Molecular Characterization of a Novel Endornavirus Conferring Hypovirulence in Rice Sheath Blight Fungus Rhizoctonia solani AG-1 IA Strain GD-2


Abstract
1. Introduction
2. Materials and Methods
2.1. Fungal Strains and Cultural Conditions
2.2. Extraction of dsRNA
2.3. cDNA Library Construction, Illumina Sequencing, and Sequence Analysis
2.4. Transmission of Mycovirus
2.5. Metabolomics Analysis
2.6. Statistical Analysis
3. Results
3.1. Nucleotide Sequence and Amino Acid Sequence of RsEV1
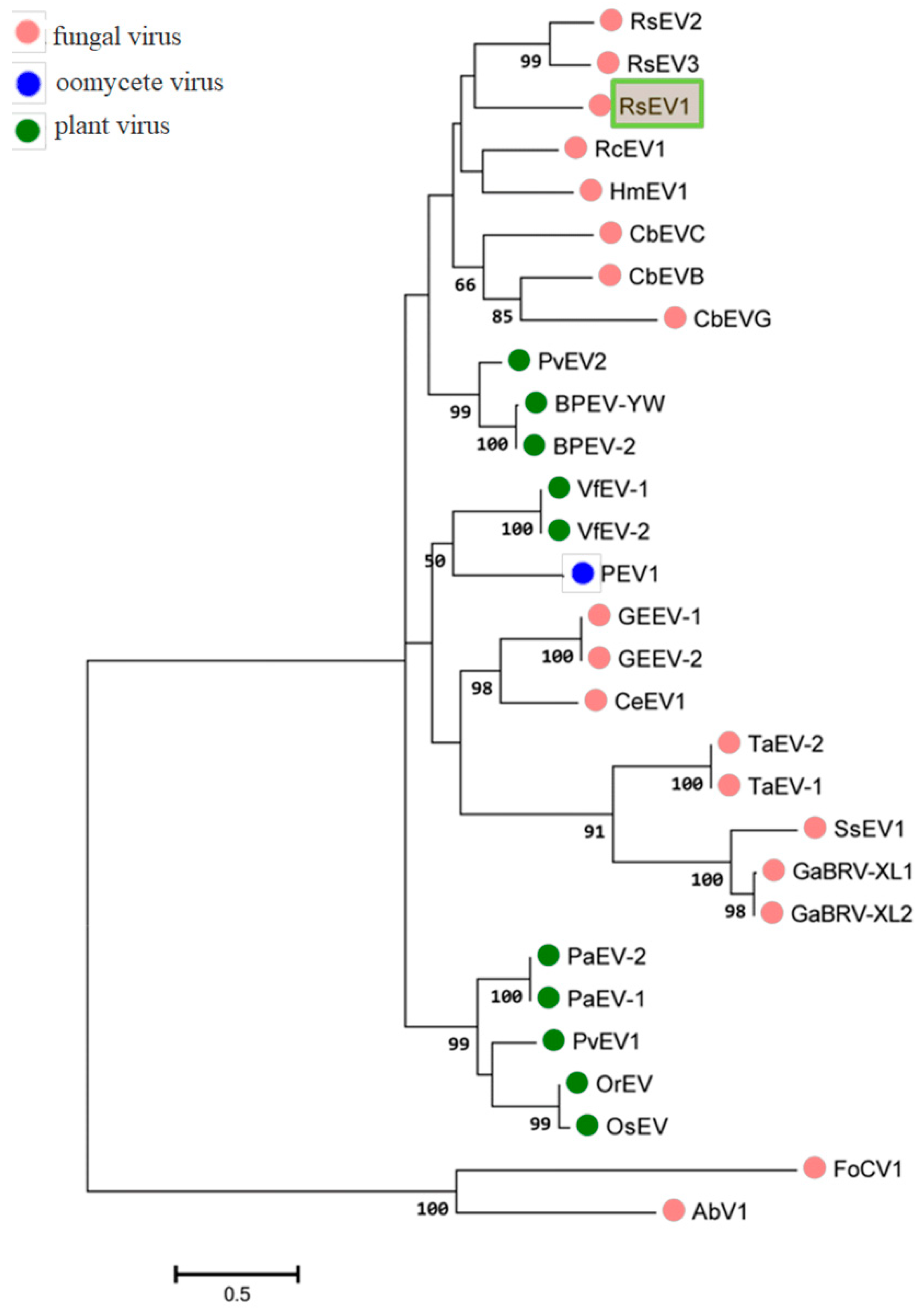
3.2. Phylogenetic Analyses
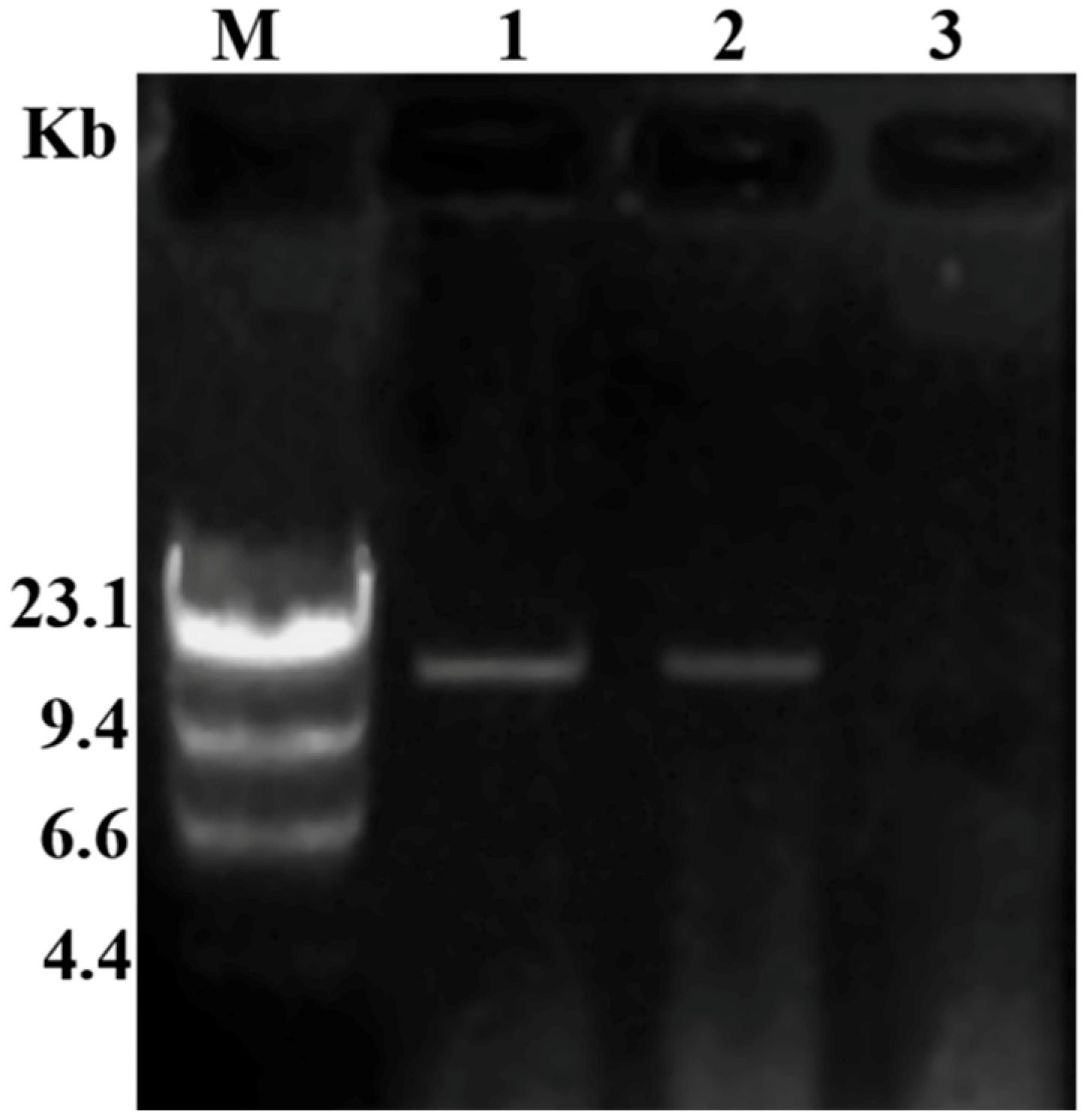
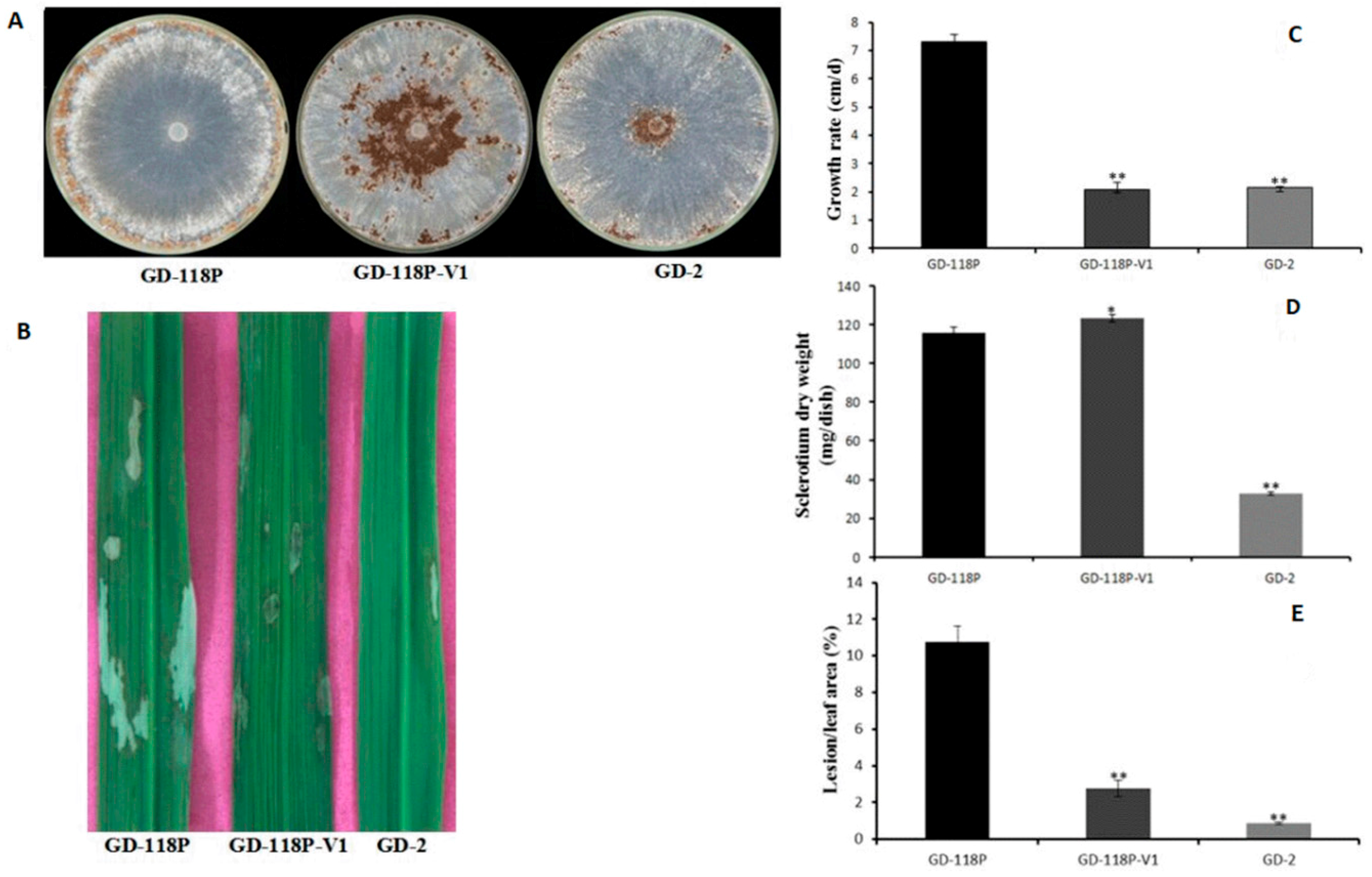
3.3. Transmission of Hypovirulence and DsRNA
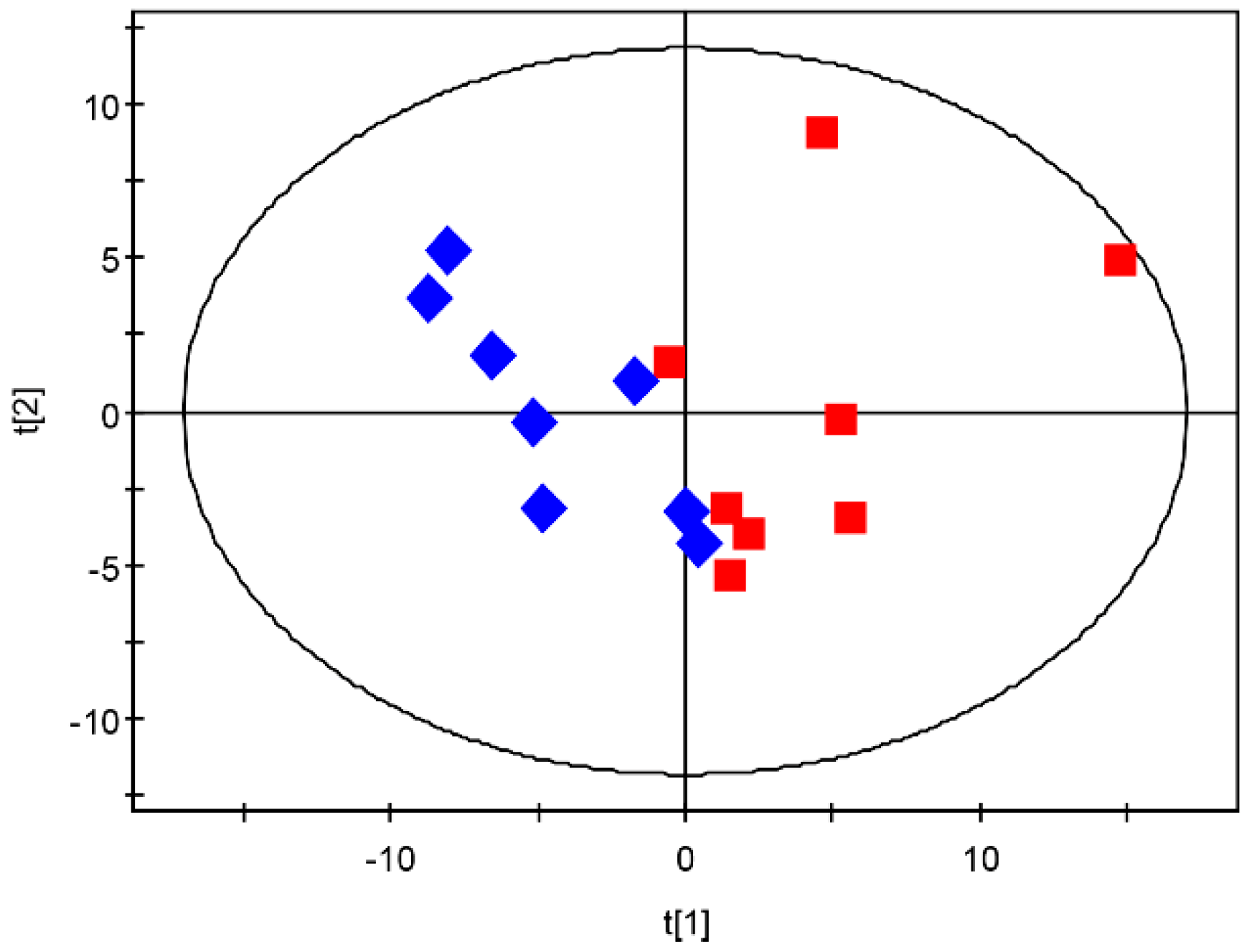
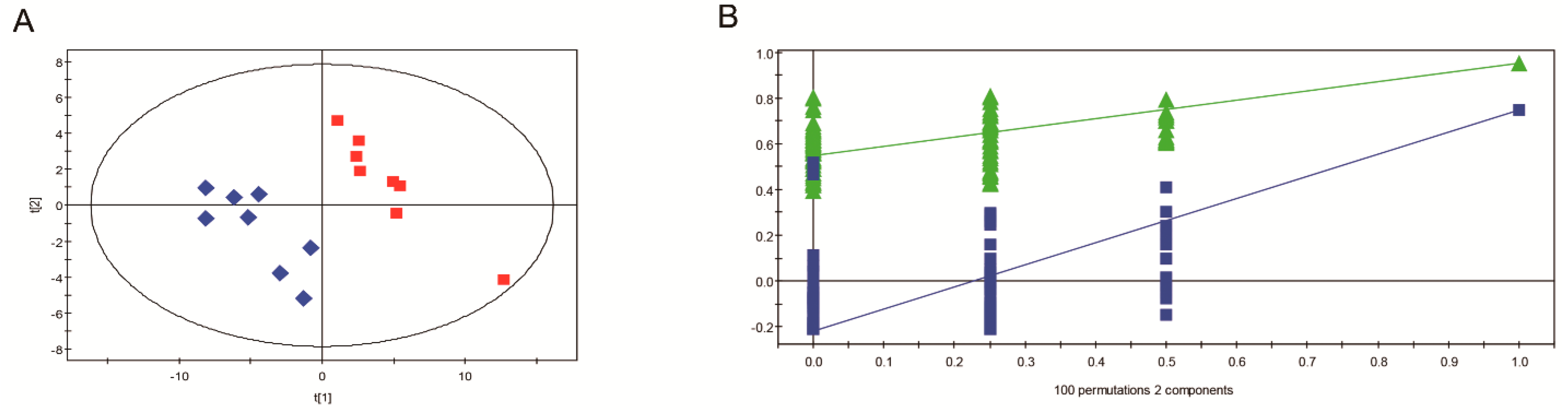
3.4. Overview of Metabolomic Profiling
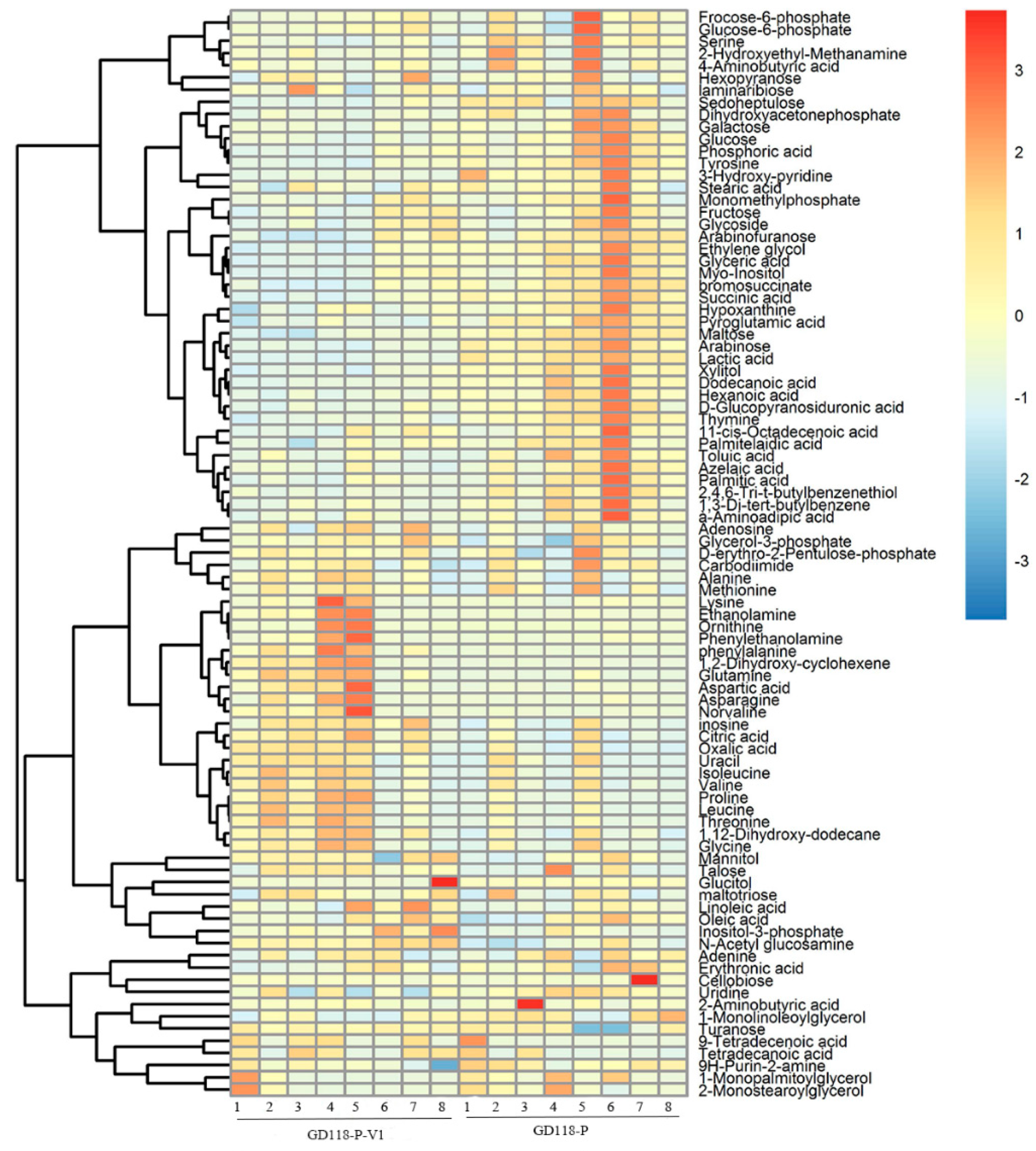
3.5. Metabolic Profile Comparison
3.6. Metabolite Changes between GD-118P and GD-118P-V1
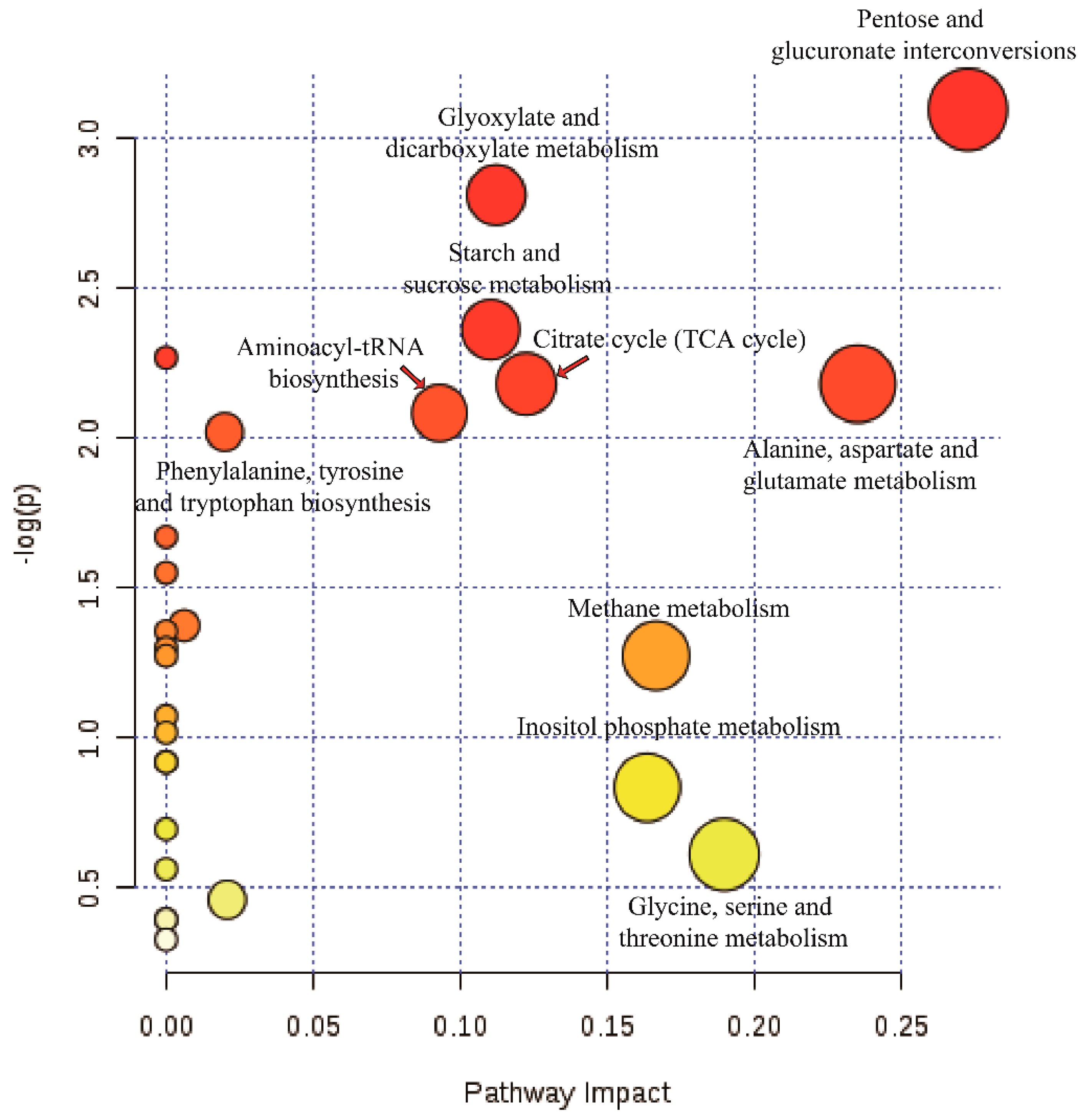
3.7. Pathway Annotation of the Differential Metabolites
4. Discussion
Author Contributions
Funding
Conflicts of Interest
References
- Ghabrial, S.A.; Suzuki, N. Viruses of plant pathogenic fungi. Annu. Rev. Phytopathol. 2009, 47, 353–384. [Google Scholar] [CrossRef]
- Pearson, M.N.; Beever, R.E.; Boine, B.; Arthur, K. Mycoviruses of filamentous fungi and their relevance to plant pathology. Mol. Plant. Pathol. 2009, 10, 115–128. [Google Scholar] [CrossRef] [PubMed]
- Ghabrial, S.A.; Castón, J.R.; Jiang, D.; Nibert, M.L.; Suzuki, N. 50-plus years of fungal viruses. Virology 2015, 356, 479–480. [Google Scholar] [CrossRef]
- Xie, J.; Jiang, D. New insights into mycoviruses and exploration for the biological control of crop fungal diseases. Annu. Rev. Phytopathol. 2014, 52, 45–68. [Google Scholar] [CrossRef] [PubMed]
- Nuss, D.L. Hypovirulence: mycoviruses at the fungal-plant interface. Nat. Rev. Microbiol. 2005, 3, 632–642. [Google Scholar] [CrossRef] [PubMed]
- Yu, X.; Li, B.; Fu, Y.; Xie, J.; Cheng, J.; Ghabrial, S.A.; Li, G.; Yi, X.; Jiang, D. Extracellular transmission of a DNA mycovirus and its use as a natural fungicide. Proc. Natl. Acad. Sci. USA 2013, 110, 1452–1457. [Google Scholar] [CrossRef] [PubMed]
- Roossinck, M.J.; Sabanadzovic, S.; Okada, R.; Valverde, R.A. The remarkable evolutionary history of endornaviruses. J. Gen. Virol. 2011, 92, 2674–2678. [Google Scholar] [CrossRef]
- Okada, R.; Yong, C.K.; Valverde, R.A.; Sabanadzovic, S.; Aoki, N.; Hotate, S.; Kiyota, E.; Moriyama, H.; Fukuhara, T. Molecular characterization of two evolutionarily distinct endornaviruses co-infecting common bean (Phaseolus vulgaris). J. Gen. Virol. 2013, 94, 220–229. [Google Scholar] [CrossRef]
- Ikeda, K.I.; Nakamura, H.; Matsumoto, N. Hypovirulent strain of the violet root rot fungus Helicobasidium mompa. J. Gen. Plant Pathol. 2003, 69, 385–390. [Google Scholar] [CrossRef]
- Tuomivirta, T.T.; Kaitera, J.; Hantula, J. A novel putative virus of Gremmeniella abietina type B (Ascomycota: Helotiaceae) has a composite genome with endornavirus affinities. J. Gen. Virol. 2009, 90, 2299–2305. [Google Scholar] [CrossRef]
- Ong, J.W.; Li, H.; Sivasithamparam, K.; Dixon, K.W.; Jones, M.G.; Wylie, S.J. Novel Endorna-like viruses, including three with two open reading frames, challenge the membership criteria and taxonomy of the Endornaviridae. Virology 2016, 499, 203–211. [Google Scholar] [CrossRef] [PubMed]
- Grill, L.K.; Garger, S.J. Identification and characterization of double-stranded RNA associated with cytoplasmic male sterility in Vicia faba. Proc. Natl. Acad. Sci. USA 1981, 78, 7043–7046. [Google Scholar] [CrossRef] [PubMed]
- Osaki, H.; Nakamura, H.; Sasaki, A.; Matsumoto, N.; Yoshida, K. An endornavirus from a hypovirulent strain of the violet root rot fungus, Helicobasidium mompa. Virus Res. 2006, 118, 143–149. [Google Scholar] [CrossRef] [PubMed]
- Zheng, L.; Zhang, M.; Chen, Q.; Zhu, M.; Zhou, E. A novel mycovirus closely related to viruses in the genus Alphapartitivirus confers hypovirulence in the phytopathogenic fungus Rhizoctonia solani. Virology 2014, 456–457, 220–226. [Google Scholar] [CrossRef] [PubMed]
- Strauss, E.E.; Lakshman, D.K.; Tavantzis, S.M. Molecular characterization of the genome of a partitivirus from the basidiomycete Rhizoctonia solani. J. Gen. Virol. 2000, 81, 255–549. [Google Scholar] [CrossRef] [PubMed]
- Castanho, B. The association of double-stranded RNA with Rhizoctonia decline. Phytopathology 1978, 68, 1515–1519. [Google Scholar] [CrossRef]
- Jian, J.H.; Lakshman, D.K.; Tavantzis, S.M. Association of distinct double-stranded RNAs with enhanced or diminished virulence in Rhizoctonia solani infecting potato. Mol. Plant. Microbe. Interact. 1997, 10, 1002–1009. [Google Scholar] [CrossRef]
- Liu, C.; Al, E. Quinic acid induces hypovirulence and expression of a hypovirulence-associated double-stranded RNA in Rhizoctonia solani. Curr. Genet. 2003, 43, 103–111. [Google Scholar]
- Bharathan, N.; Saso, H.; Gudipati, L.; Bharathan, S.; Whited, K.; Anthony, K. Double-stranded RNA: distribution and analysis among isolates of Rhizoctonia solani AG-2 to -13. Plant Pathol. 2005, 54, 196–203. [Google Scholar] [CrossRef]
- Zheng, L.; Liu, C.; Zhang, M.; Yang, M.; Zhou, E. Diversity of dsRNA viruses infecting rice sheath blight fungus Rhizoctonia solani AG-1 IA. Rice Sci. 2018, 25, 57–60. [Google Scholar]
- Zheng, L.; Liu, H.; Zhang, M.; Cao, X.; Zhou, E. The complete genomic sequence of a novel mycovirus from Rhizoctonia solani AG-1 IA strain B275. Arch. Virol. 2013, 158, 1609–1612. [Google Scholar] [CrossRef] [PubMed]
- Zhong, J.; Chen, C.Y.; Gao, B.D. Genome sequence of a novel mycovirus of Rhizoctonia solani, a plant pathogenic fungus. Virus Genes 2015, 51, 167–170. [Google Scholar] [CrossRef] [PubMed]
- Zhang, M.; Zheng, L.; Liu, C.; Shu, C.; Zhou, E. Characterization of a novel dsRNA mycovirus isolated from strain A105 of Rhizoctonia solani AG-1 IA. Arch. Virol. 2018, 163, 427–430. [Google Scholar] [CrossRef] [PubMed]
- Liu, C.; Zeng, M.; Zhang, M.; Shu, C.; Zhou, E. Complete nucleotide sequences of a partitivirus virus from Rhizoctonia solani AG-1 IA strain C24. Viruses 2018, 10, 703. [Google Scholar] [CrossRef] [PubMed]
- Morris, T.J.; Dodds, J.A. Isolation and analysis of double-stranded RNA from virus-infected plant and fungal tissue. Phytopathology 1979, 69, 854–858. [Google Scholar] [CrossRef]
- Kondo, H.; Hisano, S.; Chiba, S.; Maruyama, K.; Andika, I.B.; Toyoda, K.; Fujimori, F.; Suzuki, N. Sequence and phylogenetic analyses of novel totivirus-like double-stranded RNAs from field-collected powdery mildew fungi. Virus Res. 2016, 219, 353–364. [Google Scholar] [CrossRef]
- Zheng, L.; Lu, X.; Liang, X.; Jiang, S.; Zhao, J.; Zhan, G.; Liu, P.; Wu, J.; Kang, Z. Molecular characterization of novel Totivirus-like double-stranded RNAs from Puccinia striiformis f. sp.tritici, the causal agent of wheat stripe rust. Front. Microbiol. 2017, 8, 1960. [Google Scholar] [CrossRef]
- Wu, M.D.; Zhang, L.; Li, G.Q.; Jiang, D.H.; Hou, M.S.; Huang, H.C. Hypovirulence and double-stranded RNA in Botrytis cinerea. Phytopathology 2007, 97, 1590–1599. [Google Scholar] [CrossRef]
- Smith, C.A.; Want, E.J.; O’Maille, G.; Abagyan, R.; Siuzdak, G. XCMS: processing mass spectrometry data for metabolite profiling using nonlinear peak alignment, matching, and identification. Anal. Chem. 2006, 78, 779–787. [Google Scholar] [CrossRef]
- Kopka, J.; Schauer, N.; Krueger, S.; Birkemeyer, C.; Usadel, B.; Bergmüller, E.; Dörmann, P.; Weckwerth, W.; Gibon, Y.; Stitt, M.; et al. GMD@CSB.DB: The Golm metabolome database. Bioinformatics 2005, 21, 1635–1638. [Google Scholar] [CrossRef]
- Chong, J.; Soufan, O.; Li, C.; Caraus, I.; Li, S.; Bourque, G.; Wishart, D.S.; Xia, J. MetaboAnalyst 4.0: towards more transparent and integrative metabolomics analysis. Nucleic Acids Res. 2018, 46, W486–W494. [Google Scholar] [CrossRef] [PubMed]
- Smart, C.D.; Yuan, W.; Foglia, R.; Nuss, D.L.; Fulbright, D.W.; Hillman, B.I. Cryphonectria hypovirus 3, a virus species in the family hypoviridae with a single open reading frame. Virology 1999, 265, 66–73. [Google Scholar] [CrossRef] [PubMed]
- Linder-Basso, D.; Dynek, J.N.; Hillman, B.I. Genome analysis of Cryphonectria hypovirus 4, the most common hypovirus species in North America. Virology 2005, 337, 192–203. [Google Scholar] [CrossRef] [PubMed]
- Markinegoriaynoff, N.; Gillet, L.; Van Etten, J.L.; Korres, H.; Verma, N.; Vanderplasschen, A. Glycosyltransferases encoded by viruses. J. Gen. Virol. 2004, 85, 2741–2754. [Google Scholar] [CrossRef] [PubMed]
- Hacker, C.V.; Brasier, C.M.; Buck, K.W. A double-stranded RNA from a Phytophthora species is related to the plant endornaviruses and contains a putative UDP glycosyltransferase gene. J. Gen. Virol. 2005, 86, 1561–1570. [Google Scholar] [CrossRef]
- Koonin, E.V.; Dolja, V.V. Evolution and taxonomy of positive-strand RNA viruses: Implications of comparative analysis of amino acid sequences. Crit. Rev. Biochem. Mol. 1993, 28, 375–430. [Google Scholar] [CrossRef]
- Das, S.; Falloon, R.E.; Stewart, A.; Pitman, A.R. Molecular characterisation of an endornavirus from Rhizoctonia solani AG-3PT infecting potato. Fungal Biol. 2014, 118, 924–934. [Google Scholar] [CrossRef]
- Sneh, B.; Jabajihare, S.; Neate, S.; Dijst, G. Rhizoctonia species: Taxonomy, molecular biology, ecology, pathology and disease control; Springer: Dordrecht, The Netherlands, 1996. [Google Scholar]
- Fernie, A.R.; Trethewey, R.N.; Krotzky, A.J.; Willmitzer, L. Metabolite profiling: From diagnostics to systems biology. Nat. Rev. Mol. Cell Biol. 2004, 5, 763–769. [Google Scholar] [CrossRef]
- Nicholson, J.K.; Lindon, J.C. Systems biology: Metabonomics. Nature 2008, 455, 1054–1056. [Google Scholar] [CrossRef]
- Ling, J.; Reynolds, N.; Ibba, M. Aminoacyl-tRNA synthesis and translational quality control. Annu. Rev. Microbiol. 2016, 63, 61–78. [Google Scholar] [CrossRef]
- Banerjee, R.; Chen, S.; Dare, K.; Gilreath, M.; Praetoriusibba, M.; Raina, M.; Reynolds, N.M.; Rogers, T.; Roy, H.; Yadavalli, S.S. tRNAs: Cellular barcodes for amino acids. FEBS Lett. 2010, 584, 387–395. [Google Scholar] [CrossRef] [PubMed]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Item | Differential Metabolites | Retention Time (min) | VIP 1 | p Value 2 | LOG2(GD-118P-V1/GD-118P) 3 |
|---|---|---|---|---|---|
| 140 | N-Acetyl glucosamine | 23.46 | 2.18 | 0.002 | 0.43 |
| 19 | Lactic acid | 12.10 | 1.63 | 0.000 | −1.04 |
| 96 | Arabinose | 20.74 | 1.43 | 0.000 | −0.83 |
| 182 | Maltose | 29.57 | 1.40 | 0.000 | −1.63 |
| 47 | Succinic acid | 15.99 | 1.34 | 0.000 | −0.57 |
| 54 | Thymine | 16.76 | 1.30 | 0.000 | −0.93 |
| 75 | Pyroglutamic acid | 18.75 | 1.30 | 0.000 | −0.70 |
| 37 | Serine | 15.21 | 1.29 | 0.005 | −1.28 |
| 102 | Xylitol | 21.22 | 1.27 | 0.001 | −1.81 |
| 81 | Bromosuccinate | 19.05 | 1.26 | 0.002 | −0.54 |
| 3 | Ethylene glycol | 8.87 | 1.25 | 0.003 | −0.74 |
| 138 | Tyrosine | 23.35 | 1.22 | 0.001 | −0.37 |
| 154 | Sedoheptulose | 25.48 | 1.21 | 0.003 | −1.90 |
| 16 | 3-hydroxy-pyridine | 11.57 | 1.21 | 0.001 | −0.83 |
| 48 | Glyceric acid | 16.31 | 1.21 | 0.005 | −0.55 |
| 93 | Dodecanoic acid | 20.18 | 1.17 | 0.005 | −1.48 |
| 170 | D-Glucopyranosiduronic acid | 27.66 | 1.15 | 0.006 | −0.48 |
| 80 | 2,4,6-Tri-t-butylbenzenethiol | 19.00 | 1.15 | 0.004 | −0.94 |
| 151 | Myo-Inositol | 24.97 | 1.15 | 0.038 | −0.32 |
| 20 | Hexanoic acid | 12.25 | 1.14 | 0.011 | −0.52 |
| 40 | Phosphoric acid | 15.52 | 1.11 | 0.007 | −0.23 |
| 134 | Glucose | 23.20 | 1.11 | 0.004 | −0.21 |
| 28 | Oxalic acid | 13.44 | 1.09 | 0.003 | 1.72 |
| 110 | Arabinofuranose | 21.67 | 1.08 | 0.035 | −0.44 |
| 91 | Glutamine | 19.93 | 1.08 | 0.007 | 4.05 |
| 176 | Inosine | 28.92 | 1.06 | 0.007 | 1.57 |
| 27 | 1,2-Dihydroxy-cyclohexene | 13.14 | 1.04 | 0.013 | 5.44 |
| 148 | Palmitic acid | 24.23 | 1.01 | 0.021 | −0.55 |
| 116 | Citric acid | 22.22 | 1.01 | 0.006 | 1.58 |
| 112 | Azelaic acid | 21.80 | 1.00 | 0.009 | −0.67 |
| 113 | Hypoxanthine | 21.99 | 1.00 | 0.014 | −0.77 |
| 92 | Phenylalanine | 20.02 | 1.00 | 0.017 | 5.69 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zheng, L.; Shu, C.; Zhang, M.; Yang, M.; Zhou, E. Molecular Characterization of a Novel Endornavirus Conferring Hypovirulence in Rice Sheath Blight Fungus Rhizoctonia solani AG-1 IA Strain GD-2. Viruses 2019, 11, 178. https://doi.org/10.3390/v11020178
Zheng L, Shu C, Zhang M, Yang M, Zhou E. Molecular Characterization of a Novel Endornavirus Conferring Hypovirulence in Rice Sheath Blight Fungus Rhizoctonia solani AG-1 IA Strain GD-2. Viruses. 2019; 11(2):178. https://doi.org/10.3390/v11020178
Chicago/Turabian StyleZheng, Li, Canwei Shu, Meiling Zhang, Mei Yang, and Erxun Zhou. 2019. "Molecular Characterization of a Novel Endornavirus Conferring Hypovirulence in Rice Sheath Blight Fungus Rhizoctonia solani AG-1 IA Strain GD-2" Viruses 11, no. 2: 178. https://doi.org/10.3390/v11020178
APA StyleZheng, L., Shu, C., Zhang, M., Yang, M., & Zhou, E. (2019). Molecular Characterization of a Novel Endornavirus Conferring Hypovirulence in Rice Sheath Blight Fungus Rhizoctonia solani AG-1 IA Strain GD-2. Viruses, 11(2), 178. https://doi.org/10.3390/v11020178