High-Throughput Sequencing Analysis of Small RNAs Derived from Coleus Blumei Viroids


Abstract
1. Introduction
2. Materials and Methods
2.1. Sample Preparation
2.2. RT-PCR and Northern Blot Hybridization
2.3. Cloning and Sequencing
2.4. Library Construction and sRNA Sequencing
2.5. Bioinformatic Analysis
3. Results
3.1. sRNA Sequencing and Quality Control
3.2. Detection of CbVd-1, -5, and -6 by sRNA Sequencing
3.3. Reference Sequences of CbVd-1, -5, and -6 for Read Mapping
3.4. Characterization of sRNAs Derived from CbVd-1, -5, and -6
3.4.1. Polarity
3.4.2. Size Distribution
3.4.3. 5’-terminal nucleotide
3.5. Distribution of sRNAs Derived from CbVd-1, -5, and -6 along Their Genomes
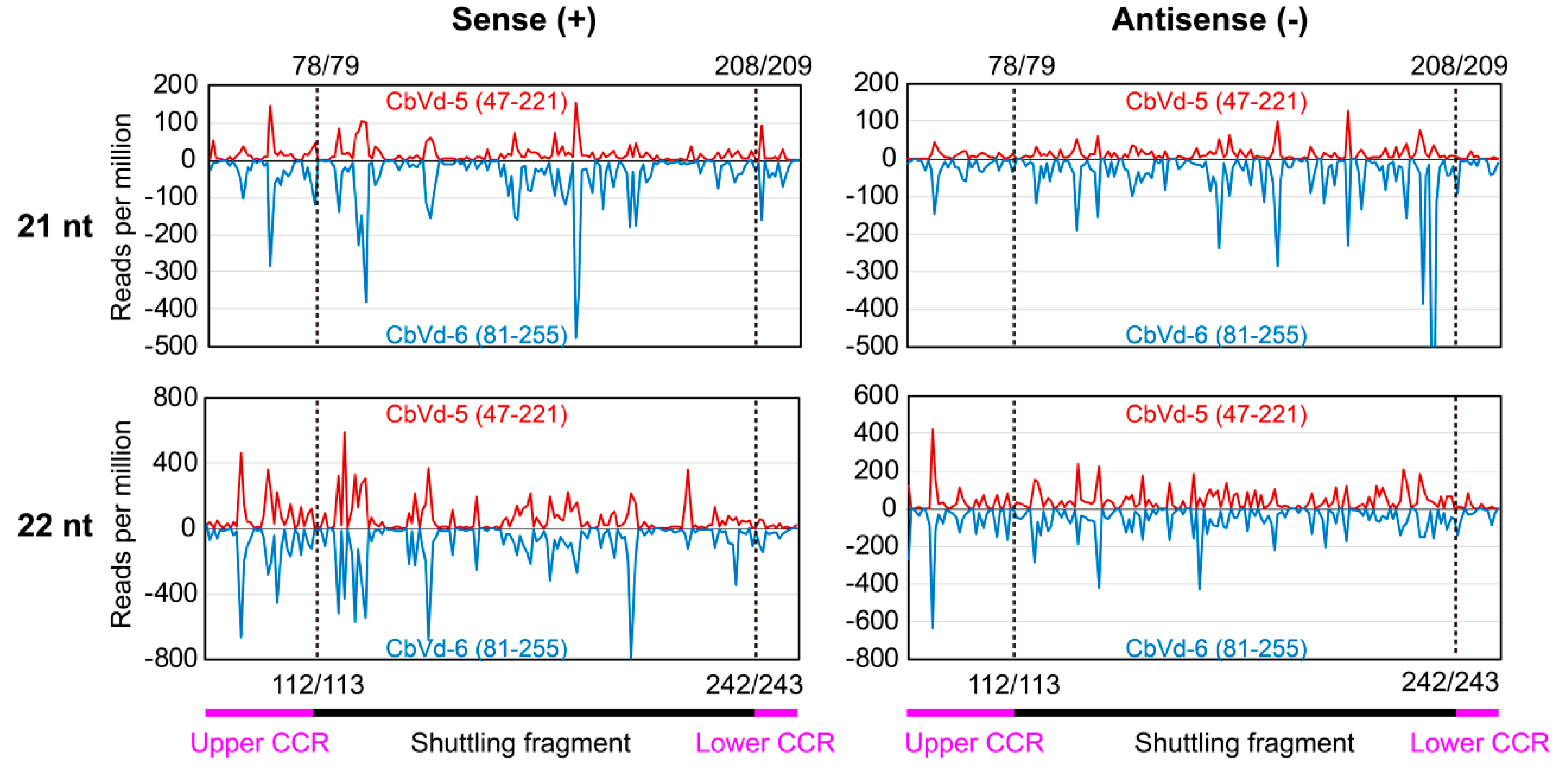
3.6. vd-sRNA Distribution on the Shuttling Genomic Fragment between CbVd-5 and -6
4. Discussion
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Nie, X.Z.; Singh, R.P. Chapter 27—coleus blumei viroids. In Viroids and satellites; Flores, R., Randles, J.W., Palukaitis, P., Eds.; Academic Press: Boston, MA, USA, 2017; pp. 289–295. [Google Scholar]
- Di Serio, F.; Flores, R.; Verhoeven, J.T.J.; Li, S.F.; Pallas, V.; Randles, J.W.; Sano, T.; Vidalakis, G.; Owens, R.A. Current status of viroid taxonomy. Arch. Virol. 2014, 159, 3467–3478. [Google Scholar] [CrossRef]
- Hou, W.Y.; Sano, T.; Li, F.; Wu, Z.J.; Li, L.; Li, S.F. Identification and characterization of a new coleviroid (cbvd-5). Arch. Virol. 2009, 154, 315–320. [Google Scholar] [CrossRef]
- Hou, W.Y.; Li, S.F.; Wu, Z.J.; Jiang, D.M.; Sano, T. Coleus blumei viroid 6: A new tentative member of the genus Coleviroid derived from natural genome shuffling. Arch. Virol. 2009, 154, 993–997. [Google Scholar] [CrossRef]
- Spieker, R.L. In vitro-generated ‘inverse’ chimeric coleus blumei viroids evolve in vivo into infectious RNA replicons. J. Gen. Virol. 1996, 77, 2839–2846. [Google Scholar] [CrossRef]
- Tsushima, T.; Sano, T. A point-mutation of coleus blumei viroid 1 switches the potential to transmit through seed. J. Gen. Virol. 2018, 99, 393–401. [Google Scholar] [CrossRef]
- Hadidi, A.; Flores, R.; Candresse, T.; Barba, M. Next-generation sequencing and genome editing in plant virology. Front. Microbiol. 2016, 7, 1325. [Google Scholar] [CrossRef]
- Barba, M.; Hadidi, A. Chapter 38—Application of next-generation sequencing technologies to viroids. In Viroids and satellites; Flores, R., Randles, J.W., Palukaitis, P., Eds.; Academic Press: Boston, MA, USA, 2017; pp. 401–412. [Google Scholar]
- Di Serio, F.; Gisel, A.; Navarro, B.; Delgado, S.; de Alba, A.E.M.; Donvito, G.; Flores, R. Deep sequencing of the small RNAs derived from two symptomatic variants of a chloroplastic viroid:Implications for their genesis and for pathogenesis. PLoS ONE 2009, 4, e7539. [Google Scholar] [CrossRef]
- Navarro, B.; Pantaleo, V.; Gisel, A.; Moxon, S.; Dalmay, T.; Bisztray, G.; di Serio, F.; Burgyan, J. Deep sequencing of viroid-derived small RNAs from grapevine provides new insights on the role of RNA silencing in plant-viroid interaction. PLoS ONE 2009, 4, e7686. [Google Scholar] [CrossRef]
- Martinez, G.; Donaire, L.; Llave, C.; Pallas, V.; Gomez, G. High-throughput sequencing of hop stunt viroid-derived small RNAs from cucumber leaves and phloem. Mol. Plant Pathol. 2010, 11, 347–359. [Google Scholar] [CrossRef]
- Bolduc, F.; Hoareau, C.; St-Pierre, P.; Perreault, J.P. In-depth sequencing of the siRNAs associated with peach latent mosaic viroid infection. BMC Mol. Biol. 2010, 11, 16. [Google Scholar] [CrossRef]
- Wang, Y.; Shibuya, M.; Taneda, A.; Kurauchi, T.; Senda, M.; Owens, R.A.; Sano, T. Accumulation of potato spindle tuber viroid-specific small RNAs is accompanied by specific changes in gene expression in two tomato cultivars. Virology 2011, 413, 72–83. [Google Scholar] [CrossRef]
- Di Serio, F.; de Alba, A.E.M.; Navarro, B.; Gisel, A.; Flores, R. RNA-dependent RNA polymerase 6 delays accumulation and precludes meristem invasion of a viroid that replicates in the nucleus. J. Virol. 2010, 84, 2477–2489. [Google Scholar] [CrossRef]
- Wu, Q.F.; Ding, S.W.; Zhang, Y.J.; Zhu, S.F. Identification of viruses and viroids by next-generation sequencing and homology-dependent and homology-independent algorithms. Ann. Rev. Phytopathol. 2015, 53, 425–444. [Google Scholar] [CrossRef]
- Massart, S.; Chiumenti, M.; de Jonghe, K.; Glover, R.; Haegeman, A.; Koloniuk, I.; Kominek, P.; Kreuze, J.; Kutnjak, D.; Lotos, L.; et al. Virus detection by high-throughput sequencing of small RNAs: Large scale performance testing of sequence analysis strategies. Phytopathology 2019, 109, 488–497. [Google Scholar] [CrossRef]
- Chiumenti, M.; Torchetti, E.M.; di Serio, F.; Minafra, A. Identification and characterization of a viroid resembling apple dimple fruit viroid in fig (ficus carica l.) by next generation sequencing of small RNAs. Virus Res. 2014, 188, 54–59. [Google Scholar] [CrossRef]
- Li, R.G.; Gao, S.; Hernandez, A.G.; Wechter, W.P.; Fei, Z.J.; Ling, K.S. Deep sequencing of small RNAs in tomato for virus and viroid identification and strain differentiation. PLoS ONE 2012, 7, e37127. [Google Scholar] [CrossRef]
- Jakse, J.; Radisek, S.; Pokorn, T.; Matousek, J.; Javornik, B. Deep-sequencing revealed citrus bark cracking viroid (cbcvd) as a highly aggressive pathogen on hop. Plant Pathol. 2015, 64, 831–842. [Google Scholar] [CrossRef]
- Wan, Y.F.; Atta, S.; Wang, X.F.; Yang, F.Y.; Zhou, C.Y.; Cao, M.J. Transcriptome sequencing reveals novel citrus bark cracking viroid (cbcvd) variants from citrus and their molecular characterization. PLoS ONE 2018, 13, e0198022. [Google Scholar]
- Zhang, Z.X.; Qi, S.S.; Tang, N.; Zhang, X.X.; Chen, S.S.; Zhu, P.F.; Ma, L.; Cheng, J.P.; Xu, Y.; Lu, M.G.; et al. Discovery of replicating circular RNAs by RNA-seq and computational algorithms. PLoS Pathog. 2014, 10, e1004553. [Google Scholar] [CrossRef]
- Wu, Q.F.; Wang, Y.; Cao, M.J.; Pantaleo, V.; Burgyan, J.; Li, W.X.; Ding, S.W. Homology-independent discovery of replicating pathogenic circular RNAs by deep sequencing and a new computational algorithm. Proc. Natl. Acad. Sci. USA 2012, 109, 3938–3943. [Google Scholar] [CrossRef]
- Jiang, D.M.; Gao, R.; Qin, L.; Wu, Z.J.; Xie, L.H.; Hou, W.Y.; Li, S.F. Infectious cDNA clones of four viroids in coleus blumei and molecular characterization of their progeny. Virus Res. 2014, 180, 97–101. [Google Scholar] [CrossRef]
- Jiang, D.M.; Wu, Z.J.; Xie, L.H.; Sano, T.; Li, S.F. Sap-direct RT-PCR for the rapid detection of coleus blumei viroids of the genus Coleviroid from natural host plants. J. Virol. Methods 2011, 174, 123–127. [Google Scholar] [CrossRef]
- Jiang, D.M.; Hou, W.Y.; Sano, T.; Kang, N.; Qin, L.; Wu, Z.J.; Li, S.F.; Xie, L.H. Rapid detection and identification of viroids in the genus Coleviroid using a universal probe. J. Virol. Methods 2013, 187, 321–326. [Google Scholar] [CrossRef]
- Li, S.F.; Su, Q.; Guo, R.; Tsuji, M.; Sano, T. First report of coleus blumei viroid from coleus in china. Plant Pathol. 2006, 55, 565. [Google Scholar] [CrossRef]
- Bolger, A.M.; Lohse, M.; Usadel, B. Trimmomatic: A flexible trimmer for illumina sequence data. Bioinformatics 2014, 30, 2114–2120. [Google Scholar] [CrossRef]
- Zerbino, D.R.; Birney, E. Velvet: Algorithms for de novo short read assembly using de bruijn graphs. Genome Res. 2008, 18, 821–829. [Google Scholar] [CrossRef]
- Brister, J.R.; Ako-adjei, D.; Bao, Y.M.; Blinkova, O. NCBI viral genomes resource. Nucleic Acids Res. 2015, 43, D571–D577. [Google Scholar] [CrossRef]
- Altschul, S.F.; Gish, W.; Miller, W.; Myers, E.W.; Lipman, D.J. Basic local alignment search tool. J. Mol. Biol. 1990, 215, 403–410. [Google Scholar] [CrossRef]
- Langmead, B.; Salzberg, S.L. Fast gapped-read alignment with bowtie 2. Nat. Methods 2012, 9, 357–359. [Google Scholar] [CrossRef]
- Domingo, E.; Sheldon, J.; Perales, C. Viral quasispecies evolution. Microbiol. Mol. Biol. R. 2012, 76, 159–216. [Google Scholar] [CrossRef]
- Giguere, T.; Raj Adkar-Purushothama, C.; Perreault, J.P. Comprehensive secondary structure elucidation of four genera of the family Pospiviroidae. PLoS ONE 2014, 9, e98655. [Google Scholar] [CrossRef]
- Pooggin, M.M. Small RNA-omics for plant virus identification, virome reconstruction, and antiviral defense characterization. Front. Microbiol. 2018, 9, 2779. [Google Scholar] [CrossRef]
- Serra, P.; Messmer, A.; Sanderson, D.; James, D.; Flores, R. Apple hammerhead viroid-like RNA is a bona fide viroid: Autonomous replication and structural features support its inclusion as a new member in the genus pelamoviroid. Virus Res. 2018, 249, 8–15. [Google Scholar] [CrossRef]
- Mi, S.J.; Cai, T.; Hu, Y.G.; Chen, Y.; Hodges, E.; Ni, F.R.; Wu, L.; Li, S.; Zhou, H.; Long, C.Z.; et al. Sorting of small RNAs into arabidopsis argonaute complexes is directed by the 5 ‘ terminal nucleotide. Cell 2008, 133, 116–127. [Google Scholar] [CrossRef]
- Minoia, S.; Carbonell, A.; di Serio, F.; Gisel, A.; Carrington, J.C.; Navarro, B.; Flores, R. Specific argonautes selectively bind small RNAs derived from potato spindle tuber viroid and attenuate viroid accumulation in vivo. J. Virol. 2014, 88, 11933–11945. [Google Scholar] [CrossRef]
- Gas, M.E.; Hernandez, C.; Flores, R.; Daros, J.A. Processing of nuclear viroids in vivo: An interplay between RNA conformations. PLoS Pathog. 2007, 3, e182. [Google Scholar] [CrossRef]
- Seguin, J.; Rajeswaran, R.; Malpica-Lopez, N.; Martin, R.R.; Kasschau, K.; Dolja, V.V.; Otten, P.; Farinelli, L.; Pooggin, M.M. De novo reconstruction of consensus master genomes of plant RNA and DNA viruses from siRNAs. PLoS ONE 2014, 9, e88513. [Google Scholar] [CrossRef]
- Flores, R.; Minoia, S.; Carbonell, A.; Gisel, A.; Delgado, S.; Lopez-Carrasco, A.; Navarro, B.; di Serio, F. Viroids, the simplest RNA replicons: How they manipulate their hosts for being propagated and how their hosts react for containing the infection. Virus Res. 2015, 209, 136–145. [Google Scholar] [CrossRef]
- Katsarou, K.; Mavrothalassiti, E.; Dermauw, W.; van Leeuwen, T.; Kalantidis, K. Combined activity of dcl2 and dcl3 is crucial in the defense against potato spindle tuber viroid. PLoS Pathog. 2016, 12, 10. [Google Scholar] [CrossRef]
- Dadami, E.; Boutla, A.; Vrettos, N.; Tzortzakaki, S.; Karakasilioti, I.; Kalantidis, K. Dicer-like 4 but not dicer-like 2 may have a positive effect on potato spindle tuber viroid accumulation in nicotiana benthamiana. Mol. plant 2013, 6, 232–234. [Google Scholar] [CrossRef]
- Leibman, D.; Wolf, D.; Saharan, V.; Zelcer, A.; Arazi, T.; Yoel, S.; Gaba, V.; Gal-On, A. A high level of transgenic viral small RNA is associated with broad potyvirus resistance in cucurbits. Mol. Plant Microbe In. 2011, 24, 1220–1238. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Viroid | Total Reads (Million) | Clean Reads (Million) | vd-sRNAs # | ||
|---|---|---|---|---|---|
| Total (%) | + (%) | − (%) | |||
| CbVd-1 * | 18.53 | 17.53 | 700,128 (3.994) | 312,876 (1.785) | 387,252 (2.209) |
| CbVd-1 | 20.11 | 19.41 | 568,154 (2.927) | 243,169 (1.253) | 324,985 (1.674) |
| CbVd-5 * | 18.53 | 17.53 | 1,006,161 (5.74) | 566,058 (3.229) | 440,103 (2.511) |
| CbVd-5 | 20.57 | 19.86 | 562,821 (2.834) | 323,465 (1.629) | 239,356 (1.205) |
| CbVd-6 | 16.55 | 15.95 | 789,359 (4.949) | 444,061 (2.784) | 345,298 (2.165) |
| Viroid | Velvet | PFOR2 | ||||
|---|---|---|---|---|---|---|
| Number | Length (nt) | Coverage | Number | Length (nt) | Coverage | |
| CbVd-1 | 28 | 31, 33 | 71.4% | 4 | 247–250 | 100% |
| CbVd-5 | 42 | 33, 53 | 64.2% | 2 | 274 | 100% |
| CbVd-6 | 38 | 31, 33 | 75.4% | 3 | 341–342 | 100% |
| CbVd-1 * | 9 | 33 | 66.7% | 1 | 247 | 100% |
| CbVd-5 * | 4 | 33 | 48.2% | 1 | 274 | 100% |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Jiang, D.-M.; Wang, M.; Li, S.-F.; Zhang, Z.-X. High-Throughput Sequencing Analysis of Small RNAs Derived from Coleus Blumei Viroids. Viruses 2019, 11, 619. https://doi.org/10.3390/v11070619
Jiang D-M, Wang M, Li S-F, Zhang Z-X. High-Throughput Sequencing Analysis of Small RNAs Derived from Coleus Blumei Viroids. Viruses. 2019; 11(7):619. https://doi.org/10.3390/v11070619
Chicago/Turabian StyleJiang, Dong-Mei, Meng Wang, Shi-Fang Li, and Zhi-Xiang Zhang. 2019. "High-Throughput Sequencing Analysis of Small RNAs Derived from Coleus Blumei Viroids" Viruses 11, no. 7: 619. https://doi.org/10.3390/v11070619
APA StyleJiang, D.-M., Wang, M., Li, S.-F., & Zhang, Z.-X. (2019). High-Throughput Sequencing Analysis of Small RNAs Derived from Coleus Blumei Viroids. Viruses, 11(7), 619. https://doi.org/10.3390/v11070619