
Geographical and Tick-Dependent Distribution of Flavi-Like Alongshan and Yanggou Tick Viruses in Russia

 ,
,  ,
,  , , , ,
, , , ,  , ,
, ,  ,
,  ,
,
Abstract
1. Introduction
2. Materials and Methods
2.1. Collection and Processing of Mosquitoes
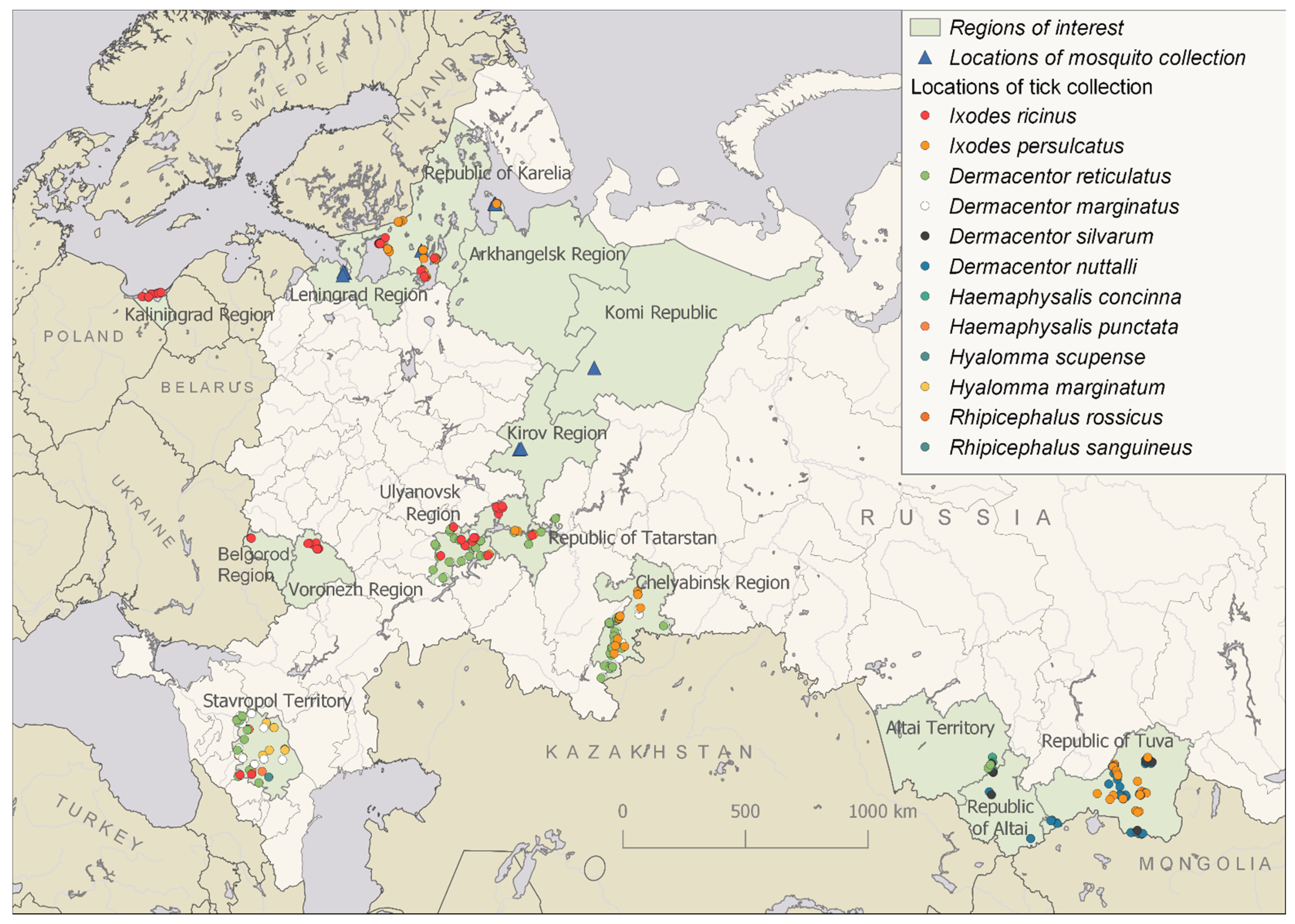
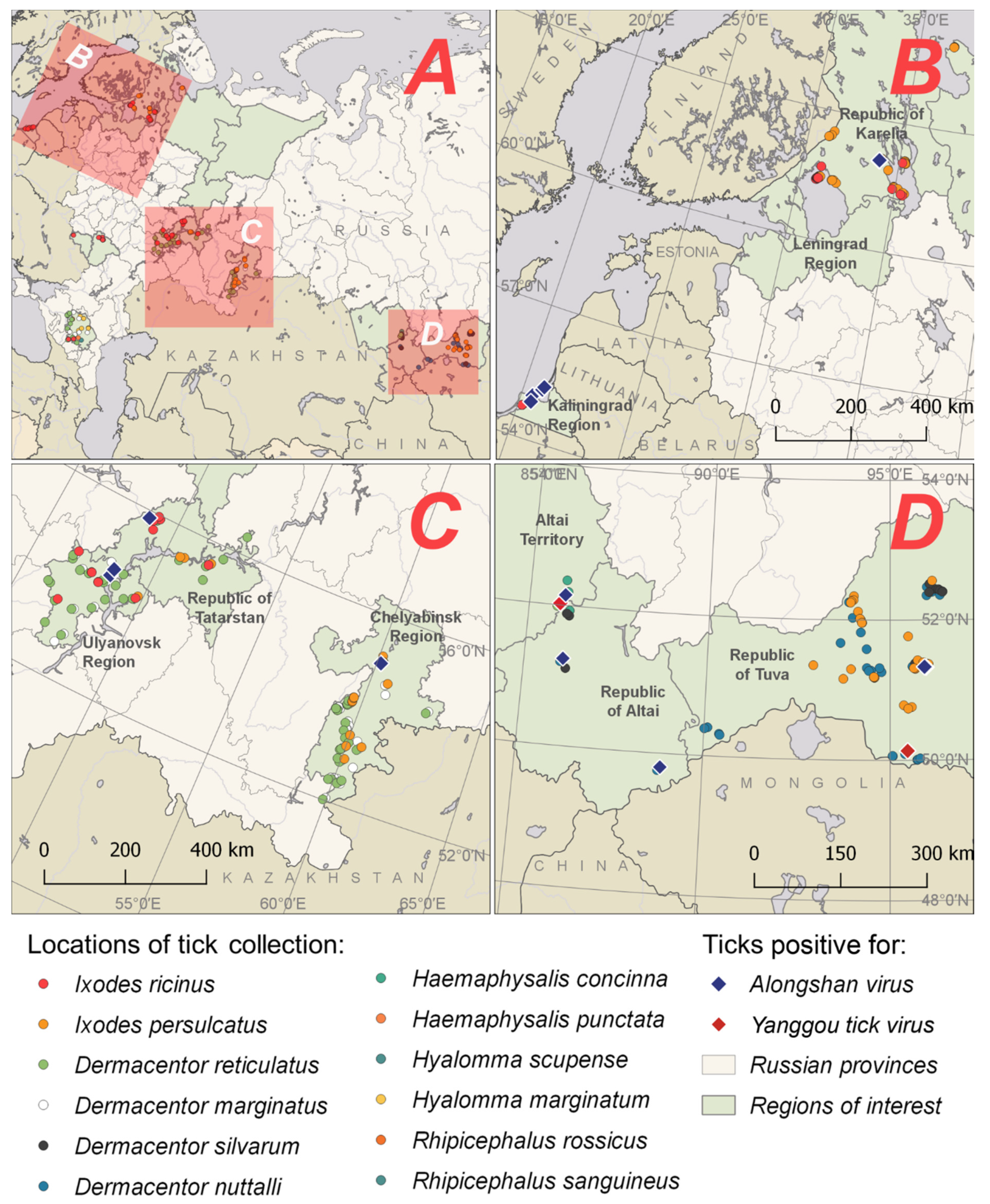
2.2. Collection and Processing of Ticks
2.3. Infection of Tick Cell Lines
2.4. Reverse-Transcriptase PCR (RT-PCR) and Sequencing of Amplified Products
2.5. High-Throughput Sequencing
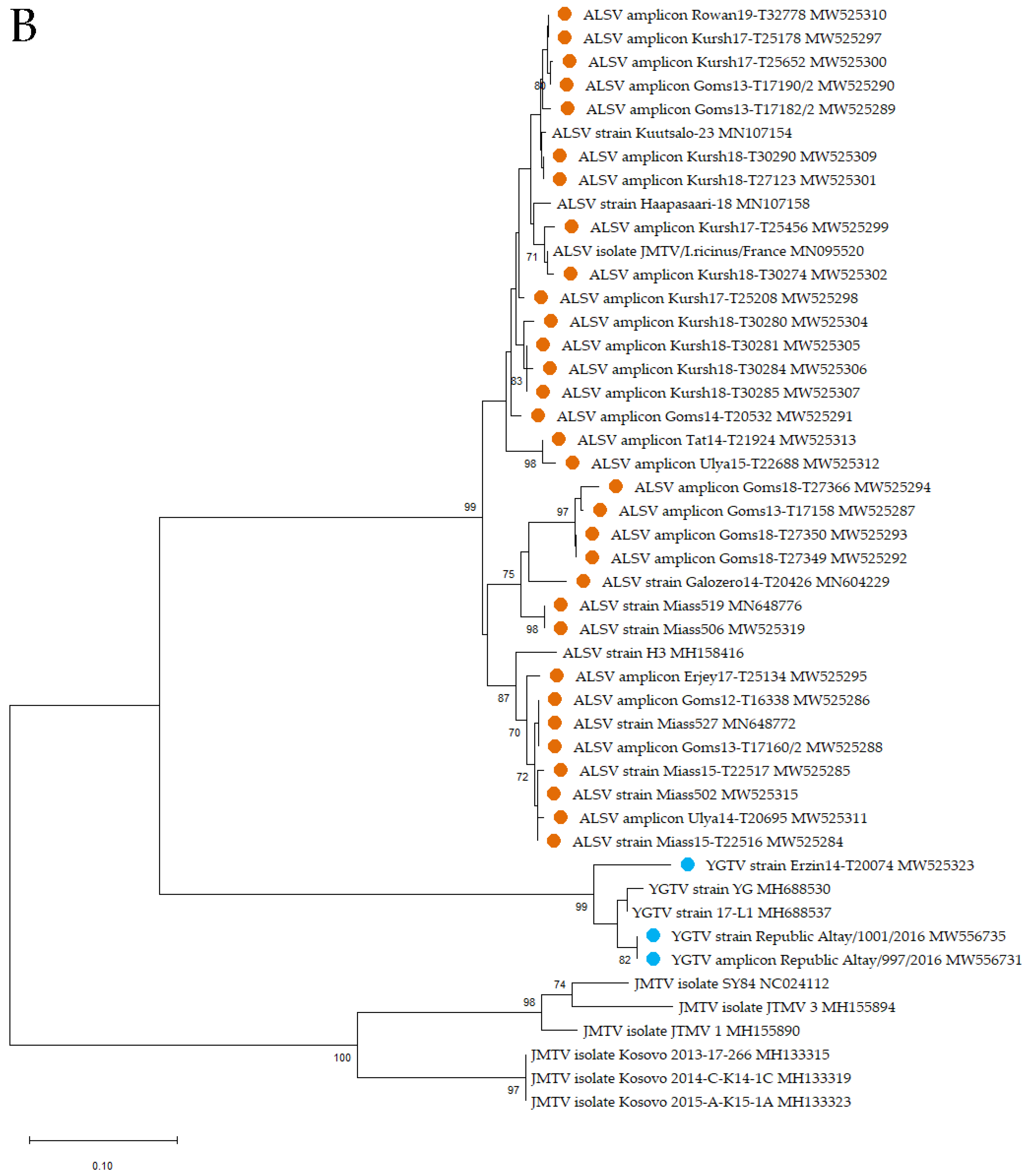
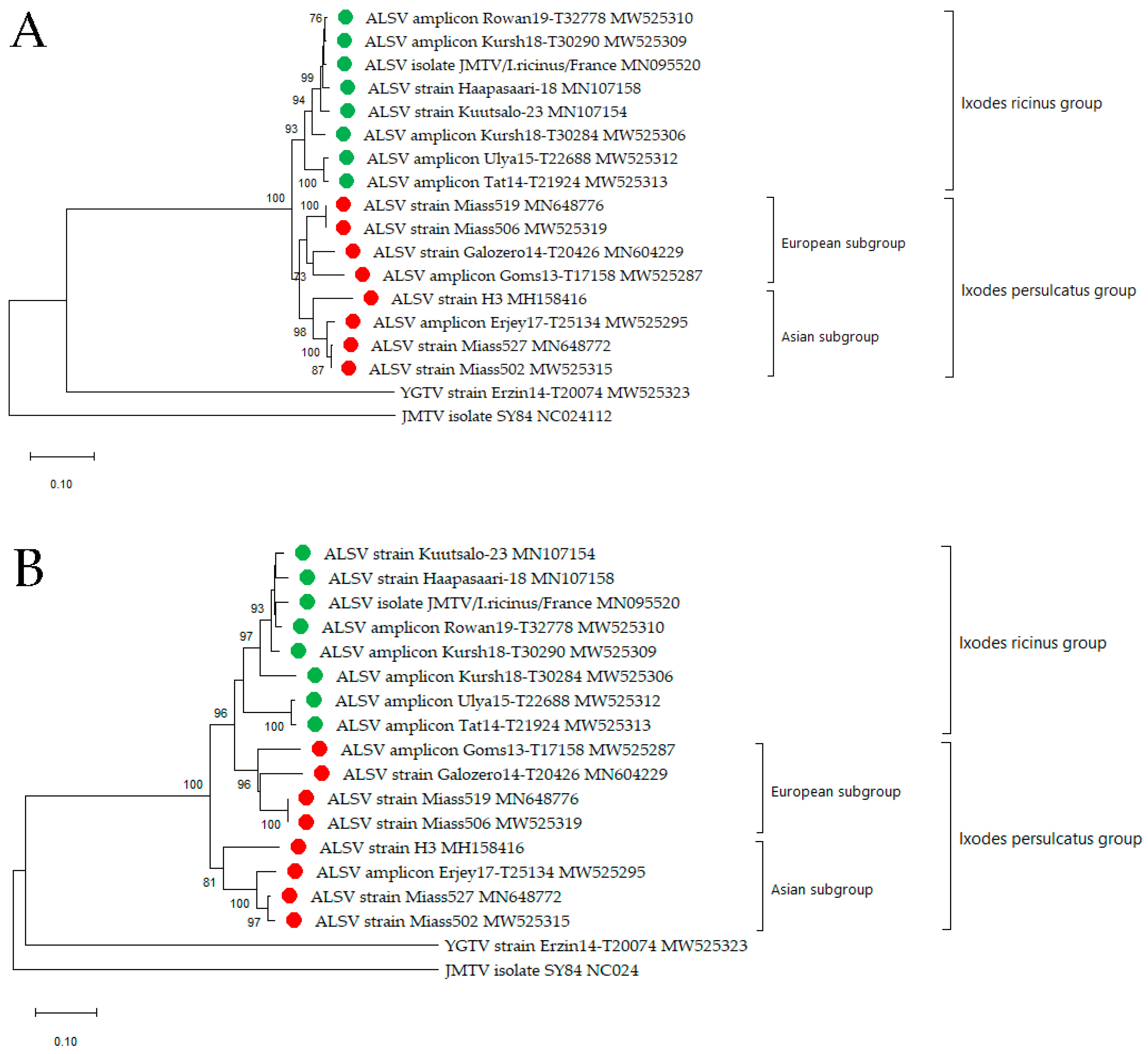
2.6. Phylogenetic Analysis
3. Results
3.1. Collection and Screening for Flavi-Like Viruses of Mosquitoes
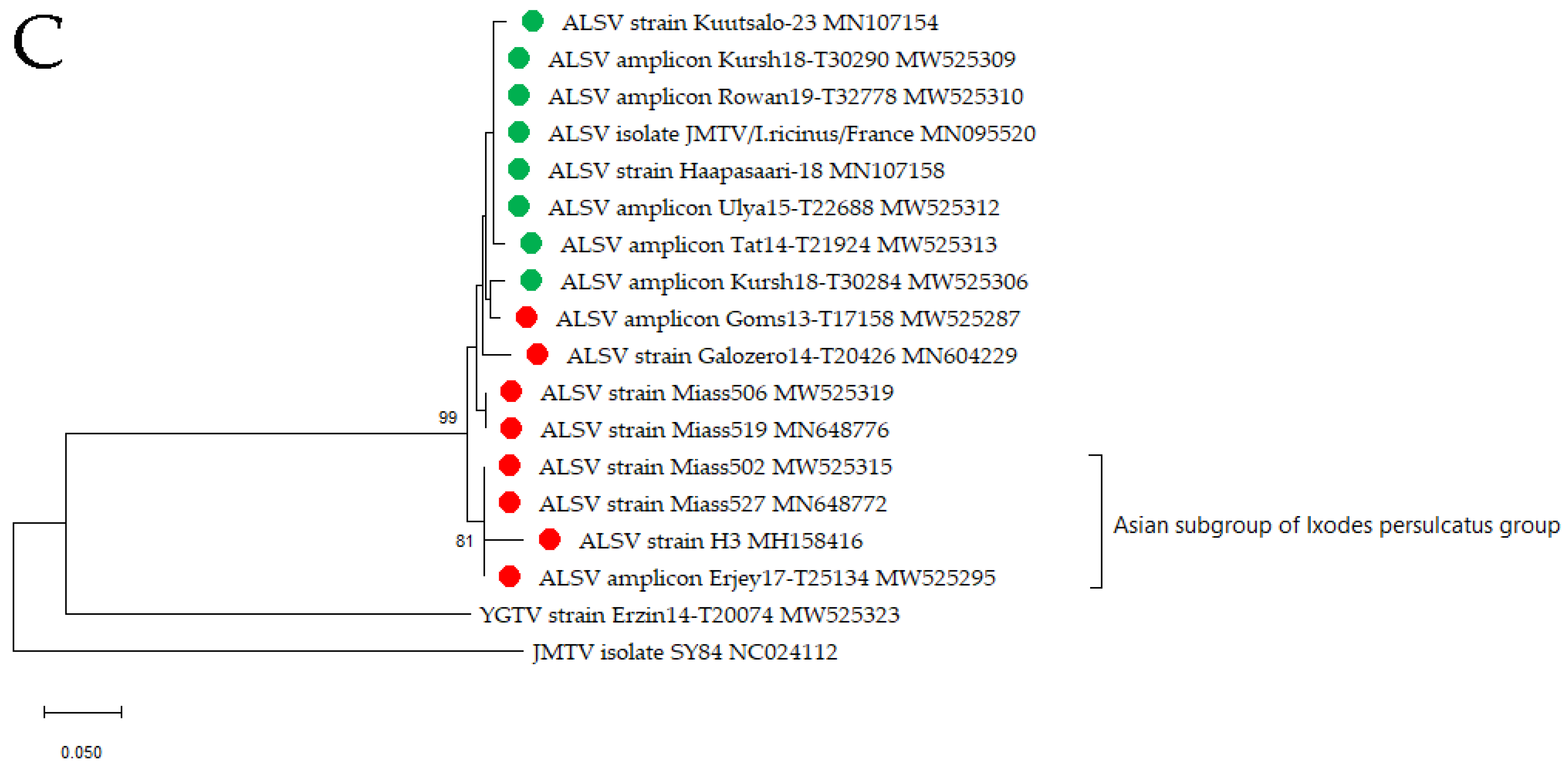
3.2. Alongshan Virus Detection in Ticks, Isolation and Phylogenetic Relationships
3.3. Yanggou Tick Virus Detection, Isolation and Phylogenetic Relationships
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Simmonds, P.; Becher, P.; Bukh, J.; Gould, E.A.; Meyers, G.; Monath, T.; Muerhoff, S.; Pletnev, A.; Rico-Hesse, R.; Smith, D.B.; et al. ICTV virus taxonomy profile: Flaviviridae. J. Gen. Virol. 2017, 98, 2–3. [Google Scholar] [CrossRef] [PubMed]
- Shi, M.; Lin, X.-D.; Vasilakis, N.; Tian, J.-H.; Li, C.-X.; Chen, L.-J.; Eastwood, G.; Diao, X.-N.; Chen, M.-H.; Chen, X.; et al. Divergent Viruses Discovered in Arthropods and Vertebrates Revise the Evolutionary History of the Flaviviridae and Related Viruses. J. Virol. 2016, 90, 659–669. [Google Scholar] [CrossRef]
- Qin, X.-C.; Shi, M.; Tian, J.-H.; Lin, X.-D.; Gao, D.-Y.; He, J.-R.; Wang, J.-B.; Li, C.-X.; Kang, Y.-J.; Yu, B.; et al. A tick-borne segmented RNA virus contains genome segments derived from unsegmented viral ancestors. Proc. Natl. Acad. Sci. USA 2014, 111, 6744–6749. [Google Scholar] [CrossRef] [PubMed]
- International Committee on Taxonomy of Viruses Genus: Flavivirus. Available online: https://talk.ictvonline.org/ictv-reports/ictv_online_report/positive-sense-rna-viruses/w/flaviviridae/360/genus-flavivirus (accessed on 27 January 2021).
- Dinçer, E.; Hacıoğlu, S.; Kar, S.; Emanet, N.; Brinkmann, A.; Nitsche, A.; Özkul, A.; Linton, Y.-M.; Ergünay, K. Survey and Characterization of Jingmen Tick Virus Variants. Viruses 2019, 11, 1071. [Google Scholar] [CrossRef]
- Garry, C.E.; Garry, R.F. Proteomics Computational Analyses Suggest that the Envelope Glycoproteins of Segmented Jingmen Flavi-Like Viruses Are Class II Viral Fusion Proteins (β-Penetrenes) with Mucin-Like Domains. Viruses 2020, 12, 260. [Google Scholar] [CrossRef] [PubMed]
- Kholodilov, I.S.; Litov, A.G.; Klimentov, A.S.; Belova, O.A.; Polienko, A.E.; Nikitin, N.A.; Shchetinin, A.M.; Ivannikova, A.Y.; Bell-Sakyi, L.; Yakovlev, A.S.; et al. Isolation and characterisation of Alongshan virus in Russia. Viruses 2020, 12, 362. [Google Scholar] [CrossRef]
- Gordon, K.H.J.; Williams, M.R.; Hendry, D.A.; Hanzlik, T.N. Sequence of the genomic RNA of Nudaurelia β virus (Tetraviridae) defines a novel virus genome organization. Virology 1999, 258, 42–53. [Google Scholar] [CrossRef]
- Yong, C.Y.; Yeap, S.K.; Omar, A.R.; Tan, W.S. Advances in the study of nodavirus. PeerJ 2017, 2017, 1–31. [Google Scholar] [CrossRef] [PubMed]
- Jia, N.; Liu, H.B.; Ni, X.B.; Bell-Sakyi, L.; Zheng, Y.C.; Song, J.L.; Li, J.; Jiang, B.G.; Wang, Q.; Sun, Y.; et al. Emergence of human infection with Jingmen tick virus in China: A retrospective study. EBioMedicine 2019, 43, 317–324. [Google Scholar] [CrossRef] [PubMed]
- Temmam, S.; Bigot, T.; Chrétien, D.; Gondard, M.; Pérot, P.; Pommelet, V.; Dufou, E.; Petres, S.; Devillers, E.; Hoem, T.; et al. Insights into the Host Range, Genetic Diversity, and Geographical Distribution of Jingmenviruses. mSphere 2019, 4, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Emmerich, P.; Jakupi, X.; von Possel, R.; Berisha, L.; Halili, B.; Günther, S.; Cadar, D.; Ahmeti, S.; Schmidt-Chanasit, J. Viral metagenomics, genetic and evolutionary characteristics of Crimean-Congo hemorrhagic fever orthonairovirus in humans, Kosovo. Infect. Genet. Evol. 2018, 65, 6–11. [Google Scholar] [CrossRef]
- Kuivanen, S.; Levanov, L.; Kareinen, L.; Sironen, T.; Jääskeläinen, A.J.; Plyusnin, I.; Zakham, F. Detection of novel tick-borne pathogen, Alongshan virus, in Ixodes ricinus ticks, south-eastern Finland. Eurosurveillance 2019, 24, 1900394. [Google Scholar] [CrossRef] [PubMed]
- Ladner, J.T.; Wiley, M.R.; Beitzel, B.; Auguste, A.J.; Dupuis, A.P.; Lindquist, M.E.; Sibley, S.D.; Kota, K.P.; Fetterer, D.; Eastwood, G.; et al. A Multicomponent Animal Virus Isolated from Mosquitoes. Cell Host Microbe 2016, 20, 357–367. [Google Scholar] [CrossRef] [PubMed]
- Maruyama, S.R.; Castro-Jorge, L.A.; Ribeiro, J.M.C.; Gardinassi, L.G.; Garcia, G.R.; Brandão, L.G.; Rodrigues, A.R.; Okada, M.I.; Abrão, E.P.; Ferreira, B.R.; et al. Characterisation of divergent flavivirus NS3 and NS5 protein sequences detected in Rhipicephalus microplus ticks from Brazil. Mem. Inst. Oswaldo Cruz 2014, 109, 38–50. [Google Scholar] [CrossRef] [PubMed]
- Villa, E.C.; Maruyama, S.R.; de Miranda-Santos, I.K.F.; Palacios, G.; Ladner, J.T. Complete Coding Genome Sequence for Mogiana Tick Virus, a Jingmenvirus Isolated from Ticks in Brazil. Genome Announc. 2017, 5, 17–18. [Google Scholar] [CrossRef]
- De Pascoal, J.O.; de Siqueira, S.M.; da Maia, R.C.; Szabó, M.P.J.; Yokosawa, J. Detection and molecular characterization of Mogiana tick virus (MGTV) in Rhipicephalus microplus collected from cattle in a savannah area, Uberlândia, Brazil. Ticks Tick. Borne. Dis. 2019, 10, 162–165. [Google Scholar] [CrossRef]
- Vandegrift, K.J.; Kumar, A.; Sharma, H.; Murthy, S.; Kramer, L.D.; Ostfeld, R.; Hudson, P.J.; Kapoor, A. Presence of segmented flavivirus infections in North America. Emerg. Infect. Dis. 2020, 26, 1810–1817. [Google Scholar] [CrossRef]
- Webster, C.L.; Waldron, F.M.; Robertson, S.; Crowson, D.; Ferrari, G.; Quintana, J.F.; Brouqui, J.M.; Bayne, E.H.; Longdon, B.; Buck, A.H.; et al. The discovery, distribution, and evolution of viruses associated with Drosophila melanogaster. PLoS Biol. 2015, 13, e1002210. [Google Scholar] [CrossRef]
- Wang, Z.-D.; Wang, B.; Wei, F.; Han, S.-Z.; Zhang, L.; Yang, Z.-T.; Yan, Y.; Lv, X.-L.; Li, L.; Wang, S.-C.; et al. A New Segmented Virus Associated with Human Febrile Illness in China. N. Engl. J. Med. 2019, 380, 2116–2125. [Google Scholar] [CrossRef]
- De Souza, W.M.; Fumagalli, M.J.; de Carrasco, A.O.T.; Romeiro, M.F.; Modha, S.; Seki, M.C.; Gheller, J.M.; Daffre, S.; Nunes, M.R.T.; Murcia, P.R.; et al. Viral diversity of Rhipicephalus microplus parasitizing cattle in southern Brazil. Sci. Rep. 2018, 8, 1–10. [Google Scholar] [CrossRef]
- Wang, Z.-D.; Wang, W.; Wang, N.N.; Qiu, K.; Zhang, X.; Tana, G.; Liu, Q. Prevalence of the emerging novel Alongshan virus infection in sheep and cattle in Inner Mongolia, northeastern China. Parasit. Vectors 2019, 12, 1–7. [Google Scholar] [CrossRef]
- Gutsevich, A.V.; Monchadskii, A.S.; Shtakel’berg, A.A. Mosquitoes Family Culicidae. In Fauna of the U.S.S.R. Diptera; No. 4.; Bykhovsky, B.E., Ed.; Nauka: Leningrad, Russia, 1970; Volume 3, p. 387. [Google Scholar]
- Rasnitsyn, S.P. Sampling methods for bloodsucking dipterans. In Guide to Medical Entomology; Derbeneva-Ukhova, V.P., Ed.; Meditsina: Moscow, Russia, 1974; pp. 163–176. [Google Scholar]
- Khalin, A.V.; Aibulatov, S.V. New investigation technique for thoracic sclerites of mosquitoes (Diptera, Culicidae) for the correct identification of genera and species. Entomol. Rev. 2012, 92, 988–993. [Google Scholar] [CrossRef]
- Becker, N.; Petrić, D.; Zgomba, M.; Boase, C.; Madon, M.; Dahl, C.; Kaiser, A. Mosquitoes and Their Control, 2nd ed.; Springer: Berlin/Heidelberg, Germany, 2010; ISBN 978-3-540-92873-7. [Google Scholar]
- Khalin, A.V. Refinements of a key to species of the genus Aedes Meigen (Diptera, Culicidae) from northwestern Russia based on the male genitalia characters. Entomol. Rev.. 2009, 89, 1137–1153. [Google Scholar] [CrossRef]
- Filippova, N.A. Ixodid Ticks of the Subfamily Ixodinae. Fauna of the USSR: Arachnoides; Nauka: Leningrad, Russia, 1977; Volume 4. [Google Scholar]
- Filippova, N.A. Fauna of Russia and Neighbouring Countries. Ixodid Ticks of Subfamily Amblyomminae. Arachnoidea; Nauka: Saint Petersburg, Russia, 1997; Volume 4. [Google Scholar]
- Bell-Sakyi, L.; Zweygarth, E.; Blouin, E.F.; Gould, E.A.; Jongejan, F. Tick cell lines: Tools for tick and tick-borne disease research. Trends Parasitol. 2007, 23, 450–457. [Google Scholar] [CrossRef]
- Bell-Sakyi, L. Continuous cell lines from the tick Hyalomma anatolicum anatolicum. J. Parasitol. 1991, 77, 1006–1008. [Google Scholar] [CrossRef] [PubMed]
- Belova, O.A.; Litov, A.G.; Kholodilov, I.S.; Kozlovskaya, L.I.; Bell-Sakyi, L.; Romanova, L.I.; Karganova, G.G. Properties of the tick-borne encephalitis virus population during persistent infection of ixodid ticks and tick cell lines. Ticks Tick. Borne. Dis. 2017, 8, 895–906. [Google Scholar] [CrossRef] [PubMed]
- Scaramozzino, N.; Crance, J.-M.; Jouan, A.; DeBriel, D.A.; Stoll, F.; Garin, D. Comparison of Flavivirus universal primer pairs and development of a rapid, highly sensitive heminested reverse transcription-PCR assay for detection of flaviviruses targeted to a conserved region of the NS5 gene sequences. J. Clin. Microbiol. 2001, 39, 1922–1927. [Google Scholar] [CrossRef] [PubMed]
- Bolger, A.M.; Lohse, M.; Usadel, B. Trimmomatic: A flexible trimmer for Illumina sequence data. Bioinformatics 2014, 30, 2114–2120. [Google Scholar] [CrossRef]
- Okonechnikov, K.; Golosova, O.; Fursov, M.; Varlamov, A.; Vaskin, Y.; Efremov, I.; German Grehov, O.G.; Kandrov, D.; Rasputin, K.; Syabro, M.; et al. Unipro UGENE: A unified bioinformatics toolkit. Bioinformatics 2012, 28, 1166–1167. [Google Scholar] [CrossRef]
- Rubel, F.; Brugger, K.; Pfeffer, M.; Chitimia-Dobler, L.; Didyk, Y.M.; Leverenz, S.; Dautel, H.; Kahl, O. Geographical distribution of Dermacentor marginatus and Dermacentor reticulatus in Europe. Ticks Tick. Borne. Dis. 2016, 7, 224–233. [Google Scholar] [CrossRef]
- Nosek, J.; Kozuch, O. Replication of tick-borne encephalitis (TBE) virus in ticks Dermacentor marginatus. Angew. Parasitol. 1985, 26, 97–101. [Google Scholar] [PubMed]
- Ličková, M.; Havlíková, S.F.; Sláviková, M.; Slovák, M.; Drexler, J.F.; Klempa, B. Dermacentor reticulatus is a vector of tick-borne encephalitis virus. Ticks Tick. Borne. Dis. 2020, 11, 101414. [Google Scholar] [CrossRef] [PubMed]
- Romanova, L.I.; Gmyl, A.P.; Dzhivanian, T.I.; Bakhmutov, D.V.; Lukashev, A.N.; Gmyl, L.V.; Rumyantsev, A.A.; Burenkova, L.A.; Lashkevich, V.A.; Karganova, G.G. Microevolution of tick-borne encephalitis virus in course of host alternation. Virology 2007, 362, 75–84. [Google Scholar] [CrossRef]
- Bell-Sakyi, L.; Kohl, A.; Bente, D.A.; Fazakerley, J.K. Tick cell lines for study of crimean-congo hemorrhagic fever virus and other arboviruses. Vector Borne Zoonotic Dis. 2012, 12, 769–781. [Google Scholar] [CrossRef] [PubMed]
- Kholodilov, I.; Belova, O.; Burenkova, L.; Korotkov, Y.; Romanova, L.; Morozova, L.; Kudriavtsev, V.; Gmyl, L.; Belyaletdinova, I.; Chumakov, A.; et al. Ixodid ticks and tick-borne encephalitis virus prevalence in the South Asian part of Russia (Republic of Tuva). Ticks Tick. Borne. Dis. 2019, 10, 959–969. [Google Scholar] [CrossRef]
- Shchuchinova, L.D.; Kozlova, I.V.; Zlobin, V.I. Influence of altitude on tick-borne encephalitis infection risk in the natural foci of the Altai Republic, Southern Siberia. Ticks Tick. Borne. Dis. 2015, 6, 322–329. [Google Scholar] [CrossRef]
- Korenberg, E.I.; Zhukov, V.I.; Shatkauskas, A.V.; Bushueva, L.K. The distribution of Ixodes persulcatus in the USSR. Zool. Zhurnal 1969, 48, 1003–1014. [Google Scholar]
- Zhang, G.; Zheng, D.; Tian, Y.; Li, S. A dataset of distribution and diversity of ticks in China. Sci. Data 2019, 6, 1–7. [Google Scholar] [CrossRef]
- Alekseev, A.N.; Dubinina, H.V.; Antykova, L.P.; Dzhivanyan, T.I.; Rijpkema, S.G.T.; Verbeek-De Kruif, N.; Cinco, M. Tick-Borne Borrelioses Pathogen Identification in Ixodes Ticks (Acarina, Ixodidae) Collected in St. Petersburg and Kaliningrad Baltic Regions of Russia. J. Med. Entomol. 1998, 35, 136–142. [Google Scholar] [CrossRef]
- Medlock, J.M.; Hansford, K.M.; Bormane, A.; Derdakova, M.; Estrada-Peña, A.; George, J.C.; Golovljova, I.; Jaenson, T.G.T.; Jensen, J.K.; Jensen, P.M.; et al. Driving forces for changes in geographical distribution of Ixodes ricinus ticks in Europe. Parasites Vectors 2013, 6, 1–11. [Google Scholar] [CrossRef]
- Bugmyrin, S.V.; Bespyatova, L.A.; Korotkov, Y.S.; Burenkova, L.A.; Belova, O.A.; Romanova, L.I.; Kozlovskaya, L.I.; Karganova, G.G.; Ieshko, E.P. Distribution of Ixodes ricinus and I. persulcatus ticks in southern Karelia (Russia). Ticks Tick. Borne. Dis. 2013, 4, 57–62. [Google Scholar] [CrossRef] [PubMed]
- Laaksonen, M.; Klemola, T.; Feuth, E.; Sormunen, J.J.; Puisto, A.; Mäkelä, S.; Penttinen, R.; Ruohomäki, K.; Hänninen, J.; Sääksjärvi, I.E.; et al. Tick-borne pathogens in Finland: Comparison of Ixodes ricinus and I. persulcatus in sympatric and parapatric areas. Parasites Vectors 2018, 11, 1–13. [Google Scholar] [CrossRef]
- Jääskeläinen, A.; Tonteri, E.; Pieninkeroinen, I.; Sironen, T.; Voutilainen, L.; Kuusi, M.; Vaheri, A.; Vapalahti, O. Siberian subtype tick-borne encephalitis virus in Ixodes ricinus in a newly emerged focus, Finland. Ticks Tick. Borne. Dis. 2016, 7, 216–223. [Google Scholar] [CrossRef] [PubMed]
- Jääskeläinen, A.E.; Tonter, E.; Sironen, T.; Pakarinen, L.; Vaheri, A.; Vapalahti, O. European Subtype Tick-borne Encephalitis Virus in Ixodes persulcatus Ticks. Emerg. Infect. Dis. 2011, 17, 323–325. [Google Scholar] [CrossRef] [PubMed]
- Ruzek, D.; Županc, T.A.; Borde, J.; Chrdle, A.; Eyer, L.; Karganova, G.; Kholodilov, I.; Knap, N.; Kozlovskaya, L.; Matveev, A.; et al. Tick-borne encephalitis in Europe and Russia: Review of pathogenesis, clinical features, therapy, and vaccines. Antivir. Res. 2019, 164, 23–51. [Google Scholar] [CrossRef] [PubMed]
- Guo, J.-J.; Lin, X.-D.; Chen, Y.-M.; Hao, Z.-Y.; Wang, Z.-X.; Yu, Z.-M.; Lu, M.; Li, K.; Qin, X.-C.; Wang, W.; et al. Diversity and circulation of Jingmen tick virus in ticks and mammals. Virus Evol. 2020, 6, 1–12. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Strain (s)/Amplicon (a) | Tick Species | Year, Region (GPS) | GenBank Access. No. |
|---|---|---|---|
| Alongshan virus | |||
| a. Miass501 | I. persulcatus | 2014, Chelyabinsk Region (55.02145°, 60.168283°) | MT210222 |
| s. Miass502 | I. persulcatus | MW525314–MW525317 | |
| s. Miass506 | I. persulcatus | MW525318–MW525321 | |
| a. Miass508 | I. persulcatus | MT210221 | |
| a. Miass510 | I. persulcatus | MT210225 | |
| a. Miass515 | I. persulcatus | MT210223 | |
| s. Miass519 | I. persulcatus | MN648774–MN648777 | |
| a. Miass523 | I. persulcatus | MT210224 | |
| s. Miass527 | I. persulcatus | MN648770–MN648773 | |
| s. Miass15-T22516 | I. persulcatus | 2015, Chelyabinsk Region (55.021583°, 60.169783°) | MW525284 |
| s. Miass15-T22516 | I. persulcatus | MW525285 | |
| a. Goms12-T16338 | I. persulcatus | 2012, Republic of Karelia (62,0690667°, 33,96141667°) | MW525286 |
| a. Goms13-T17158 | I. persulcatus | 2013, Republic of Karelia (62.069381°, 33.964528°) | MW525287 |
| a. Goms13-T17160/2 | I. persulcatus | MW525288 | |
| a. Goms13-T17182/2 | I. persulcatus | 2013, Republic of Karelia (62.068557°, 33.962730°) | MW525289 |
| a. Goms13-T17190/2 | I. persulcatus | MW525290 | |
| s. Galozero-14-T20426 | I. persulcatus | 2014, Republic of Karelia (62.075051°, 33.951404°) | MN604229, MW584331 |
| a. Goms14-T20532 | I. persulcatus | 2014, Republic of Karelia (62.063838°, 33.943815°) | MW525291 |
| a. Goms18-T27349 | I. persulcatus | 2018, Republic of Karelia (62.075230°, 33.946687°) | MW525292 |
| a. Goms18-T27350 | I. persulcatus | MW525293 | |
| a. Goms18-T27366 | I. persulcatus | 2018, Republic of Karelia (62.067203°, 33.933651°) | MW525294 |
| a. Erjey17-T25134 | I. persulcatus | 2017, Republic of Tuva (51.32646°, 95.98224°) | MW525295 |
| a. Sizim17-T25125 | I. persulcatus | 2017, Republic of Tuva (51.33110°, 95.94315°) | MW525296 |
| a. Republic Altay/777/2016 | D. nuttalli | 2016, Republic of Altai (51.24428°, 86.06387°) | MW556740 |
| a. Republic Altay/1395/2016 | D. nuttalli | 2016, Republic of Altai (49.8006°, 88.8875°) | MW556741 |
| a. Altay region/1710/2016 | H. concinna | 2016, Altai Territory (52.14821°, 85.97828°) | MW556738 |
| a. Altay region/1713/2016 | H. concinna | MW556739 | |
| a. Kursh17-T25178 | I. ricinus | 2017, Kaliningrad Region (54.972030°, 20.508669°) | MW525297 |
| a. Kursh17-T25208 | I. ricinus | MW525298 | |
| a. Kursh17-T25456 | I. ricinus | 2017, Kaliningrad Region (55.18242°, 20.85957°) | MW525299 |
| a. Kursh17-T25652 | I. ricinus | 2017, Kaliningrad Region (54.97020°, 20.50458°) | MW525300 |
| a. Kursh18-T30280 | I. ricinus | 2018, Kaliningrad Region (55.146110°, 20.824530°) | MW525304 |
| a. Kursh18-T30274 | I. ricinus | 2018, Kaliningrad Region (55.15465°, 20.82790°) | MW525302 |
| a. Kursh18-T27123 | I. ricinus | 2018, Kaliningrad Region (55.183363°, 20.857993°) | MW525301 |
| a. Kursh18-T30281 | I. ricinus | 2018, Kaliningrad Region (55.175897°, 20.846458°) | MW525305 |
| a. Kursh18-T30284 | I. ricinus | 2018, Kaliningrad Region (55.1591837°, 20.8432753°) | MW525306 |
| a. Kursh18-T30285 | I. ricinus | MW525307 | |
| a. Kursh18-T30286 | I. ricinus | MW525308 | |
| a. Kursh18-T30278 | I. ricinus | 2018, Kaliningrad Region (55.222354°, 20.891556°) | MW525303 |
| a. Kursh18-T30290 | I. ricinus | MW525309 | |
| a. Rowan19-T32778 | I. ricinus | 2019, Kaliningrad Region (54.827417°, 20.517167°) | MW525310 |
| a. Ulya14-T20695 | D. reticulatus | 2014, Ulyanovsk Region (54.446204°, 48.379524°) | MW525311 |
| a. Ulya15-T22688 | I. ricinus | 2015, Ulyanovsk Region (54.588423°, 48.416590°) | MW525312 |
| a. Tat14-T21924 | I. ricinus | 2014, Republic of Tatarstan (55.85397°, 48.7577°) | MW525313 |
| Yanggou tick virus | |||
| s. Erzin14-T20074 | D. nuttalli | 2014, Republic of Tuva (50.13304°, 95.4722°) | MW525322–MW525325 |
| s. Republic Altay/1001/2016 | D. marginatus | 2016, Republic of Altai (52.02653°, 85.85219°) | MW556734–MW556737 |
| a. Republic Altay/997/2016 | D. marginatus | MW556730–MW556733 | |
| Region of Russia | Tick Species | Number of Analysed Ticks (Total) | Number of Analysed Pools/Individual Ticks | Number of Positive Pools/Individual Ticks | MIR 1 (%) |
|---|---|---|---|---|---|
| Alongshan virus | |||||
| Republic of Tuva | I. persulcatus | 348 | 73/12 | 2/0 | 0.6 |
| Republic of Altai | D. nuttalli | 19 | 0/19 | 0/1 | 5.2 2 |
| D. nuttalli | 26 | 0/26 | 0/1 | 3.8 | |
| Altai Territory | H. concinna | 16 | 0/16 | 0/2 | 12.5 2 |
| Chelyabinsk Region | I. persulcatus | 254 | 62/0 | 11/0 | 4.3 |
| Republic of Tatarstan | I. ricinus | 74 | 16/0 | 1/0 | 1.4 |
| Ulyanovsk Region | I. ricinus | 20 | 0/20 | 0/1 | 5.0 2 |
| D. reticulatus | 9 | 2/0 | 1/0 | 11.1 2 | |
| Republic of Karelia | I. persulcatus | 1265 | 265/0 | 10/0 | 0.8 |
| Kaliningrad Region | I. ricinus (1) 3 | 57 | 23/0 | 1/0 | 1.8 |
| I. ricinus (2) 3 | 59 | 14/0 | 3/0 | 5.1 | |
| I. ricinus (3) 3 | 129 | 30/0 | 10/0 | 7.8 | |
| Yanggou tick virus | |||||
| Republic of Tuva | D. nuttalli | 205 | 49/20 | 1/0 | 0.5 |
| Republic of Altai | D. marginatus | 25 | 0/25 | 0/2 | 8.0 |
| Amino Acid Position | Ixodes ricinus Group | Ixodes persulcatus Group | |
|---|---|---|---|
| European Subgroup | Asian Subgroup | ||
| VP1a | |||
| 8 | Ala | Ala | Thr |
| 72 | Val | Ala | Ala |
| 115 | Ala | Val | Val |
| 135 | Val | Lys | Lys |
| 138 | Pro | Ser | Ser |
| 153 | Lys | Arg | Arg |
| 210 | Gly | Gly | Ser |
| 216 | Thr | Ala | Ala |
| 321 | Val | Val | Thr |
| 460 | Thr | Met | Thr |
| 472 | Arg | His | His |
| 476 | Arg | Arg | Gln |
| 479 | Arg/His | Arg | His |
| VP1b | |||
| 118 | Met | Met | Leu |
| 172 | Ile | Ile | Va |
| 187 | Lys | Lys | Arg |
| 195 | Ser | Ser | Gly |
| 276 | Val | Ile | Ile |
| nuORF | |||
| 4 | Lys | Lys | Gln |
| 15 | Asp | Asp | Asn |
| 132 | Thr | Ala | Thr |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kholodilov, I.S.; Belova, O.A.; Morozkin, E.S.; Litov, A.G.; Ivannikova, A.Y.; Makenov, M.T.; Shchetinin, A.M.; Aibulatov, S.V.; Bazarova, G.K.; Bell-Sakyi, L.; et al. Geographical and Tick-Dependent Distribution of Flavi-Like Alongshan and Yanggou Tick Viruses in Russia. Viruses 2021, 13, 458. https://doi.org/10.3390/v13030458
Kholodilov IS, Belova OA, Morozkin ES, Litov AG, Ivannikova AY, Makenov MT, Shchetinin AM, Aibulatov SV, Bazarova GK, Bell-Sakyi L, et al. Geographical and Tick-Dependent Distribution of Flavi-Like Alongshan and Yanggou Tick Viruses in Russia. Viruses. 2021; 13(3):458. https://doi.org/10.3390/v13030458
Chicago/Turabian StyleKholodilov, Ivan S., Oxana A. Belova, Evgeny S. Morozkin, Alexander G. Litov, Anna Y. Ivannikova, Marat T. Makenov, Alexey M. Shchetinin, Sergey V. Aibulatov, Galina K. Bazarova, Lesley Bell-Sakyi, and et al. 2021. "Geographical and Tick-Dependent Distribution of Flavi-Like Alongshan and Yanggou Tick Viruses in Russia" Viruses 13, no. 3: 458. https://doi.org/10.3390/v13030458
APA StyleKholodilov, I. S., Belova, O. A., Morozkin, E. S., Litov, A. G., Ivannikova, A. Y., Makenov, M. T., Shchetinin, A. M., Aibulatov, S. V., Bazarova, G. K., Bell-Sakyi, L., Bespyatova, L. A., Bugmyrin, S. V., Chernetsov, N., Chernokhaeva, L. L., Gmyl, L. V., Khaisarova, A. N., Khalin, A. V., Klimentov, A. S., Kovalchuk, I. V., ... Karganova, G. G. (2021). Geographical and Tick-Dependent Distribution of Flavi-Like Alongshan and Yanggou Tick Viruses in Russia. Viruses, 13(3), 458. https://doi.org/10.3390/v13030458