
Antiviral Effects of Adipose Tissue-Derived Mesenchymal Stem Cells Secretome against Feline Calicivirus and Feline Herpesvirus Type 1


Abstract
1. Introduction
2. Materials and Methods
2.1. Isolation and Expansion of Feline AD-MSCs
2.2. Characterization of Feline AD-MSCs
2.3. Production of fADSC-CM
2.4. Cells and Viruses
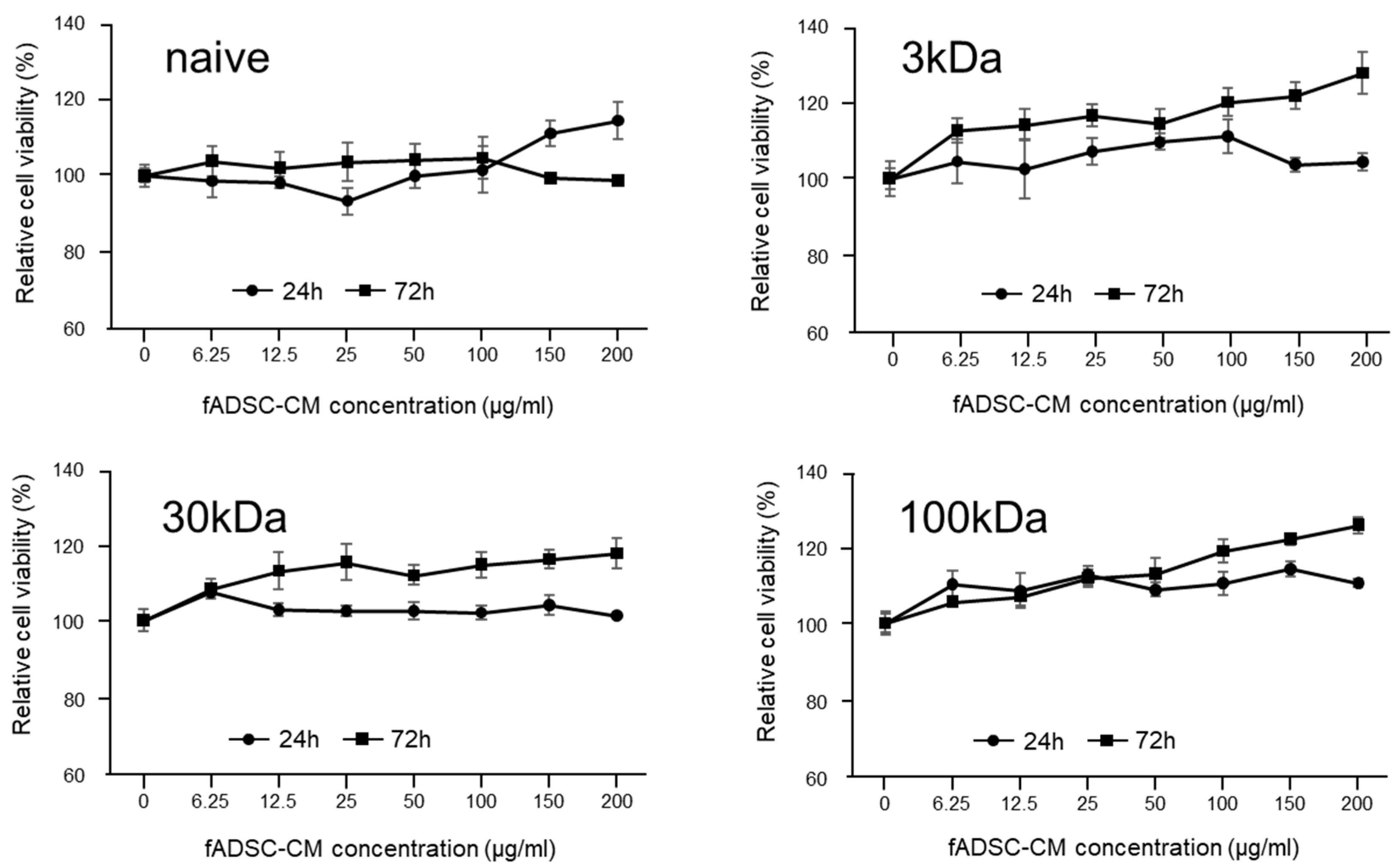
2.5. Cytotoxicity Assay for fADSC-CM
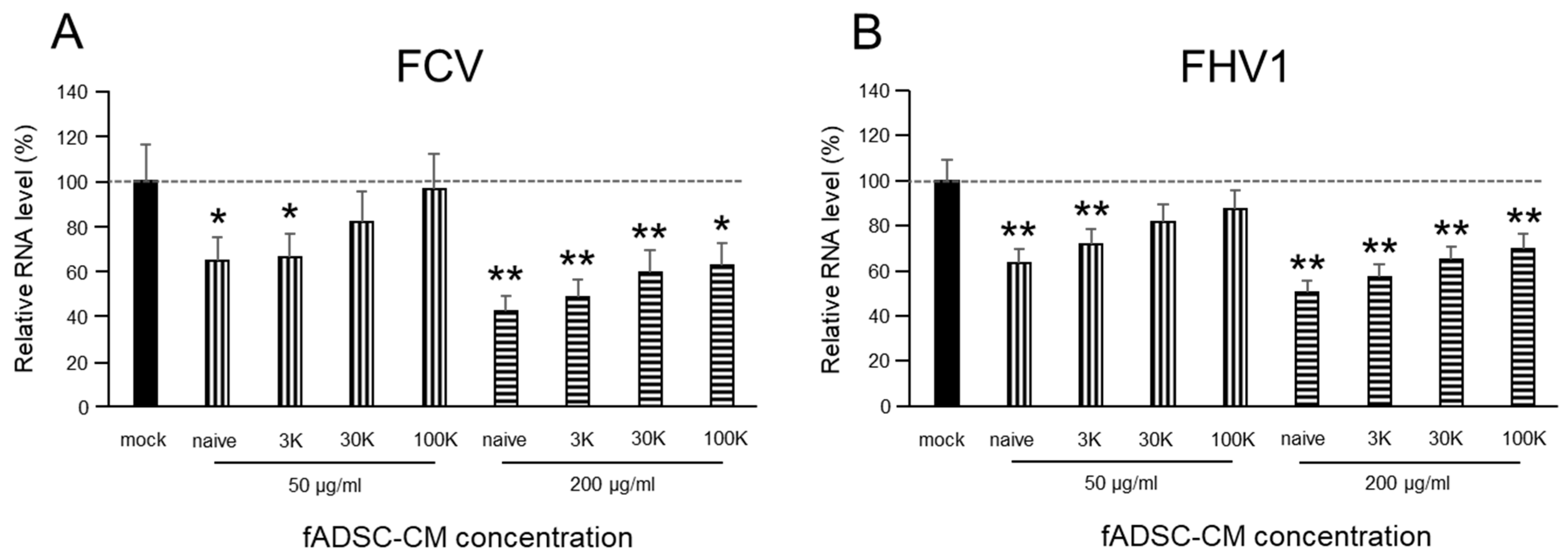
2.6. Effect of fADSC-CM on Viral Replication
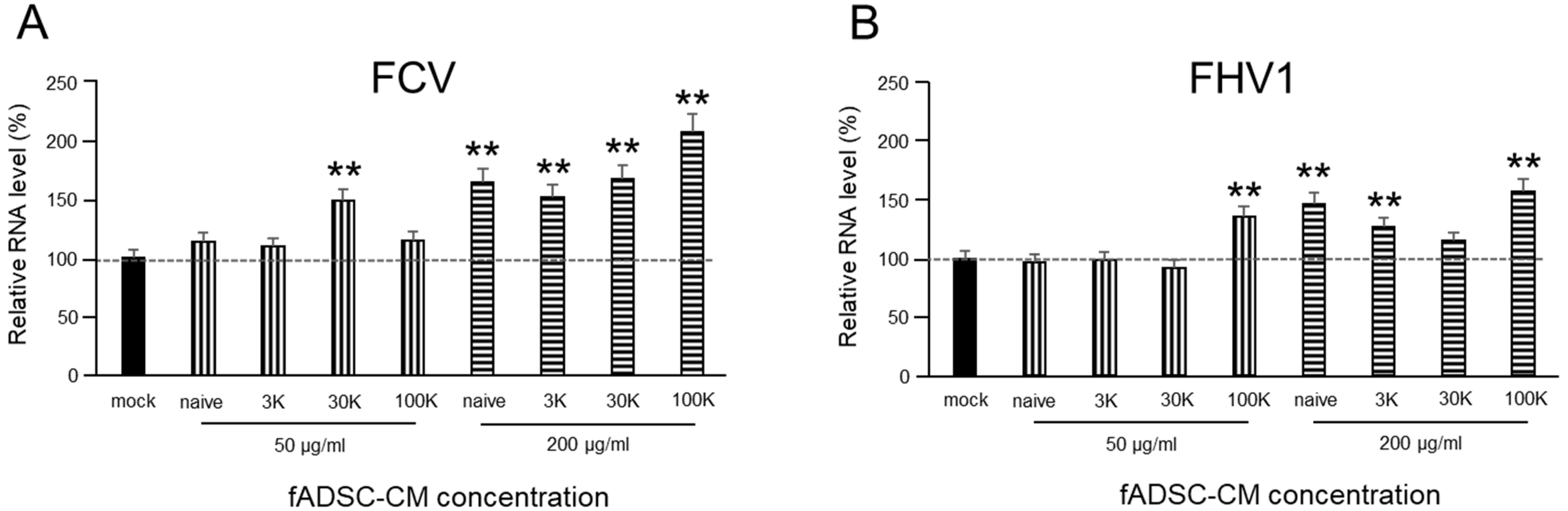
2.7. Effect of fADSC-CM on Viral Entry
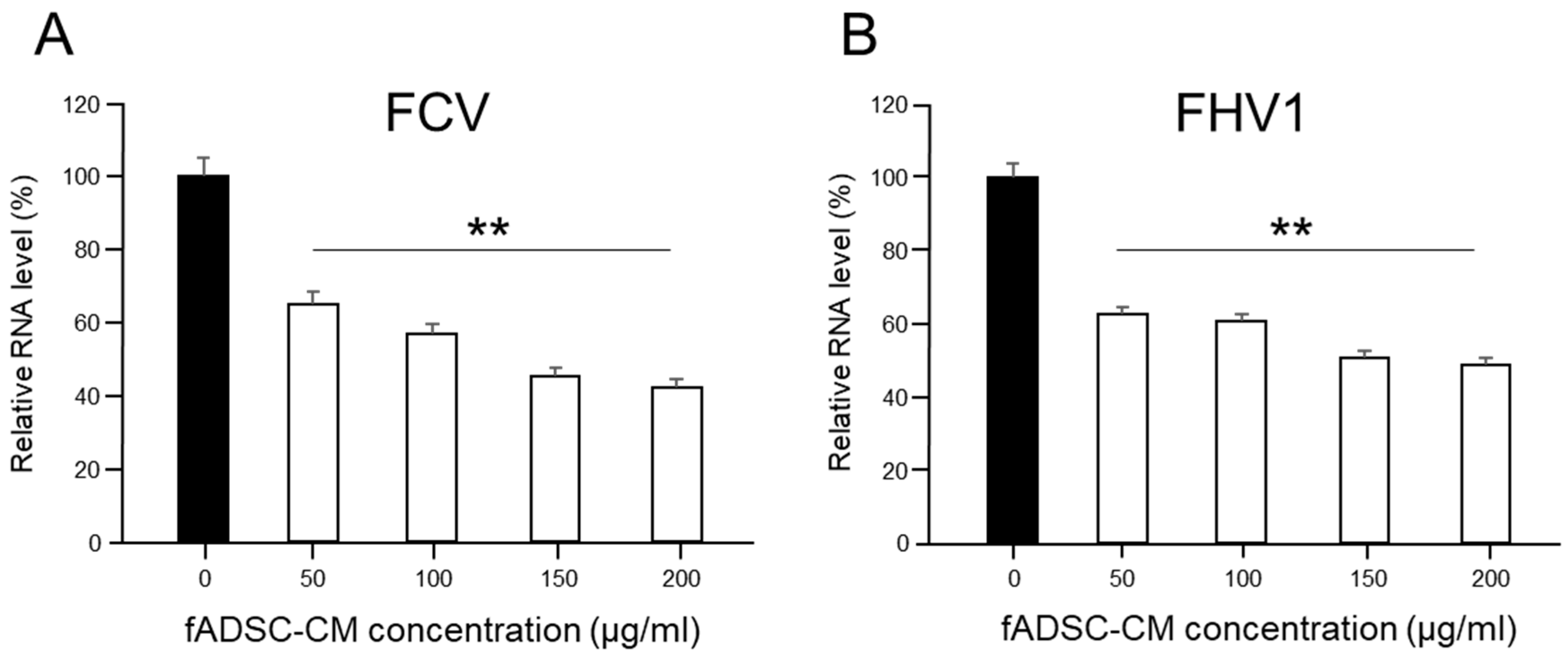
2.8. Effects of Different Amounts of fADSC-CM on Viral Replication
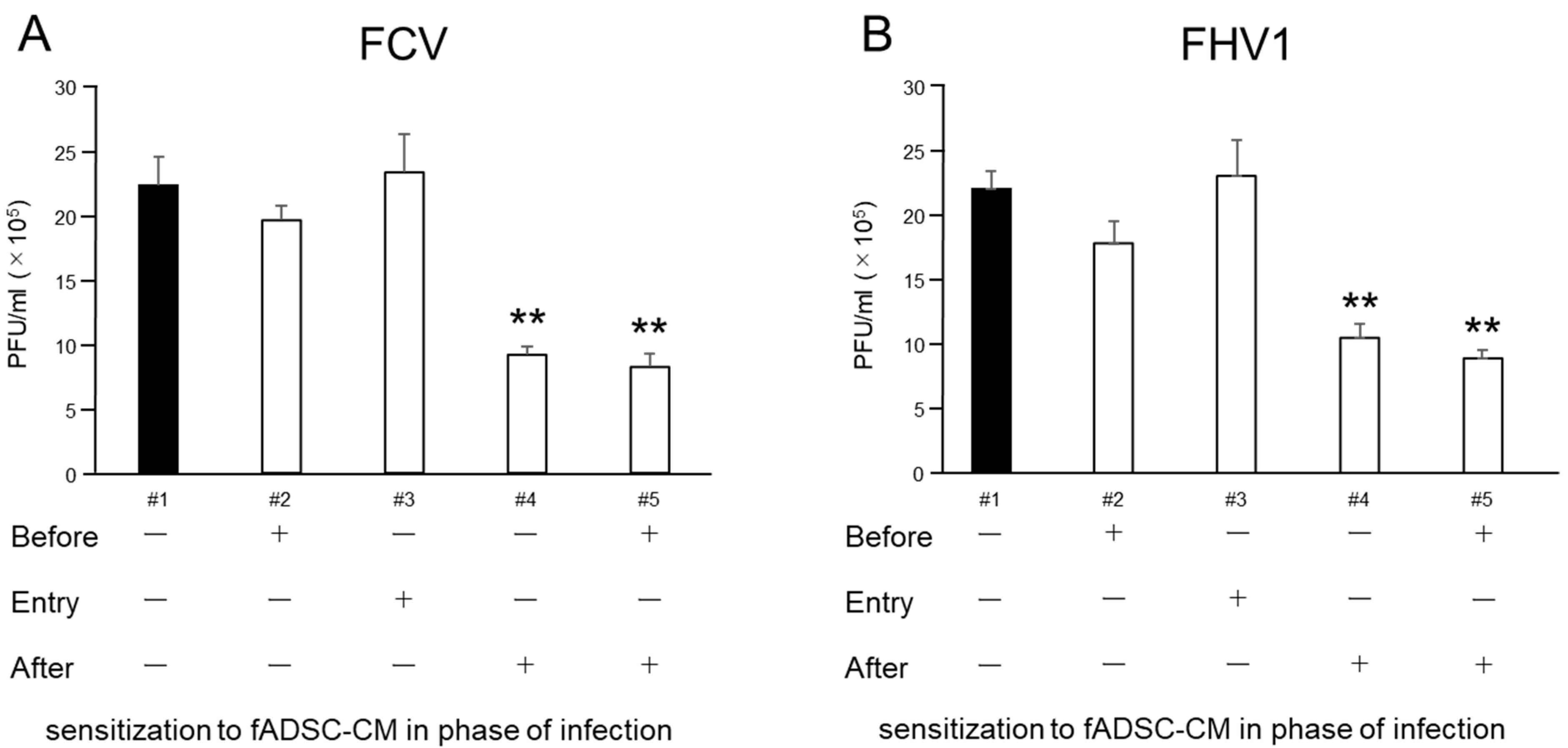
2.9. Effect of fADSC-CM on Different Phases of CRFK Cells under Infection Conditions
2.10. Plaque Assay
2.11. Real-Time RT-PCR
2.12. RNA Sequencing
2.13. Statistical Analysis
3. Results
3.1. Characterization of Feline AD-MSCs
3.2. Cytotoxicity Assay of CRFK Cells in fADSC-CM
3.3. fADSC-CM Concentrated by MWCO Ultrafiltration Inhibits Virus Replication
3.4. fADSC-CM Does Not Prevent FCV and FHV1 Entry
3.5. Inhibition of Viral Replication Depends on Amount of Naïve fADSC-CM
3.6. Naïve fAD-MSC-CM Treatment Inhibits Viral Replication of Infected CRFK Cells
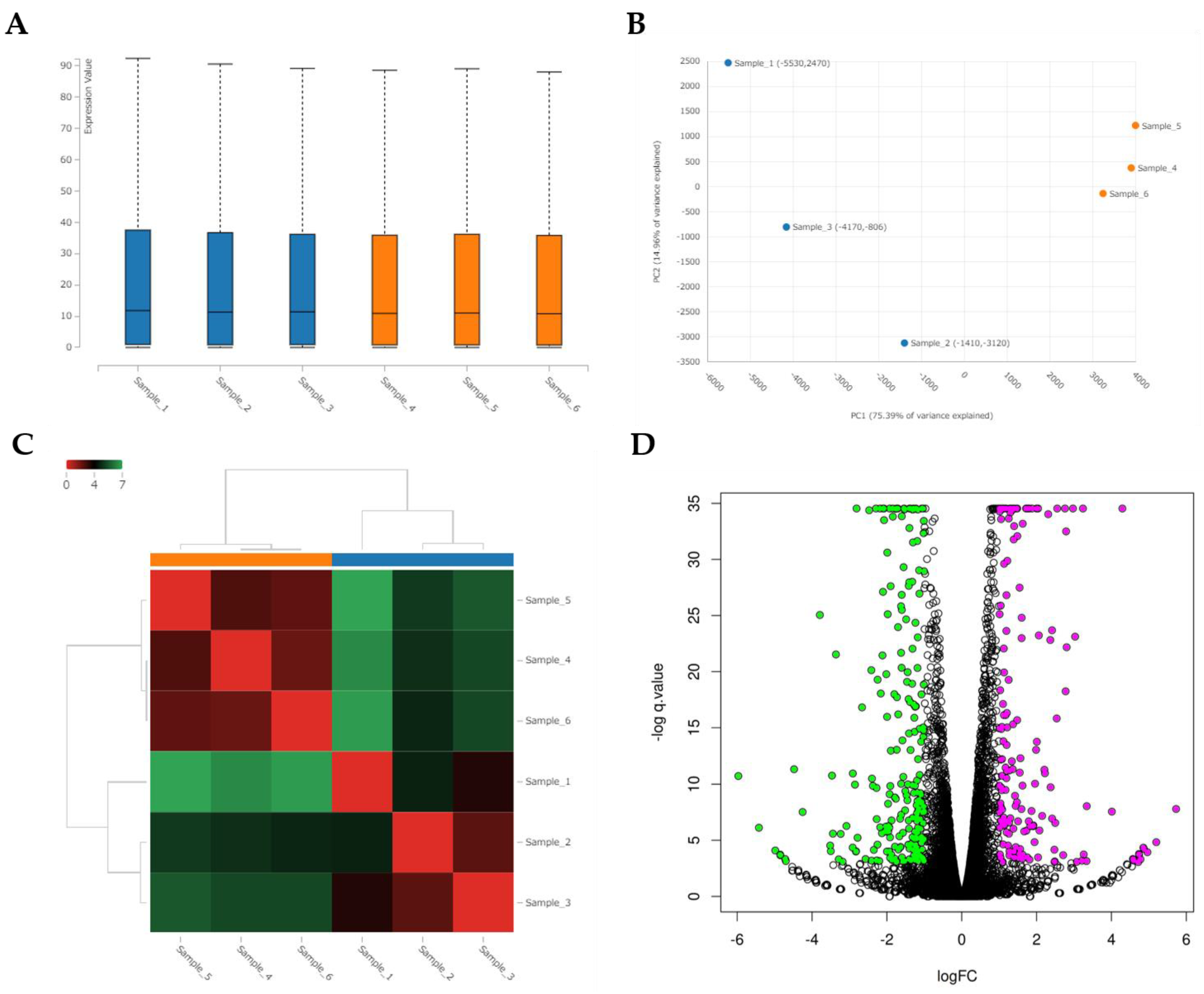
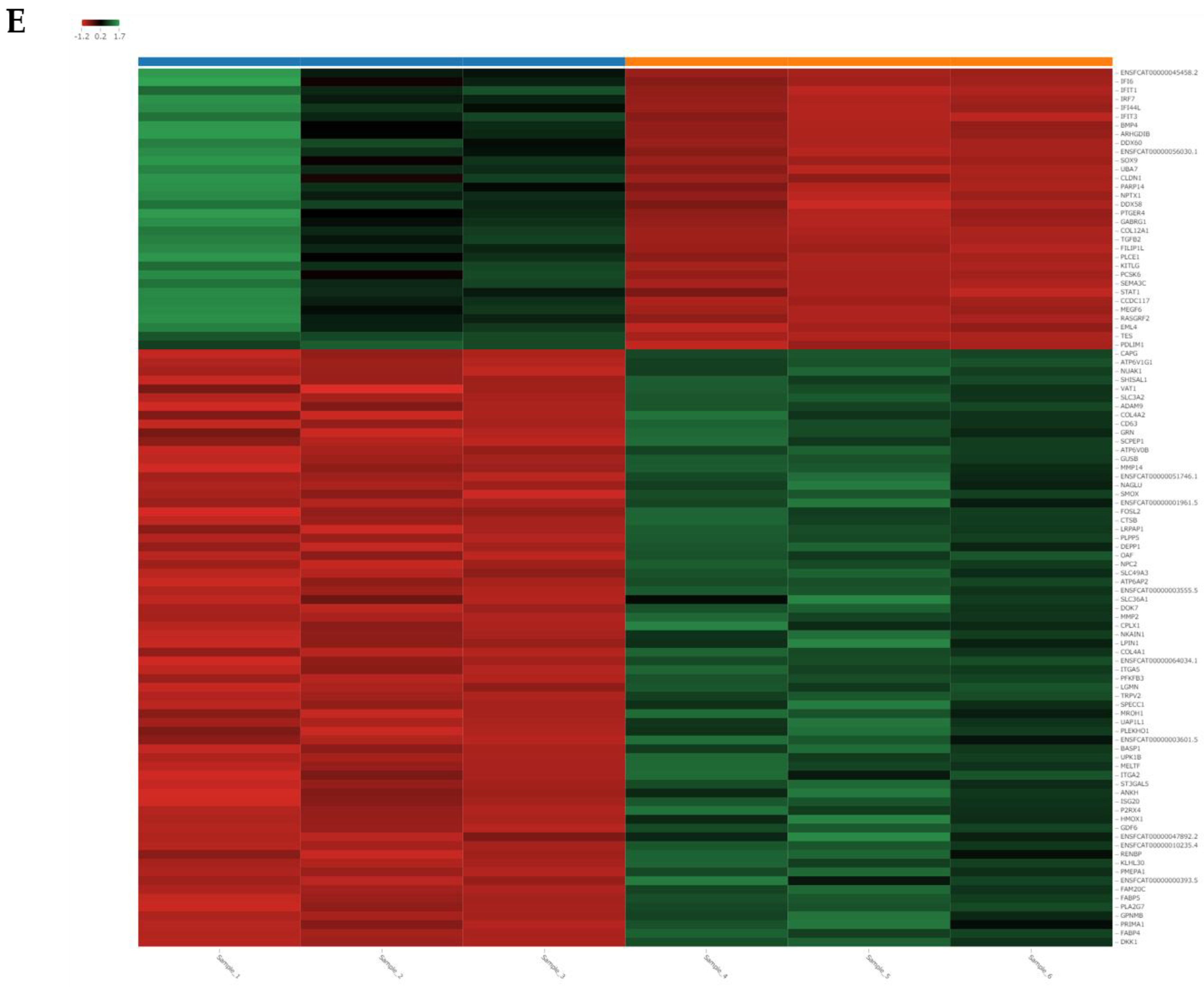
3.7. RNA Sequencing
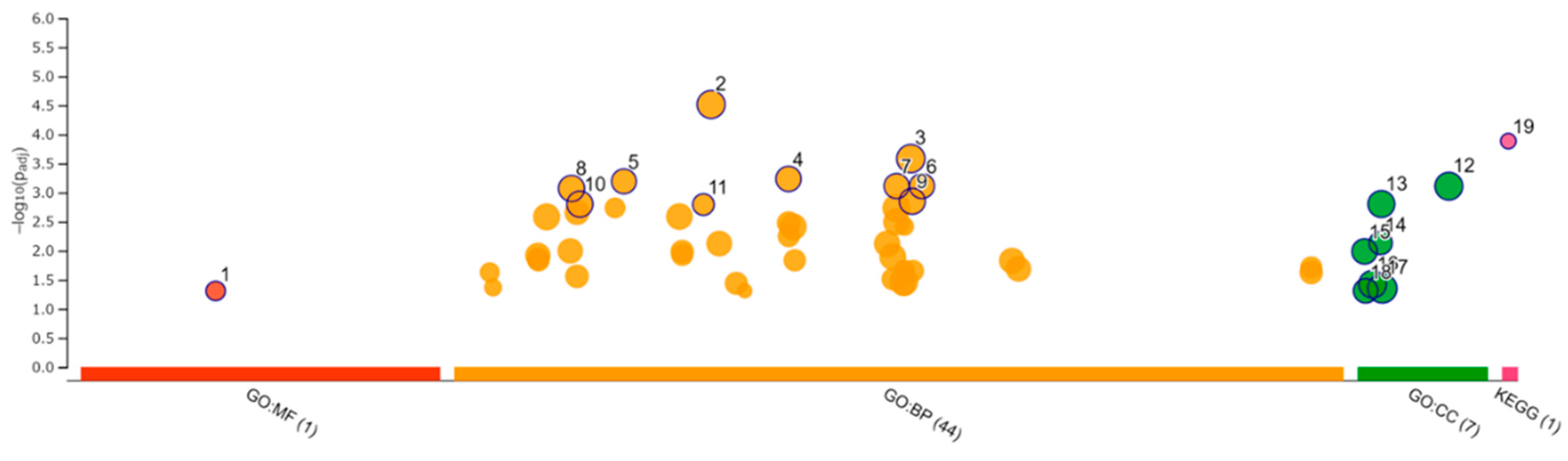
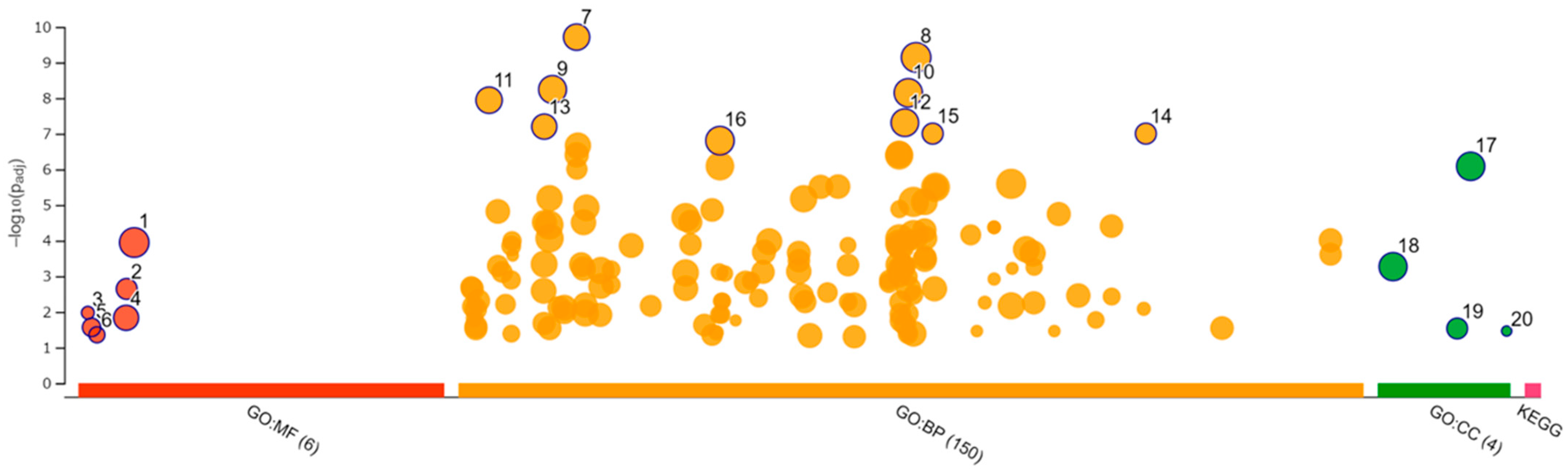
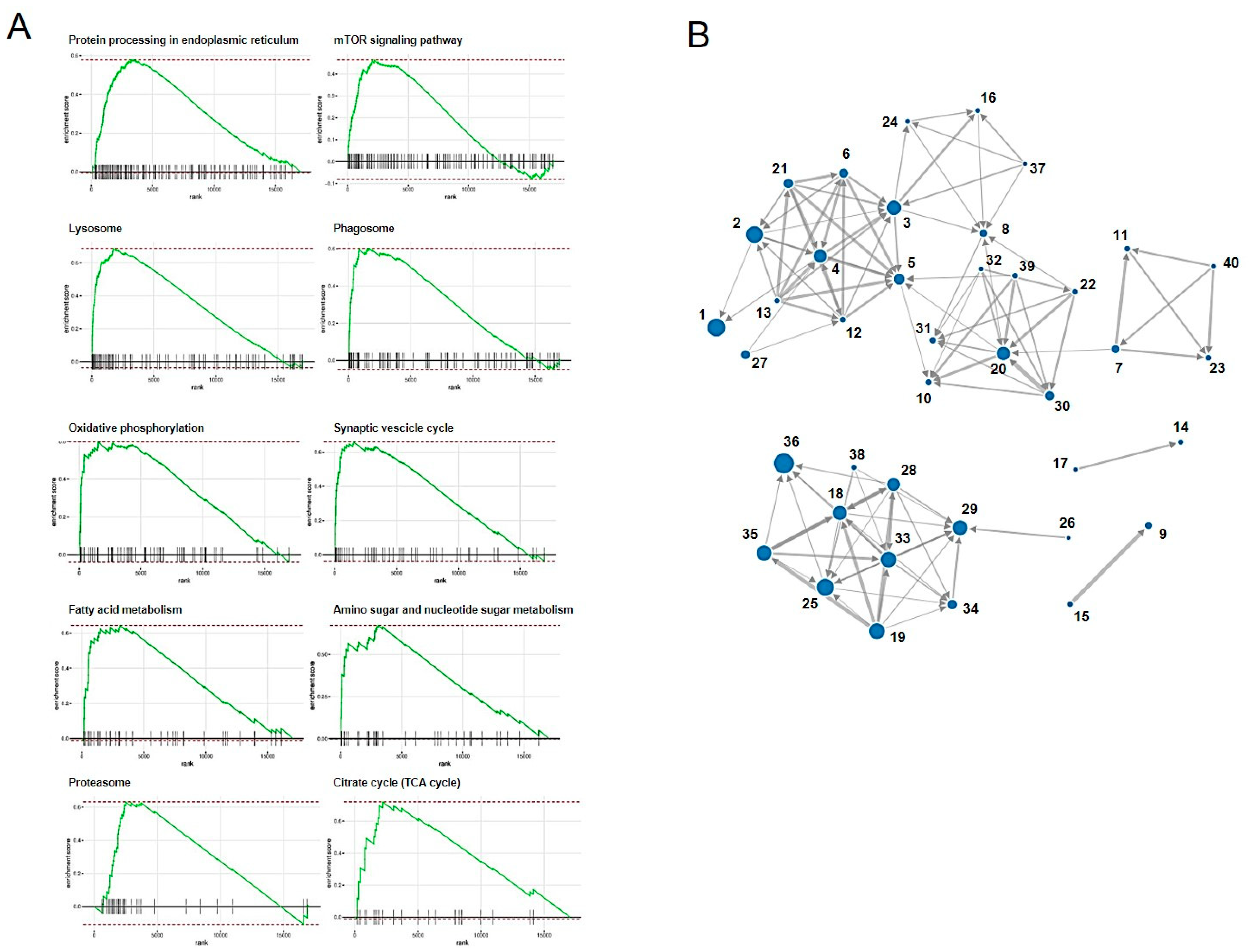
3.8. Enrichment Analysis
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Abdelrazik, H.; Giordano, E.; Barbanti Brodano, G.; Griffoni, C.; De Falco, E.; Pelagalli, A. Substantial Overview on Mesenchymal Stem Cell Biological and Physical Properties as an Opportunity in Translational Medicine. Int. J. Mol. Sci. 2019, 20, 5386. [Google Scholar] [CrossRef] [PubMed]
- Pittenger, M.F.; Discher, D.E.; Péault, B.M.; Phinney, D.G.; Hare, J.M.; Caplan, A.I. Mesenchymal stem cell perspective: Cell biology to clinical progress. NPJ Regen. Med. 2019, 4, 22. [Google Scholar] [CrossRef] [PubMed]
- D’souza, N.; Rossignoli, F.; Golinelli, G.; Grisendi, G.; Spano, C.; Candini, O.; Osturu, S.; Catani, F.; Paolucci, P.; Horwitz, E.M.; et al. Mesenchymal stem/stromal cells as a delivery platform in cell and gene therapies. BMC Med. 2015, 13, 186. [Google Scholar] [CrossRef] [PubMed]
- Zaher, W.; Harkness, L.; Jafari, A.; Kassem, M. An update of human mesenchymal stem cell biology and their clinical uses. Arch. Toxicol. 2014, 88, 1069–1082. [Google Scholar] [CrossRef] [PubMed]
- Dias, I.E.; Pinto, P.O.; Barros, L.C.; Viegas, C.A.; Dias, I.R.; Carvalho, P.P. Mesenchymal stem cells therapy in companion animals: Useful for immune-mediated diseases? BMC Vet. Res. 2019, 15, 358. [Google Scholar] [CrossRef]
- Vizoso, F.J.; Eiro, N.; Cid, S.; Schneider, J.; Perez-Fernandez, R. Mesenchymal Stem Cell Secretome: Toward Cell-Free Therapeutic Strategies in Regenerative Medicine. Int. J. Mol. Sci. 2017, 18, 1852. [Google Scholar] [CrossRef]
- Nikfarjam, S.; Rezaie, J.; Zolbanin, N.M.; Jafari, R. Mesenchymal stem cell derived-exosomes: A modern approach in translational medicine. J. Transl. Med. 2020, 18, 449. [Google Scholar] [CrossRef]
- Van de Vyver, M.; Powrie, Y.S.L.; Smith, C. Targeting Stem Cells in Chronic Inflammatory Diseases. Adv. Exp. Med. Biol. 2021, 1286, 163–181. [Google Scholar]
- Taechangam, N.; Kol, A.; Arzi, B.; Borjesson, D.L. Multipotent Stromal Cells and Viral Interaction: Current Implications for Therapy. Stem Cell Rev. Rep. 2022, 18, 214–227. [Google Scholar] [CrossRef] [PubMed]
- Radford, A.D.; Coyne, K.P.; Dawson, S.; Porter, C.J.; Gaskell, R.M. Feline calicivirus. Vet. Res. 2007, 38, 319–335. [Google Scholar] [CrossRef]
- Gaskell, R.; Dawson, S.; Radford, A.; Thiry, E. Feline herpesvirus. Vet. Res. 2007, 38, 337–354. [Google Scholar] [CrossRef] [PubMed]
- Bergmann, M.; Speck, S.; Rieger, A.; Truyen, U.; Hartmann, K. Antibody response to feline herpesvirus-1 vaccination in healthy adult cats. J. Feline Med. Surg. 2020, 22, 329–338. [Google Scholar] [CrossRef] [PubMed]
- Porter, C.J.; Radford, A.D.; Gaskell, R.M.; Ryvar, R.; Coyne, K.P.; Pinchbeck, G.L.; Dawson, S. Comparison of the ability of feline calicivirus (FCV) vaccines to neutralise a panel of current UK FCV isolates. J. Feline Med. Surg. 2008, 10, 32–40. [Google Scholar] [CrossRef]
- Haller, O.; Kochs, G.; Weber, F. The interferon response circuit: Induction and suppression by pathogenic viruses. Virology 2006, 344, 119–130. [Google Scholar] [CrossRef] [PubMed]
- De Los Santos, T.; de Avila Botton, S.; Weiblen, R.; Grubman, M.J. The leader proteinase of foot-and-mouth disease virus inhibits the induction of beta interferon mRNA and blocks the host innate immune response. J. Virol. 2006, 80, 1906–1914. [Google Scholar] [CrossRef] [PubMed]
- Didcock, L.; Young, D.F.; Goodbourn, S.; Randall, R.E. The V protein of simian virus 5 inhibits interferon signalling by targeting STAT1 for proteasome-mediated degradation. J. Virol. 1999, 73, 9928–9933. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Li, M.; Zheng, H.; Muster, T.; Palese, P.; Beg, A.; García-Sastre, A. Influenza A virus NS1 protein prevents activation of NF-kappaB and induction of alpha/beta interferon. J. Virol. 2000, 74, 11566–11577. [Google Scholar] [CrossRef]
- Tian, J.; Zhang, X.; Wu, H.; Liu, C.; Liu, J.; Hu, X.; Qu, L. Assessment of the IFN-β response to four feline caliciviruses: Infection in CRFK cells. Infect. Genet. Evol. 2015, 34, 352–360. [Google Scholar] [CrossRef]
- Van der Meulen, K.; Garré, B.; Croubels, S.; Nauwynck, H. In vitro comparison of antiviral drugs against feline herpesvirus 1. BMC Vet. Res. 2006, 2, 13. [Google Scholar] [CrossRef]
- Fried, W.A.; Soltero-Rivera, M.; Ramesh, A.; Lommer, M.J.; Arzi, B.; DeRisi, J.L.; Horst, J.A. Use of unbiased metagenomic and transcriptomic analyses to investigate the association between feline calicivirus and feline chronic gingivostomatitis in domestic cats. Am. J. Vet. Res. 2021, 82, 381–394. [Google Scholar] [CrossRef] [PubMed]
- Arzi, B.; Mills-Ko, E.; Verstraete, F.J.; Kol, A.; Walker, N.J.; Badgley, M.R.; Fazel, N.; Murphy, W.J.; Vapniarsky, N.; Borjesson, D.L. Therapeutic Efficacy of Fresh, Autologous Mesenchymal Stem Cells for Severe Refractory Gingivostomatitis in Cats. Stem Cells Transl. Med. 2016, 5, 75–86. [Google Scholar] [CrossRef]
- Arzi, B.; Clark, K.C.; Sundaram, A.; Spriet, M.; Verstraete, F.J.M.; Walker, N.J.; Loscar, M.R.; Fazel, N.; Murphy, W.J.; Vapniarsky, N.; et al. Therapeutic Efficacy of Fresh, Allogeneic Mesenchymal Stem Cells for Severe Refractory Feline Chronic Gingivostomatitis. Stem Cells Transl. Med. 2017, 6, 1710–1722. [Google Scholar] [CrossRef]
- Arzi, B.; Peralta, S.; Fiani, N.; Vapniarsky, N.; Taechangam, N.; Delatorre, U.; Clark, K.C.; Walker, N.J.; Loscar, M.R.; Lommer, M.J.; et al. A multicenter experience using adipose-derived mesenchymal stem cell therapy for cats with chronic, non-responsive gingivostomatitis. Stem Cell Res. Ther. 2020, 11, 115. [Google Scholar] [CrossRef] [PubMed]
- Escobar-Herrera, J.; Medina-Ramírez, F.J.; Gutiérrez-Escolano, A.L. A carboxymethyl-cellulose plaque assay for feline calicivirus. J. Virol. Methods 2007, 146, 393–396. [Google Scholar] [CrossRef] [PubMed]
- Ueki, Y.; Shoji, M.; Suto, A.; Tanabe, T.; Okimura, Y.; Kikuchi, Y.; Saito, N.; Sano, D.; Omura, T. Persistence of caliciviruses in artificially contaminated oysters during depuration. Appl. Environ. Microbiol. 2007, 73, 5698–5701. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Kessler, Y.; Helfer-Hungerbuehler, A.K.; Cattori, V.; Meli, M.L.; Zellweger, B.; Ossent, P.; Riond, B.; Reusch, C.E.; Lutz, H.; Hofmann-Lehmann, R. Quantitative TaqMan real-time PCR assays for gene expression normalisation in feline tissues. BMC Mol. Biol. 2009, 10, 106. [Google Scholar] [CrossRef] [PubMed]
- Hussein, I.T.; Field, H.J. Development of a quantitative real-time TaqMan PCR assay for testing the susceptibility of feline herpesvirus-1 to antiviral compounds. J. Virol. Methods 2008, 152, 85–90. [Google Scholar] [CrossRef]
- Khatri, M.; Richardson, L.A.; Meulia, T. Mesenchymal stem cell-derived extracellular vesicles attenuate influenza virus-induced acute lung injury in a pig model. Stem Cell Res. Ther. 2018, 9, 17. [Google Scholar] [CrossRef]
- Qian, X.; Xu, C.; Fang, S.; Zhao, P.; Wang, Y.; Liu, H.; Yuan, W.; Qi, Z. Exosomal MicroRNAs Derived from Umbilical Mesenchymal Stem Cells Inhibit Hepatitis C Virus Infection. Stem Cells Transl. Med. 2016, 5, 1190–1203. [Google Scholar] [CrossRef]
- Oh, S.J.; Lee, E.N.; Park, J.H.; Lee, J.K.; Cho, G.J.; Park, I.H.; Shin, O.S. Anti-Viral Activities of Umbilical Cord Mesenchymal Stem Cell-Derived Small Extracellular Vesicles Against Human Respiratory Viruses. Front. Cell Infect. Microbiol. 2022, 12, 850744. [Google Scholar] [CrossRef]
- Tang, Y.; Zhou, Y.; Li, H.J. Advances in mesenchymal stem cell exosomes: A review. Stem Cell Res. Ther. 2021, 12, 71. [Google Scholar] [CrossRef] [PubMed]
- Chen, J.; Li, P.; Zhang, T.; Xu, Z.; Huang, X.; Wang, R.; Du, L. Review on Strategies and Technologies for Exosome Isolation and Purification. Front. Bioeng. Biotechnol. 2022, 9, 811971. [Google Scholar] [CrossRef] [PubMed]
- Makino, A.; Shimojima, M.; Miyazawa, T.; Kato, K.; Tohya, Y.; Akashi, H. Junctional adhesion molecule 1 is a functional receptor for feline calicivirus. J. Virol. 2006, 80, 4482–4490. [Google Scholar] [CrossRef] [PubMed]
- Agelidis, A.M.; Shukla, D. Cell entry mechanisms of HSV: What we have learned in recent years. Future Virol. 2015, 10, 1145–1154. [Google Scholar] [CrossRef] [PubMed]
- Maeda, K.; Yokoyama, N.; Fujita, K.; Maejima, M.; Mikami, T. Heparin-binding activity of feline herpesvirus type 1 glycoproteins. Virus Res. 1997, 52, 169–176. [Google Scholar] [CrossRef]












{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Target | Oligonucleotide Sequence | |
|---|---|---|
| FCV F4 | Forward | 5′-TCGATTCCTTCGGACCTGATC-3′ |
| Reverse | 5′-AAGTCGAAATGACGGTTTGCTT-3′ | |
| Probe | FAM-TAATCGCTACTGGACTGAC-TAMRA | |
| FHV1 gC | Forward | 5′-ACGGGAAGCCAATAGAAAG-3′ |
| Reverse | 5′-CGGAATAGCCAACACAGAA-3′ | |
| Probe | FAM-ATGAGTCCATCTATCCATACACTTGCCG-TAMRA | |
| GUSB | Forward | 5′-CTACATCGATGACATCACCATCAG-3′ |
| Reverse | 5′-CGCCTTCAACAAAAATCTGGTAA-3′ | |
| Probe | FAM-ACCAGCGTGAACCAAGACACTGGGC-TAMRA | |
| Gene ID | Gene Symbol | Fold Change | Padj | Gene Function |
|---|---|---|---|---|
| ENSFCAG00000035883 | PKLR | 53.00 | 0.0004143 | pyruvate kinase L/R |
| ENSFCAG00000010889 | ADAMTS20 | 36.72 | 0.0079952 | ADAM metallopeptidase with thrombospondin type 1 motif 20 |
| ENSFCAG00000034557 | PDX1 | 31.17 | 0.0195328 | pancreatic and duodenal homeobox 1 |
| ENSFCAG00000031865 | CFAP97D1 | 29.18 | 0.0144898 | CFAP97 domain containing 1 |
| ENSFCAG00000031341 | TMPRSS11E | 28.93 | 0.0130241 | transmembrane serine protease 11E |
| ENSFCAG00000004768 | SHANK2 | 27.41 | 0.0366154 | SH3 and multiple ankyrin repeat domains 2 |
| ENSFCAG00000037311 | RNase_MRP | 26.70 | 0.0240336 | RNase MRP |
| ENSFCAG00000024035 | PAQR9 | 26.63 | 0.0228903 | progestin and adipoQ receptor family member 9 |
| ENSFCAG00000043275 | CAMSAP3 | 26.29 | 0.0470668 | calmodulin regulated spectrin associated protein family member 3 |
| ENSFCAG00000041431 | CXCR5 | 24.09 | 0.0370509 | C-X-C motif chemokine receptor 5 |
| ENSFCAG00000011344 | SLC13A3 | 24.09 | 0.0410041 | solute carrier family 13 member 3 |
| ENSFCAG00000015185 | BPIFB4 | 24.04 | 0.0369054 | BPI fold containing family B member 4 |
| ENSFCAG00000045063 | HES3 | 23.82 | 0.0356022 | hes family bHLH transcription factor 3 |
| ENSFCAG00000041480 | SLAMF7 | 19.60 | 2.676 × 10−27 | SLAM family member 7 |
| ENSFCAG00000010157 | NCKAP1L | 16.13 | 0.0005246 | NCK associated protein 1 like |
| Gene ID | Gene Symbol | Fold Change | Padj | Gene Function |
|---|---|---|---|---|
| ENSFCAG00000045199 | ARX | 0.016 | 2.198 × 10−5 | aristaless related homeobox |
| ENSFCAG00000013814 | SPTSSB | 0.023 | 0.002199 | serine palmitoyltransferase small subunit B |
| ENSFCAG00000043019 | TNMD | 0.032 | 0.0167252 | tenomodulin |
| ENSFCAG00000008166 | S100A14 | 0.035 | 0.024189 | S100 calcium binding protein A14 |
| ENSFCAG00000001649 | CDHR1 | 0.035 | 0.0257187 | cadherin related family member 1 |
| ENSFCAG00000004983 | HTR3A | 0.038 | 0.0357949 | 5-hydroxytryptamine receptor 3A |
| ENSFCAG00000028760 | CB2H6orf52 | 0.038 | 0.0429303 | chromosome B2 C6orf52 homolog |
| ENSFCAG00000000794 | CCDC85A | 0.045 | 1.199 × 10−5 | coiled-coil domain containing 85A |
| ENSFCAG00000004322 | TENM4 | 0.052 | 0.0005413 | teneurin transmembrane protein 4 |
| ENSFCAG00000022100 | PLEKHS1 | 0.072 | 1.323 × 10−11 | pleckstrin homology domain containing S1 |
| ENSFCAG00000033516 | ZC2HC1B | 0.087 | 0.0107755 | zinc finger C2HC-type containing 1B |
| ENSFCAG00000029875 | RERG | 0.088 | 0.0177269 | RAS like estrogen regulated growth inhibitor |
| ENSFCAG00000027949 | ART4 | 0.090 | 2.117 × 10−5 | ADP-ribosyltransferase 4 |
| ENSFCAG00000004490 | PLEKHB1 | 0.092 | 0.0037363 | pleckstrin homology domain containing B1 |
| ENSFCAG00000045651 | ANGPT1 | 0.097 | 4.458 × 10−10 | angiopoietin 1 |
| ID | Category | GO ID | Term | Padj |
|---|---|---|---|---|
| 1 | GO:MF | GO:0019955 | cytokine binding | 4.93 × 10−2 |
| 2 | GO:BP | GO:0032502 | developmental process | 3.06 × 10−5 |
| 3 | GO:BP | GO:0051179 | localization | 2.61 × 10−4 |
| 4 | GO:BP | GO:0040011 | locomotion | 5.82 × 10−4 |
| 5 | GO:BP | GO:0016477 | cell migration | 6.42 × 10−4 |
| 6 | GO:BP | GO:0051674 | localization of cell | 7.73 × 10−4 |
| 7 | GO:BP | GO:0048870 | cell motility | 7.73 × 10−4 |
| 8 | GO:BP | GO:0009653 | anatomical structure morphogenesis | 8.51 × 10−4 |
| 9 | GO:BP | GO:0051239 | regulation of multicellular organismal process | 1.41 × 10−3 |
| 10 | GO:BP | GO:0010033 | response to organic substance | 1.58 × 10−3 |
| 11 | GO:BP | GO:0032103 | positive regulation of response to external stimulus | 1.61 × 10−3 |
| 12 | GO:CC | GO:0071944 | cell periphery | 7.78 × 10−4 |
| 13 | GO:CC | GO:0012505 | endomembrane system | 1.58 × 10−3 |
| 14 | GO:CC | GO:0009986 | cell surface | 7.39 × 10−3 |
| 15 | GO:CC | GO:0005576 | extracellular region | 1.03 × 10−2 |
| 16 | GO:CC | GO:0005886 | plasma membrane | 3.77 × 10−2 |
| 17 | GO:CC | GO:0016020 | membrane | 4.46 × 10−2 |
| 18 | GO:CC | GO:0005615 | extracellular space | 4.91 × 10−2 |
| 19 | KEGG | KEGG:03320 | PPAR signaling pathway | 1.30 × 10−4 |
| ID | Category | GO ID | Term | Padj |
|---|---|---|---|---|
| 1 | GO:MF | GO:0005515 | protein binding | 1.13 × 10−4 |
| 2 | GO:MF | GO:0005126 | cytokine receptor binding | 2.27 × 10−3 |
| 3 | GO:MF | GO:0001730 | 2′−5′-oligoadenylate synthetase activity | 1.06 × 10−2 |
| 4 | GO:MF | GO:0005102 | signaling receptor binding | 1.47 × 10−2 |
| 5 | GO:MF | GO:0003725 | double-stranded RNA binding | 2.72 × 10−2 |
| 6 | GO:MF | GO:0003950 | NAD + ADP-ribosyltransferase activity | 4.41 × 10−2 |
| 7 | GO:BP | GO:0009605 | response to external stimulus | 1.95 × 10−10 |
| 8 | GO:BP | GO:0050896 | response to stimulus | 7.23 × 10−10 |
| 9 | GO:BP | GO:0007275 | multicellular organism development | 5.75 × 10−9 |
| 10 | GO:BP | GO:0048856 | anatomical structure development | 7.12 × 10−9 |
| 11 | GO:BP | GO:0002376 | immune system process | 1.15 × 10−8 |
| 12 | GO:BP | GO:0048731 | system development | 4.90 × 10−8 |
| 13 | GO:BP | GO:0006955 | immune response | 6.33 × 10−8 |
| 14 | GO:BP | GO:0140546 | defense response to symbiont | 9.95 × 10−8 |
| 15 | GO:BP | GO:0051607 | defense response to virus | 9.95 × 10−8 |
| 16 | GO:BP | GO:0032502 | developmental process | 1.57 × 10−7 |
| 17 | GO:CC | GO:0071944 | cell periphery | 8.23 × 10−7 |
| 18 | GO:CC | GO:0005886 | plasma membrane | 5.39 × 10−4 |
| 19 | GO:CC | GO:0062023 | collagen-containing extracellular matrix | 2.93 × 10−2 |
| 20 | GO:CC | GO:1990584 | cardiac Troponin complex | 3.45 × 10−2 |
| No | Pathway_ID | Pathway | NumGenes | ES | NES | Size | Padj |
|---|---|---|---|---|---|---|---|
| 1 | fca04141 | Protein processing in endoplasmic reticulum | 67 | 0.577 | 2.182 | 142 | 0.005 |
| 2 | fca04150 | mTOR signaling pathway | 40 | 0.464 | 1.736 | 132 | 0.005 |
| 3 | fca04142 | Lysosome | 52 | 0.688 | 2.498 | 108 | 0.005 |
| 4 | fca04145 | Phagosome | 31 | 0.601 | 2.143 | 94 | 0.005 |
| 5 | fca00190 | Oxidative phosphorylation | 25 | 0.597 | 2.053 | 75 | 0.005 |
| 6 | fca04721 | Synaptic vesicle cycle | 20 | 0.66 | 2.148 | 55 | 0.005 |
| 7 | fca01212 | Fatty acid metabolism | 23 | 0.645 | 2.023 | 45 | 0.005 |
| 8 | fca00520 | Amino sugar and nucleotide sugar metabolism | 21 | 0.674 | 2.061 | 39 | 0.005 |
| 9 | fca03050 | Proteasome | 28 | 0.631 | 1.894 | 35 | 0.005 |
| 10 | fca00020 | Citrate cycle (TCA cycle) | 13 | 0.719 | 2.038 | 27 | 0.005 |
| 11 | fca01040 | Biosynthesis of unsaturated fatty acids | 13 | 0.726 | 1.926 | 20 | 0.005 |
| 12 | fca04966 | Collecting duct acid secretion | 10 | 0.764 | 2.048 | 21 | 0.005 |
| 13 | fca_M00160 | V-type ATPase, eukaryotes | 12 | 0.815 | 2.185 | 21 | 0.005 |
| 14 | fca00100 | Steroid biosynthesis | 11 | 0.866 | 2.185 | 16 | 0.005 |
| 15 | fca_M00341 | Proteasome, 19S regulatory particle (PA700) | 17 | 0.782 | 1.999 | 17 | 0.005 |
| 16 | fca00531 | Glycosaminoglycan degradation | 10 | 0.83 | 2.032 | 14 | 0.005 |
| 17 | fca_M00101 | Cholesterol biosynthesis, squalene 2,3-epoxide => cholesterol | 8 | 0.965 | 2.096 | 9 | 0.005 |
| 18 | fca05160 | Hepatitis C | 25 | −0.465 | −1.814 | 107 | 0.007 |
| 19 | fca05168 | Herpes simplex infection | 30 | −0.443 | −1.766 | 122 | 0.007 |
| 20 | fca01200 | Carbon metabolism | 35 | 0.496 | 1.783 | 99 | 0.007 |
| 21 | fca05323 | Rheumatoid arthritis | 17 | 0.571 | 1.885 | 59 | 0.007 |
| 22 | fca_M00001 | Glycolysis (Embden-Meyerhof pathway), glucose => pyruvate | 11 | 0.733 | 1.921 | 19 | 0.008 |
| 23 | fca00062 | Fatty acid elongation | 12 | 0.697 | 1.868 | 21 | 0.01 |
| 24 | fca00604 | Glycosphingolipid biosynthesis—ganglio series | 7 | 0.786 | 1.887 | 13 | 0.01 |
| 25 | fca04390 | Hippo signaling pathway | 52 | −0.406 | −1.642 | 134 | 0.01 |
| 26 | fca_M00285 | MCM complex | 6 | −0.919 | −1.838 | 6 | 0.011 |
| 27 | fca04976 | Bile secretion | 14 | 0.55 | 1.779 | 53 | 0.011 |
| 28 | fca05162 | Measles | 20 | −0.454 | −1.733 | 94 | 0.013 |
| 29 | fca04110 | Cell cycle | 45 | −0.434 | −1.7 | 112 | 0.013 |
| 30 | fca01230 | Biosynthesis of amino acids | 23 | 0.552 | 1.808 | 57 | 0.013 |
| 31 | fca00051 | Fructose and mannose metabolism | 15 | 0.636 | 1.826 | 29 | 0.013 |
| 32 | fca_M00003 | Gluconeogenesis, oxaloacetate => fructose-6P | 7 | 0.797 | 1.876 | 12 | 0.016 |
| 33 | fca05161 | Hepatitis B | 23 | −0.405 | −1.608 | 121 | 0.019 |
| 34 | fca04115 | p53 signaling pathway | 19 | −0.506 | −1.777 | 58 | 0.023 |
| 35 | fca05164 | Influenza A | 24 | −0.412 | −1.632 | 116 | 0.024 |
| 36 | fca04015 | Rap1 signaling pathway | 27 | −0.364 | −1.519 | 165 | 0.024 |
| 37 | fca_M00079 | Keratan sulfate degradation | 5 | 0.92 | 1.717 | 5 | 0.024 |
| 38 | fca_M00679 | BMP signaling | 8 | −0.707 | −1.884 | 17 | 0.024 |
| 39 | fca_M00009 | Citrate cycle (TCA cycle, Krebs cycle) | 11 | 0.668 | 1.789 | 21 | 0.024 |
| 40 | fca_M00415 | Fatty acid biosynthesis, elongation, endoplasmic reticulum | 10 | 0.757 | 1.82 | 13 | 0.03 |
| 41 | fca_M00077 | Chondroitin sulfate degradation | 4 | 0.909 | 1.698 | 5 | 0.032 |
| 42 | fca04152 | AMPK signaling pathway | 31 | 0.435 | 1.586 | 110 | 0.035 |
| 43 | fca04964 | Proximal tubule bicarbonate reclamation | 5 | 0.725 | 1.775 | 14 | 0.04 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Teshima, T.; Yasumura, Y.; Suzuki, R.; Matsumoto, H. Antiviral Effects of Adipose Tissue-Derived Mesenchymal Stem Cells Secretome against Feline Calicivirus and Feline Herpesvirus Type 1. Viruses 2022, 14, 1687. https://doi.org/10.3390/v14081687
Teshima T, Yasumura Y, Suzuki R, Matsumoto H. Antiviral Effects of Adipose Tissue-Derived Mesenchymal Stem Cells Secretome against Feline Calicivirus and Feline Herpesvirus Type 1. Viruses. 2022; 14(8):1687. https://doi.org/10.3390/v14081687
Chicago/Turabian StyleTeshima, Takahiro, Yuyo Yasumura, Ryohei Suzuki, and Hirotaka Matsumoto. 2022. "Antiviral Effects of Adipose Tissue-Derived Mesenchymal Stem Cells Secretome against Feline Calicivirus and Feline Herpesvirus Type 1" Viruses 14, no. 8: 1687. https://doi.org/10.3390/v14081687
APA StyleTeshima, T., Yasumura, Y., Suzuki, R., & Matsumoto, H. (2022). Antiviral Effects of Adipose Tissue-Derived Mesenchymal Stem Cells Secretome against Feline Calicivirus and Feline Herpesvirus Type 1. Viruses, 14(8), 1687. https://doi.org/10.3390/v14081687