
Plasma Membrane-Derived Liposomes Exhibit Robust Antiviral Activity against HSV-1



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Cells, Viruses, Media, Buffers and Antibodies
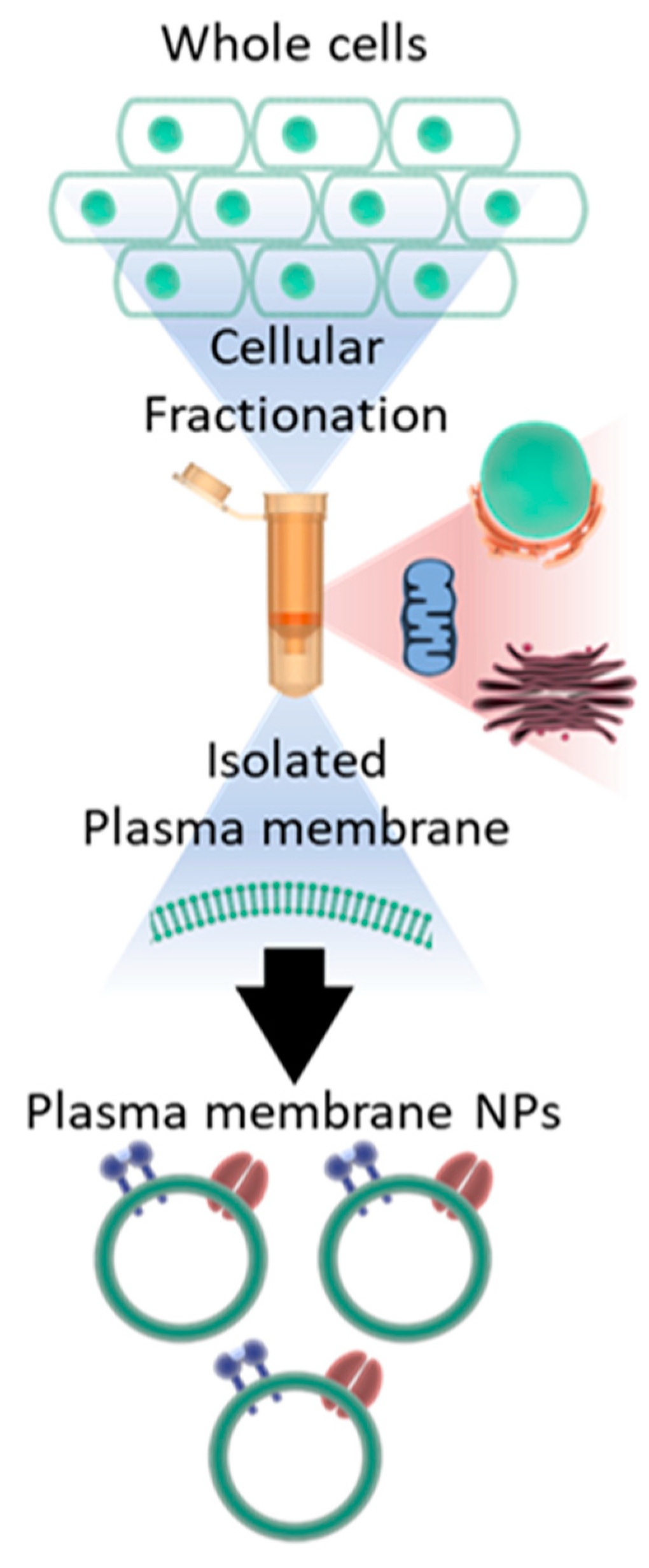
2.2. Plasma Membrane Isolation
2.2.1. Manual Isolation
2.2.2. Plasma Membrane Isolation Kit
2.3. Neutralization Assay
2.4. Viral Entry Assay
2.5. Transmission Electron Microscopy Imaging
2.6. Cell Viability Assay
2.7. Flow Cytometry
2.8. Western Blot
2.9. Plaque Assay
2.10. Quantitative PCR Assay
3. Results
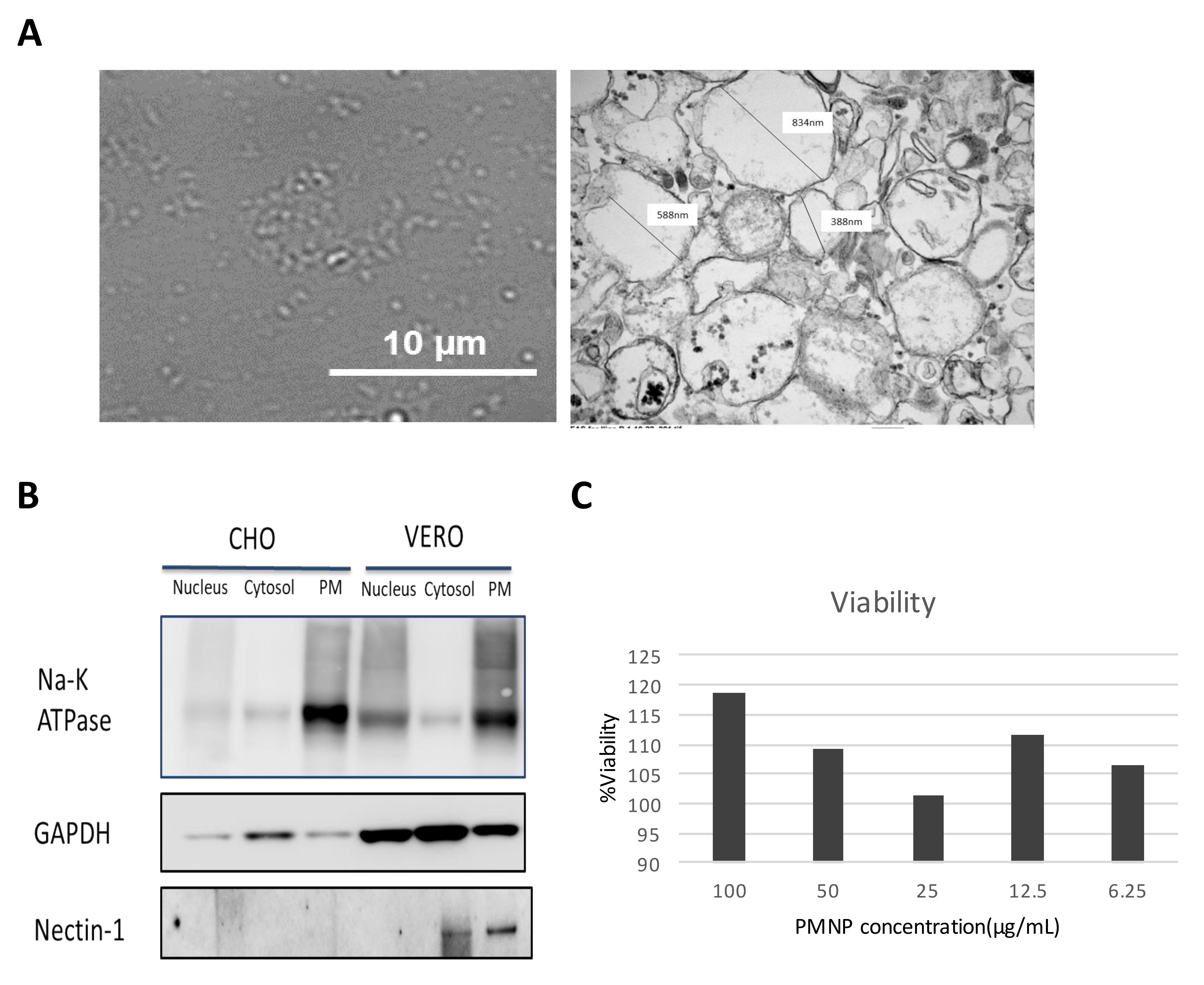
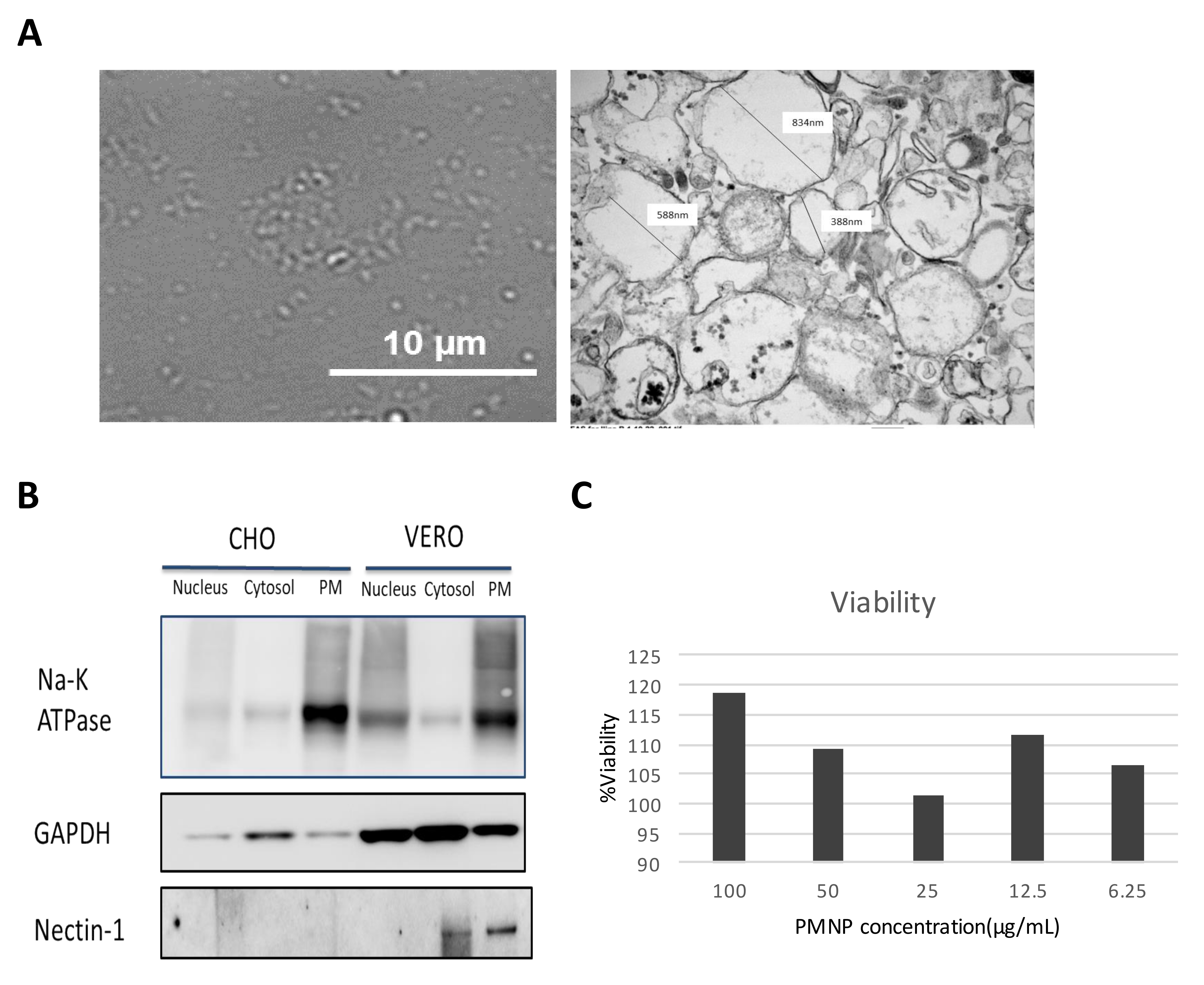
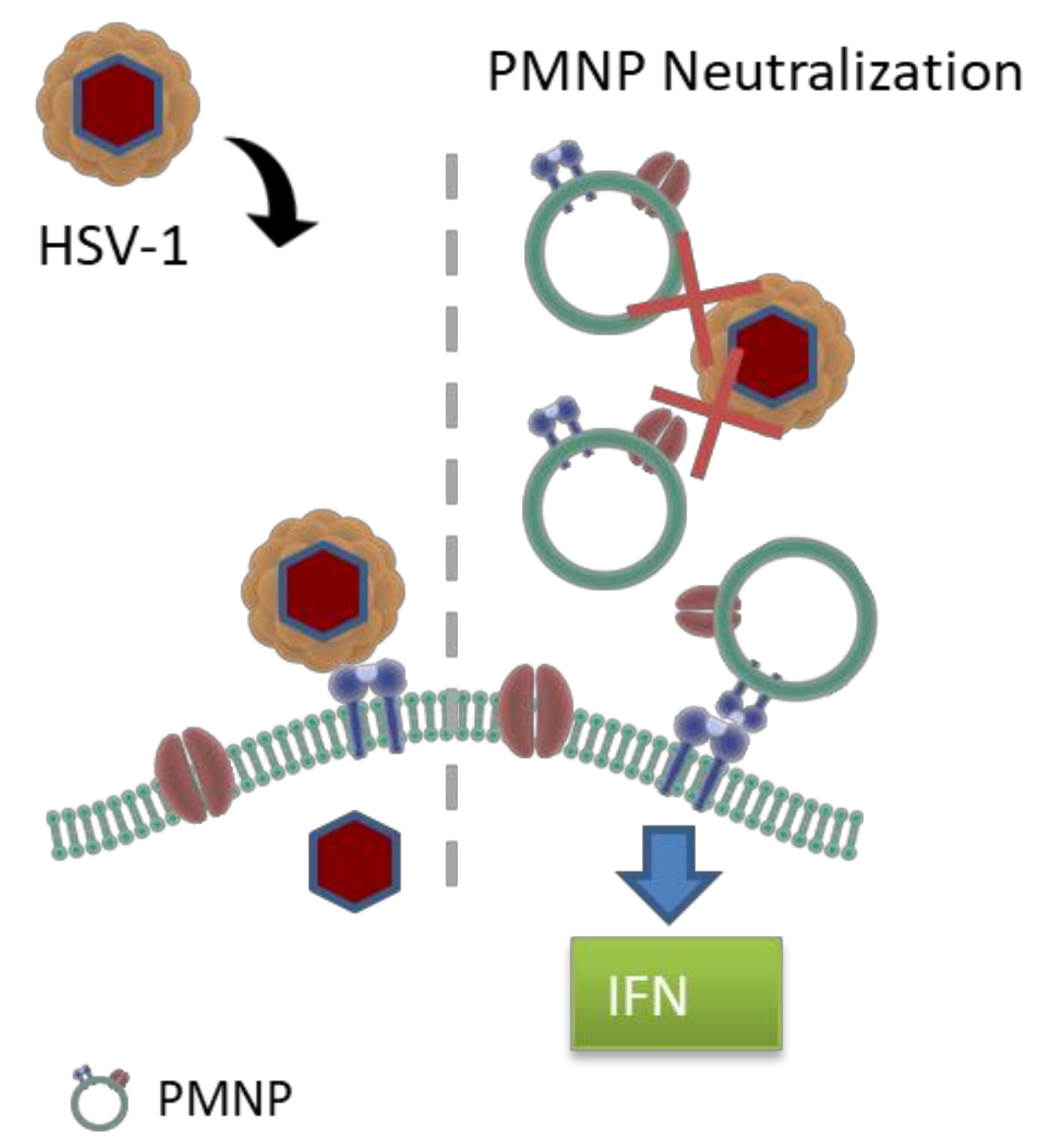
3.1. Preparation of Plasma Membrane Derived Neutralizing Particles (PMNP)
3.2. Characterization of Plasma Membrane Derive Nanoparticles (PMNP)
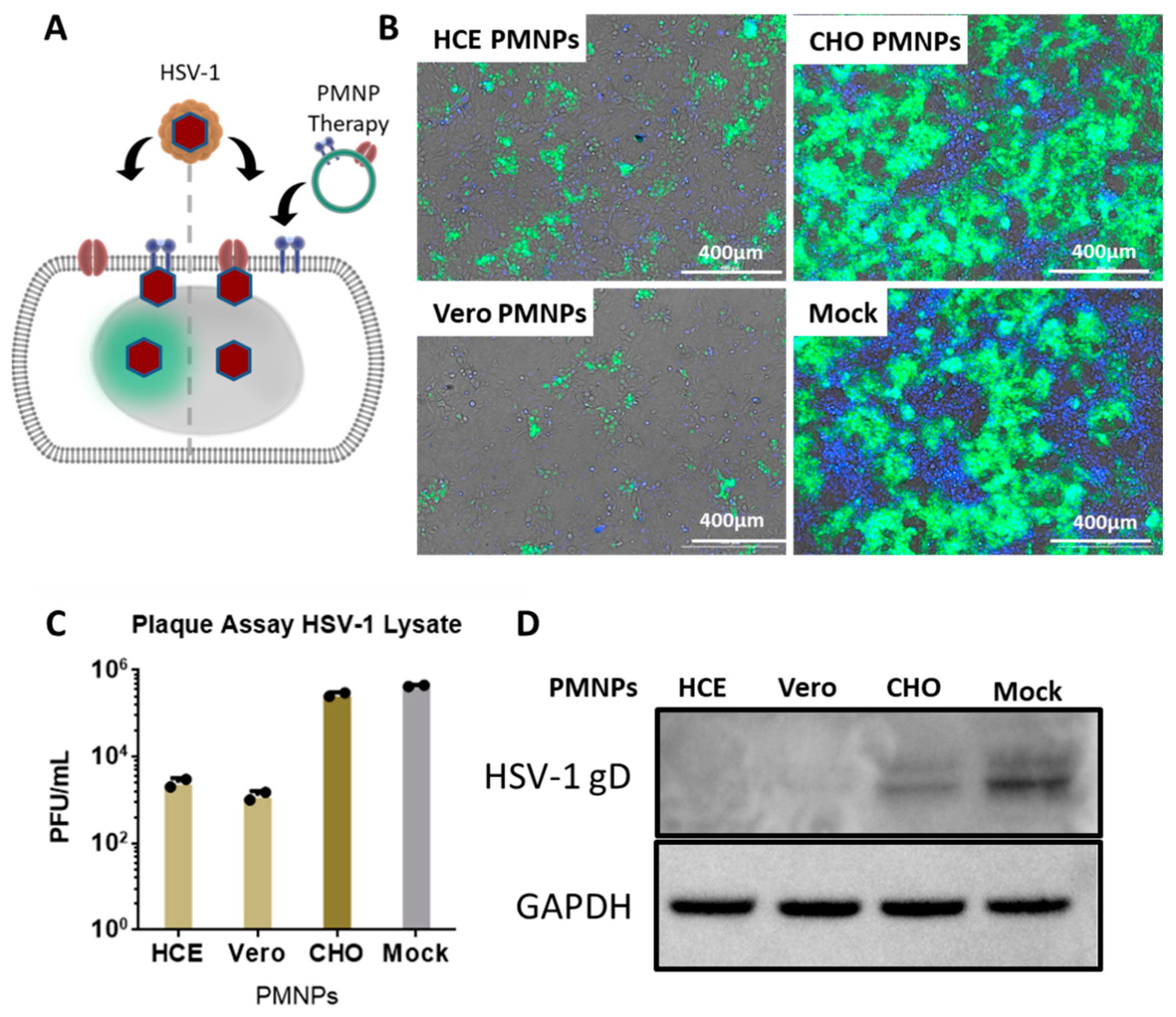
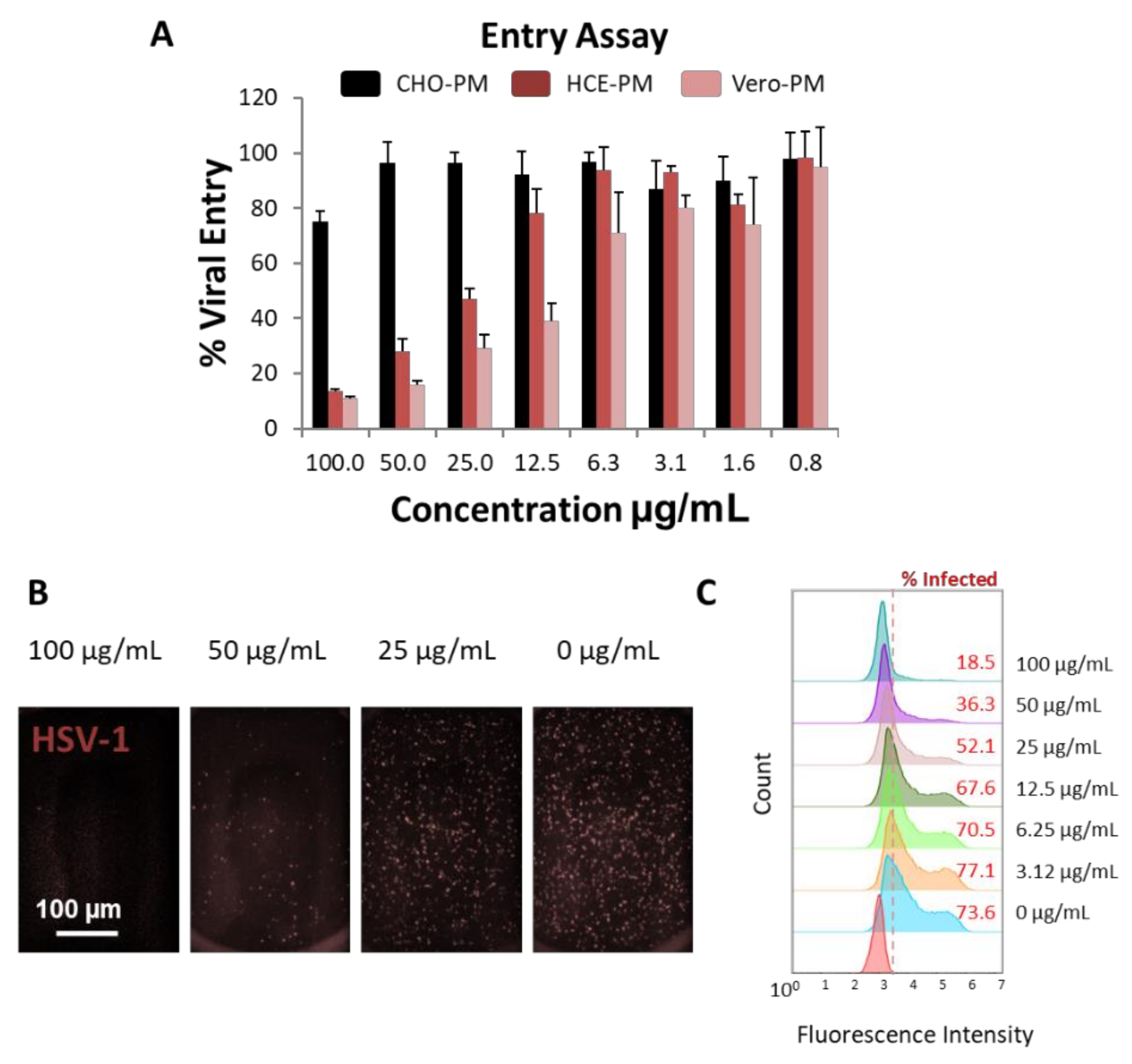
3.3. PMNPs Block Viral Entry into Target Cells
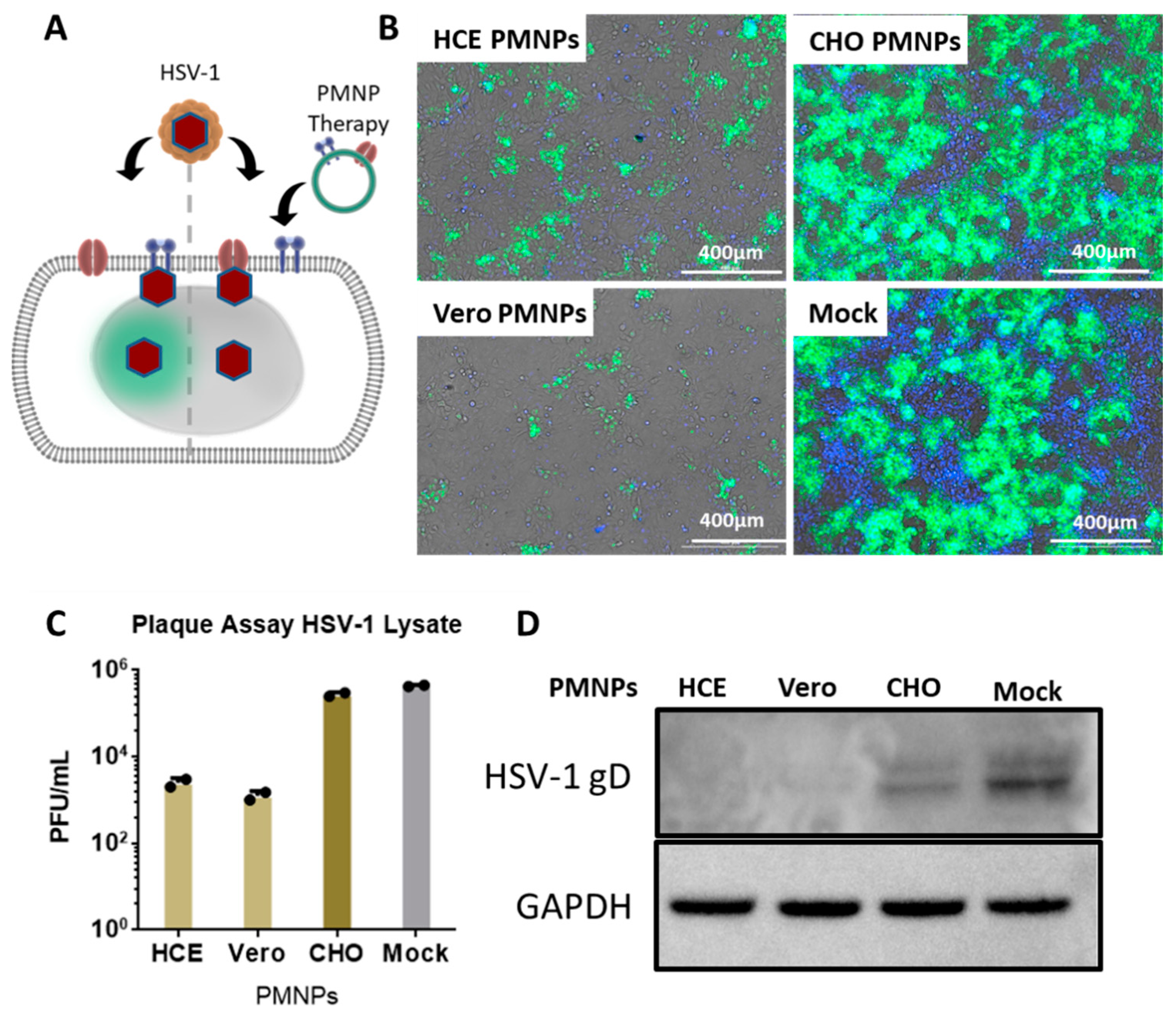
3.4. Therapeutic Treatment with PMNPs Protects Human Corneal Epithelial Cells from Viral Spread
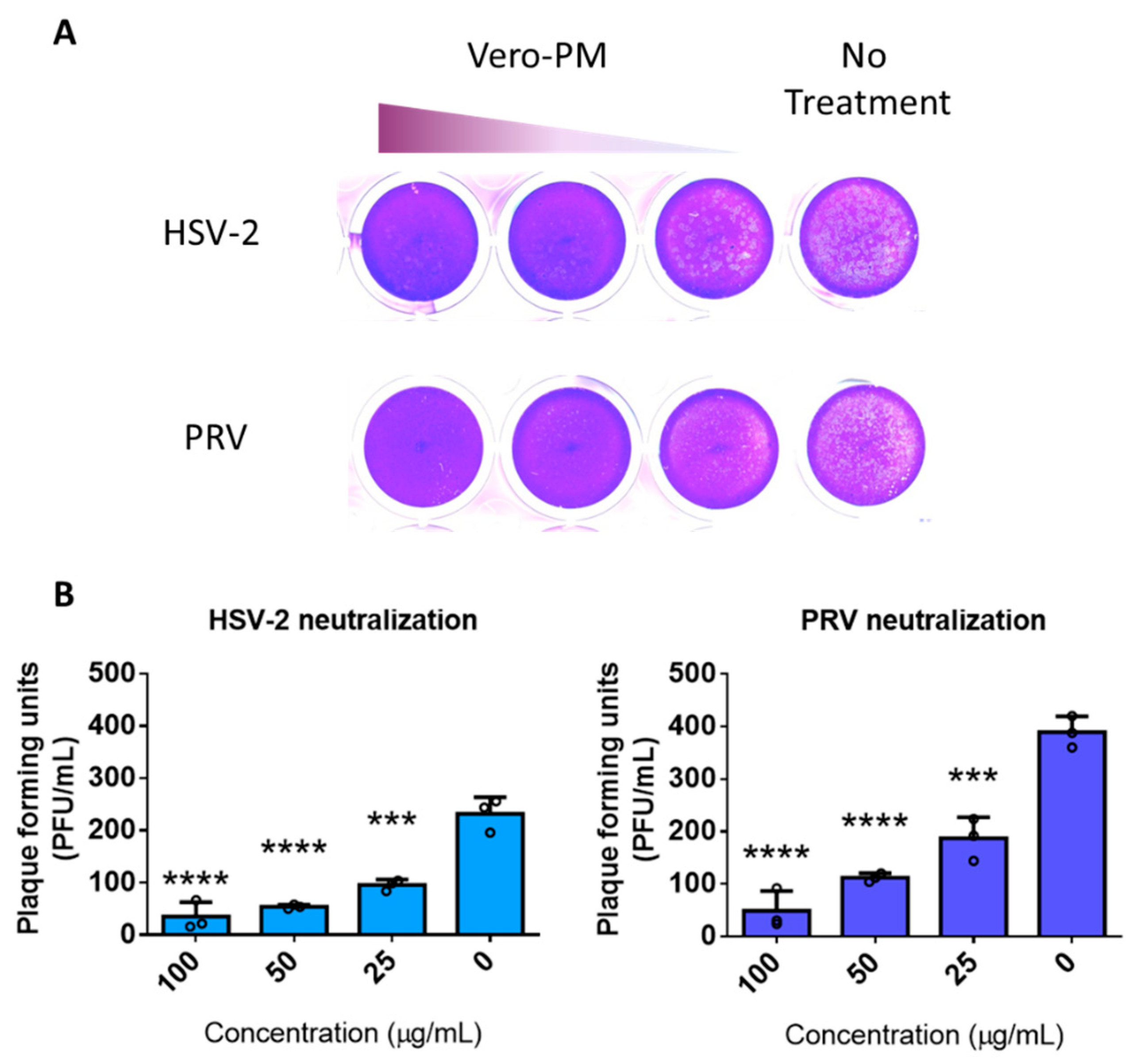
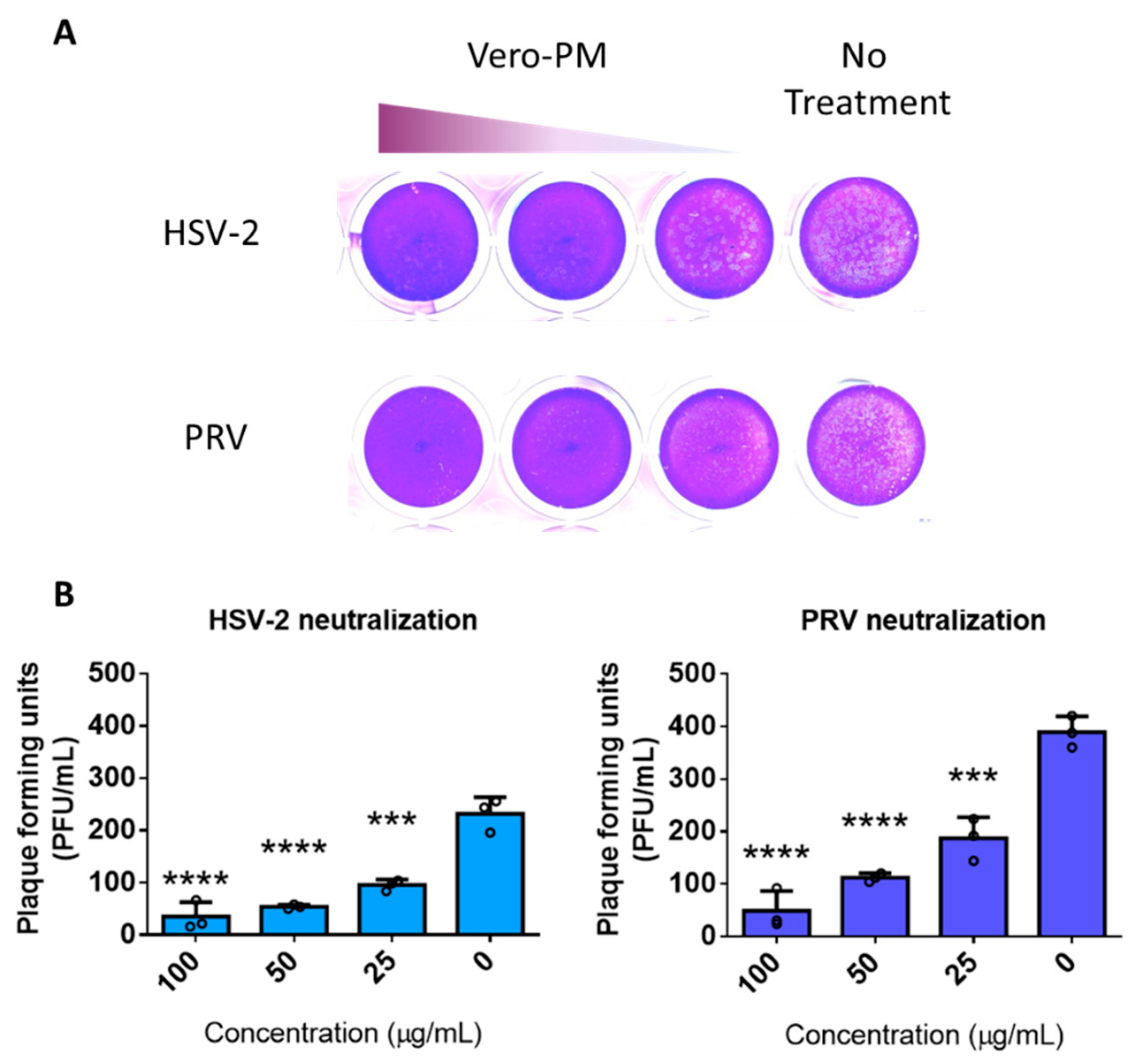
3.5. PMNPs Are Effective against Other Herpesviruses
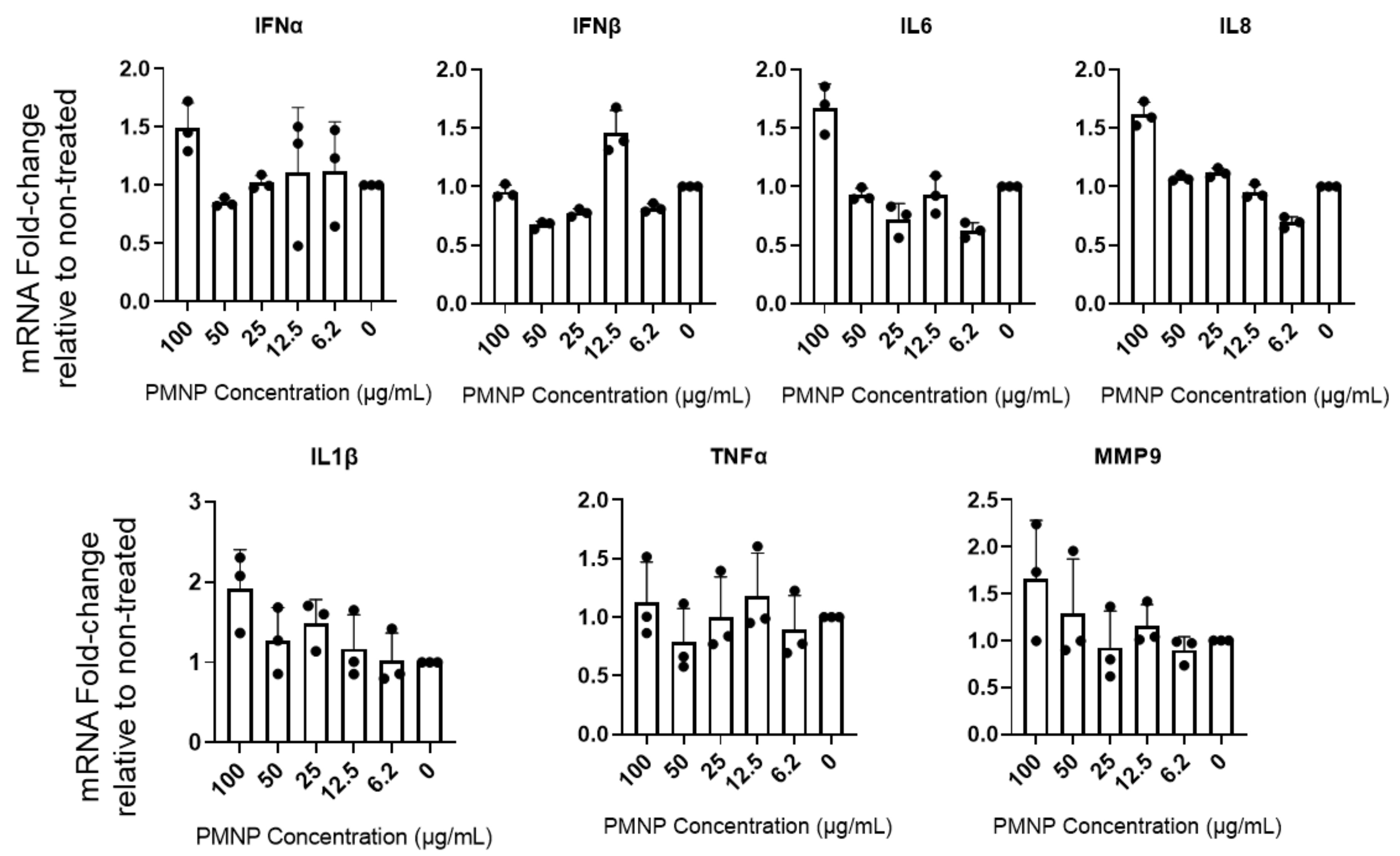
3.6. PMNPs Stimulate Interferon Response
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Conflicts of Interest
References
- Cooper, G.M. Structure of the Plasma Membrane. In The Cell: A Molecular Approach, 2nd ed.; ASM Press: Washington, DC, USA, 2000. Available online: https://www.ncbi.nlm.nih.gov/books/NBK9898/ (accessed on 2 April 2022).
- Thorley, J.; McKeating, J.; Rappoport, J. Mechanisms of Viral Entry: Sneaking in the Front Door. Protoplasma 2010, 244, 15–24. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Destache, C.J.; Mandal, S.; Yuan, Z.; Kang, G.; Date, A.A.; Lu, W.; Shibata, A.; Pham, R.; Bruck, P.; Rezich, M.; et al. Topical Tenofovir Disoproxil Fumarate Nanoparticles Prevent HIV-1 Vaginal Transmission in a Humanized Mouse Model. Antimicrob. Agents Chemother. 2016, 60, 3633–3639. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Madavaraju, K.; Koganti, R.; Volety, I.; Yadavalli, T.; Shukla, D. Herpes Simplex Virus Cell Entry Mechanisms: An Update. Front. Cell. Infect. Microbiol. 2020, 10, 617578. [Google Scholar] [CrossRef] [PubMed]
- Shang, J.; Wan, Y.; Luo, C.; Ye, G.; Geng, Q.; Auerbach, A.; Li, F. Cell entry mechanisms of SARS-CoV-2. Proc. Natl. Acad. Sci. USA 2020, 117, 11727–11734. [Google Scholar] [CrossRef]
- Coleman, C.M.; Liu, Y.V.; Mu, H.; Taylor, J.K.; Massare, M.; Flyer, D.C.; Smith, G.E.; Frieman, M.B. Purified coronavirus spike protein nanoparticles induce coronavirus neutralizing antibodies in mice. Vaccine 2014, 32, 3169–3174. [Google Scholar] [CrossRef]
- Fan, Y.; Cui, Y.; Hao, W.; Chen, M.; Liu, Q.; Wang, Y.; Yang, M.; Li, Z.; Gong, W.; Song, S.; et al. Carrier-Free Highly Drug-Loaded Biomimetic Nanosuspensions Encapsulated by Cancer Cell Membrane Based on Homology and Active Targeting for the Treatment of Glioma. Bioact. Mater. 2021, 6, 4402–4414. [Google Scholar] [CrossRef]
- Zhao, Q.; Sun, X.; Wu, B.; Shang, Y.; Huang, X.; Dong, H.; Liu, H.; Chen, W.; Gui, R.; Li, J. Construction of Homologous Cancer Cell Membrane Camouflage in a Nano-Drug Delivery System for the Treatment of Lymphoma. J. Nanobiotechnol. 2021, 19, 8. [Google Scholar] [CrossRef]
- Xia, Q.; Zhang, Y.; Li, Z.; Hou, X.; Feng, N. Red Blood Cell Membrane-Camouflaged Nanoparticles: A Novel Drug Delivery System for Antitumor Application. Acta Pharm. Sin. B 2019, 9, 675–689. [Google Scholar] [CrossRef]
- Zan, M.-H.; Rao, L.; Xie, W.; Zhu, D.-M.; Qie, X.-W.; Dong, W.-F.; Liu, W. Advances in Cell Membrane-Camouflaged Nano-Carrier for Photothermal Therapy. Chin. Opt. 2018, 11, 392–400. [Google Scholar] [CrossRef]
- Kyluik-Price, D.L.; Li, L.; Scott, M.D. Comparative Efficacy of Blood Cell Immunocamouflage by Membrane Grafting of Methoxypoly (Ethylene Glycol) and Polyethyloxazoline. Biomaterials 2014, 35, 412–422. [Google Scholar] [CrossRef]
- Hu, Q.; Bu, Y.; Cao, R.; Zhang, G.; Xie, X.; Wang, S. Stability Designs of Cell Membrane Cloaked Magnetic Carbon Nanotubes for Improved Life Span in Screening Drug Leads. Anal. Chem. 2019, 91, 13062–13070. [Google Scholar] [CrossRef]
- Spanjers, J.M.; Städler, B. Cell Membrane Coated Particles. Adv. Biosyst. 2020, 4, 2000174. [Google Scholar] [CrossRef]
- Liu, Z.; Wang, F.; Liu, X.; Sang, Y.; Zhang, L.; Ren, J.; Qu, X. Cell Membrane–Camouflaged Liposomes for Tumor Cell–Selective Glycans Engineering and Imaging In Vivo. Proc. Natl. Acad. Sci. USA 2021, 118, e2022769118. [Google Scholar] [CrossRef]
- Nishiguchi, A.; Taguchi, T. Development of an Immunosuppressive Camouflage-Coating Platform with Nanocellulose and Cell Membrane Vesicles. J. Biomater. Sci. Polym. Ed. 2020, 31, 1912–1924. [Google Scholar] [CrossRef]
- Shieh, M.T.; WuDunn, D.; Montgomery, R.I.; Esko, J.D.; Spear, P.G. Cell surface receptors for herpes simplex virus are heparan sulfate proteoglycans. J. Cell Biol. 1992, 116, 1273–1281. [Google Scholar] [CrossRef]
- Araki-Sasaki, K.; Ohashi, Y.; Sasabe, T.; Hayashi, K.; Watanabe, H.; Tano, Y.; Handa, H. An SV40-immortalized human corneal epithelial cell line and its characterization. Investig. Ophthalmol. Vis. Sci. 1995, 36, 614–621. [Google Scholar]
- Thompson, R.L.; Preston, C.M.; Sawtell, N.M. De Novo Synthesis of VP16 Coordinates the Exit from HSV Latency in Vivo. PLoS Pathog. 2009, 5, e1000352. [Google Scholar] [CrossRef]
- Jett, M.; Seed, T.M.; Jamieson, G.A. Isolation and characterization of plasma membranes and intact nuclei from lymphoid cells. J. Biol. Chem. 1977, 252, 2134–2142. [Google Scholar] [CrossRef]
- Yadavalli, T.; Agelidis, A.; Jaishankar, D.; Mangano, K.; Thakkar, N.; Penmetcha, K.; Shukla, D. Targeting Herpes Simplex Virus-1 gD by a DNA Aptamer Can Be an Effective New Strategy to Curb Viral Infection. Mol. Ther. Nucl. Acids 2017, 9, 365–378. [Google Scholar] [CrossRef] [Green Version]
- Montgomery, R.I.; Warner, M.S.; Lum, B.J.; Spear, P.G. Herpes Simplex Virus-1 Entry into Cells Mediated by a Novel Member of the TNF/NGF Receptor Family. Cell 1996, 87, 427–436. [Google Scholar] [CrossRef] [Green Version]
- Guo, D.; Zhu, Q.; Zhang, H.; Sun, D. Proteomic analysis of membrane proteins of vero cells: Exploration of potential proteins responsible for virus entry. DNA Cell Biol. 2014, 33, 20–28. [Google Scholar] [CrossRef] [Green Version]
- Luk, B.T.; Zhang, L. Cell membrane-camouflaged nanoparticles for drug delivery. J. Control. Release 2015, 220, 600–607. [Google Scholar] [CrossRef] [Green Version]
- Xie, M.; Deng, T.; Li, J.; Shen, H. The Camouflage of Graphene Oxide by Red Blood Cell Membrane with High Dispersibility for Cancer Chemotherapy. J. Colloid Interface Sci. 2021, 591, 290–299. [Google Scholar] [CrossRef]
- Li, R.; He, Y.; Zhang, S.; Qin, J.; Wang, J. Cell Membrane-Based Nanoparticles: A New Biomimetic Platform for Tumor Diagnosis and Treatment. Acta Pharm. Sin. B 2018, 8, 14–22. [Google Scholar] [CrossRef]
- Wu, M.; Mei, T.; Lin, C.; Wang, Y.; Chen, J.; Le, W.; Sun, M.; Xu, J.; Dai, H.; Zhang, Y.; et al. Melanoma Cell Membrane Biomimetic Versatile CuS Nanoprobes for Homologous Targeting Photoacoustic Imaging and Photothermal Chemotherapy. ACS Appl. Mater. Interfaces 2020, 12, 16031–16039. [Google Scholar] [CrossRef]
- Narain, A.; Asawa, S.; Chhabria, V.; Patil-Sen, Y. Cell Membrane Coated Nanoparticles: Next-Generation Therapeutics. Nanomedicine 2017, 12, 2677–2692. [Google Scholar] [CrossRef]
- Shang, Y.; Wang, Q.; Li, J.; Zhao, Q.; Huang, X.; Dong, H.; Liu, H.; Gui, R.; Nie, X. Platelet-Membrane-Camouflaged Zirconia Nanoparticles Inhibit the Invasion and Metastasis of Hela Cells. Front. Chem. 2020, 8, 377. [Google Scholar] [CrossRef]
- Zhai, Y.; Su, J.; Ran, W.; Zhang, P.; Yin, Q.; Zhang, Z.; Yu, H.; Li, Y. Preparation and Application of Cell Membrane-Camouflaged Nanoparticles for Cancer Therapy. Theranostics 2017, 7, 2575–2592. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; Wang, W.; Zhang, X.; Yao, H.; Wei, Z.; Li, X.; Mu, X.; Jiang, J.; Zhang, H. Seedless Preparation of Au Nanorods by Hydroquinone Assistant and Red Blood Cell Membrane Camouflage. RSC Adv. 2018, 8, 21316–21325. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Q.; Honko, A.; Zhou, J.; Gong, H.; Downs, S.N.; Vasquez, J.H.; Fang, R.H.; Gao, W.; Griffiths, A.; Zhang, L. Cellular Nanosponges Inhibit SARS-CoV-2 Infectivity. Nano Lett. 2020, 20, 5570–5574. [Google Scholar] [CrossRef] [PubMed]
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bhattacharya, I.; Yadavalli, T.; Wu, D.; Shukla, D. Plasma Membrane-Derived Liposomes Exhibit Robust Antiviral Activity against HSV-1. Viruses 2022, 14, 799. https://doi.org/10.3390/v14040799
Bhattacharya I, Yadavalli T, Wu D, Shukla D. Plasma Membrane-Derived Liposomes Exhibit Robust Antiviral Activity against HSV-1. Viruses. 2022; 14(4):799. https://doi.org/10.3390/v14040799
Chicago/Turabian StyleBhattacharya, Ilina, Tejabhiram Yadavalli, David Wu, and Deepak Shukla. 2022. "Plasma Membrane-Derived Liposomes Exhibit Robust Antiviral Activity against HSV-1" Viruses 14, no. 4: 799. https://doi.org/10.3390/v14040799
APA StyleBhattacharya, I., Yadavalli, T., Wu, D., & Shukla, D. (2022). Plasma Membrane-Derived Liposomes Exhibit Robust Antiviral Activity against HSV-1. Viruses, 14(4), 799. https://doi.org/10.3390/v14040799