur, D.S.; Smith, G.L.; Ferguson, B.J. Intracellular sensing of viral DNA by the innate immune system. Microbes Infect. 2014, 16, 1002–1012. [Google Scholar] [CrossRef] [PubMed]
- Ljungberg, K.; Whitmore, A.C.; Fluet, M.E.; Moran, T.P.; Shabman, R.S.; Collier, M.L.; Kraus, A.A.; Thompson, J.M.; Montefiori, D.C.; Beard, C.; et al. Increased immunogenicity of a DNA-launched Venezuelan equine encephalitis virus-based replicon DNA vaccine. J. Virol. 2007, 81, 13412–13423. [Google Scholar] [CrossRef] [PubMed]
- Bhat, N.; Fitzgerald, K.A. Recognition of cytosolic DNA by cGAS and other STING-dependent sensors. Eur. J. Immunol. 2014, 44, 634–640. [Google Scholar] [CrossRef] [PubMed]
- Coban, C.; Kobiyama, K.; Jounai, N.; Tozuka, M.; Ishii, K.J. DNA vaccines: A simple DNA sensing matter? Hum. Vaccin. Immunother. 2013, 9, 2216–2221. [Google Scholar] [CrossRef] [PubMed]
- Sharma, S.; Schmid, M.A.; Sanchez Felipe, L.; Grenelle, J.; Kaptein, S.J.F.; Coelmont, L.; Neyts, J.; Dallmeier, K. Small-molecule inhibitors of TBK1 serve as an adjuvant for a plasmid-launched live-attenuated yellow fever vaccine. Hum. Vaccines Immunother. 2020, 16, 2196–2203. [Google Scholar] [CrossRef] [PubMed]
- Wicker, S.; Seale, H.; von Gierke, L.; Maltezou, H. Vaccination of healthcare personnel: Spotlight on groups with underlying conditions. Vaccine 2014, 32, 4025–4031. [Google Scholar] [CrossRef]
- Haber, P.; Moro, P.L.; Cano, M.; Lewis, P.; Stewart, B.; Shimabukuro, T.T. Post-licensure surveillance of quadrivalent live attenuated influenza vaccine United States, Vaccine Adverse Event Reporting System (VAERS), July 2013–June 2014. Vaccine 2015, 33, 1987–1992. [Google Scholar] [CrossRef]
- Turner, P.J.; Southern, J.; Andrews, N.J.; Miller, E.; Erlewyn-Lajeunesse, M.; Investigators, S.-S. Safety of live attenuated influenza vaccine in young people with egg allergy: Multicentre prospective cohort study. BMJ 2015, 351, h6291. [Google Scholar] [CrossRef]
- Franceschini, F.; Bottau, P.; Caimmi, S.; Crisafulli, G.; Lucia, L.; Peroni, D.; Saretta, F.; Vernich, M.; Povesi Dascola, C.; Caffarelli, C. Vaccination in children with allergy to non active vaccine components. Clin. Transl. Med. 2015, 4, 3. [Google Scholar] [CrossRef]
- Lee, J.; Woodruff, M.C.; Kim, E.H.; Nam, J.H. Knife’s edge: Balancing immunogenicity and reactogenicity in mRNA vaccines. Exp. Mol. Med. 2023, 55, 1305–1313. [Google Scholar] [CrossRef]
- Tebas, P.; Roberts, C.C.; Muthumani, K.; Reuschel, E.L.; Kudchodkar, S.B.; Zaidi, F.I.; White, S.; Khan, A.S.; Racine, T.; Choi, H.; et al. Safety and Immunogenicity of an Anti-Zika Virus DNA Vaccine. N. Engl. J. Med. 2021, 385, e35. [Google Scholar] [CrossRef]
- Tretyakova, I.; Tomai, M.; Vasilakos, J.; Pushko, P. Live-Attenuated VEEV Vaccine Delivered by iDNA Using Microneedles Is Immunogenic in Rabbits. Front. Trop. Dis. 2022, 3, 813671. [Google Scholar] [CrossRef]
- Yamshchikov, V. Infectious DNA as a Vaccine against West Nile and Other Flavivirues. U.S. Patent US20050276816A1, 2 December 2008. [Google Scholar]
- Yamshchikov, V.; Manuvakhova, M.; Rodriguez, E. Development of a human live attenuated West Nile infectious DNA vaccine: Suitability of attenuating mutations found in SA14-14-2 for WN vaccine design. Virology 2015, 487, 198–206. [Google Scholar] [CrossRef]
- Yamshchikov, V.; Mishin, V.; Cominelli, F. A new strategy in design of +RNA virus infectious clones enabling their stable propagation in E. coli. Virology 2001, 281, 272–280. [Google Scholar] [CrossRef]
- Schwartz, O.; Albert, M.L. Biology and pathogenesis of chikungunya virus. Nat. Rev. Microbiol. 2010, 8, 491–500. [Google Scholar] [CrossRef]
- Strauss, J.H.; Strauss, E.G. The alphaviruses: Gene expression, replication, and evolution. Microbiol. Rev. 1994, 58, 491–562. [Google Scholar] [CrossRef]
- Bettis, A.A.; L’Azou Jackson, M.; Yoon, I.K.; Breugelmans, J.G.; Goios, A.; Gubler, D.J.; Powers, A.M. The global epidemiology of chikungunya from 1999 to 2020: A systematic literature review to inform the development and introduction of vaccines. PLoS Negl. Trop. Dis. 2022, 16, e0010069. [Google Scholar] [CrossRef]
- ECDC. Chikungunya Worldwide Overview. 2024. Available online: https://www.ecdc.europa.eu/en/chikungunya-monthly (accessed on 1 February 2024).
- Pumpens, P.; Pushko, P. Virus-Like Particles: A Comprehensive Guide; CRC Press: Boca Raton, FL, USA, 2022. [Google Scholar]
- Arankalle, V.A.; Shrivastava, S.; Cherian, S.; Gunjikar, R.S.; Walimbe, A.M.; Jadhav, S.M.; Sudeep, A.B.; Mishra, A.C. Genetic divergence of Chikungunya viruses in India (1963–2006) with special reference to the 2005–2006 explosive epidemic. J. Gen. Virol. 2007, 88, 1967–1976. [Google Scholar] [CrossRef]
- Weaver, S.C.; Reisen, W.K. Present and future arboviral threats. Antivir. Res. 2010, 85, 328–345. [Google Scholar] [CrossRef] [PubMed]
- Couderc, T.; Khandoudi, N.; Grandadam, M.; Visse, C.; Gangneux, N.; Bagot, S.; Prost, J.F.; Lecuit, M. Prophylaxis and therapy for Chikungunya virus infection. J. Infect. Dis. 2009, 200, 516–523. [Google Scholar] [CrossRef] [PubMed]
- Queyriaux, B.; Simon, F.; Grandadam, M.; Michel, R.; Tolou, H.; Boutin, J.P. Clinical burden of chikungunya virus infection. Lancet Infect. Dis. 2008, 8, 2–3. [Google Scholar] [CrossRef] [PubMed]
- Burt, F.J.; Rolph, M.S.; Rulli, N.E.; Mahalingam, S.; Heise, M.T. Chikungunya: A re-emerging virus. Lancet 2012, 379, 662–671. [Google Scholar] [CrossRef]
- Queyriaux, B.; Armengaud, A.; Jeannin, C.; Couturier, E.; Peloux-Petiot, F. Chikungunya in Europe. Lancet 2008, 371, 723–724. [Google Scholar] [CrossRef] [PubMed]
- Enserink, M. Entomology. A mosquito goes global. Science 2008, 320, 864–866. [Google Scholar] [CrossRef] [PubMed]
- Weaver, S.C.; Osorio, J.E.; Livengood, J.A.; Chen, R.; Stinchcomb, D.T. Chikungunya virus and prospects for a vaccine. Expert Rev. Vaccines 2012, 11, 1087–1101. [Google Scholar] [CrossRef] [PubMed]
- Dhiman, R.C.; Pahwa, S.; Dhillon, G.P.; Dash, A.P. Climate change and threat of vector-borne diseases in India: Are we prepared? Parasitol. Res. 2010, 106, 763–773. [Google Scholar] [CrossRef] [PubMed]
- Harris, E. Amid Paraguay Chikungunya Outbreak, Vaccine Candidate Safe, Immunogenic. JAMA 2023, 330, 111. [Google Scholar] [CrossRef] [PubMed]
- Padane, A.; Tegally, H.; Ramphal, Y.; Seyni, N.; Sarr, M.; Diop, M.M.; Diedhiou, C.K.; Mboup, A.; Diouf, N.D.; Souare, A.; et al. An emerging clade of Chikungunya West African genotype discovered in real-time during 2023 outbreak in Senegal. medRxiv 2023. [Google Scholar] [CrossRef]
- Pistone, T.; Ezzedine, K.; Schuffenecker, I.; Receveur, M.C.; Malvy, D. An imported case of Chikungunya fever from Madagascar: Use of the sentinel traveller for detecting emerging arboviral infections in tropical and European countries. Travel Med. Infect. Dis. 2009, 7, 52–54. [Google Scholar] [CrossRef] [PubMed]
- Randolph, S.E.; Rogers, D.J. The arrival, establishment and spread of exotic diseases: Patterns and predictions. Nat. Rev. Microbiol. 2010, 8, 361–371. [Google Scholar] [CrossRef]
- Weaver, S.C. Urbanization and geographic expansion of zoonotic arboviral diseases: Mechanisms and potential strategies for prevention. Trends Microbiol. 2013, 21, 360–363. [Google Scholar] [CrossRef]
- Thiboutot, M.M.; Kannan, S.; Kawalekar, O.U.; Shedlock, D.J.; Khan, A.S.; Sarangan, G.; Srikanth, P.; Weiner, D.B.; Muthumani, K. Chikungunya: A potentially emerging epidemic? PLoS Negl. Trop. Dis. 2010, 4, e623. [Google Scholar] [CrossRef] [PubMed]
- Petersen, L.R.; Stramer, S.L.; Powers, A.M. Chikungunya virus: Possible impact on transfusion medicine. Transfus. Med. Rev. 2010, 24, 15–21. [Google Scholar] [CrossRef]
- Schwameis, M.; Buchtele, N.; Wadowski, P.P.; Schoergenhofer, C.; Jilma, B. Chikungunya vaccines in development. Hum. Vaccin. Immunother. 2015, 12, 716–731. [Google Scholar] [CrossRef] [PubMed]
- Gorchakov, R.; Wang, E.; Leal, G.; Forrester, N.L.; Plante, K.; Rossi, S.L.; Partidos, C.D.; Adams, A.P.; Seymour, R.L.; Weger, J.; et al. Attenuation of Chikungunya virus vaccine strain 181/clone 25 is determined by two amino acid substitutions in the E2 envelope glycoprotein. J. Virol. 2012, 86, 6084–6096. [Google Scholar] [CrossRef]
- Plante, K.S.; Rossi, S.L.; Bergren, N.A.; Seymour, R.L.; Weaver, S.C. Extended Preclinical Safety, Efficacy and Stability Testing of a Live-attenuated Chikungunya Vaccine Candidate. PLoS Negl. Trop. Dis. 2015, 9, e0004007. [Google Scholar] [CrossRef]
- Edelman, R.; Tacket, C.O.; Wasserman, S.S.; Bodison, S.A.; Perry, J.G.; Mangiafico, J.A. Phase II safety and immunogenicity study of live chikungunya virus vaccine TSI-GSD-218. Am. J. Trop. Med. Hyg. 2000, 62, 681–685. [Google Scholar] [CrossRef] [PubMed]
- Levitt, N.H.; Ramsburg, H.H.; Hasty, S.E.; Repik, P.M.; Cole, F.E., Jr.; Lupton, H.W. Development of an attenuated strain of chikungunya virus for use in vaccine production. Vaccine 1986, 4, 157–162. [Google Scholar] [CrossRef]
- Mallilankaraman, K.; Shedlock, D.J.; Bao, H.; Kawalekar, O.U.; Fagone, P.; Ramanathan, A.A.; Ferraro, B.; Stabenow, J.; Vijayachari, P.; Sundaram, S.G.; et al. A DNA vaccine against chikungunya virus is protective in mice and induces neutralizing antibodies in mice and nonhuman primates. PLoS Negl. Trop. Dis. 2011, 5, e928. [Google Scholar] [CrossRef]
- Inovio. Inovio Pharmaceuticals Breakthrough DNA-Based Monoclonal Antibody Therapy Completely Protects Animals from Lethal Viral Challenge. 2014. Available online: http://ir.inovio.com/news/news-releases/news-releases-details/2014/Inovio-Pharmaceuticals-Breakthrough-DNA-Based-Monoclonal-Antibody-Therapy-Completely-Protects-Animals-From-Lethal-Viral-Challenge/ (accessed on 19 March 2016).
- Hallengard, D.; Kakoulidou, M.; Lulla, A.; Kummerer, B.M.; Johansson, D.X.; Mutso, M.; Lulla, V.; Fazakerley, J.K.; Roques, P.; Le Grand, R.; et al. Novel attenuated Chikungunya vaccine candidates elicit protective immunity in C57BL/6 mice. J. Virol. 2014, 88, 2858–2866. [Google Scholar] [CrossRef]
- Tretyakova, I.; Joh, J.; Lukashevich, I.S.; Alejandro, B.; Gearon, M.; Chung, D.; Pushko, P. Live-Attenuated CHIKV Vaccine with Rearranged Genome Replicates in vitro and Induces Immune Response in Mice. bioRxiv 2023. [Google Scholar] [CrossRef]
- Scherer, W.F.; Cupp, E.W.; Dziem, G.M.; Breener, R.J.; Ordonez, J.V. Mesenteronal infection threshold of an epizootic strain of Venezuelan encephalitis virus in Culex (Melanoconion) taeniopus mosquitoes and its implication to the apparent disappearance of this virus strain from an enzootic habitat in Guatemala. Am. J. Trop. Med. Hyg. 1982, 31, 1030–1037. [Google Scholar] [CrossRef]
- Brault, A.C.; Powers, A.M.; Ortiz, D.; Estrada-Franco, J.G.; Navarro-Lopez, R.; Weaver, S.C. Venezuelan equine encephalitis emergence: Enhanced vector infection from a single amino acid substitution in the envelope glycoprotein. Proc. Natl. Acad. Sci. USA 2004, 101, 11344–11349. [Google Scholar] [CrossRef]
- Deardorff, E.R.; Weaver, S.C. Vector competence of Culex (Melanoconion) taeniopus for equine-virulent subtype IE strains of Venezuelan equine encephalitis virus. Am. J. Trop. Med. Hyg. 2010, 82, 1047–1052. [Google Scholar] [CrossRef] [PubMed]
- Weaver, S.C.; Ferro, C.; Barrera, R.; Boshell, J.; Navarro, J.C. Venezuelan equine encephalitis. Annu. Rev. Entomol. 2004, 49, 141–174. [Google Scholar] [CrossRef] [PubMed]
- Scherer, W.F.; Weaver, S.C.; Taylor, C.A.; Cupp, E.W.; Dickerman, R.W.; Rubino, H.H. Vector competence of Culex (Melanoconion) taeniopus for allopatric and epizootic Venezuelan equine encephalomyelitis viruses. Am. J. Trop. Med. Hyg. 1987, 36, 194–197. [Google Scholar] [CrossRef]
- Smart, D.L.; Trainer, D.O. Serologic evidence of Venezuelan equine encephalitis in some wild and domestic populations of southern Texas. J. Wildl. Dis. 1975, 11, 195–200. [Google Scholar] [CrossRef]
- Weaver, S.C.; Pfeffer, M.; Marriott, K.; Kang, W.; Kinney, R.M. Genetic evidence for the origins of Venezuelan equine encephalitis virus subtype IAB outbreaks. Am. J. Trop. Med. Hyg. 1999, 60, 441–448. [Google Scholar] [CrossRef] [PubMed]
- Rivera, L.F.; Lezcano-Coba, C.; Galue, J.; Rodriguez, X.; Juarez, Y.; de Souza, W.M.; Capitan-Barrios, Z.; Valderrama, A.; Abrego, L.; Cedeno, H.; et al. Clinical and epidemiological characteristics of Madariaga and Venezuelan equine encephalitis virus infections. medRxiv 2024. [Google Scholar] [CrossRef]
- Kitchen, L.W.; Vaughn, D.W. Role of U.S. military research programs in the development of U.S.-licensed vaccines for naturally occurring infectious diseases. Vaccine 2007, 25, 7017–7030. [Google Scholar] [CrossRef]
- Lemon, S.M. Protecting Our Forces: Improving Vaccine Acquisition and Availability in the U.S. Military. Institute of Medicine (U.S.). Committee on a Strategy for Minimizing the Impact of Naturally Occurring Infectious Diseases of Military Importance; National Academies Press: Washington, DC, USA, 2002. [Google Scholar]
- Paessler, S.; Weaver, S.C. Vaccines for Venezuelan equine encephalitis. Vaccine 2009, 27 (Suppl. S4), D80–D85. [Google Scholar] [CrossRef]
- Sharma, A.; Knollmann-Ritschel, B. Current Understanding of the Molecular Basis of Venezuelan Equine Encephalitis Virus Pathogenesis and Vaccine Development. Viruses 2019, 11, 164. [Google Scholar] [CrossRef]
- Kinney, R.M.; Chang, G.J.; Tsuchiya, K.R.; Sneider, J.M.; Roehrig, J.T.; Woodward, T.M.; Trent, D.W. Attenuation of Venezuelan equine encephalitis virus strain TC-83 is encoded by the 5′-noncoding region and the E2 envelope glycoprotein. J. Virol. 1993, 67, 1269–1277. [Google Scholar] [CrossRef]
- Phillpotts, R.J.; Wright, A.J. TC-83 vaccine protects against airborne or subcutaneous challenge with heterologous mouse-virulent strains of Venezuelan equine encephalitis virus. Vaccine 1999, 17, 982–988. [Google Scholar] [CrossRef]
- Grabenstein, J.D.; Pittman, P.R.; Greenwood, J.T.; Engler, R.J. Immunization to protect the US Armed Forces: Heritage, current practice, and prospects. Epidemiol. Rev. 2006, 28, 3–26. [Google Scholar] [CrossRef]
- Hoke, C.H., Jr. History of U.S. military contributions to the study of viral encephalitis. Mil. Med. 2005, 170, 92–105. [Google Scholar] [CrossRef]
- Pittman, P.R.; Makuch, R.S.; Mangiafico, J.A.; Cannon, T.L.; Gibbs, P.H.; Peters, C.J. Long-term duration of detectable neutralizing antibodies after administration of live-attenuated VEE vaccine and following booster vaccination with inactivated VEE vaccine. Vaccine 1996, 14, 337–343. [Google Scholar] [CrossRef] [PubMed]
- Perri, S.; Greer, C.E.; Thudium, K.; Doe, B.; Legg, H.; Liu, H.; Romero, R.E.; Tang, Z.; Bin, Q.; Dubensky, T.W., Jr.; et al. An alphavirus replicon particle chimera derived from venezuelan equine encephalitis and sindbis viruses is a potent gene-based vaccine delivery vector. J. Virol. 2003, 77, 10394–10403. [Google Scholar] [CrossRef] [PubMed]
- Polo, J.M.; Gardner, J.P.; Ji, Y.; Belli, B.A.; Driver, D.A.; Sherrill, S.; Perri, S.; Liu, M.A.; Dubensky, T.W., Jr. Alphavirus DNA and particle replicons for vaccines and gene therapy. Dev. Biol. 2000, 104, 181–185. [Google Scholar]
- Riabov, V.; Tretyakova, I.; Alexander, R.B.; Pushko, P.; Klyushnenkova, E.N. Anti-tumor effect of the alphavirus-based virus-like particle vector expressing prostate-specific antigen in a HLA-DR transgenic mouse model of prostate cancer. Vaccine 2015, 33, 5386–5395. [Google Scholar] [CrossRef] [PubMed]
- Nelson, E.L.; Prieto, D.; Alexander, T.G.; Pushko, P.; Lofts, L.A.; Rayner, J.O.; Kamrud, K.I.; Fralish, B.; Smith, J.F. Venezuelan equine encephalitis replicon immunization overcomes intrinsic tolerance and elicits effective anti-tumor immunity to the ‘self’ tumor-associated antigen, neu in a rat mammary tumor model. Breast Cancer Res. Treat. 2003, 82, 169–183. [Google Scholar] [CrossRef] [PubMed]
- Pushko, P.; Tretyakova, I. Alphavirus Replicon Vectors for Prophylactic Applications and Cancer Interventions. In Novel Technologies for Vaccine Development; Lukashevich, I.S., Shirwan, H., Eds.; Springer: Vienna, Austria, 2014; pp. 61–87. [Google Scholar]
- Pushko, P.; Parker, M.; Ludwig, G.V.; Davis, N.L.; Johnston, R.E.; Smith, J.F. Replicon-helper systems from attenuated Venezuelan equine encephalitis virus: Expression of heterologous genes in vitro and immunization against heterologous pathogens in vivo. Virology 1997, 239, 389–401. [Google Scholar] [CrossRef]
- Geisbert, T.W.; Pushko, P.; Anderson, K.; Smith, J.; Davis, K.J.; Jahrling, P.B. Evaluation in nonhuman primates of vaccines against Ebola virus. Emerg. Infect. Dis. 2002, 8, 503–507. [Google Scholar] [CrossRef]
- Pushko, P.; Geisbert, J.; Parker, M.; Jahrling, P.; Smith, J. Individual and bivalent vaccines based on alphavirus replicons protect guinea pigs against infection with Lassa and Ebola viruses. J. Virol. 2001, 75, 11677–11685. [Google Scholar] [CrossRef]
- Pushko, P.; Bray, M.; Ludwig, G.V.; Parker, M.; Schmaljohn, A.; Sanchez, A.; Jahrling, P.B.; Smith, J.F. Recombinant RNA replicons derived from attenuated Venezuelan equine encephalitis virus protect guinea pigs and mice from Ebola hemorrhagic fever virus. Vaccine 2000, 19, 142–153. [Google Scholar] [CrossRef]
- Ong, E.Z.; Yee, J.X.; Ooi, J.S.G.; Syenina, A.; de Alwis, R.; Chen, S.; Sim, J.X.Y.; Kalimuddin, S.; Leong, Y.S.; Chan, Y.F.Z.; et al. Immune gene expression analysis indicates the potential of a self-amplifying COVID-19 mRNA vaccine. NPJ Vaccines 2022, 7, 154. [Google Scholar] [CrossRef] [PubMed]
- Oda, Y.; Kumagai, Y.; Kanai, M.; Iwama, Y.; Okura, I.; Minamida, T.; Yagi, Y.; Kurosawa, T.; Greener, B.; Zhang, Y.; et al. Immunogenicity and safety of a booster dose of a self-amplifying RNA COVID-19 vaccine (ARCT-154) versus BNT162b2 mRNA COVID-19 vaccine: A double-blind, multicentre, randomised, controlled, phase 3, non-inferiority trial. Lancet Infect. Dis. 2023. [Google Scholar] [CrossRef] [PubMed]
- Johnson, D.M.; Sokoloski, K.J.; Jokinen, J.D.; Pfeffer, T.L.; Chu, Y.K.; Adcock, R.S.; Chung, D.; Tretyakova, I.; Pushko, P.; Lukashevich, I.S. Advanced Safety and Genetic Stability in Mice of a Novel DNA-Launched Venezuelan Equine Encephalitis Virus Vaccine with Rearranged Structural Genes. Vaccines 2020, 8, 114. [Google Scholar] [CrossRef] [PubMed]
- Tretyakova, I.; Plante, K.S.; Rossi, S.L.; Lawrence, W.S.; Peel, J.E.; Gudjohnsen, S.; Wang, E.; Mirchandani, D.; Tibbens, A.; Lamichhane, T.N.; et al. Venezuelan equine encephalitis vaccine with rearranged genome resists reversion and protects non-human primates from viremia after aerosol challenge. Vaccine 2020, 38, 3378–3386. [Google Scholar] [CrossRef] [PubMed]
- Tretyakova, I.; Tibbens, A.; Jokinen, J.D.; Johnson, D.M.; Lukashevich, I.S.; Pushko, P. Novel DNA-launched Venezuelan equine encephalitis virus vaccine with rearranged genome. Vaccine 2019, 37, 3317–3325. [Google Scholar] [CrossRef] [PubMed]
- Berge, T.O.; Banks, I.S.; Tigertt, W.D. Attenuation of Venezuelan equine encephalomyelitis virus by in vitro cultivation in guinea-pig heart cells. Am. J. Trop. Med. Hyg. 1961, 73, 209–218. [Google Scholar] [CrossRef]
- Cheng, B.Y.; Ortiz-Riano, E.; de la Torre, J.C.; Martinez-Sobrido, L. Arenavirus Genome Rearrangement for the Development of Live Attenuated Vaccines. J. Virol. 2015, 89, 7373–7384. [Google Scholar] [CrossRef]
- Lowen, A.C.; Boyd, A.; Fazakerley, J.K.; Elliott, R.M. Attenuation of bunyavirus replication by rearrangement of viral coding and noncoding sequences. J. Virol. 2005, 79, 6940–6946. [Google Scholar] [CrossRef] [PubMed]
- Pushko, P.; Parker, M.D.; Smith, J.F.; Crise, B.J.; Hart, M.K. Live Attenuated Venezuelan Equine Encephalitis Vaccine. U.S. Patent 6296854, 2 October 2001. [Google Scholar]
- Reisen, W.K. Ecology of West Nile virus in North America. Viruses 2013, 5, 2079–2105. [Google Scholar] [CrossRef]
- CDC. West Nile Virus. 2023. Available online: https://www.cdc.gov/westnile/index.html (accessed on 1 February 2024).
- Petersen, L.R.; Brault, A.C.; Nasci, R.S. West Nile virus: Review of the literature. JAMA 2013, 310, 308–315. [Google Scholar] [CrossRef] [PubMed]
- Frost, M.J.; Zhang, J.; Edmonds, J.H.; Prow, N.A.; Gu, X.; Davis, R.; Hornitzky, C.; Arzey, K.E.; Finlaison, D.; Hick, P.; et al. Characterization of virulent West Nile virus Kunjin strain, Australia, 2011. Emerg. Infect. Dis. 2012, 18, 792–800. [Google Scholar] [CrossRef] [PubMed]
- Zhang, W.; Yin, Q.; Wang, H.; Liang, G. The reemerging and outbreak of genotypes 4 and 5 of Japanese encephalitis virus. Front. Cell Infect. Microbiol. 2023, 13, 1292693. [Google Scholar] [CrossRef] [PubMed]
- Halstead, S.B.; Thomas, S.J. New Japanese encephalitis vaccines: Alternatives to production in mouse brain. Expert Rev. Vaccines 2011, 10, 355–364. [Google Scholar] [CrossRef] [PubMed]
- CDC. Japanese Encephalitis Vaccine. 2023. Available online: https://www.cdc.gov/japaneseencephalitis/vaccine/index.html (accessed on 1 February 2024).
- Nickols, B.; Tretyakova, I.; Tibbens, A.; Klyushnenkova, E.; Pushko, P. Plasmid DNA launches live-attenuated Japanese encephalitis virus and elicits virus-neutralizing antibodies in BALB/c mice. Virology 2017, 512, 66–73. [Google Scholar] [CrossRef]
- Tsetsarkin, K.A.; Kenney, H.; Chen, R.; Liu, G.; Manukyan, H.; Whitehead, S.S.; Laassri, M.; Chumakov, K.; Pletnev, A.G. A Full-Length Infectious cDNA Clone of Zika Virus from the 2015 Epidemic in Brazil as a Genetic Platform for Studies of Virus-Host Interactions and Vaccine Development. MBio 2016, 7, e01114-16. [Google Scholar] [CrossRef]
- Oyono, M.G.; Kenmoe, S.; Abanda, N.N.; Takuissu, G.R.; Ebogo-Belobo, J.T.; Kenfack-Momo, R.; Kengne-Nde, C.; Mbaga, D.S.; Tchatchouang, S.; Kenfack-Zanguim, J.; et al. Epidemiology of yellow fever virus in humans, arthropods, and non-human primates in sub-Saharan Africa: A systematic review and meta-analysis. PLoS Negl. Trop. Dis. 2022, 16, e0010610. [Google Scholar] [CrossRef]
- Hansen, C.A.; Barrett, A.D.T. The Present and Future of Yellow Fever Vaccines. Pharmaceuticals 2021, 14, 891. [Google Scholar] [CrossRef] [PubMed]
- Tuells, J.; Henao-Martinez, A.F.; Franco-Paredes, C. Yellow Fever: A Perennial Threat. Arch. Med. Res. 2022, 53, 649–657. [Google Scholar] [CrossRef]
- WHO. Eliminate Yellow Fever Epidemics (EYE) Strategy 2017–2026. Available online: https://www.who.int/initiatives/eye-strategy (accessed on 1 February 2024).
- Rice, C.M.; Grakoui, A.; Galler, R.; Chambers, T.J. Transcription of infectious yellow fever RNA from full-length cDNA templates produced by in vitro ligation. New Biol. 1989, 1, 285–296. [Google Scholar]
- Kilchert, C.; Wittmann, S.; Vasiljeva, L. The regulation and functions of the nuclear RNA exosome complex. Nat. Rev. Mol. Cell Biol. 2016, 17, 227–239. [Google Scholar] [CrossRef] [PubMed]
- Guirakhoo, F.; Arroyo, J.; Pugachev, K.V.; Miller, C.; Zhang, Z.X.; Weltzin, R.; Georgakopoulos, K.; Catalan, J.; Ocran, S.; Soike, K.; et al. Construction, safety, and immunogenicity in nonhuman primates of a chimeric yellow fever-dengue virus tetravalent vaccine. J. Virol. 2001, 75, 7290–7304. [Google Scholar] [CrossRef]
- Sharma, S.; Dallmeier, K. Use of Optical In Vivo Imaging to Monitor and Optimize Delivery of Novel Plasmid-Launched Live-Attenuated Vaccines. Methods Mol. Biol. 2022, 2412, 283–294. [Google Scholar] [CrossRef] [PubMed]
- Cohen, J. Unfilled Vials. Science 2016, 351, 16–19. [Google Scholar] [CrossRef]
- Lukashevich, I.S.; Carrion, R., Jr.; Salvato, M.S.; Mansfield, K.; Brasky, K.; Zapata, J.; Cairo, C.; Goicochea, M.; Hoosien, G.E.; Ticer, A.; et al. Safety, immunogenicity, and efficacy of the ML29 reassortant vaccine for Lassa fever in small non-human primates. Vaccine 2008, 26, 5246–5254. [Google Scholar] [CrossRef]
- Sullivan, N.J.; Hensley, L.; Asiedu, C.; Geisbert, T.W.; Stanley, D.; Johnson, J.; Honko, A.; Olinger, G.; Bailey, M.; Geisbert, J.B.; et al. CD8+ cellular immunity mediates rAd5 vaccine protection against Ebola virus infection of nonhuman primates. Nat. Med. 2011, 17, 1128–1131. [Google Scholar] [CrossRef]
- Safronetz, D.; Mire, C.; Rosenke, K.; Feldmann, F.; Haddock, E.; Geisbert, T.; Feldmann, H. A recombinant vesicular stomatitis virus-based Lassa fever vaccine protects guinea pigs and macaques against challenge with geographically and genetically distinct Lassa viruses. PLoS Negl. Trop. Dis. 2015, 9, e0003736. [Google Scholar] [CrossRef]
- Kang, S.M.; Pushko, P.; Bright, R.A.; Smith, G.; Compans, R.W. Influenza virus-like particles as pandemic vaccines. Curr. Top. Microbiol. Immunol. 2009, 333, 269–289. [Google Scholar] [PubMed]
- Tretyakova, I.; Hidajat, R.; Hamilton, G.; Horn, N.; Nickols, B.; Prather, R.O.; Tumpey, T.M.; Pushko, P. Preparation of quadri-subtype influenza virus-like particles using bovine immunodeficiency virus gag protein. Virology 2016, 487, 163–171. [Google Scholar] [CrossRef] [PubMed]
- Akahata, W.; Yang, Z.Y.; Andersen, H.; Sun, S.; Holdaway, H.A.; Kong, W.P.; Lewis, M.G.; Higgs, S.; Rossmann, M.G.; Rao, S.; et al. A virus-like particle vaccine for epidemic Chikungunya virus protects nonhuman primates against infection. Nat. Med. 2010, 16, 334–338. [Google Scholar] [CrossRef] [PubMed]



{kind=link}
{kind=link}
| Vaccine Feature | Live-Attenuated Virus | mRNA | Traditional DNA | iDNA |
|---|---|---|---|---|
| Vaccine Formula | Live Virus | mRNA | Plasmid DNA | Plasmid DNA |
| Vaccine Antigen | Live Virus | Subunit | Subunit | Live Virus |
| Genetic stability | + | + | + | |
| Simple production | + | + | + | |
| High purity | + | + | + | |
| No cold chain | + | + | ||
| Single-dose vaccine | + | + | ||
| Rapid protection | + | + |
| Virus Family | Vaccine | Dose, In Vitro | Dose, In Vivo | Vaccine Route | Viremia | Neutralizing Antibody | Protection | References |
|---|---|---|---|---|---|---|---|---|
| Alpha | CHIKV | 5 ng | 10 μg | IM/EP | +/− | + (10/10) | + (10/10) | [25] |
| VEEV | 8 ng | 50 μg | IM/EP | +/− | + (10/10) | + (10/10) | [26] | |
| 10 μg | IV | +/− | + | |||||
| 20 μg | TD | − | + | nt | [64] | |||
| Flavi | WNV | 0.1–10 μg | IM | + | + | + | [28,30,65,66] | |
| JEV | 2 μg | nt | n/a | n/a | n/a | n/a | [29,67] | |
| YFV | 10 ng | 20 μg | IM/EP | − | + (10/10) | nt | [27] | |
| 5 μg | 0.1–10 μg | IM/EP | + | + | nt | [45] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Pushko, P.; Lukashevich, I.S.; Johnson, D.M.; Tretyakova, I. Single-Dose Immunogenic DNA Vaccines Coding for Live-Attenuated Alpha- and Flaviviruses. Viruses 2024, 16, 428. https://doi.org/10.3390/v16030428
Pushko P, Lukashevich IS, Johnson DM, Tretyakova I. Single-Dose Immunogenic DNA Vaccines Coding for Live-Attenuated Alpha- and Flaviviruses. Viruses. 2024; 16(3):428. https://doi.org/10.3390/v16030428
Chicago/Turabian StylePushko, Peter, Igor S. Lukashevich, Dylan M. Johnson, and Irina Tretyakova. 2024. "Single-Dose Immunogenic DNA Vaccines Coding for Live-Attenuated Alpha- and Flaviviruses" Viruses 16, no. 3: 428. https://doi.org/10.3390/v16030428
APA StylePushko, P., Lukashevich, I. S., Johnson, D. M., & Tretyakova, I. (2024). Single-Dose Immunogenic DNA Vaccines Coding for Live-Attenuated Alpha- and Flaviviruses. Viruses, 16(3), 428. https://doi.org/10.3390/v16030428