
Detection of Feline Coronavirus Variants in Cats without Feline Infectious Peritonitis

 ,
,  and
and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Study Population and Exclusion of FIP
2.2. Sample Collection
2.3. FCoV 7b Gene RT-qPCR
2.4. FCoV S Gene Mutation RT-qPCR
2.5. Sanger Sequencing
3. Results
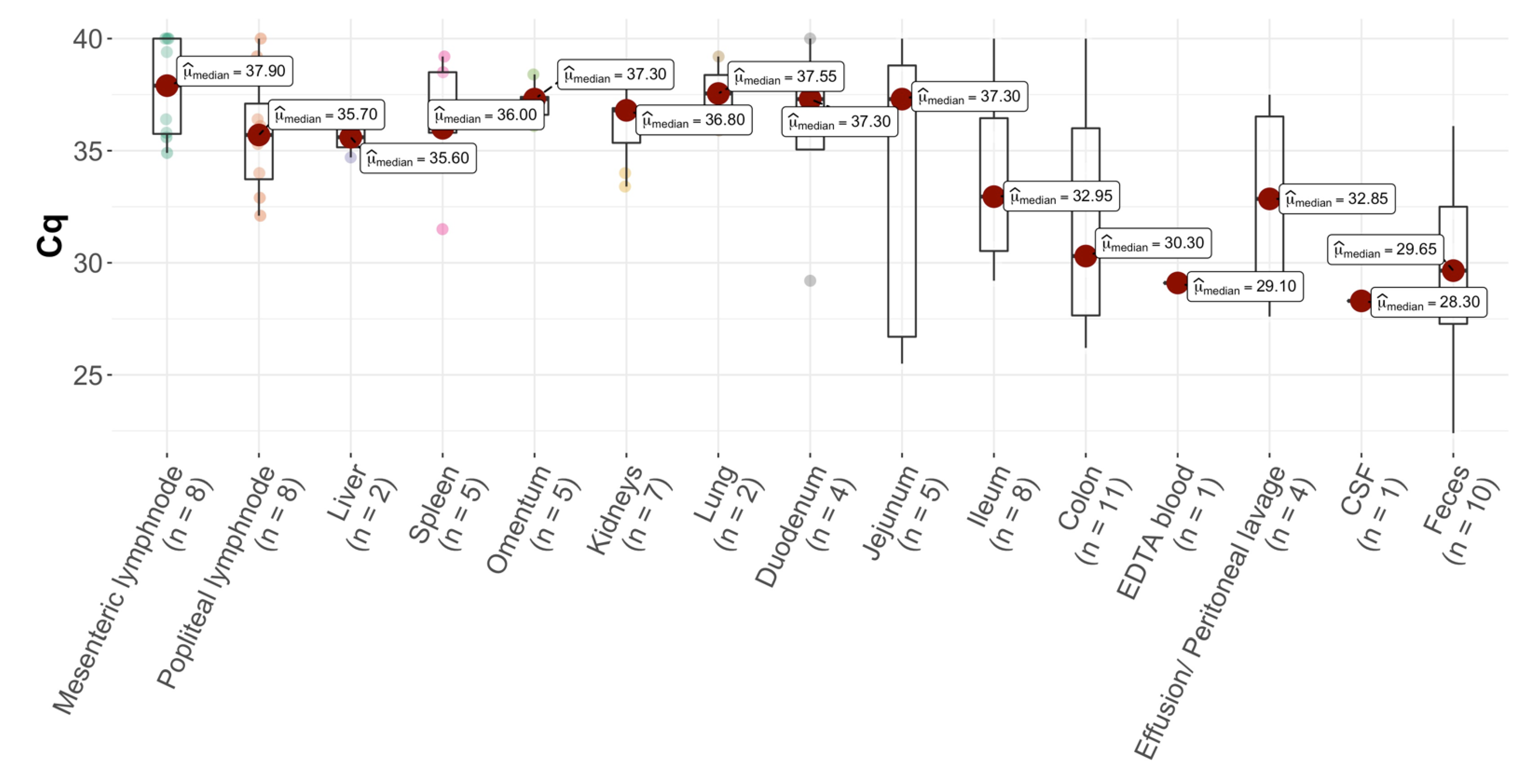
3.1. Presence of FCoV RNA in Cats without FIP
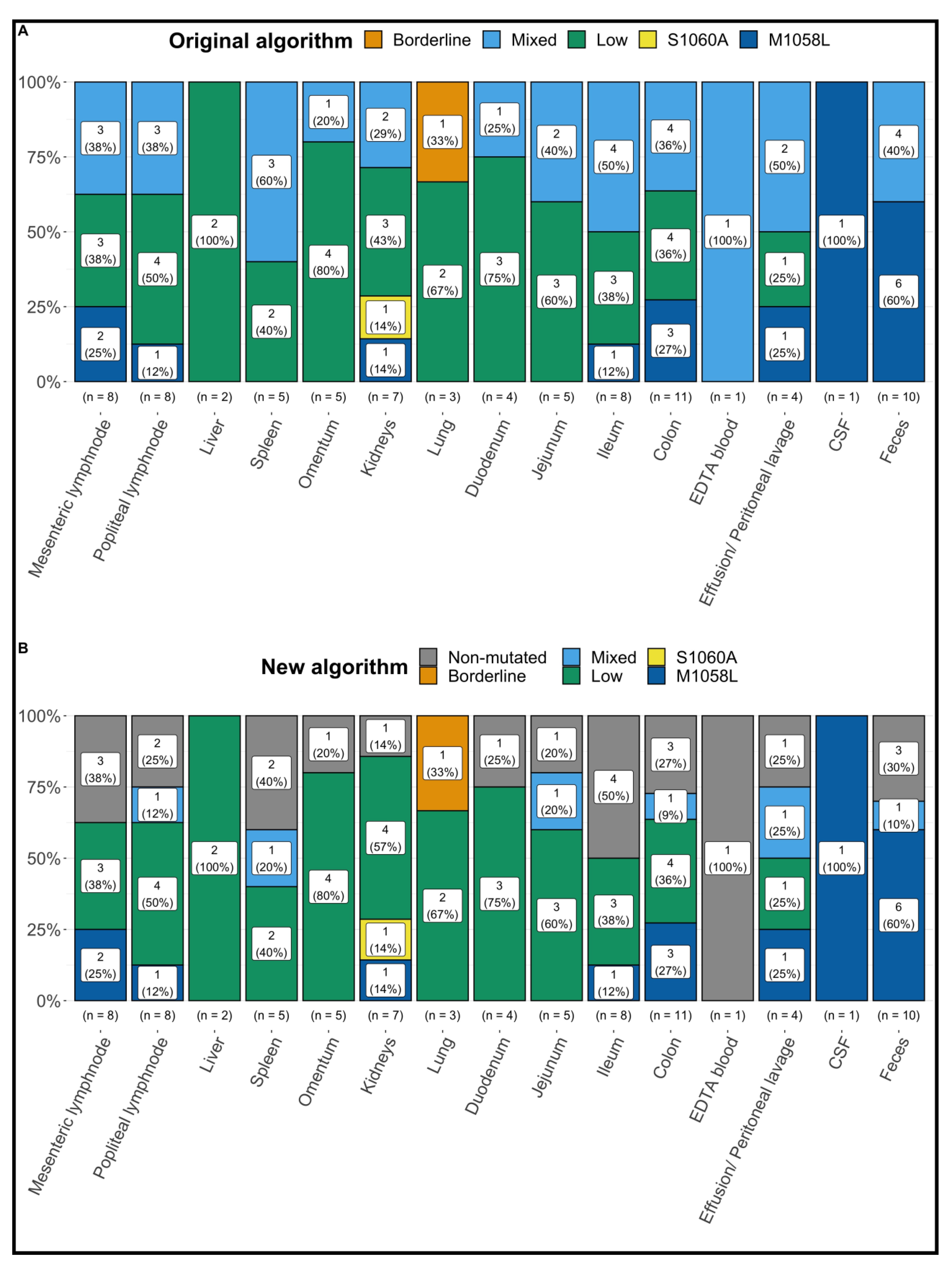
3.2. Results of FCoV S Gene Mutation RT-qPCR following the Original Algorithm
3.3. Results of FCoV S Gene Mutation RT-qPCR following the New Algorithm
3.4. Results of Sanger Sequencing
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Addie, D.D.; Jarrett, O. A Study of Naturally Occurring Feline Coronavirus Infections in Kittens. Vet. Rec. 1992, 130, 133–137. [Google Scholar] [CrossRef] [PubMed]
- Kipar, A.; Meli, M.L.; Baptiste, K.E.; Bowker, L.J.; Lutz, H. Sites of Feline Coronavirus Persistence in Healthy Cats. J. Gen. Virol. 2010, 91, 1698–1707. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Desmarets, L.M.B.; Vermeulen, B.L.; Theuns, S.; Conceicąõ-Neto, N.; Zeller, M.; Roukaerts, I.D.M.; Acar, D.D.; Olyslaegers, D.A.J.; van Ranst, M.; Matthijnssens, J.; et al. Experimental Feline Enteric Coronavirus Infection Reveals an Aberrant Infection Pattern and Shedding of Mutants with Impaired Infectivity in Enterocyte Cultures. Sci. Rep. 2016, 6, 20022. [Google Scholar] [CrossRef] [PubMed]
- Meli, M.; Kipar, A.; Müller, C.; Jenal, K.; Gönczi, E.; Borel, N.; Gunn-Moore, D.; Chalmers, S.; Lin, F.; Reinacher, M.; et al. High Viral Loads despite Absence of Clinical and Pathological Findings in Cats Experimentally Infected with Feline Coronavirus (FCoV) Type I and in Naturally FCoV-Infected Cats. J. Feline Med. Surg. 2004, 6, 69–81. [Google Scholar] [CrossRef] [Green Version]
- Kipar, A.; Meli, M.L.; Failing, K.; Euler, T.; Gomes-Keller, M.A.; Schwartz, D.; Lutz, H.; Reinacher, M. Natural Feline Coronavirus Infection: Differences in Cytokine Patterns in Association with the Outcome of Infection. Vet. Immunol. Immunopathol. 2006, 112, 141–155. [Google Scholar] [CrossRef] [PubMed]
- Malbon, A.J.; Meli, M.L.; Barker, E.N.; Davidson, A.D.; Tasker, S.; Kipar, A. Inflammatory Mediators in the Mesenteric Lymph Nodes, Site of a Possible Intermediate Phase in the Immune Response to Feline Coronavirus and the Pathogenesis of Feline Infectious Peritonitis? J. Comp. Pathol. 2019, 166, 69–86. [Google Scholar] [CrossRef] [PubMed]
- Hu, C.M.J.; Chang, W.S.; Fang, Z.S.; Chen, Y.T.; Wang, W.L.; Tsai, H.H.; Chueh, L.L.; Takano, T.; Hohdatsu, T.; Chen, H.W. Nanoparticulate Vacuolar ATPase Blocker Exhibits Potent Host-Targeted Antiviral Activity against Feline Coronavirus. Sci. Rep. 2017, 7, 13043. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Herrewegh, A.A.P.M.; Smeenk, I.; Horzinek, M.C.; Rottier, P.J.M.; de Groot, R.J. Feline Coronavirus Type II Strains 79-1683 and 79-1146 Originate from a Double Recombination between Feline Coronavirus Type I and Canine Coronavirus. J. Virol. 1998, 72, 4508–4514. [Google Scholar] [CrossRef] [Green Version]
- Xia, H.; Li, X.; Zhao, W.; Jia, S.; Zhang, X.; Irwin, D.M.; Zhang, S. Adaptive Evolution of Feline Coronavirus Genes Based on Selection Analysis. BioMed Res. Int. 2020, 2020, 9089768. [Google Scholar] [CrossRef] [PubMed]
- Vennema, H.; Poland, A.; Foley, J.; Pedersen, N.C. Feline Infectious Peritonitis Viruses Arise by Mutation from Endemic Feline Enteric Coronaviruses. Virology 1998, 243, 150–157. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chang, H.W.; Egberink, H.F.; Halpin, R.; Spiro, D.J.; Rottie, P.J.M. Spike Protein Fusion Peptide and Feline Coronavirus Virulence. Emerg. Infect. Dis. 2012, 18, 1089–1095. [Google Scholar] [CrossRef]
- Pedersen, N.C.; Liu, H.; Scarlett, J.; Leutenegger, C.M.; Golovko, L.; Kennedy, H.; Kamal, F.M. Feline Infectious Peritonitis: Role of the Feline Coronavirus 3c Gene in Intestinal Tropism and Pathogenicity Based upon Isolates from Resident and Adopted Shelter Cats. Virus. Res. 2012, 165, 17–28. [Google Scholar] [CrossRef] [PubMed]
- Bank-Wolf, B.R.; Stallkamp, I.; Wiese, S.; Moritz, A.; Tekes, G.; Thiel, H.J. Mutations of 3c and Spike Protein Genes Correlate with the Occurrence of Feline Infectious Peritonitis. Vet. Microbiol. 2014, 173, 177–188. [Google Scholar] [CrossRef]
- Borschensky, C.M.; Reinacher, M. Mutations in the 3c and 7b Genes of Feline Coronavirus in Spontaneously Affected FIP Cats. Res. Vet. Sci. 2014, 97, 333–340. [Google Scholar] [CrossRef] [PubMed]
- Bosch, B.J.; van der Zee, R.; de Haan, C.A.M.; Rottier, P.J.M. The Coronavirus Spike Protein Is a Class I Virus Fusion Protein: Structural and Functional Characterization of the Fusion Core Complex. J. Virol. 2003, 77, 8801–8811. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lewis, C.S.; Porter, E.; Matthews, D.; Kipar, A.; Tasker, S.; Helps, C.R.; Siddell, S.G. Genotyping Coronaviruses Associated with Feline Infectious Peritonitis. J. Gen. Virol. 2015, 96, 1358–1368. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Licitra, B.N.; Millet, J.K.; Regan, A.D.; Hamilton, B.S.; Rinaldi, V.D.; Duhamel, G.E.; Whittaker, G.R. Mutation in Spike Protein Cleavage Site and Pathogenesis of Feline Coronavirus. Emerg. Infect. Dis. 2013, 19, 1066–1073. [Google Scholar] [CrossRef] [PubMed]
- Shirato, K.; Chang, H.W.; Rottier, P.J.M. Differential Susceptibility of Macrophages to Serotype II Feline Coronaviruses Correlates with Differences in the Viral Spike Protein. Virus Res. 2018, 255, 14–23. [Google Scholar] [CrossRef]
- Porter, E.; Tasker, S.; Day, M.J.; Harley, R.; Kipar, A.; Siddell, S.G.; Helps, C.R. Amino Acid Changes in the Spike Protein of Feline Coronavirus Correlate with Systemic Spread of Virus from the Intestine and Not with Feline Infectious Peritonitis. Vet. Res. 2014, 45, 49–59. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hartmann, K.; Binder, C.; Hirschberger, J.; Cole, D.; Reinacher, M.; Schroo, S.; Frost, J.; Egberink, H.; Lutz, H.; Hermanns, W. Comparison of Different Tests to Diagnose Feline Infectious Peritonitis. J. Vet. Intern. Med. 2003, 17, 781–790. [Google Scholar] [CrossRef] [PubMed]
- Kipar, A.; Bellmann, S.; Kremendahl, J.; Köhler, K.; Reinacher, M. Cellular Composition, Coronavirus Antigen Expression and Production of Specific Antibodies in Lesions in Feline Infectious Peritonitis. Vet. Immunol. Immunopathol. 1998, 65, 243–257. [Google Scholar] [CrossRef]
- Sharif, S.; Arshad, S.S.; Hair-Bejo, M.; Omar, A.R.; Zeenathul, N.A.; Alazawy, A. Diagnostic Methods for Feline Coronavirus: A Review. Vet. Med. Int. 2010, 2010, 809480. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Barker, E.N.; Stranieri, A.; Helps, C.R.; Porter, E.L.; Davidson, A.D.; Day, M.J.; Knowles, T.; Kipar, A.; Tasker, S. Limitations of Using Feline Coronavirus Spike Protein Gene Mutations to Diagnose Feline Infectious Peritonitis. Vet. Res. 2017, 48, 60. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dunbar, D.; Kwok, W.; Graham, E.; Armitage, A.; Irvine, R.; Johnston, P.; McDonald, M.; Montgomery, D.; Nicolson, L.; Robertson, E.; et al. Diagnosis of Non-Effusive Feline Infectious Peritonitis by Reverse Transcriptase Quantitative PCR from Mesenteric Lymph Node Fine-Needle Aspirates. J. Feline Med. Surg. 2019, 21, 910–921. [Google Scholar] [CrossRef] [Green Version]
- Felten, S.; Leutenegger, C.M.; Balzer, H.J.; Pantchev, N.; Matiasek, K.; Wess, G.; Egberink, H.; Hartmann, K. Sensitivity and Specificity of a Real-Time Reverse Transcriptase Polymerase Chain Reaction Detecting Feline Coronavirus Mutations in Effusion and Serum/Plasma of Cats to Diagnose Feline Infectious Peritonitis. BMC Vet. Res. 2017, 13, 228. [Google Scholar] [CrossRef] [PubMed]
- Longstaff, L.; Porter, E.; Crossley, V.J.; Hayhow, S.E.; Helps, C.R.; Tasker, S. Feline Coronavirus Quantitative Reverse Transcriptase Polymerase Chain Reaction on Effusion Samples in Cats with and without Feline Infectious Peritonitis. J. Feline Med. Surg. 2017, 19, 240–245. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Addie, D.; Belák, S.; Boucraut-Baralon, C.; Egberink, H.; Frymus, T.; Gruffydd-Jones, T.; Hartmann, K.; Hosie, M.J.; Lloret, A.; Lutz, H.; et al. Feline Infectious Peritonitis ABCD Guidelines on Prevention and Management. J. Feline Med. Surg. 2009, 11, 594–604. [Google Scholar] [CrossRef] [PubMed]
- Sangl, L.; Matiasek, K.; Felten, S.; Gründl, S.; Bergmann, M.; Balzer, H.J.; Pantchev, N.; Leutenegger, C.M.; Hartmann, K. Detection of Feline Coronavirus Mutations in Paraffin-Embedded Tissues in Cats with Feline Infectious Peritonitis and Controls. J. Feline Med. Surg. 2019, 21, 133–142. [Google Scholar] [CrossRef] [Green Version]
- Emmler, L.; Felten, S.; Matiasek, K.; Balzer, H.J.; Pantchev, N.; Leutenegger, C.; Hartmann, K. Feline Coronavirus with and without Spike Gene Mutations Detected by Real-Time RT-PCRs in Cats with Feline Infectious Peritonitis. J. Feline Med. Surg. 2020, 22, 791–799. [Google Scholar] [CrossRef] [PubMed]
- Felten, S.; Matiasek, K.; Leutenegger, C.M.; Sangl, L.; Herre, S.; Dörfelt, S.; Fischer, A.; Hartmann, K. Diagnostic Value of Detecting Feline Coronavirus Rna and Spike Gene Mutations in Cerebrospinal Fluid to Confirm Feline Infectious Peritonitis. Viruses 2021, 13, 186. [Google Scholar] [CrossRef]
- Sangl, L.; Felten, S.; Matiasek, K.; Dörfelt, S.; Bergmann, M.; Balzer, H.J.; Pantchev, N.; Leutenegger, C.; Hartmann, K. Detection of Feline Coronavirus RNA, Spike Gene Mutations, and Feline Coronavirus Antigen in Macrophages in Aqueous Humor of Cats in the Diagnosis of Feline Infectious Peritonitis. J. Vet. Diagn. 2020, 32, 527–534. [Google Scholar] [CrossRef] [PubMed]
- Felten, S.; Hartmann, K. Diagnosis of Feline Infectious Peritonitis: A Review of the Current Literature. Viruses 2019, 11, 1068. [Google Scholar] [CrossRef] [Green Version]
- Felten, S.; Matiasek, K.; Gruendl, S.; Sangl, L.; Wess, G.; Hartmann, K. Investigation into the Utility of an Immunocytochemical Assay in Body Cavity Effusions for Diagnosis of Feline Infectious Peritonitis. J. Feline Med. Surg. 2017, 19, 410–418. [Google Scholar] [CrossRef]
- Gut, M.; Leutenegger, C.M.; Huder, J.B.; Pedersen, N.C.; Lutz, H. One-Tube Fluorogenic Reverse Transcription-Polymerase Chain Reaction for the Quantitation of Feline Coronaviruses. J. Virol. Methods 1999, 77, 37–46. [Google Scholar] [CrossRef]
- Lutz, M.; Steiner, A.R.; Cattori, V.; Hofmann-Lehmann, R.; Lutz, H.; Kipar, A.; Meli, M.L. Fcov Viral Sequences of Systemically Infected Healthy Cats Lack Gene Mutations Previously Linked to the Development of Fip. Pathogens 2020, 9, 603. [Google Scholar] [CrossRef] [PubMed]
- Meli, M.L.; Spiri, A.M.; Zwicklbauer, K.; Krentz, D.; Felten, S.; Bergmann, M.; Dorsch, R.; Matiasek, K.; Alberer, M.; Kolberg, L.; et al. Fecal Feline Coronavirus RNA Shedding and Spike Gene Mutations in Cats with Feline Infectious Peritonitis Treated with GS-441524. Viruses 2022, 14, 1069. [Google Scholar] [CrossRef] [PubMed]
- Barker, E.N.; Tasker, S.; Gruffydd-Jones, T.J.; Tuplin, C.K.; Burton, K.; Porter, E.; Day, M.J.; Harley, R.; Fews, D.; Helps, C.R.; et al. Phylogenetic Analysis of Feline Coronavirus Strains in an Epizootic Outbreak of Feline Infectious Peritonitis. J. Vet. Intern. Med. 2013, 27, 445–450. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vogel, L.; van der Lubben, M.; te Lintelo, E.G.; Bekker, C.P.J.; Geerts, T.; Schuijff, L.S.; Grinwis, G.C.M.; Egberink, H.F.; Rottier, P.J.M. Pathogenic Characteristics of Persistent Feline Enteric Coronavirus Infection in Cats. Vet. Res. 2010, 41, 71–82. [Google Scholar] [CrossRef] [PubMed] [Green Version]


{kind=link}
{kind=link}
| Original Algorithm | New Algorithm | |||
|---|---|---|---|---|
| Total number of cats | 87 | 87 | ||
| FCoV 7b Gene RT-qPCR | FCoV 7b gene RT-qPCR- negative in all samples | 66 | 66 | |
| FCoV 7b gene RT-qPCR- positive in at least 1 sample | 21 | 21 | ||
| FCoV S Gene Mutation RT-qPCR0 | Same Results in All Samples | 1Only Non-mutated | 0 | 0 |
| 2Only Mutated (M1058L *) | 2 | 2 | ||
| 3Only Mutated (S1060A ¶) | 0 | 0 | ||
| 4Only Mixed § | 1 | 0 | ||
| Only Low ‡ | 7 | 7 | ||
| Different Results in Different Samples | Mixed § and Mutated (M1058L *) | 1 | 0 | |
| Mixed § and Low ‡ | 4 | 0 | ||
| Borderline Ω, Mixed § and Low ‡ | 1 | 0 | ||
| Mutated (M1058L *) and Low ‡ | 3 | 3 | ||
| Mutated (both M1058L * and S1060A ¶) and Low ‡ | 1 | 1 | ||
| Mixed § and Mutated (M1058L *) and Low ‡ | 1 | 0 | ||
| Non-mutated and Low ‡ | 0 | 4 | ||
| Non-mutated and M1058L * | 0 | 1 | ||
| Non-mutated and Mixed § | 0 | 1 | ||
| Non-mutated, M1058L *, Mixed § and Low ‡ | 0 | 1 | ||
| Non-mutated, Borderline Ω, Mixed § and Low ‡ | 0 | 1 |
| Cat | Mesenteric Lymph Node | Popliteal Lymph Node | Liver | Spleen | Omentum | Kidneys | Lung | Duodenum | Jejunum | Ileum | Colon | EDTA Blood | Effusion/Peritoneal Lavage | CSF | Aqueous Humor | Feces | |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 1 | Cq | 39.4 | >40.0 | 39.0 | |||||||||||||
| Origi-nal | Low ‡ | - | - | - | - | - | - | Low ‡ | - | Low ‡ | - | - | - | - | - | - | |
| New | Low ‡ | - | - | - | - | - | - | Low ‡ | - | Low ‡ | - | - | - | - | - | - | |
| 2 | Cq | 31.5 | 39.2 | ||||||||||||||
| Origi-nal | - | - | - | Mixed§ | - | - | Low ‡ | - | - | - | - | - | - | - | - | - | |
| New | Non-mutated | Low ‡ | |||||||||||||||
| 3 | Cq | 34.9 | 35.3 | 34.7 | 38.5 | 34.0 | |||||||||||
| Origi-nal | Mixed§ | Low ‡ | Low ‡ | Low ‡ | - | Low ‡ | - | - | - | - | - | - | - | - | - | - | |
| New | Non-mutated | Low ‡ | Low ‡ | Low ‡ | Low ‡ | ||||||||||||
| 4 | Cq | 38.3 | |||||||||||||||
| Origi-nal | - | - | - | - | - | Low ‡ | - | - | - | - | - | - | - | - | - | - | |
| New | Low ‡ | ||||||||||||||||
| 5 | Cq | 35.6 | 34.0 | 36.5 | 35.8 | 36.1 | 36.7 | 35.9 | 29.2 | 26.7 | 29.2 | 26.2 | 29.1 | 27.6 | 27.5 | ||
| Origi-nal | Mixed § | Mixed § | Low ‡ | Mixed § | Mixed § | Mixed § | Mixed § | Mixed § | Mixed § | Mixed § | Mixed § | Mixed § | - | - | Mixed § | ||
| New | Non-mutated | Non-mutated | Low ‡ | Non-mutated | Non-mutated | Non-mutated | Low ‡ | Non-mutated | Non-mutated | Non-mutated | Non-mutated | Non-mutated | Non-mutated | Non-mutated | |||
| 6 | Cq | >40.0 | >40.0 | ||||||||||||||
| Origi-nal | Low ‡ | Low ‡ | - | - | - | - | - | - | - | - | - | - | - | - | - | - | |
| New | Low ‡ | Low ‡ | |||||||||||||||
| 7 | Cq | >40.0 | |||||||||||||||
| Origi-nal | - | - | - | - | - | - | - | - | - | Low ‡ | - | - | - | - | - | - | |
| New | Low ‡ | ||||||||||||||||
| 8 | Cq | >40.0 | |||||||||||||||
| Origi-nal | - | - | - | - | - | - | - | - | - | - | Low ‡ | - | - | - | - | - | |
| New | Low ‡ | ||||||||||||||||
| 9 | Cq | 38.4 | |||||||||||||||
| Origi-nal | - | - | - | - | Low ‡ | - | - | - | - | - | - | - | - | - | - | - | |
| New | Low ‡ | ||||||||||||||||
| 10 | Cq | 29.5 | 28.3 | ||||||||||||||
| Origi-nal | - | - | - | - | - | - | - | - | - | - | - | - | M1058L * | M1058L * | - | - | |
| New | M1058L * | M1058L * | |||||||||||||||
| 11 | Cq | Cq 35.8 | 35.2 (FNA) 32.1 | 37.3 | 36.8 | >40.0 | 30.7 | 28.1 | 36.2 | ||||||||
| Origi-nal | M1058L * | M1058L * | - | - | Low ‡ | M1058L * | - | - | Low ‡ | M1058L * | M1058L * | - | Low ‡ | - | - | - | |
| New | M1058L * | M1058L * | Low ‡ | M1058L * | Low ‡ | M1058L * | M1058L * | Low ‡ | |||||||||
| 12 | Cq | 38.8 | |||||||||||||||
| Origi-nal | - | - | - | - | - | - | - | - | Low ‡ | - | - | - | - | - | - | - | |
| New | Low ‡ | ||||||||||||||||
| 13 | Cq | >40.0 | 36.4 | 39.2 | 36.6 | 36.8 | No Cq value | 37.0 | 25.5 | 30.0 | 26.3 | 23.2 | |||||
| Origi-nal | Mixed § | Mixed § | - | Mixed § | Low ‡ | Mixed § | Borderline Ω | Low ‡ | Mixed § | Mixed § | Mixed § | - | - | - | - | Mixed § | |
| New | Non-mutated | Non-mutated | Mixed § | Low ‡ | Low ‡ | Borderline Ω | Low ‡ | Mixed § | Non-mutated | Mixed § | Mixed § | ||||||
| 14 | Cq | 36.4 | 39.2 | 36.0 | 37.4 | 33.4 | 35.6 | 27.2 | 22.4 | ||||||||
| Origi-nal | M1058L * | Low ‡ | - | Low ‡ | Low ‡ | S1060A ¶ | - | - | - | Low ‡ | M1058L * | - | - | - | - | M1058L * | |
| New | M1058L * | Low ‡ | Low ‡ | Low ‡ | S1060A ¶ | Low ‡ | M1058L * | M1058L * | |||||||||
| 15 | Cq | >40.0 | 37.8 | 27.2 | |||||||||||||
| Origi-nal | Low ‡ | - | - | - | - | - | - | - | - | - | Low ‡ | - | - | - | M1058L * | ||
| New | Low ‡ | Low ‡ | M1058L * | ||||||||||||||
| 16 | Cq | 32.9 | 37.0 | 37.6 | 37.3 | 33.1 | 29.6 | 32.2 | |||||||||
| Origi-nal | - | Mixed § | - | - | - | Low ‡ | - | Low ‡ | Low ‡ | Mixed § | M1058L * | - | - | - | - | M1058L * | |
| New | Mixed § | Low ‡ | Low ‡ | Low ‡ | Non-mutated | M1058L * | M1058L * | ||||||||||
| 17 | Cq | 32.8 | 34.2 | 30.6 | |||||||||||||
| Origi-nal | - | - | - | - | - | - | - | - | - | Mixed § | Low ‡ | - | - | - | - | Mixed § | |
| New | Non-mutated | Low ‡ | Non-mutated | ||||||||||||||
| 18 | Cq | 33.8 | 32.6 | ||||||||||||||
| Origi-nal | - | - | - | - | - | - | - | - | - | - | Low ‡ | - | - | - | - | M1058L * | |
| New | Low ‡ | M1058L * | |||||||||||||||
| 19 | Cq | 38.7 | 36.1 | ||||||||||||||
| Origi-nal | - | - | - | - | - | - | - | - | - | - | Mixed § | - | - | - | - | M1058L * | |
| New | Non-mutated | M1058L * | |||||||||||||||
| 20 | Cq | 36.1 | 30.3 | 37.5 | 28.7 | ||||||||||||
| Origi-nal | - | Low ‡ | - | - | - | - | - | - | - | - | Mixed § | - | Mixed § | - | - | Mixed § | |
| New | Low ‡ | Non-mutated | Mixed § | Non-mutated | |||||||||||||
| 21 | Cq value | 34.2 | |||||||||||||||
| Origi-nal | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | M1058L * | |
| New | M1058L * |
| Cat | Material | Commercial FCoV S Gene Mutation RT-qPCR According to the Original Algorithm | Commercial FCoV S Gene Mutation RT-qPCR According to the New Algorithm | Sanger Sequencing | Sequencing after Cloning |
|---|---|---|---|---|---|
| 5 | Feces | Mixed § | Non-mutated | Non-mutated | Non-mutated |
| 11 | Popliteal lymph node | M1058L * | M1058L * | Non-mutated | Non-mutated |
| 11 | Kidney | M1058L * | M1058L * | Non-mutated | |
| 11 | Ileum | M1058L * | M1058L * | Non-mutated | Non-mutated |
| 11 | Mesenteric lymph node | M1058L * | M1058L * | Non-mutated | |
| 13 | Feces | Mixed § | Mixed § | Non-mutated | Non-mutated |
| 14 | Kidney | M1058L * | M1058L * | Non-mutated | Non-mutated |
| 14 | Colon | M1058L * | M1058L * | Non-mutated | Non-mutated |
| 14 | Feces | M1058L * | M1058L * | Non-mutated | Non-mutated |
| 16 | Ileum | Mixed § | Non-mutated | Non-mutated | |
| 16 | Colon | M1058L * | M1058L * | Non-mutated | |
| 16 | Feces | M1058L * | M1058L * | Non-mutated | Non-mutated |
| 17 | Feces | Mixed § | Non-mutated | Non-mutated | |
| 18 | Feces | M1058L * | M1058L * | Non-mutated | |
| 19 | Feces | M1058L * | M1058L * | Non-mutated | |
| 20 | Feces | Mixed § | Non-mutated | Non-mutated |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Jähne, S.; Felten, S.; Bergmann, M.; Erber, K.; Matiasek, K.; Meli, M.L.; Hofmann-Lehmann, R.; Hartmann, K. Detection of Feline Coronavirus Variants in Cats without Feline Infectious Peritonitis. Viruses 2022, 14, 1671. https://doi.org/10.3390/v14081671
Jähne S, Felten S, Bergmann M, Erber K, Matiasek K, Meli ML, Hofmann-Lehmann R, Hartmann K. Detection of Feline Coronavirus Variants in Cats without Feline Infectious Peritonitis. Viruses. 2022; 14(8):1671. https://doi.org/10.3390/v14081671
Chicago/Turabian StyleJähne, Stéphanie, Sandra Felten, Michèle Bergmann, Katharina Erber, Kaspar Matiasek, Marina L. Meli, Regina Hofmann-Lehmann, and Katrin Hartmann. 2022. "Detection of Feline Coronavirus Variants in Cats without Feline Infectious Peritonitis" Viruses 14, no. 8: 1671. https://doi.org/10.3390/v14081671
APA StyleJähne, S., Felten, S., Bergmann, M., Erber, K., Matiasek, K., Meli, M. L., Hofmann-Lehmann, R., & Hartmann, K. (2022). Detection of Feline Coronavirus Variants in Cats without Feline Infectious Peritonitis. Viruses, 14(8), 1671. https://doi.org/10.3390/v14081671