
Structural and Functional Diversity among Five RING Finger Proteins from Carassius Auratus Herpesvirus (CaHV)


Abstract
1. Introduction
2. Materials and Methods
2.1. Virus, Cell Lines, and Reagents
2.2. Sequence Analysis of RING Finger Protein Homologs
2.3. Polymerase Chain Reaction (PCR) and Plasmid Construction
2.4. Transfection and Fluorescence Observation
2.5. Ubiquitination Detection in Cell Culture
2.6. Prokaryotic Expression and Purification of Proteins
2.7. Ubiquitination Assays of Purified Proteins
3. Results
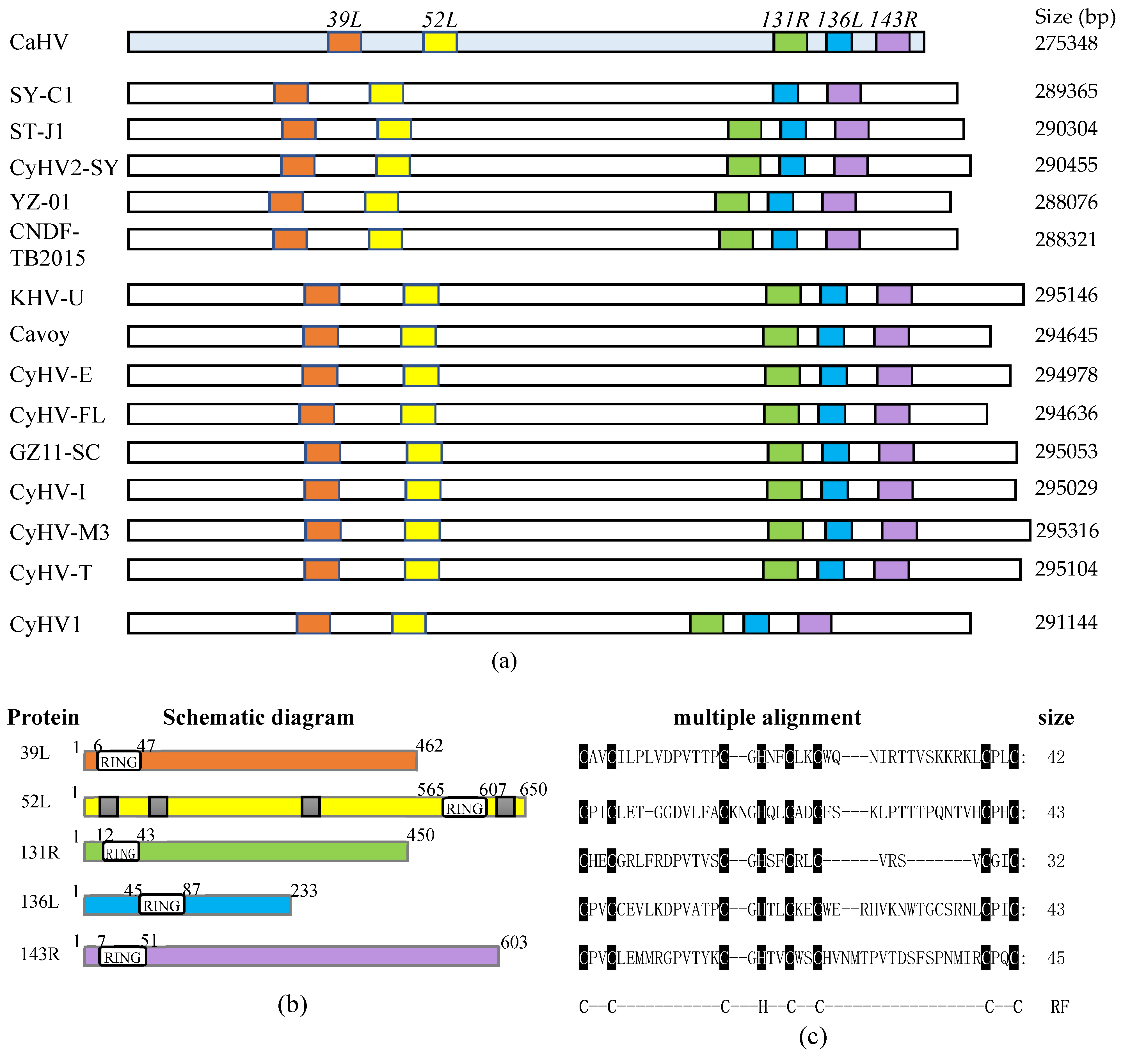
3.1. Fish Herpesvirus Encoding RING Finger Proteins
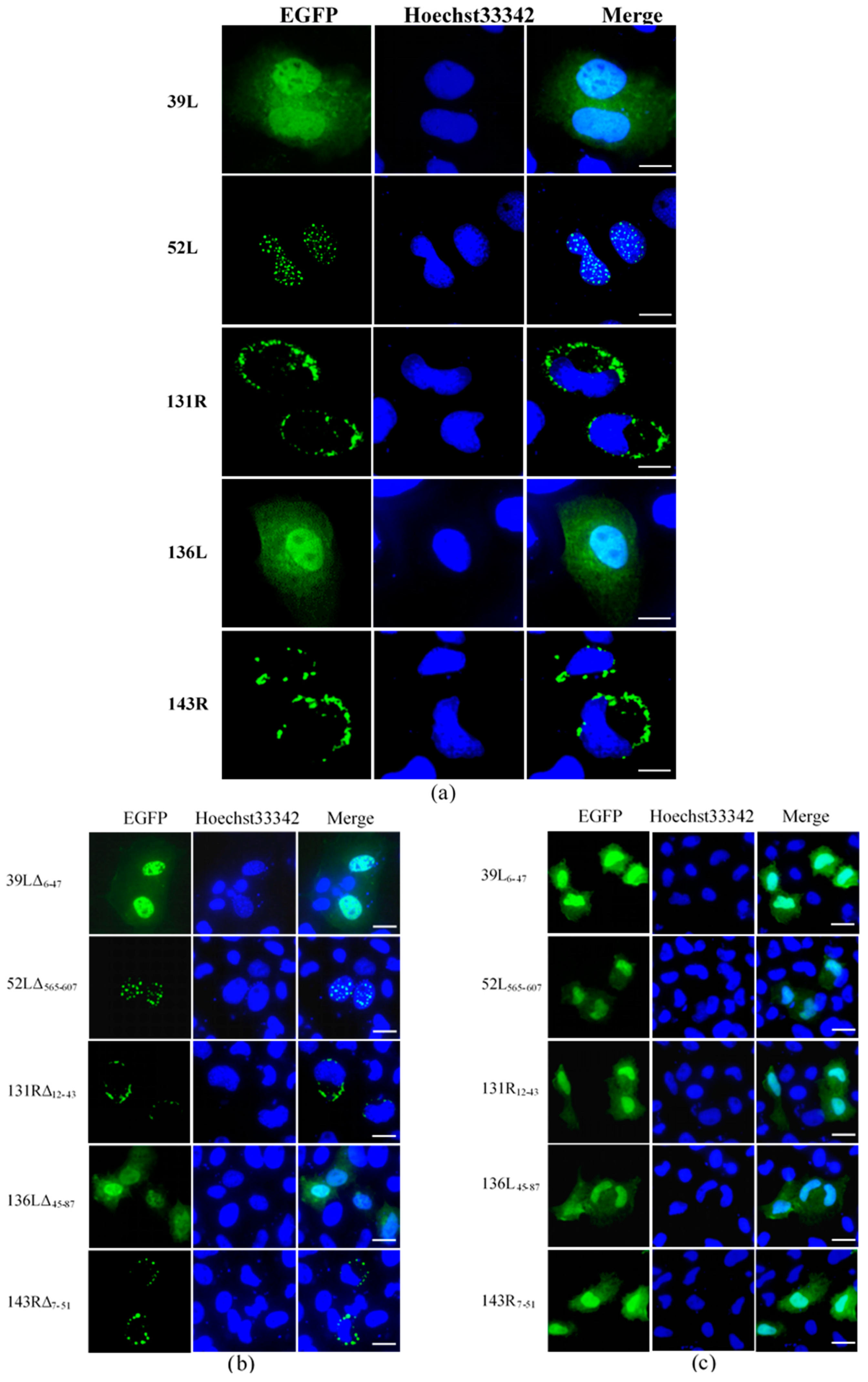
3.2. Subcellular Localization of RING Finger Proteins
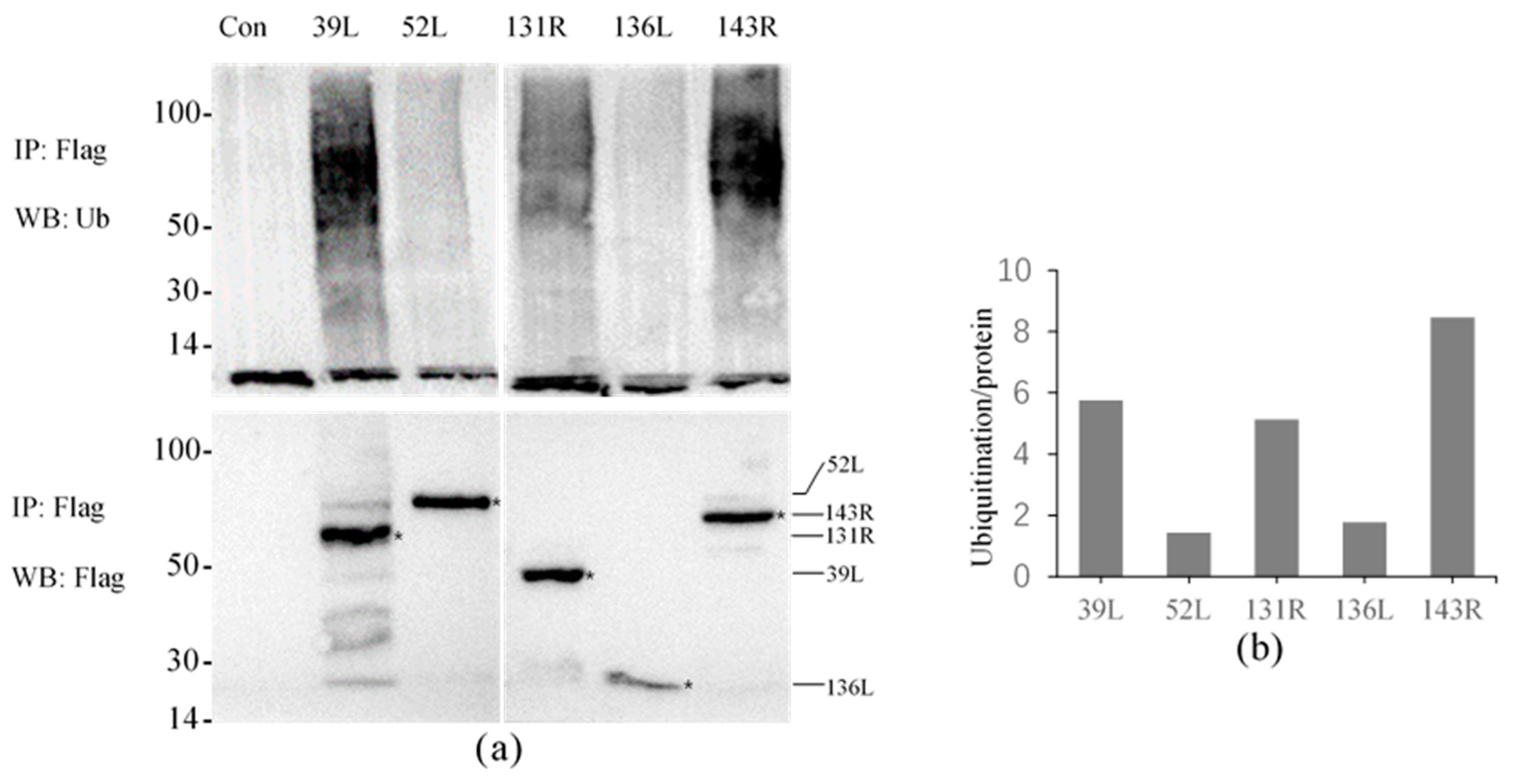
3.3. Ubiquitination Activity of RING Finger Proteins in Cell Culture
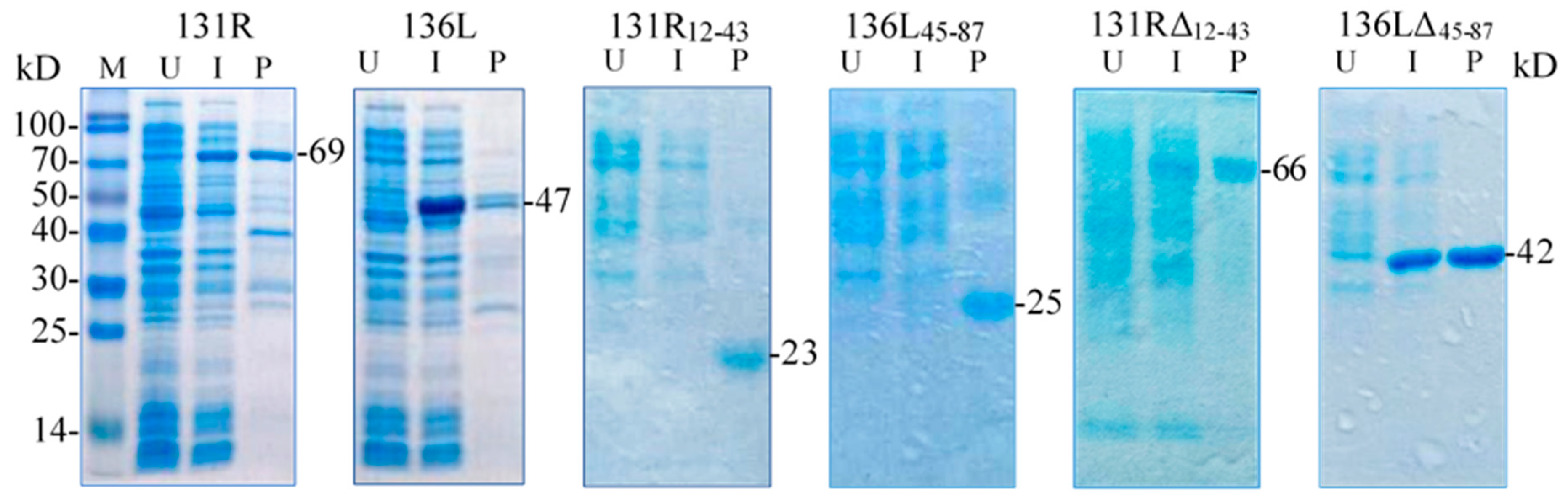
3.4. Identification of Prokaryotic Expression Products
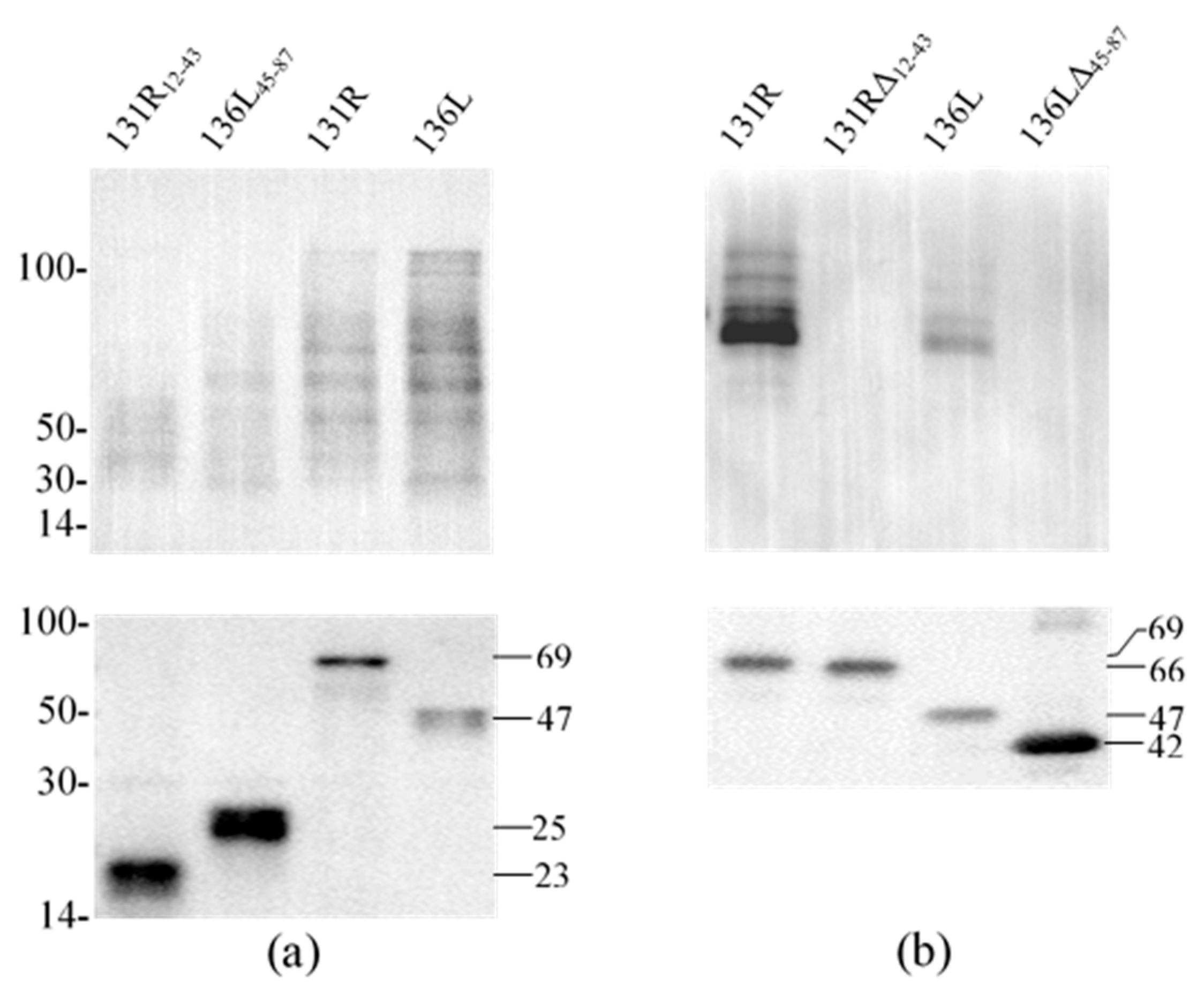
3.5. Ubiquitination Activity of the Recombinant Proteins
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Naylor, R.L.; Goldburg, R.J.; Primavera, J.H.; Kautsky, N.; Beveridge, M.C.; Clay, J.; Folke, C.; Lubchenco, J.; Mooney, H.; Troell, M. Effect of aquaculture on world fish supplies. Nature 2000, 405, 1017–1024. [Google Scholar] [CrossRef]
- Gui, J.F.; Zhou, L.; Zhang, X.J. Research Advances and Prospects for Fish Genetic Breeding. Bull Chin. Acad. Sci. 2018, 33, 932–939. [Google Scholar]
- Belton, B.; Little, D.C.; Zhang, W.; Edwards, P.; Skladany, M.; Thilsted, S.H. Farming fish in the sea will not nourish the world. Nat. Commun. 2020, 11, 5804. [Google Scholar] [CrossRef]
- Lu, M.; Li, X.Y.; Li, Z.; Du, W.X.; Zhou, L.; Wang, Y.; Zhang, X.J.; Wang, Z.W.; Gui, J.F. Regain of sex determination system and sexual reproduction ability in a synthetic octoploid male fish. Sci. China Life Sci. 2020, 63. [Google Scholar] [CrossRef]
- Zhang, X.J.; Zhou, L.; Gui, J.F. Biotechnological innovation in genetic breeding and sustainable green development in Chinese aquaculture. Sci. China Life Sci. 2019, 49, 1409–1429. [Google Scholar] [CrossRef]
- Hanson, L.; Dishon, A.; Kotler, M. Herpesviruses that infect fish. Viruses 2011, 3, 2160–2191. [Google Scholar] [CrossRef]
- Kibenge, F.S. Emerging viruses in aquaculture. Curr. Opin. Virol. 2019, 34, 97–103. [Google Scholar] [CrossRef]
- Gui, L.; Zhang, Q.Y. A brief review on aquatic animal virology researches in China. J. Fish. China 2019, 43, 168–187, (In Chinese with English abstract). [Google Scholar]
- Davison, A.J.; Kurobe, T.; Gatherer, D.; Cunningham, C.; Korf, I.; Fukuda, H.; Hedrick, R.P.; Waltzek, T.B. Comparative genomics of carp herpesviruses. J. Virol. 2013, 87, 2908–2922. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Q.Y.; Gui, J.F. Virus genomes and virus-host interactions in aquaculture animals. Sci. China Life Sci. 2015, 58, 156–169. [Google Scholar] [CrossRef] [PubMed]
- Gui, L.; Zhang, Q.Y. Disease prevention and control. In Aquaculture in China: Success Stories and Modern Trends; Gui, J.F., Tang, Q.S., Li, Z.J., Liu, J.S., Sena, S.S.D., Eds.; Wiley-Blackwell: Chichester, UK, 2018; pp. 577–598. [Google Scholar]
- Zhang, Q.Y. An overview on several large DNA viruses in freshwater ecosystems. Acta Hydrobiologica Sinica 2020, 5, 28–42, (Chinese with English abstract). [Google Scholar]
- Zeng, X.T.; Chen, Z.Y.; Deng, Y.S.; Gui, J.F.; Zhang, Q.Y. Complete genome sequence and architecture of crucian carp Carassius auratus herpesvirus (CaHV). Arch. Virol. 2016, 161, 3577–3581. [Google Scholar] [CrossRef]
- Wang, J.; Gui, L.; Chen, Z.Y.; Zhang, Q.Y. Mutations in the Cterminal region affect subcellular localization of crucian carp herpesvirus (CaHV) GPCR. Virus Genes 2016, 52, 484–494. [Google Scholar] [CrossRef]
- Mou, C.Y.; Wang, Y.; Zhang, Q.Y.; Gao, F.X.; Li, Z.; Tong, J.F.; Zhou, L.; Gui, J.F. Differential interferon system gene expression profiles in susceptible and resistant gynogenetic clones of gibel carp challenged with herpesvirus CaHV. Dev. Comp. Immunol. 2018, 86, 52–64. [Google Scholar] [CrossRef]
- Wang, Z.H.; Zhang, Q.Y. Characterization of Carassius auratus herpesvirus ORF31R (CaHV-31R) and the encoded protein colocalize with cellular organs. J. Fish. China 2019, 43, 1263–1270, (In Chinese with English abstract). [Google Scholar]
- Lu, W.J.; Gao, F.X.; Wang, Y.; Zhang, Q.Y.; Li, Z.; Zhang, X.J.; Zhou, L.; Gui, J.F. Differential expression of innate and adaptive immune genes in the survivors of three gibel carp gynogenetic clones after herpesvirus challenge. BMC Genom. 2019, 20, 432. [Google Scholar] [CrossRef]
- Zhao, Y.H.; Zeng, X.T.; Zhang, Q.Y. Fish herpesvirus protein (CaHV-138L) can target to mitochondrial protein FoF1 ATPase. Virus Res. 2020, 275, 197754. [Google Scholar] [CrossRef]
- Inman, M.; Zhang, Y.; Geiser, V.; Jones, C. The zinc ring finger in the bICP0 protein encoded by bovine herpesvirus-1 mediates toxicity and activates productive infection. J. Gen. Virol. 2001, 82, 483–492. [Google Scholar] [CrossRef]
- Madigan, V.J.; Yuziuk, J.A.; Chiarella, A.M.; Tyson, T.O.; Meganck, R.M.; Elmore, Z.C.; Tse, L.V.; Hathaway, N.A.; Asokan, A. Ring finger protein 121 is a potent regulator of adeno-associated viral genome transcription. PLoS Pathog. 2019, 15, e1007988. [Google Scholar] [CrossRef] [PubMed]
- Joazeiro, C.A.; Weissman, A.M. RING finger proteins: Mediators of ubiquitin ligase activity. Cell 2000, 102, 549–552. [Google Scholar] [CrossRef]
- Nakazawa, Y.; Hara, Y.; Oka, Y.; Komine, O.; Van den Heuvel, D.; Guo, C.; Daigaku, Y.; Isono, M.; He, Y.; Shimada, M.; et al. Ubiquitination of DNA Damage-Stalled RNAPII Promotes Transcription-Coupled Repair. Cell 2020, 180, 1228–1244. [Google Scholar] [CrossRef]
- Rodríguez, M.C.; Dybas, J.M.; Hughes, J.; Weitzman, M.D.; Boutell, C. The HSV-1 ubiquitin ligase ICP0: Modifying the cellular proteome to promote infection. Virus Res. 2020, 285, 198015. [Google Scholar] [CrossRef] [PubMed]
- Deshaies, R.J.; Joazeiro, C.A.P. RING domain E3 ubiquitin ligases. Annu. Rev. Biochem. 2009, 78, 399–434. [Google Scholar] [CrossRef] [PubMed]
- Xie, J.; Zhu, J.; Yang, H.; Weng, S.; Chen, C.; Chen, H.; Xie, Q.; Li, M.; He, J. RNFs of infectious spleen and kidney necrosis virus (ISKNV) function as ubiquitin ligase enzymes. Virus Res. 2007, 123, 170–177. [Google Scholar] [CrossRef]
- Shekar, M.; Venugopal, M.N. Identification and characterization of novel double zinc fingers encoded by putative proteins in genome of white spot syndrome virus. Arch. Virol. 2019, 164, 961–969. [Google Scholar] [CrossRef] [PubMed]
- Margolis, D.M.; Archin, N.M.; Cohen, M.S.; Eron, J.J.; Ferrari, G.; Garcia, J.V.; Gay, C.L.; Goonetilleke, N.; Joseph, S.B.; Swanstrom, R.; et al. Curing HIV: Seeking to Target and Clear Persistent Infection. Cell 2020, 181, 189–206. [Google Scholar] [CrossRef]
- Sommerset, I.; Krossøy, B.; Biering, E.; Frost, P. Vaccines for fish in aquaculture. Expert Rev. Vaccines 2005, 4, 89–101. [Google Scholar] [CrossRef] [PubMed]
- Chen, Z.Y.; Gui, J.F.; Gao, X.C.; Pei, C.; Hong, Y.J.; Zhang, Q.Y. Genome architecture changes and major gene variations of Andrias davidianus ranavirus (ADRV). Vet. Res. 2013, 44, 101. [Google Scholar] [CrossRef]
- Fang, J.; Deng, Y.S.; Wang, J.; Li, T.; Chen, Z.Y.; Zhou, L.; Zhang, Q.Y. Pathological changes of acute viral hemorrhages in the gills of crucian carp. J. Fish. Sci. China 2016, 23, 336–343, (In Chinese with English abstract). [Google Scholar]
- Zeng, X.T.; Gao, X.C.; Zhang, Q.Y. Rana grylio virus 43R encodes an envelope protein involved in virus entry. Virus Genes 2018, 54, 779–791. [Google Scholar] [CrossRef]
- Zeng, X.T.; Zhang, Q.Y. Interaction between Two Iridovirus Core Proteins and Their Effects on Ranavirus (RGV) Replication in Cells from Different Species. Viruses 2019, 11, 416. [Google Scholar] [CrossRef]
- Deschamps, T.; Waisner, H.; Dogrammatzis, C.; Roy, A.; Chacko, S.; Perera, C.; Prisinzano, T.E.; Kalamvoki, M. Discovery of Small-Molecule Inhibitors Targeting the E3 Ubiquitin Ligase Activity of the Herpes Simplex Virus 1 ICP0 Protein Using an in vitro High-Throughput Screening Assay. J. Virol. 2019, 93, e00619-19. [Google Scholar] [CrossRef]
- Aoki, T.; Hirono, I.; Kurokawa, K.; Fukuda, H.; Nahary, R.; Eldar, A.; Davison, A.J.; Waltzek, T.B.; Bercovier, H.; Hedrick, R.P. Genome sequences of three koi herpesvirus isolates representing the expanding distribution of an emerging disease threatening koi and common carp worldwide. J. Virol. 2007, 81, 5058–5065. [Google Scholar] [CrossRef]
- Li, L.L.; Luo, Y.Z.; Gao, Z.X.; Huang, J.; Zheng, X.H.; Nie, H.H.; Zhang, J.M.; Lin, L.; Yuan, J.F. Molecular characterisation and prevalence of a new genotype of Cyprinid herpesvirus 2 in mainland China. Can. J. Microbiol. 2015, 61, 1–7. [Google Scholar] [CrossRef]
- Gao, Y.; Suárez, N.M.; Wilkie, G.S.; Dong, C.; Bergmann, S.; Lee, P.A.; Davison, A.J.; Vanderplasschen, A.F.C.; Boutier, M. Genomic and biologic comparisons of cyprinid herpesvirus 3 strains. Vet. Res. 2018, 49, 40–50. [Google Scholar] [CrossRef] [PubMed]
- Zhu, R.; Zhang, Y.B.; Zhang, Q.Y.; Gui, J.F. Functional domains and the antiviral effect of the double-stranded RNA-dependent protein kinase PKR from Paralichthys olivaceus. J. Virol. 2008, 82, 6889–6901. [Google Scholar] [CrossRef] [PubMed]
- Zhao, Z.; Ke, F.; Huang, Y.H.; Zhao, J.G.; Gui, J.F.; Zhang, Q.Y. Identification and characterization of a novel envelope protein in Rana grylio virus. J. Gen. Virol. 2008, 89, 1866–1872. [Google Scholar] [CrossRef] [PubMed]
- Ke, F.; Wang, Z.H.; Ming, C.Y.; Zhang, Q.Y. Ranaviruses Bind Cells from Different Species through Interaction with Heparan Sulfate. Viruses 2019, 11, 593. [Google Scholar] [CrossRef]
- Wang, Z.H. (State Key Laboratory of Freshwater Ecology and Biotechnology, Institute of Hydrobiology, Chinese Academy of Sciences, Wuhan, China). Personal communication, 2020.
- Van Beurden, S.J.; Bossers, A.; Voorbergen-Laarman, M.H.; Haenen, O.L.; Peters, S.; Abma-Henkens, M.H.; Peeters, B.P.; Rottier, P.J.; Engelsma, M.Y. Complete genome sequence and taxonomic position of anguillid herpesvirus 1. J. Gen. Virol. 2010, 91, 880–887. [Google Scholar] [CrossRef]
- Budhidarmo, R.; Nakatani, Y.; Day, C.L. RINGs hold the key to ubiquitin transfer. Trends Biochem. Sci. 2012, 37, 58–65. [Google Scholar] [CrossRef]
- Han, Y.; Li, R.; Gao, J.; Miao, S.; Wang, L. Characterisation of human RING finger protein TRIM69, a novel testis E3 ubiquitin ligase and its subcellular localisation. Biochem. Biophys. Res. Commun. 2012, 429, 6–11. [Google Scholar] [CrossRef]
- Lorick, K.L.; Jensen, J.P.; Fang, S.; Ong, A.M.; Hatakeyama, S.; Weissman, A.M. 1999. RING fingers mediate ubiquitin-conjugating enzyme (E2)-dependent ubiquitination. Proc. Natl. Acad. Sci. USA 1999, 96, 11364–11369. [Google Scholar] [CrossRef]
- Branigan, E.; Penedo, J.C.; Hay, R.T. Ubiquitin transfer by a RING E3 ligase occurs from a closed E2~ubiquitin conformation. Nat. Commun. 2020, 11, 2846. [Google Scholar] [CrossRef]
- Martinez-Chacin, R.C.; Bodrug, T.; Bolhuis, D.L.; Kedziora, K.M.; Bonacci, T.; Ordureau, A.; Gibbs, M.E.; Weissmann, F.; Qiao, R.; Grant, G.D.; et al. Ubiquitin chain-elongating enzyme UBE2S activates the RING E3 ligase APC/C for substrate priming. Nat. Struct. Mol. Biol. 2020, 27, 550–560. [Google Scholar] [CrossRef]
- He, F.; Fenner, B.J.; Godwin, A.K.; Kwang, J. White spot syndrome virus open reading frame 222 encodes a viral E3 ligase and mediates degradation of a host tumor suppressor via ubiquitination. J. Virol. 2006, 80, 3884–3892. [Google Scholar] [CrossRef] [PubMed]
- Vanni, E.; Gatherer, D.; Tong, L.; Everett, R.D.; Boutell, C. Functional characterization of residues required for the herpes simplex virus 1 E3 ubiquitin ligase ICP0 to interact with the cellular E2 ubiquitin-conjugating enzyme UBE2D1 (UbcH5a). J. Virol. 2012, 86, 6323–6333. [Google Scholar] [CrossRef]
- Cao, C.; An, R.; Yu, Y.; Dai, H.; Qu, Z.; Gao, M.; Wang, J. BICP0 Negatively Regulates TRAF6-Mediated NF-κB and Interferon Activation by Promoting K48-Linked Polyubiquitination of TRAF6. Front. Microbiol. 2020, 8, 3040. [Google Scholar] [CrossRef] [PubMed]
- Bergmann, S.M.; Sadowski, J.; Kiełpiński, M.; Bartłomiejczyk, M.; Fichtner, D.; Riebe, R.; Lenk, M.; Kempter, J. Susceptibility of koi x crucian carp and koi x goldfish hybrids to koi herpesvirus (KHV) and the development of KHV disease (KHVD). J. Fish Dis. 2010, 33, 267–272. [Google Scholar] [CrossRef] [PubMed]
- Thul, P.J.; Åkesson, L.; Wiking, M.; Mahdessian, D.; Geladaki, A.; Ait Blal, H.; Alm, T.; Asplund, A.; Björk, L.; Breckels, L.M.; et al. A subcellular map of the human proteome. Science 2017, 356, eaal3321. [Google Scholar] [CrossRef]
- Halford, W.P.; Schaffer, P.A. ICP0 is required for efficient reactivation of herpessimplex virus type 1 from neuronal latency. J. Virol. 2001, 75, 3240–3249. [Google Scholar] [CrossRef]
- Zheng, Y.; Gu, H. Identification of three redundant segments responsible for herpes simplex virus 1 ICP0 to fuse with ND10 nuclear bodies. J. Virol. 2015, 89, 4214–4226. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Mostafa, H.H.; Thompson, T.W.; Davido, D.J. N-terminal phosphorylation sites of herpes simplex virus 1 ICP0 differentially regulate its activities and enhance viral replication. J. Virol. 2013, 87, 2109–2119. [Google Scholar] [CrossRef]
- Nerenberg, B.T.; Taylor, J.; Bartee, E.; Gouveia, K.; Barry, M.; Früh, K. The poxviral RING protein p28 is a ubiquitin ligase that targets ubiquitin to viral replication factories. J. Virol. 2005, 79, 597–601. [Google Scholar] [CrossRef] [PubMed]
- Pardieu, C.; Vigan, R.; Wilson, S.J.; Calvi, A.; Zang, T.; Bieniasz, P.; Kellam, P.; Towers, G.J.; Neil, S.J. The RING-CH ligase K5 antagonizes restriction of KSHV and HIV-1 particle release by mediating ubiquitin-dependent endosomal degradation of tetherin. PLoS Pathog. 2010, 6, e1000843. [Google Scholar] [CrossRef] [PubMed]
- Bareiss, B.; Barry, M. Fowlpox virus encodes two p28-like ubiquitin ligases that are expressed early and late during infection. Virology 2014, 462–463, 60–70. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Said, A.; Osterrieder, N. Equine herpesvirus type 1 (EHV-1) open reading frame 59 encodes an early protein that is localized to the cytosol and required for efficient virus growth. Virology 2014, 449, 263–269. [Google Scholar] [CrossRef]
- Zhang, K.; Brownlie, R.; Snider, M.; Van Drunen, L.; Van den Hurk, S. Phosphorylation of Bovine Herpesvirus 1 VP8 Plays a Role in Viral DNA Encapsidation and Is Essential for Its Cytoplasmic Localization and Optimal Virion Incorporation. J. Virol. 2016, 90, 4427–4440. [Google Scholar] [CrossRef]
- Coscoy, L.; Sanchez, D.J.; Ganem, D. A novel class of herpesvirus-encoded membrane-bound E3 ubiquitin ligases regulates endocytosis of proteins involved in immune recognition. J. Cell Biol. 2001, 155, 1265–1273. [Google Scholar] [CrossRef]
- Cadwell, K.; Coscoy, L. Ubiquitination on nonlysine residues by a viral E3 ubiquitin ligase. Science 2005, 309, 127–130. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Primers | Primer Sequence (5′ to 3′) | Plasmids | Size of DNA Fragments (bp) |
|---|---|---|---|
| 39L-F | AACAAGCTTAATAATGGAAGACACG (HindIII) | pEGFP-39L | 1389 |
| 39L-R | GATGGTACCGACATGGTTCGAC (KpnI) | ||
| 52L-F | AGAAAGCTTAACACGATGCCGAC (HindIII) | pEGFP-52L | 1953 |
| 51L-R | TCTGAGGTACCACATGTGACCATAG (KpnI) | ||
| 52L-linkerF | AGGCCTCAACAGGTGCCCGATACC | ||
| 52L-linkerR | GGTATCGGGCACCTGTTGAGGCCT | ||
| 131R-F | ACAAAGCTTCAACCATGGATCGT (HindIII) | pEGFP-131R | 1353 |
| 131R-R | AATCCGGATCCGAGTGTTAGAGT (BamHI) | ||
| 136L-F | TTTGAATTCGACAATGTCCACCT (EcoRI) | pEGFP-136L | 702 |
| 136L-R | CGCGGATCCGAGTATGCTGCAGAAGC (BamHI) | ||
| 143R-F | CAGCAGAAGCTTCTAAAAATGACG (HindIII) | pEGFP-143R | 1812 |
| 143R-R | CACAGGATCCGATATTAGCTACAAT (BamHI) | ||
| 39LΔ6–47-F | TCAGATCTCGAGCTCAAGCTTATGGAAGACACGGACAGAGAAAAGGTGAAGGGCACG (HindIII) | pEGFP-39LΔ6–47 | 1263 |
| 39LΔ6–47-R | GGATCCCGGGCCCGCGGTACCGACATGGTTCGACGTGACAA (KpnI) | ||
| 52LΔ565–607-F | TCAGATCTCGAGCTCAAGCTTATGCCGACCTGGCCCATGTTT (HindIII) | pEGFP-52LΔ565–607 | 1824 |
| 52LΔ565–607-R | GGATCCCGGGCCCGCGGTACCACATGTGACCATAGACTTAAAGG (KpnI) | ||
| 52LΔ565–607-F1 | GCTACCGTTCTCGGA GACGTTGCCCTGGTCAATTAT | ||
| 52LΔ565–607-R1 | GACCAGGGCAACGTC TCCGAGAACGGTAGCCACTTC | ||
| 131RΔ12–43-F | TCAGATCTCGAGCTCAAGCTTATGGATCGTGAGACTCTACTCGGGCACCTGAGCCTGATAGAGTGCAGTCAACCGT (HindIII) | pEGFP-131RΔ12–43 | 1257 |
| 131RΔ12–43-R | GGATCCCGGGCCCGCGGTACCGAGTGTTAGAGTTATGGAAGC (KpnI) | ||
| 136LΔ45–87-F | TCAGATCTCGAGCTCAAGCTTATGTCCACCTACGTTGATATG (HindIII) | pEGFP-136LΔ45–87 | 573 |
| 136LΔ45–87-R | GGATCCCGGGCCCGCGGTACCGAGTATGCTGCAGAAGC (KpnI) | ||
| 136LΔ45–87-F1 | GTCAAGTCGTTGATG AACAGGGACGTACCCGATAAT | ||
| 136LΔ45–87-R1 | GGGTACGTCCCTGTT CATCAACGACTTGACAAAGTTG | ||
| 143RΔ7–51-F | TCAGATCTCGAGCTCAAGCTTATGACGGAGCCTCTGGATAGGACCATGGTGTGCAAAAAC (HindIII) | pEGFP-143RΔ7–51 | 1677 |
| 143RΔ7–51-R | GGATCCCGGGCCCGCGGTACCGATATTAGCTACAATAGTGGC (KpnI) | ||
| 39L6–47-F | TCAGATCTCGAGCTCAAGCTTATGGAAGACACGGACTGTGC (HindIII) | pEGFP-39L6–47 | 126 |
| 39L6–47-R | GGATCCCGGGCCCGCGGTACCTCTGAGCGTGCCCTTCAC (KpnI) | ||
| 52L565–607-F | TCAGATCTCGAGCTCAAGCTTCCGTTCTCGGATGTCCCAT (HindIII) | pEGFP-52L565–607 | 129 |
| 52L565–607-R | GGATCCCGGGCCCGCGGTACCATAATTGACCAGGGCAACG (KpnI) | ||
| 131R12–43-F | TCAGATCTCGAGCTCAAGCTTTGCCACGAGTGCGGCAGAC (HindIII) | pEGFP-131R12–43 | 96 |
| 131R12–43-R | GGATCCCGGGCCCGCGGTACCACAGATTCCACACACCGATCTG (KpnI) | ||
| 136L45–87-F | TCAGATCTCGAGCTCAAGCTTTGTCCCGTGTGCTGCGAG (HindIII) | pEGFP-136L45–87 | 129 |
| 136L45–87-R | GGATCCCGGGCCCGCGGTACCACAGATGGGACACAGGTTACG (KpnI) | ||
| 143R7–51-F | TCAGATCTCGAGCTCAAGCTTTGTCCCGTGTGCCTGGAAATG (HindIII) | pEGFP-143R7–51 | 135 |
| 143R7–51-R | GGATCCCGGGCCCGCGGTACCACACTGTGGGCACCTGATC (KpnI) | ||
| 131R-E-F | ACAGGATCCATGGATCGTGAGACTCTACTC (BamHI) | pET131R | 1353 |
| 131R-E-R | TCCAAGCTTGAGTGTTAGAGTTATGGAAGC (HindIII) | ||
| 136L-E-F | CGCGGATCCATGTCCACCTACGTTGATATG (BamHI) | pET136L | 702 |
| 136L-E-R | CCGCTCGAGGAGTATGCTGCAGAAGC (XhoI) | ||
| 131R12–43-E-F | GCCATGGCTGATATCGGATCCTGCCACGAGTGCGGCAGAC (BamHI) | pET131R12–43 | 96 |
| 131R12–43-E-R | CTCGAGTGCGGCCGCAAGCTTACAGATTCCACACACCGATCTG (HindIII) | ||
| 136L45–87-E-F | GCCATGGCTGATATCGGATCCTGTCCCGTGTGCTGCGAG (BamHI) | pET136L45–87 | 129 |
| 136L45–87-E-R | CTCGAGTGCGGCCGCAAGCTTACAGATGGGACACAGGTTACG (HindIII) | ||
| 131RΔ12–43-E-F | ATCGGATCCGAATTCGAGCTCATGGATCGTGAGACTCTACTC (SacI) | pET131RΔ12–43 | 1257 |
| 131RΔ12–43-E-R | CTCGAGTGCGGCCGCAAGCTTGAGTGTTAGAGTTATGGAAGC (HindIII) | ||
| 136LΔ45–87-E-F | ATCGGATCCGAATTCGAGCTCATGTCCACCTACGTTGATATG (SacI) | pET136LΔ45–87 | 573 |
| 136LΔ45–87-E-R | CTCGAGTGCGGCCGCAAGCTTGAGTATGCTGCAGAAGC (HindIII) | ||
| 39L-C-F | CCGGAATTCGATGGAAGACACGGACTGTGC (EcoRI) | pCMV-39L | 1389 |
| 39L-C-R | CCCAAGCTTGACATGGTTCGACGTGACAA (HindIII) | ||
| 52L-C-F | CCCAAGCTTGATGCCGACCTGGCCCATGTTT (HindIII) | pCMV-52L | 1953 |
| 52L-C-R | ACGCGTCGACCTAACATGTGACCATAGACTT (SalI) | ||
| 131R-C-F | CGCGGATCCGATGGATCGTGAGACTCTACTC (BamHI) | pCMV-131R | 1353 |
| 131R-C-R | CCCAAGCTTGAGTGTTAGAGTTATGGAAGC (HindIII) | ||
| 136L-C-F | CGCGGATCCGATGTCCACCTACGTTGATATG (BamHI) | pCMV-136L | 702 |
| 136L-C-R | CCGCTCGAGGAGTATGCTGCAGAAGC (XhoI) | ||
| 143R-C-F | CGCGGATCCGATGACGGAGCCTCTGGATTG (BamHI) | pCMV-143R | 1812 |
| 143R-C-R | CCCAAGCTTGATATTAGCTACAATAGTGGC (HindIII) | ||
| Ub-F | CCCAAGCTTATGCAGATCTTCGTGAAGACTC (HindIII) | pcDNA3.1-Ub-His | 231 |
| Ub-His-R | CGGGGTACCTTAATGGTGATGGTGATGATGCCCACCTCTGAGACGGAGC (KpnI) |
| Open Reading Frames (ORFs) | Position of RING Domain | Remained Amino Acid Sizes | Subcellular Localization | Ubiquitination in Cell Culture | Ubiquitination of Purified Proteins |
|---|---|---|---|---|---|
| 39L | N-terminal | 1–462 | Whole cell | ++ | nd |
| 52L | C-terminal | 1–650 | Cellular nucleus | + | nd |
| 131R | N-terminal | 1–450 | Cyto-plasmembrane | ++ | ++ |
| 136L | N-terminal | 1–233 | Whole cell | + | ++ |
| 143R | N-terminal | 1–603 | Cyto-plasmembrane | ++ | nd |
| Isolates | |||||
| 131R12–43 | Remain | 12–43 | Whole cell | nd | + |
| 136L45–87 | Remain | 45–87 | Whole cell | nd | + |
| Mutants | |||||
| 131RΔ12–43 | Deletion | 1–11, 44–450 | Cyto-plasmembrane | nd | − |
| 136LΔ45–87 | Deletion | 1–44, 88–450 | Whole cell | nd | − |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, Z.-H.; Ke, F.; Zhang, Q.-Y.; Gui, J.-F. Structural and Functional Diversity among Five RING Finger Proteins from Carassius Auratus Herpesvirus (CaHV). Viruses 2021, 13, 254. https://doi.org/10.3390/v13020254
Wang Z-H, Ke F, Zhang Q-Y, Gui J-F. Structural and Functional Diversity among Five RING Finger Proteins from Carassius Auratus Herpesvirus (CaHV). Viruses. 2021; 13(2):254. https://doi.org/10.3390/v13020254
Chicago/Turabian StyleWang, Zi-Hao, Fei Ke, Qi-Ya Zhang, and Jian-Fang Gui. 2021. "Structural and Functional Diversity among Five RING Finger Proteins from Carassius Auratus Herpesvirus (CaHV)" Viruses 13, no. 2: 254. https://doi.org/10.3390/v13020254
APA StyleWang, Z.-H., Ke, F., Zhang, Q.-Y., & Gui, J.-F. (2021). Structural and Functional Diversity among Five RING Finger Proteins from Carassius Auratus Herpesvirus (CaHV). Viruses, 13(2), 254. https://doi.org/10.3390/v13020254