Follow-Up of Viral Parameters in FeLV- or FIV-Naturally Infected Cats Treated Orally with Low Doses of Human Interferon Alpha

 ,
,
Abstract

1. Introduction
2. Materials and Methods
2.1. Feline Patients and Administration of Treatment
2.2. Clinical Evaluation and Sampling
2.3. Detection of FeLV p27CA, FIV p24CA and Retrotranscriptase Activity (RT)
2.4. Evaluation of FeLV and FIV Proviral load by Real Time PCR (rtPCR)
2.5. Statistical Analysis
3. Results
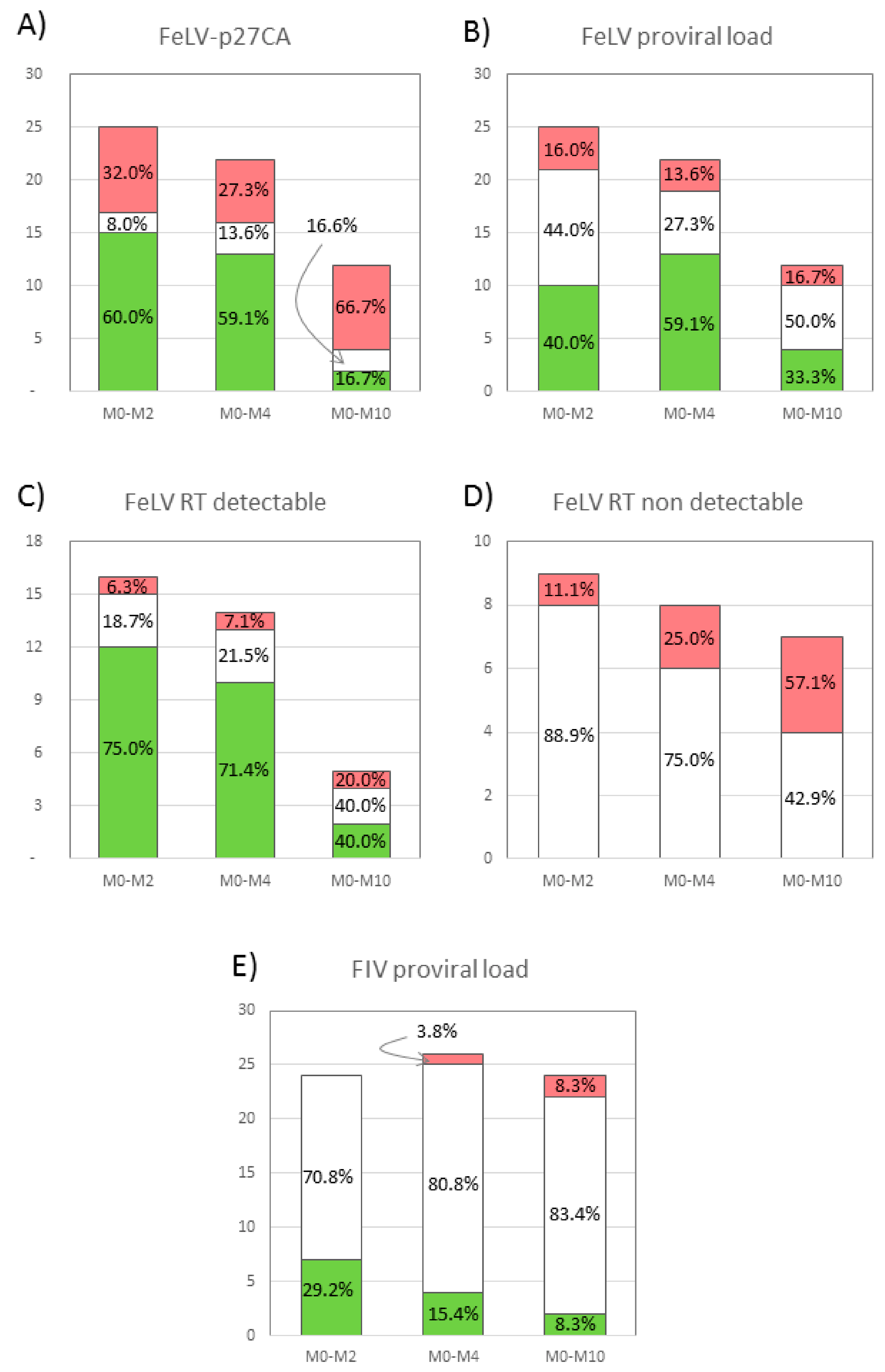
3.1. Plasma Antigenemia
3.2. Reverse Transcriptase (RT) Activity
3.3. Proviral Load
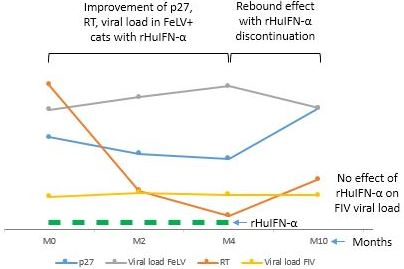
3.4. Combined Results of the Viral Parameters in FeLV+ Cats
4. Discussion
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Bęczkowski, P.M.; Litster, A.; Lin, T.L.; Mellor, D.J.; Willett, B.J.; Hosie, M.J. Contrasting clinical outcomes in two cohorts of cats naturally infected with feline immunodeficiency virus (FIV). Vet. Microbiol. 2015, 176, 50–60. [Google Scholar] [CrossRef] [PubMed]
- Bęczkowski, P.M.; Harris, M.; Techakriengkrai, N.; Beatty, J.A.; Willett, B.J.; Hosie, M.J. Neutralising antibody response in domestic cats immunised with a commercial feline immunodeficiency virus (FIV) vaccine. Vaccine 2015, 33, 977–984. [Google Scholar] [CrossRef] [PubMed]
- Miller, C.; Abdo, Z.; Ericsson, A.; Elder, J.; VandeWoude, S. Applications of the FIV Model to Study HIV Pathogenesis. Viruses 2018, 10, 206. [Google Scholar] [CrossRef] [PubMed]
- Collado, V.M.; Domenech, A.; Miró, G.; Martin, S.; Escolar, E.; Gomez-Lucia, E. Epidemiological aspects and clinicopathological findings in cats naturally infected with feline leukemia virus (FeLV) and/or feline immunodeficiency virus (FIV). Open J. Vet. Med. 2012, 2, 13–20. [Google Scholar] [CrossRef][Green Version]
- Hartmann, K. Clinical aspects of feline retroviruses: A review. Viruses 2012, 4, 2684–2710. [Google Scholar] [CrossRef]
- Dunham, S.P.; Graham, E. Retroviral infections of small animals. Vet. Clin. N. Am. Small Anim. Pract. 2008, 38, 879–901. [Google Scholar] [CrossRef] [PubMed]
- Tejerizo, G.; Domenech, A.; Illera, J.C.; Collado, V.M.; Gomez-Lucia, E. Effect of 17beta-estradiol and progesterone on the expression of FeLV in chronically infected cells. Vet. Microbiol. 2005, 109, 191–199. [Google Scholar] [CrossRef] [PubMed]
- Jarrett, O. Feline leukaemia virus neutralising antibodies. Vet. Rec. 2001, 149, 632. [Google Scholar]
- Hartmann, K. Efficacy of antiviral chemotherapy for retrovirus-infected cats: What does the current literature tell us? J. Feline Med. Surg. 2015, 17, 925–939. [Google Scholar] [CrossRef]
- Gómez-Lucía, E.; Collado, V.M.; Miró, G.; Doménech, A. Effect of type-I interferon on retroviruses. Viruses 2009, 1, 545–573. [Google Scholar] [CrossRef]
- Zeidner, N.S.; Myles, M.H.; Mathiason-DuBard, C.K.; Dreitz, M.J.; Mullins, J.I.; Hoover, E.A. Alpha interferon (2b) in combination with zidovudine for the treatment of presymptomatic feline leukemia virus-induced immunodeficiency syndrome. Antimicrob. Agents Chemother. 1990, 34, 1749–1756. [Google Scholar] [CrossRef] [PubMed]
- Cummins, J.M.; Tompkins, M.B.; Olsen, R.G.; Tompkins, W.A.; Lewis, M.G. Oral use of human alpha interferon in cats. J. Biol. Response Mod. 1988, 7, 513–523. [Google Scholar] [PubMed]
- Collado, V.M.; Gómez-Lucía, E.; Tejerizo, G.; Miró, G.; Escolar, E.; Martín, S.; Doménech, A. Effect of type I interferons on the expression of feline leukaemia virus. Vet. Microbiol. 2007, 123, 180–186. [Google Scholar] [CrossRef] [PubMed]
- Collado Alcalá, V.M. Efecto “in vitro” de Interferón de Tipo I Sobre la Expresión de Retrovirus Felinos y Evaluación de su Aplicación Terapéutica en Gatos con Infección Natural. Ph.D. Thesis, Universidad Complutense de Madrid, Madrid, Spain, 2017. [Google Scholar]
- Stuetzer, B.; Brunner, K.; Lutz, H.; Hartmann, K. A trial with 3′-azido-2′,3′-dideoxythymidine and human interferon-α in cats naturally infected with feline leukaemia virus. J. Feline Med. Surg. 2013, 15, 667–671. [Google Scholar] [CrossRef] [PubMed]
- Doménech, A.; Miró, G.; Collado, V.M.; Ballesteros, N.; Sanjosé, L.; Escolar, E.; Martin, S.; Gomez-Lucia, E. Use of recombinant interferon omega in feline retrovirosis: From theory to practice. Vet. Immunol. Immunopathol. 2011, 143, 301–306. [Google Scholar] [CrossRef] [PubMed]
- Gil, S.; Leal, R.O.; Duarte, A.; McGahie, D.; Sepúlveda, N.; Siborro, I.; Cravo, J.; Cartaxeiro, C.; Tavares, L.M. Relevance of feline interferon omega for clinical improvement and reduction of concurrent viral excretion in retrovirus infected cats from a rescue shelter. Res. Vet. Sci. 2013, 94, 753–763. [Google Scholar] [CrossRef] [PubMed]
- Leal, R.O.; Gil, S.; Duarte, A.; McGahie, D.; Sepúlveda, N.; Niza, M.M.; Tavares, L. Evaluation of viremia, proviral load and cytokine profile in naturally feline immunodeficiency virus infected cats treated with two different protocols of recombinant feline interferon omega. Res. Vet. Sci. 2015, 99, 87–95. [Google Scholar] [CrossRef] [PubMed]
- Pedretti, E.; Passeri, B.; Amadori, M.; Isola, P.; Di Pede, P.; Telera, A.; Vescovini, R.; Quintavalla, F.; Pistello, M. Low-dose interferon-alpha treatment for feline immunodeficiency virus infection. Vet. Immunol. Immunopathol. 2006, 109, 245–254. [Google Scholar] [CrossRef] [PubMed]
- Arjona, A.; Barquero, N.; Doménech, A.; Tejerizo, G.; Collado, V.M.; Toural, C.; Martín, D.; Gomez-Lucia, E. Evaluation of a novel nested PCR for the routine diagnosis of feline leukemia virus (FeLV) and feline immunodeficiency virus (FIV). J. Feline Med. Surg. 2007, 9, 14–22. [Google Scholar] [CrossRef] [PubMed]
- Leutenegger, C.M.; Klein, D.; Hofmann-Lehmann, R.; Mislin, C.; Hummel, U.; Böni, J.; Boretti, F.; Guenzburg, W.H.; Lutz, H. Rapid feline immunodeficiency virus provirus quantitation by polymerase chain reaction using the TaqMan® fluorogenic real-time detection system. J. Virol. Methods 1999, 78, 105–116. [Google Scholar] [CrossRef]
- Pinches, M.; Helps, C.; Gruffyddjones, T.; Egan, K.; Jarrett, O.; Tasker, S. Diagnosis of feline leukaemia virus infection by semi-quantitative real-time polymerase chain reaction. J. Feline Med. Surg. 2007, 9, 8–13. [Google Scholar] [CrossRef] [PubMed]
- Basi, D.L.; Ross, K.F.; Hodges, J.S.; Herzberg, M.C. The Modulation of Tissue Factor by Endothelial Cells during Heat Shock. J. Biol. Chem. 2003, 278, 11065–11071. [Google Scholar] [CrossRef] [PubMed]
- Leal, R.O.; Gil, S.; Sepúlveda, N.; McGahie, D.; Duarte, A.; Niza, M.M.R.E.; Tavares, L. Monitoring acute phase proteins in retrovirus infected cats undergoing feline interferon-ω therapy. J. Small Anim. Pract. 2014, 55, 39–45. [Google Scholar] [CrossRef] [PubMed]
- Cattori, V.; Tandon, R.; Riond, B.; Pepin, A.C.; Lutz, H.; Hofmann-Lehmann, R. The kinetics of feline leukaemia virus shedding in experimentally infected cats are associated with infection outcome. Vet. Microbiol. 2009, 133, 292–296. [Google Scholar] [CrossRef] [PubMed]
- Willett, B.J.; Hosie, M.J. Feline leukaemia virus: Half a century since its discovery. Vet. J. 2013, 195, 16–23. [Google Scholar] [CrossRef] [PubMed]
- Boesch, A.; Cattori, V.; Riond, B.; Willi, B.; Meli, M.L.; Rentsch, K.M.; Hosie, M.J.; Hofmann-Lehmann, R.; Lutz, H. Evaluation of the effect of short-term treatment with the integrase inhibitor raltegravir (Isentress) on the course of progressive feline leukemia virus infection. Vet. Microbiol. 2015, 175, 167–178. [Google Scholar] [CrossRef] [PubMed]
- Feng, X.; Heyden, N.V.; Ratner, L. Alpha interferon inhibits human T-cell leukemia virus type 1 assembly by preventing Gag interaction with rafts. J. Virol. 2003, 77, 13389–13395. [Google Scholar] [CrossRef] [PubMed]
- Haas, D.W.; Lavelle, J.; Nadler, J.P.; Greenberg, S.B.; Frame, P.; Mustafa, N.; St Clair, M.; McKinnis, R.; Dix, L.; Elkins, M.; et al. A randomized trial of interferon alpha therapy for HIV type 1 infection. AIDS Res. Hum. Retrovir. 2000, 16, 183–190. [Google Scholar] [CrossRef] [PubMed]
- Murphy, B.; Hillman, C.; Castillo, D.; Vapniarsky, N.; Rowe, J. The presence or absence of the gamma-activated site determines IFN gamma-mediated transcriptional activation in CAEV promoters cloned from the mammary gland and joint synovium of a single CAEV-infected goat. Virus Res. 2012, 163, 537–545. [Google Scholar] [CrossRef]
- Eckstrand, C.D.; Sparger, E.E.; Pitt, K.A.; Murphy, B.G. Peripheral and central immune cell reservoirs in tissues from asymptomatic cats chronically infected with feline immunodeficiency virus. PLoS ONE 2017, 12, e0175327. [Google Scholar] [CrossRef]
- Bęczkowski, P.M.; Logan, N.; McMonagle, E.; Litster, A.; Willett, B.J.; Hosie, M.J. An investigation of the breadth of neutralizing antibody response in cats naturally infected with feline immunodeficiency virus. J. Gen. Virol. 2015, 96, 671–680. [Google Scholar] [CrossRef] [PubMed]
- McDonnel, S.J.; Liepnieks, M.L.; Murphy, B.G. Treatment of chronically FIV-infected cats with suberoylanilide hydroxamic acid. Antivir. Res. 2014, 108, 74–78. [Google Scholar] [CrossRef] [PubMed]
- Gómez, N.V.; Fontanals, A.; Castillo, V.; Gisbert, M.A.; Suraniti, A.; Mira, G.; Pisano, P.B. Evaluation of different antiretroviral drug protocols on naturally infected feline immunodeficiency virus (FIV) cats in the late phase of the asymptomatic stage of infection. Viruses 2012, 4, 924–939. [Google Scholar] [CrossRef] [PubMed]
- Yokota, M.; Tatsumi, N.; Nathalang, O.; Yamada, T.; Tsuda, I. Effects of Heparin on Polymerase Chain Reaction for Blood White Cells. Available online: https://onlinelibrary.wiley.com/doi/abs/10.1002/(SICI)1098-2825(1999)13:3%3C133::AID-JCLA8%3E3.0.CO;2-0 (accessed on 19 August 2019).



{kind=link}
{kind=link}
{kind=link}
| CS | p27CA | RT | Proviral Load | |
|---|---|---|---|---|
| CG1 | ||||
| M0 | 0.00 (0.00) | 0.60 (0.13) | 0.86 (0.52) | 0.72 (0.04) |
| M2 | 0.00 (0.00) | 0.49 (0.08) | 0.11 (0.09) | 0.78 (0.05) |
| M4 | 0.00 (0.00) | 0.55 (0.18) | 0.12 (0.12) | 0.88 (0.10) |
| M10 | 0.00 (0.00) | 0.63 (0.16) | 0.65 (0.39) | 0.66 (0.06) |
| CG2 | ||||
| M0 | 3.50 (0.05) | 0.46 (0.13) | 1.22 (0.59) | 0.61 (0.06) |
| M2 | 3.25 (0.84) | 0.22 (0.07) | 0.28 (0.28) | 0.72 (0.06) |
| M4 | 1.75 (0.62) | 0.40 (0.17) | 0.62 (0.40) | 0.70 (0.08) |
| M10 | 2.00 (0.00) | 0.85 (0.19) | 3.29 (3.29) | 0.77 (0.04) |
| CG3 | ||||
| M0 | 8.00 (0.84) | 0.49 (0.08) | 3.13 (1.46) | 0.73 (0.06) |
| M2 | 6.17 (1.48) | 0.58 (0.16) | 2.51 (1.55) | 0.81 (0.07) |
| M4 | 3.00 (1.91) | 0.20 (0.10) | 0.25 (0.15) | 0.88 (0.04) |
| M10 | 0.00 (0.00) | 0.66 (0.40) | 0.40 (0.37) | 0.69 (0.03) |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gomez-Lucia, E.; Collado, V.M.; Miró, G.; Martín, S.; Benítez, L.; Doménech, A. Follow-Up of Viral Parameters in FeLV- or FIV-Naturally Infected Cats Treated Orally with Low Doses of Human Interferon Alpha. Viruses 2019, 11, 845. https://doi.org/10.3390/v11090845
Gomez-Lucia E, Collado VM, Miró G, Martín S, Benítez L, Doménech A. Follow-Up of Viral Parameters in FeLV- or FIV-Naturally Infected Cats Treated Orally with Low Doses of Human Interferon Alpha. Viruses. 2019; 11(9):845. https://doi.org/10.3390/v11090845
Chicago/Turabian StyleGomez-Lucia, Esperanza, Victorio M. Collado, Guadalupe Miró, Sonsoles Martín, Laura Benítez, and Ana Doménech. 2019. "Follow-Up of Viral Parameters in FeLV- or FIV-Naturally Infected Cats Treated Orally with Low Doses of Human Interferon Alpha" Viruses 11, no. 9: 845. https://doi.org/10.3390/v11090845
APA StyleGomez-Lucia, E., Collado, V. M., Miró, G., Martín, S., Benítez, L., & Doménech, A. (2019). Follow-Up of Viral Parameters in FeLV- or FIV-Naturally Infected Cats Treated Orally with Low Doses of Human Interferon Alpha. Viruses, 11(9), 845. https://doi.org/10.3390/v11090845