

The Importance of Including Non-Household Environments in Dengue Vector Control Activities

 , , and
, , and
Abstract
1. Introduction
2. Materials and Methods
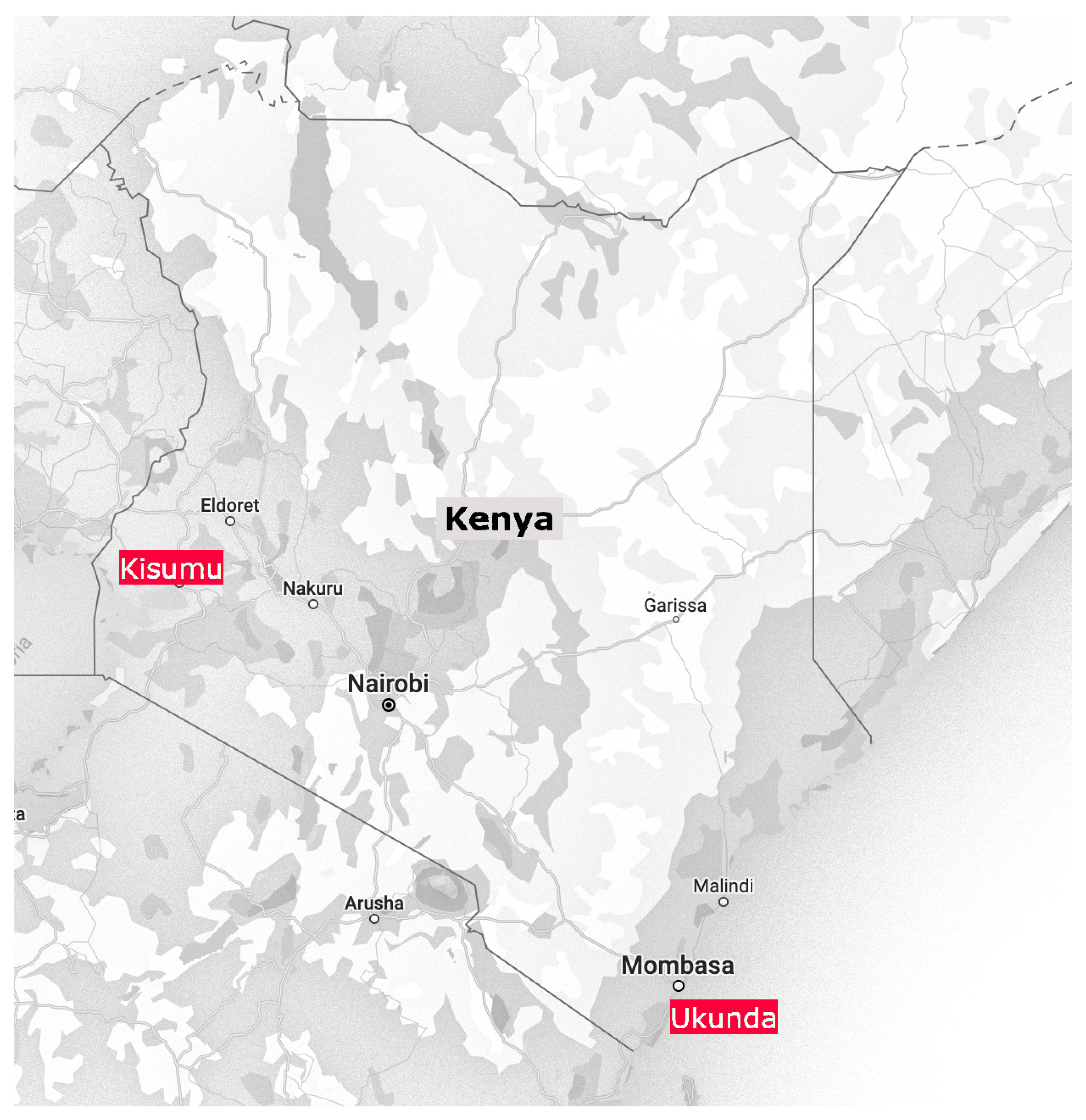
2.1. Vector Sampling and Data Collection
2.2. Statistical Analysis
2.3. Mosquito Density Simulations
3. Results
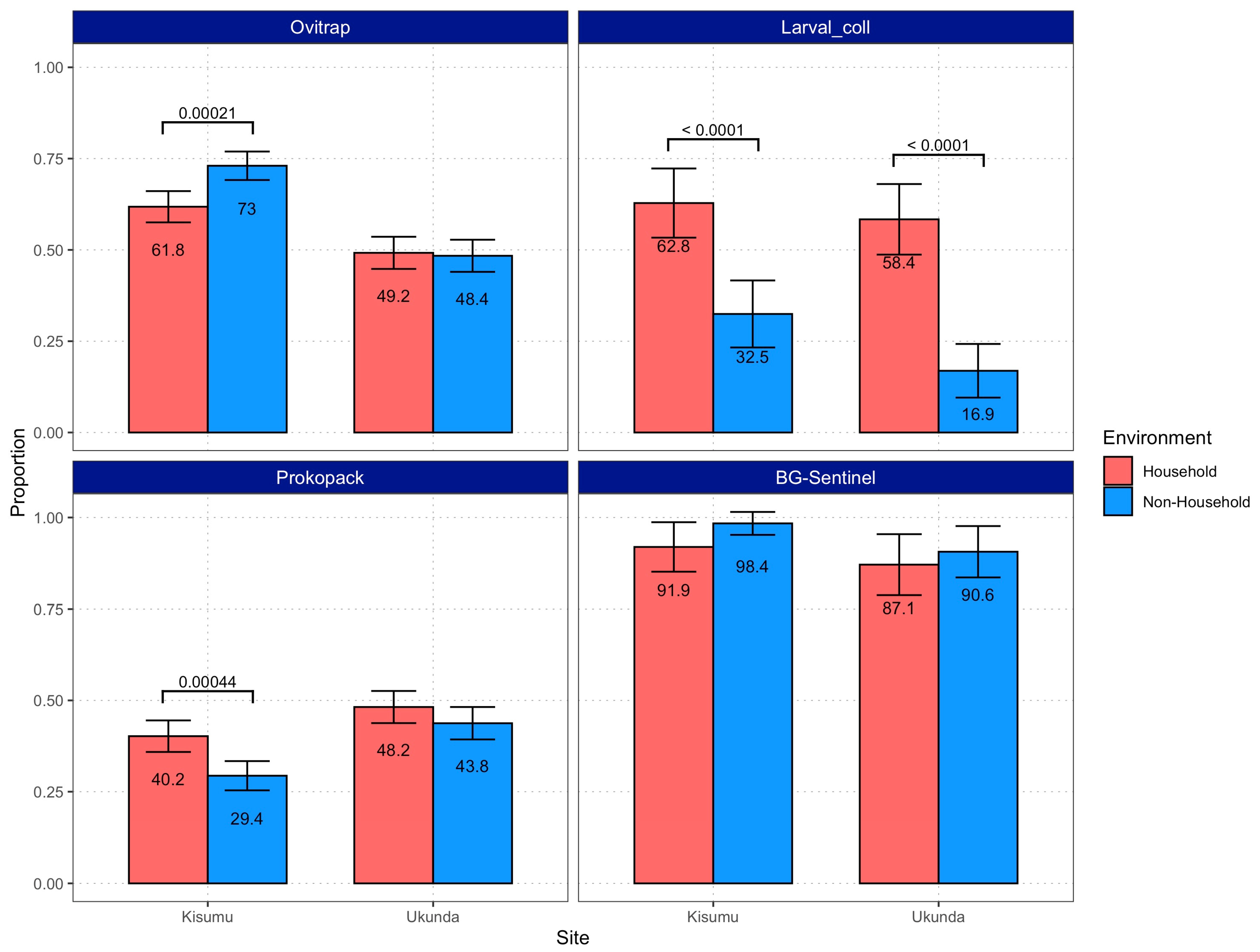
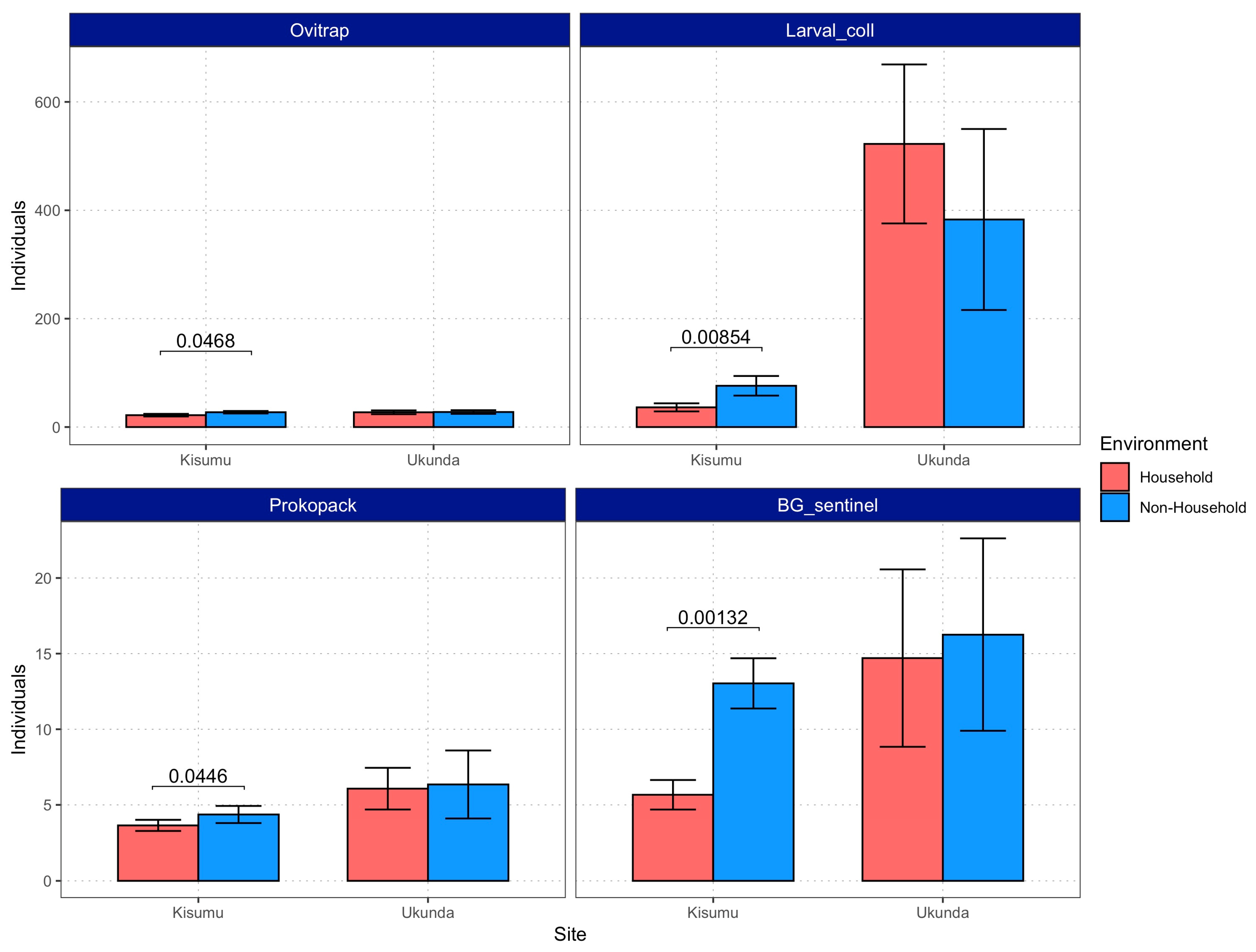
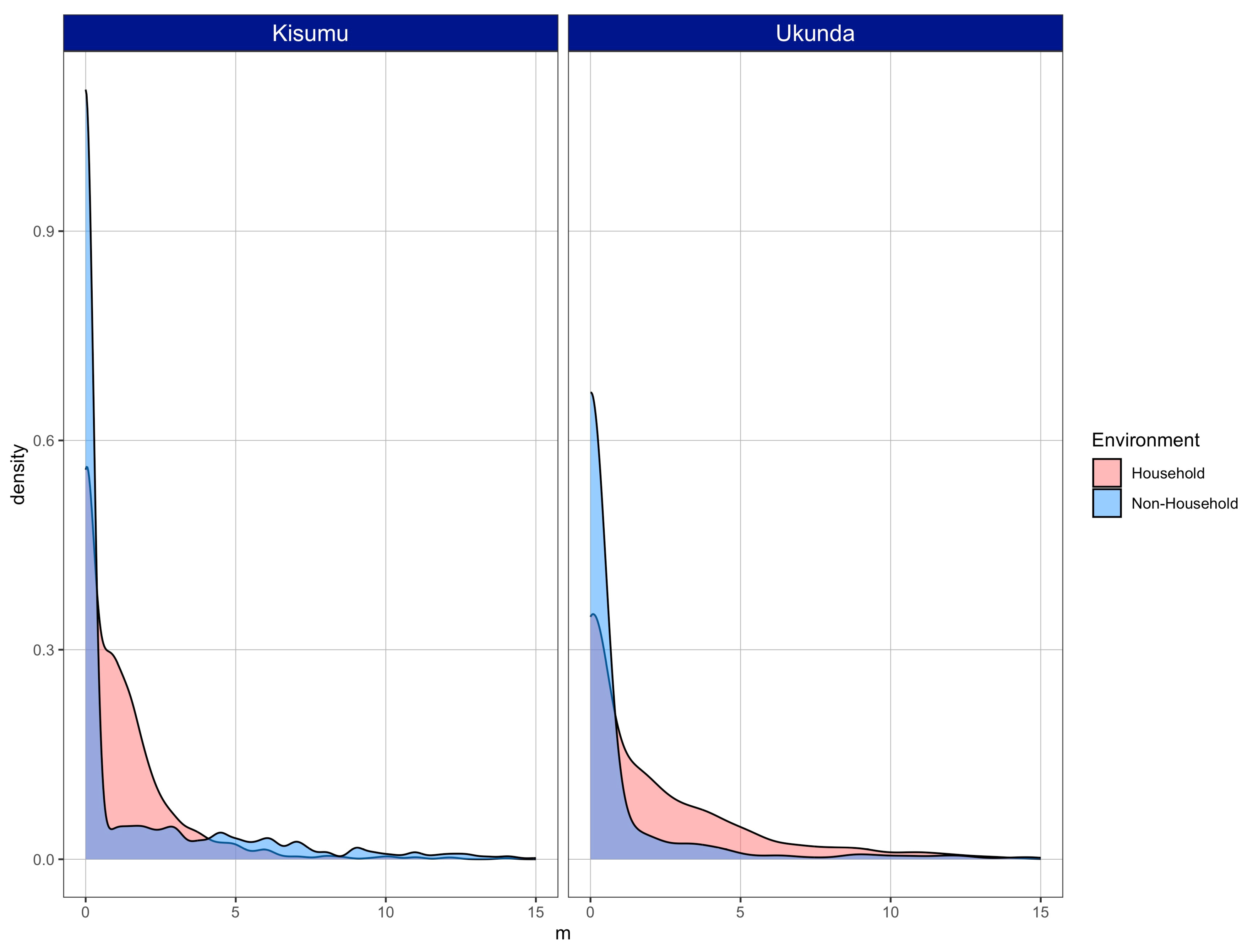
3.1. Mosquitoes Were Present and Abundant in Both Environments
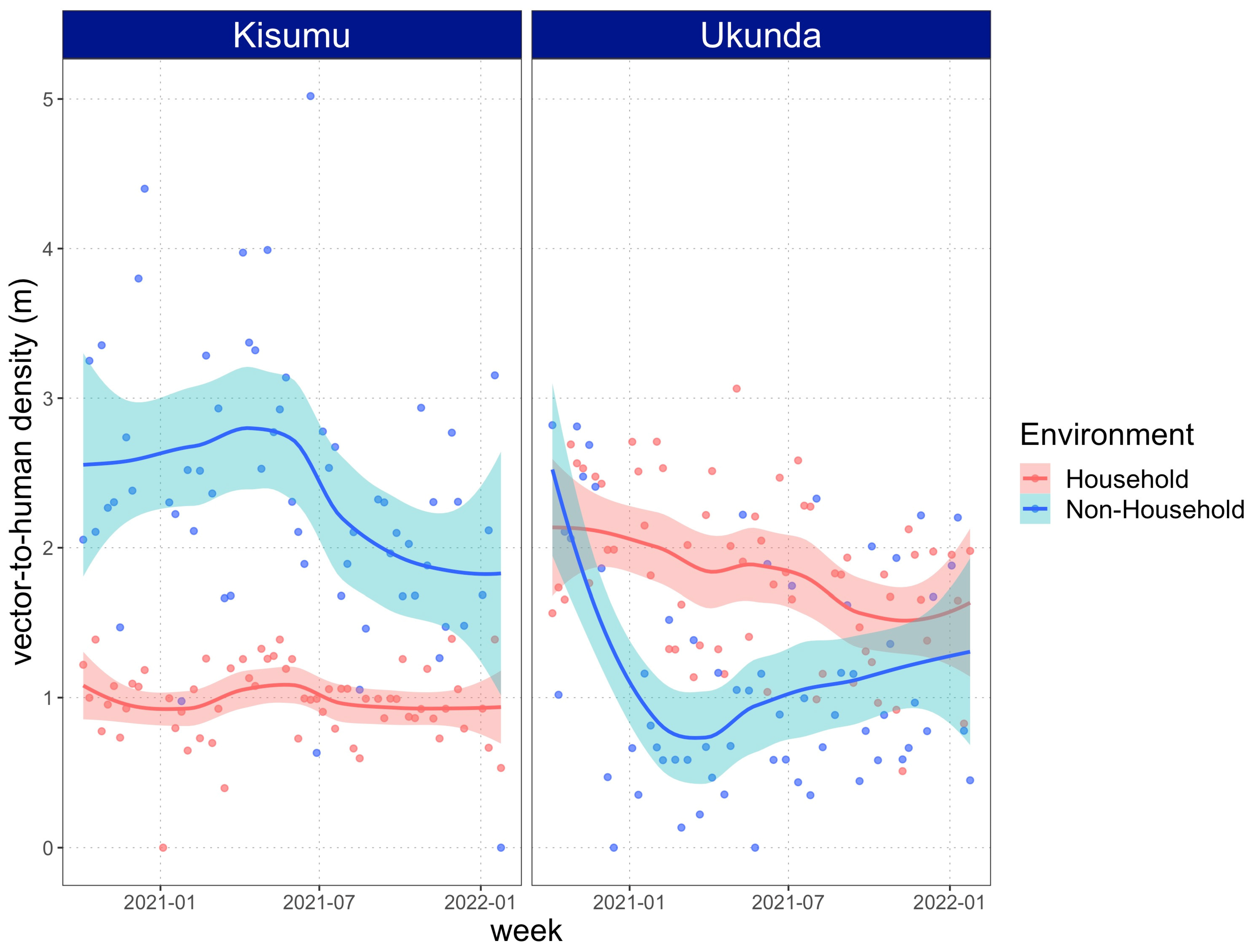
3.2. Vector Density Is Equal or Higher in Non-Household Spaces Compared to Households
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Sow, A.; Loucoubar, C.; Diallo, D.; Faye, O.; Ndiaye, Y.; Senghor, C.S.; Dia, A.T.; Faye, O.; Weaver, S.C.; Diallo, M.; et al. Concurrent malaria and arbovirus infections in Kedougou, southeastern Senegal. Malar. J. 2016, 15, 47. [Google Scholar] [CrossRef] [PubMed]
- LaBeaud, A.D.; Banda, T.; Brichard, J.; Muchiri, E.M.; Mungai, P.L.; Mutuku, F.M.; Borland, E.; Gildengorin, G.; Pfeil, S.; Teng, C.Y.; et al. High Rates of O’Nyong Nyong and Chikungunya Virus Transmission in Coastal Kenya. PLoS Negl. Trop. Dis. 2015, 9, e0003436. [Google Scholar] [CrossRef] [PubMed]
- Ali, I.M.; Tchuenkam, V.P.K.; Colton, M.; Stittleburg, V.; Mitchell, C.; Gaither, C.; Thwai, K.; Espinoza, D.O.; Zhu, Y.; Jamal, H.; et al. Arboviruses as an unappreciated cause of non-malarial acute febrile illness in the Dschang Health District of western Cameroon. PLoS Negl. Trop. Dis. 2022, 16, e0010790. [Google Scholar] [CrossRef]
- Gainor, E.M.; Harris, E.; LaBeaud, A.D. Uncovering the Burden of Dengue in Africa: Considerations on Magnitude, Misdiagnosis, and Ancestry. Viruses 2022, 14, 233. [Google Scholar] [CrossRef] [PubMed]
- Mwanyika, G.O.; Sindato, C.; Rugarabamu, S.; Rumisha, S.F.; Karimuribo, E.D.; Misinzo, G.; Rweyemamu, M.M.; Hamid, M.M.A.; Haider, N.; Vairo, F.; et al. Seroprevalence and associated risk factors of chikungunya, dengue, and Zika in eight districts in Tanzania. Int. J. Infect. Dis. 2021, 111, 271–280. [Google Scholar] [CrossRef] [PubMed]
- Konongoi, L.; Ofula, V.; Nyunja, A.; Owaka, S.; Koka, H.; Makio, A.; Koskei, E.; Eyase, F.; Langat, D.; Schoepp, R.J.; et al. Detection of dengue virus serotypes 1, 2 and 3 in selected regions of Kenya: 2011–2014. Virol. J. 2016, 13, 182. [Google Scholar] [CrossRef] [PubMed]
- Shah, M.M.; Ndenga, B.A.; Mutuku, F.M.; Vu, D.M.; Grossi-Soyster, E.N.; Okuta, V.; Ronga, C.O.; Chebii, P.K.; Maina, P.; Jembe, Z.; et al. High Dengue Burden and Circulation of 4 Virus Serotypes among Children with Undifferentiated Fever, Kenya, 2014–2017. Emerg. Infect. Dis. 2020, 26, 2638–2650. [Google Scholar] [CrossRef] [PubMed]
- Grossi-Soyster, E.N.; Cook, E.A.J.; de Glanville, W.A.; Thomas, L.F.; Krystosik, A.R.; Lee, J.; Wamae, C.N.; Kariuki, S.; Fèvre, E.M.; LaBeaud, A.D. Serological and spatial analysis of alphavirus and flavivirus prevalence and risk factors in a rural community in western Kenya. PLoS Negl. Trop. Dis. 2017, 11, e0005998. [Google Scholar] [CrossRef]
- Vu, D.M.; Banda, T.; Teng, C.Y.; Heimbaugh, C.; Muchiri, E.M.; Mungai, P.L.; Mutuku, F.M.; Brichard, J.; Gildengorin, G.; Borland, E.M.; et al. Dengue and West Nile Virus Transmission in Children and Adults in Coastal Kenya. Am. J. Trop. Med. Hyg. 2017, 96, 141–143. [Google Scholar] [CrossRef]
- Vu, D.M.; Mutai, N.; Heath, C.J.; Vulule, J.M.; Mutuku, F.M.; Ndenga, B.A.; LaBeaud, A.D. Unrecognized Dengue Virus Infections in Children, Western Kenya, 2014–2015. Emerg. Infect. Dis. 2017, 23, 1915–1917. [Google Scholar] [CrossRef]
- Hortion, J.; Mutuku, F.M.; Eyherabide, A.L.; Vu, D.M.; Boothroyd, D.B.; Grossi-Soyster, E.N.; King, C.H.; Ndenga, B.A.; LaBeaud, A.D. Acute Flavivirus and Alphavirus Infections among Children in Two Different Areas of Kenya, 2015. Am. J. Trop. Med. Hyg. 2019, 100, 170–173. [Google Scholar] [CrossRef]
- Sutherland, L.J.; Sang, R.C.; King, C.H.; LaBeaud, A.D.; Cash, A.A.; King, C.L.; Weaver, S.C.; Malhotra, I.; Huang, Y.-J.S.; Moormann, A.M. Serologic Evidence of Arboviral Infections among Humans in Kenya. Am. J. Trop. Med. Hyg. 2011, 85, 158–161. [Google Scholar] [CrossRef] [PubMed]
- Ngugi, H.N.; Mutuku, F.M.; Ndenga, B.A.; Musunzaji, P.S.; Mbakaya, J.O.; Aswani, P.; Irungu, L.W.; Mukoko, D.; Vulule, J.; Kitron, U.; et al. Characterization and productivity profiles of Aedes aegypti (L.) breeding habitats across rural and urban landscapes in western and coastal Kenya. Parasites Vectors 2017, 10, 331. [Google Scholar] [CrossRef] [PubMed]
- Ndenga, B.A.; Mutuku, F.M.; Ngugi, H.N.; Mbakaya, J.O.; Aswani, P.; Musunzaji, P.S.; Vulule, J.; Mukoko, D.; Kitron, U.; LaBeaud, A.D. Characteristics of Aedes aegypti adult mosquitoes in rural and urban areas of western and coastal Kenya. PLoS ONE 2017, 12, e0189971. [Google Scholar] [CrossRef] [PubMed]
- Ndenga, B.A.; Mutuku, F.M.; Ngugi, H.N.; Mbakaya, J.O.; Mukoko, D.; Kitron, U.; LaBeaud, A.D. Night Time Extension of Aedes aegypti Human Blood Seeking Activity. Am. J. Trop. Med. Hyg. 2022, 107, 208–210. [Google Scholar] [CrossRef]
- Scott, T.W.; Amerasinghe, P.H.; Morrison, A.C.; Lorenz, L.H.; Clark, G.G.; Strickman, D.; Kittayapong, P.; Edman, J.D. Longitudinal studies of Aedes aegypti (Diptera: Culicidae) in Thailand and Puerto Rico: Blood feeding frequency. J. Med. Entomol. 2000, 37, 89–101. [Google Scholar] [CrossRef]
- Scott, T.W.; Chow, E.; Strickman, D.; Kittayapong, P.; Wirtz, R.A.; Lorenz, L.H.; Edman, J.D. Blood-feeding patterns of Aedes aegypti (Diptera: Culicidae) collected in a rural Thai village. J. Med. Entomol. 1993, 30, 922–927. [Google Scholar] [CrossRef]
- Roiz, D.; Wilson, A.L.; Scott, T.W.; Fonseca, D.M.; Jourdain, F.; Müller, P.; Velayudhan, R.; Corbel, V. Integrated Aedes management for the control of Aedes-borne diseases. PLoS Negl. Trop. Dis. 2018, 12, e0006845. [Google Scholar] [CrossRef]
- WHO. Vector Control Operations Framework for Zika Virus; World Health Organization: Geneva, Switzerland, 2016. [Google Scholar]
- Christophers, S.R. Aedes aegypti (L.), the Yellow Fever Mosquito; Its Life History, Bionomics, and Structure; University Press: Cambridge, MA, USA, 1960; p. 738. [Google Scholar]
- Trpis, M.; McClelland, G.A.; Gillett, J.D.; Teesdale, C.; Rao, T.R. Diel periodicity in the landing of Aedes aegypti on man. Bull. World Health Organ. 1973, 48, 623–629. [Google Scholar]
- McBride, C.S.; Baier, F.; Omondi, A.B.; Spitzer, S.A.; Lutomiah, J.; Sang, R.; Ignell, R.; Vosshall, L.B. Evolution of mosquito preference for humans linked to an odorant receptor. Nature 2014, 515, 222–227. [Google Scholar] [CrossRef]
- Huang, C.H.; Lin, C.Y.; Yang, C.Y.; Chan, T.C.; Chiang, P.H.; Chen, Y.H. Relationship between the Incidence of Dengue Virus Transmission in Traditional Market and Climatic Conditions in Kaohsiung City. Can. J. Infect. Dis. Med. Microbiol. 2021, 2021, 9916642. [Google Scholar] [CrossRef]
- Suwanbamrung, C.; Promsupa, S.; Doungsin, T.; Tongjan, S. Risk factors related to dengue infections in primary school students: Exploring students’ basic knowledge of dengue and examining the larval indices in southern Thailand. J. Infect. Public Health 2013, 6, 347–357. [Google Scholar] [CrossRef] [PubMed]
- Pérez-Pérez, J.; Peña-García, V.H.; Calle-Tobón, A.; Quimbayo-Forero, M.; Rojo, R.; Henao, E.; Shragai, T.; Rúa-Uribe, G. Entomovirological Surveillance in Schools: Are They a Source for Arboviral Diseases Transmission? Int. J. Environ. Res. Public Health 2021, 18, 6137. [Google Scholar] [CrossRef]
- Kampango, A.; Furu, P.; Sarath, D.L.; Haji, K.A.; Konradsen, F.; Schiøler, K.L.; Alifrangis, M.; Saleh, F.; Weldon, C.W. Risk factors for occurrence and abundance of Aedes aegypti and Aedes bromeliae at hotel compounds in Zanzibar. Parasit Vectors 2021, 14, 544. [Google Scholar] [CrossRef] [PubMed]
- Barrera, R.; Acevedo, V.; Amador, M. Role of Abandoned and Vacant Houses on Aedes aegypti Productivity. Am. J. Trop. Med. Hyg. 2021, 104, 145–150. [Google Scholar] [CrossRef] [PubMed]
- Troyo, A.; Calderón-Arguedas, O.; Fuller, D.O.; Solano, M.E.; Avendaño, A.; Arheart, K.L.; Chadee, D.D.; Beier, J.C. Seasonal profiles of Aedes aegypti (Diptera: Culicidae) larval habitats in an urban area of Costa Rica with a history of mosquito control. J. Vector Ecol. 2008, 33, 76–88. [Google Scholar] [CrossRef]
- Macdonald, G. The Epidemiology and Control of Malaria; Oxford University Press: London, UK, 1957; p. 201. [Google Scholar]
- Smith, D.L.; Perkins, T.A.; Reiner, R.C.; Barker, C.M.; Niu, T.; Chaves, L.F.; Ellis, A.M.; George, D.B.; Le Menach, A.; Pulliam, J.R.; et al. Recasting the theory of mosquito-borne pathogen transmission dynamics and control. Trans. R. Soc. Trop. Med. Hyg. 2014, 108, 185–197. [Google Scholar] [CrossRef]
- Kramer, L.D.; Ciota, A.T. Dissecting vectorial capacity for mosquito-borne viruses. Curr. Opin. Virol. 2015, 15, 112–118. [Google Scholar] [CrossRef]
- Garrett-Jones, C. Prognosis for interruption of malaria transmission through assessment of the mosquito’s vectorial capacity. Nature 1964, 204, 1173–1175. [Google Scholar] [CrossRef]
- Forsyth, J.E.; Mutuku, F.M.; Kibe, L.; Mwashee, L.; Bongo, J.; Egemba, C.; Ardoin, N.M.; LaBeaud, A.D. Source reduction with a purpose: Mosquito ecology and community perspectives offer insights for improving household mosquito management in coastal Kenya. PLoS Negl. Trop. Dis. 2020, 14, e0008239. [Google Scholar] [CrossRef]
- Huang, Y.-M. The subgenus Stegomyia of Aedes in the Afrotropical Region with keys to the species (Diptera: Culicidae). Zootaxa 2004, 700, 1–120. [Google Scholar] [CrossRef]
- Gillett, J.D. Common African Mosquitoes and Their Medical Importance; William Heinemann Medical Books Ltd.: London, UK, 1972; p. 106. [Google Scholar]
- Bhatt, S.; Gething, P.W.; Brady, O.J.; Messina, J.P.; Farlow, A.W.; Moyes, C.L.; Drake, J.M.; Brownstein, J.S.; Hoen, A.G.; Sankoh, O.; et al. The global distribution and burden of dengue. Nature 2013, 496, 504–507. [Google Scholar] [CrossRef] [PubMed]
- Yang, X.; Quam, M.B.M.; Zhang, T.; Sang, S. Global burden for dengue and the evolving pattern in the past 30 years. J. Travel Med. 2021, 28, taab146. [Google Scholar] [CrossRef] [PubMed]
- Labeaud, A.D.; Bashir, F.; King, C.H. Measuring the burden of arboviral diseases: The spectrum of morbidity and mortality from four prevalent infections. Popul. Health Metr. 2011, 9, 1. [Google Scholar] [CrossRef]
- Torres-Flores, J.M.; Reyes-Sandoval, A.; Salazar, M.I. Dengue Vaccines: An Update. BioDrugs 2022, 36, 325–336. [Google Scholar] [CrossRef]
- Ogunlade, S.T.; Meehan, M.T.; Adekunle, A.I.; Rojas, D.P.; Adegboye, O.A.; McBryde, E.S. A Review: Aedes-Borne Arboviral Infections, Controls and Wolbachia-Based Strategies. Vaccines 2021, 9, 32. [Google Scholar] [CrossRef]
- Wilder-Smith, A. Dengue during the COVID-19 pandemic. J. Travel Med. 2021, 28, taab183. [Google Scholar] [CrossRef]
- Sasmono, R.T.; Santoso, M.S. Movement dynamics: Reduced dengue cases during the COVID-19 pandemic. Lancet Infect. Dis. 2022, 22, 570–571. [Google Scholar] [CrossRef]
- Forsyth, J.E.; Kempinsky, A.; Pitchik, H.O.; Alberts, C.J.; Mutuku, F.M.; Kibe, L.; Ardoin, N.M.; LaBeaud, A.D. Larval source reduction with a purpose: Designing and evaluating a household- and school-based intervention in coastal Kenya. PLoS Negl. Trop. Dis. 2022, 16, e0010199. [Google Scholar] [CrossRef]
- Peña-García, V.H.; Luvall, J.C.; Christofferson, R.C. Arbovirus Transmission Predictions Are Affected by Both Temperature Data Source and Modeling Methodologies across Cities in Colombia. Microorganisms 2023, 11, 1249. [Google Scholar] [CrossRef]
- Peña-García, V.H.; Sánchez-Vargas, I.; Christofferson, R.; Black Iv, W.C.; Arboleda, S.; Triana-Chavez, O. Estimation of DENV-2 Transmission as a Function of Site-Specific Entomological Parameters from Three Cities in Colombia. Ann. Glob. Health 2019, 85, 32. [Google Scholar] [CrossRef] [PubMed]
- Harrington, L.C.; Fleisher, A.; Ruiz-Moreno, D.; Vermeylen, F.; Wa, C.V.; Poulson, R.L.; Edman, J.D.; Clark, J.M.; Jones, J.W.; Kitthawee, S.; et al. Heterogeneous feeding patterns of the dengue vector, Aedes aegypti, on individual human hosts in rural Thailand. PLoS Negl. Trop. Dis. 2014, 8, e3048. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| City | Environment | Factor | Coefficient | OR | p-Value |
|---|---|---|---|---|---|
| Kisumu | Household | Intercept | −2.28682 | 0.101589 | 1.64 × 10−09 |
| Type Drum | 1.95693 | 7.07756555 | 0.00652 | ||
| Type Pot | 2.94528 | 19.0159861 | 8.81 × 10−05 | ||
| Purpose Sanitation | 1.37002 | 3.9354294 | 0.00899 | ||
| Non-Household | Intercept | −3.493 | 0.03040951 | 0.029187 | |
| Size large | −28.16 | 5.89 × 10−13 | 0.001062 | ||
| Size medium | −25.54 | 8.09 × 10−12 | 7.06 × 10−04 | ||
| Size small | −26 | 5.11 × 10−12 | 7.22 × 10−04 | ||
| Type small plastic domestic | 3.986 | 53.8391017 | 0.015713 | ||
| Type tire | 5.812 | 334.287032 | 0.009137 | ||
| Source rain | 26.76 | 4.18 × 1011 | 4.47 × 10−04 | ||
| Ukunda | Household | Intercept | −2.508 | 0.08143094 | 0.002533 |
| Type small domestic | −4.864 | 0.00771954 | 0.003098 | ||
| Type animal feeding | −4.18 | 0.01529851 | 0.020602 | ||
| Type Bucket | −8.607 | 1.83 × 10−04 | 4.87 × 10−04 | ||
| Type Drum | −3.965 | 0.01896804 | 0.048362 | ||
| Type jerrycan | −5.016 | 0.006631 | 0.013445 | ||
| Purpose animal | 3.588 | 36.1616802 | 0.011661 | ||
| Non-Household | None | NA | NA | NA |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Peña-García, V.H.; Mutuku, F.M.; Ndenga, B.A.; Mbakaya, J.O.; Ndire, S.O.; Agola, G.A.; Mutuku, P.S.; Malumbo, S.L.; Ng’ang’a, C.M.; Andrews, J.R.; et al. The Importance of Including Non-Household Environments in Dengue Vector Control Activities. Viruses 2023, 15, 1550. https://doi.org/10.3390/v15071550
Peña-García VH, Mutuku FM, Ndenga BA, Mbakaya JO, Ndire SO, Agola GA, Mutuku PS, Malumbo SL, Ng’ang’a CM, Andrews JR, et al. The Importance of Including Non-Household Environments in Dengue Vector Control Activities. Viruses. 2023; 15(7):1550. https://doi.org/10.3390/v15071550
Chicago/Turabian StylePeña-García, Víctor Hugo, Francis M. Mutuku, Bryson A. Ndenga, Joel Omari Mbakaya, Samwuel Otieno Ndire, Gladys Adhiambo Agola, Paul S. Mutuku, Said L. Malumbo, Charles M. Ng’ang’a, Jason R. Andrews, and et al. 2023. "The Importance of Including Non-Household Environments in Dengue Vector Control Activities" Viruses 15, no. 7: 1550. https://doi.org/10.3390/v15071550
APA StylePeña-García, V. H., Mutuku, F. M., Ndenga, B. A., Mbakaya, J. O., Ndire, S. O., Agola, G. A., Mutuku, P. S., Malumbo, S. L., Ng’ang’a, C. M., Andrews, J. R., Mordecai, E. A., & LaBeaud, A. D. (2023). The Importance of Including Non-Household Environments in Dengue Vector Control Activities. Viruses, 15(7), 1550. https://doi.org/10.3390/v15071550