Molecular and Antigenic Characterization of Piscine orthoreovirus (PRV) from Rainbow Trout (Oncorhynchus mykiss)

 , ,
, ,  ,
,  and
and
Abstract
1. Introduction
2. Materials and Methods
2.1. Challenge Experiments
2.2. Virus Purification
2.3. Transmission Electron Microscopy (TEM)
2.4. RNA Isolation and RT-qPCR
2.5. Illumina Sequencing and Genome Assembly
2.6. Sequence and Phylogenetic Analyses
2.7. SDS-PAGE and Western Blot
3. Results
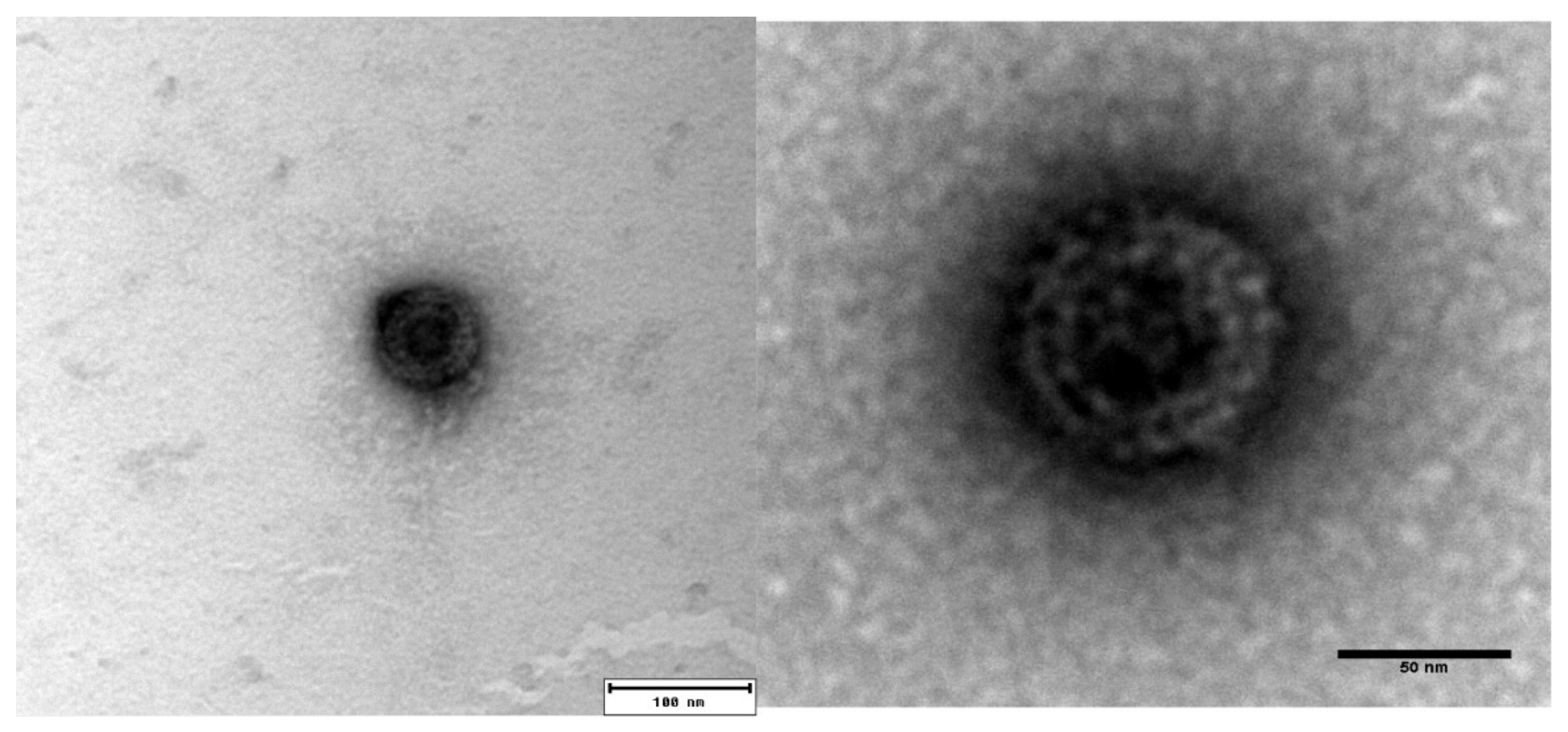
3.1. Morphology of Purified PRV-3 Viral Particles
3.2. The PRV-3 Genome
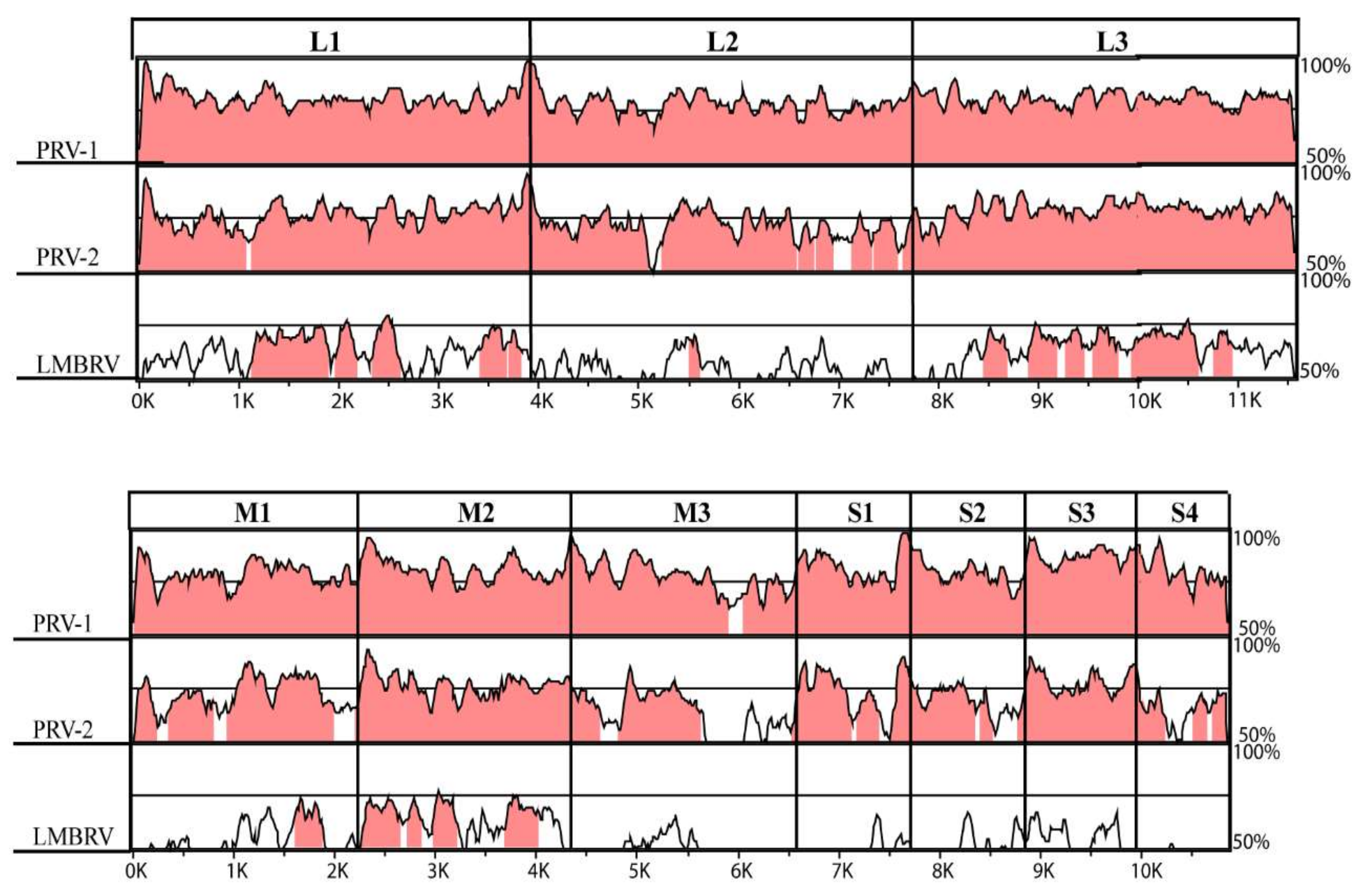
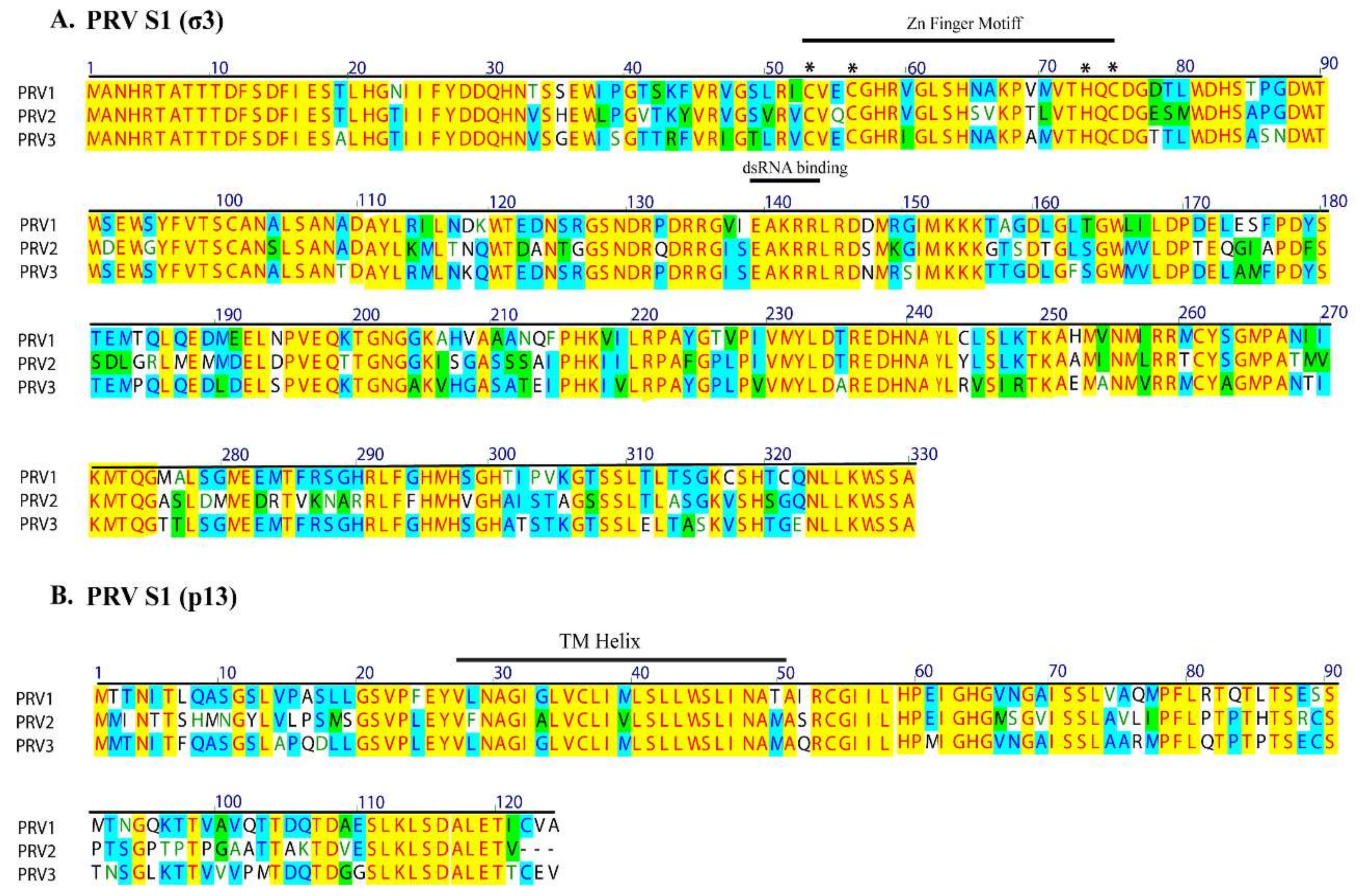
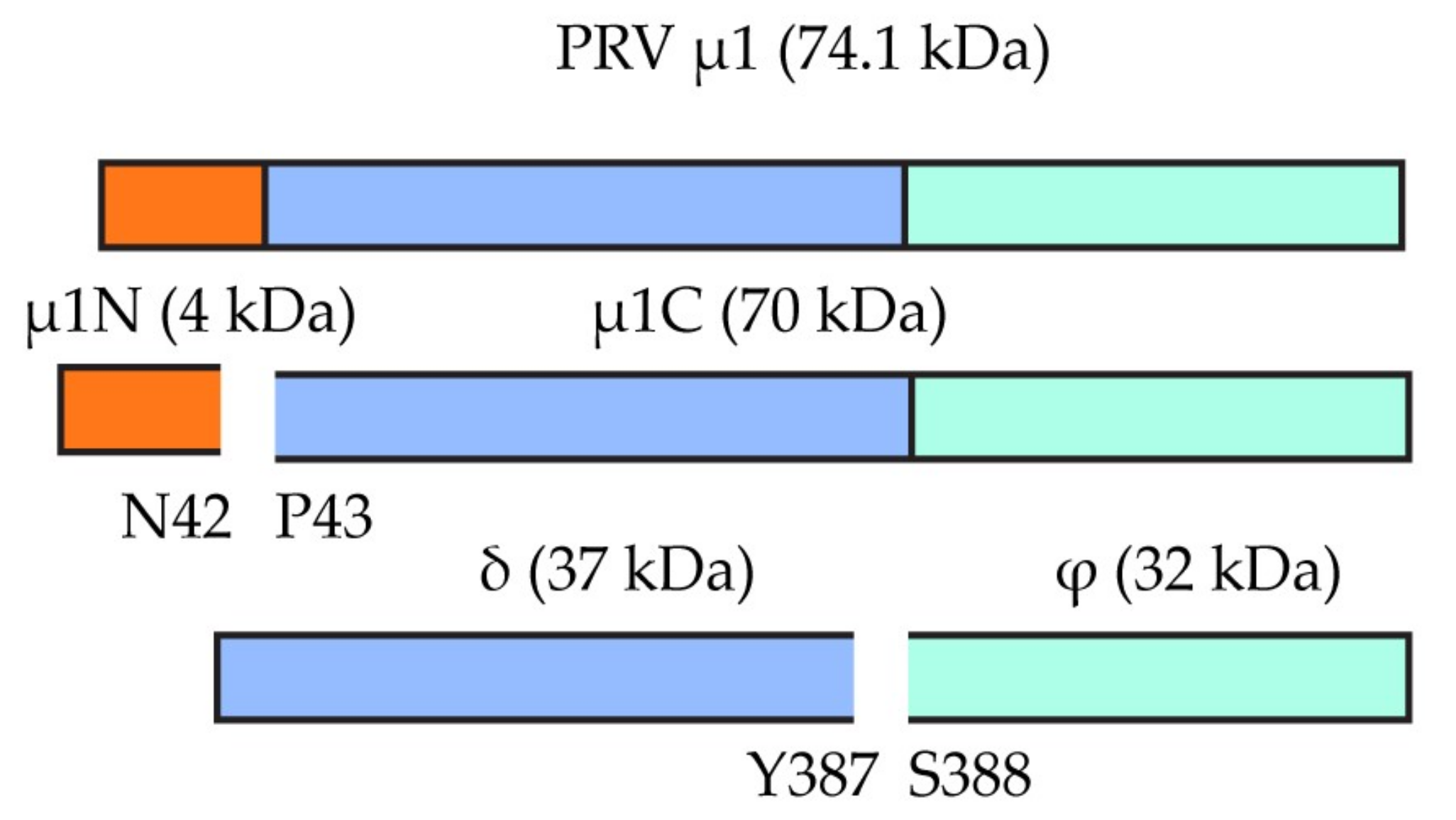
3.3. High Conservation of Putative Functional Protein Domains between the Three PRV Subtypes
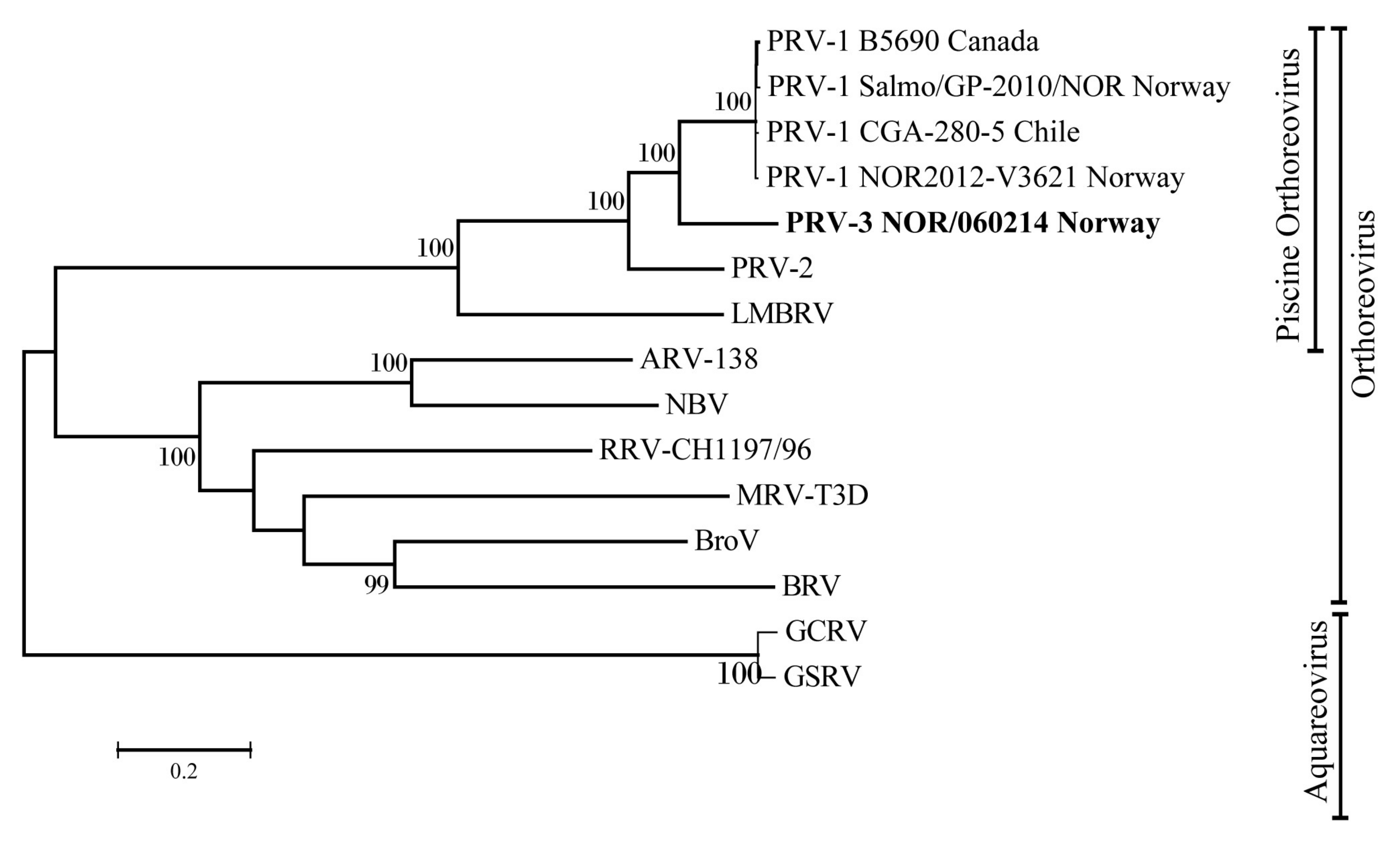
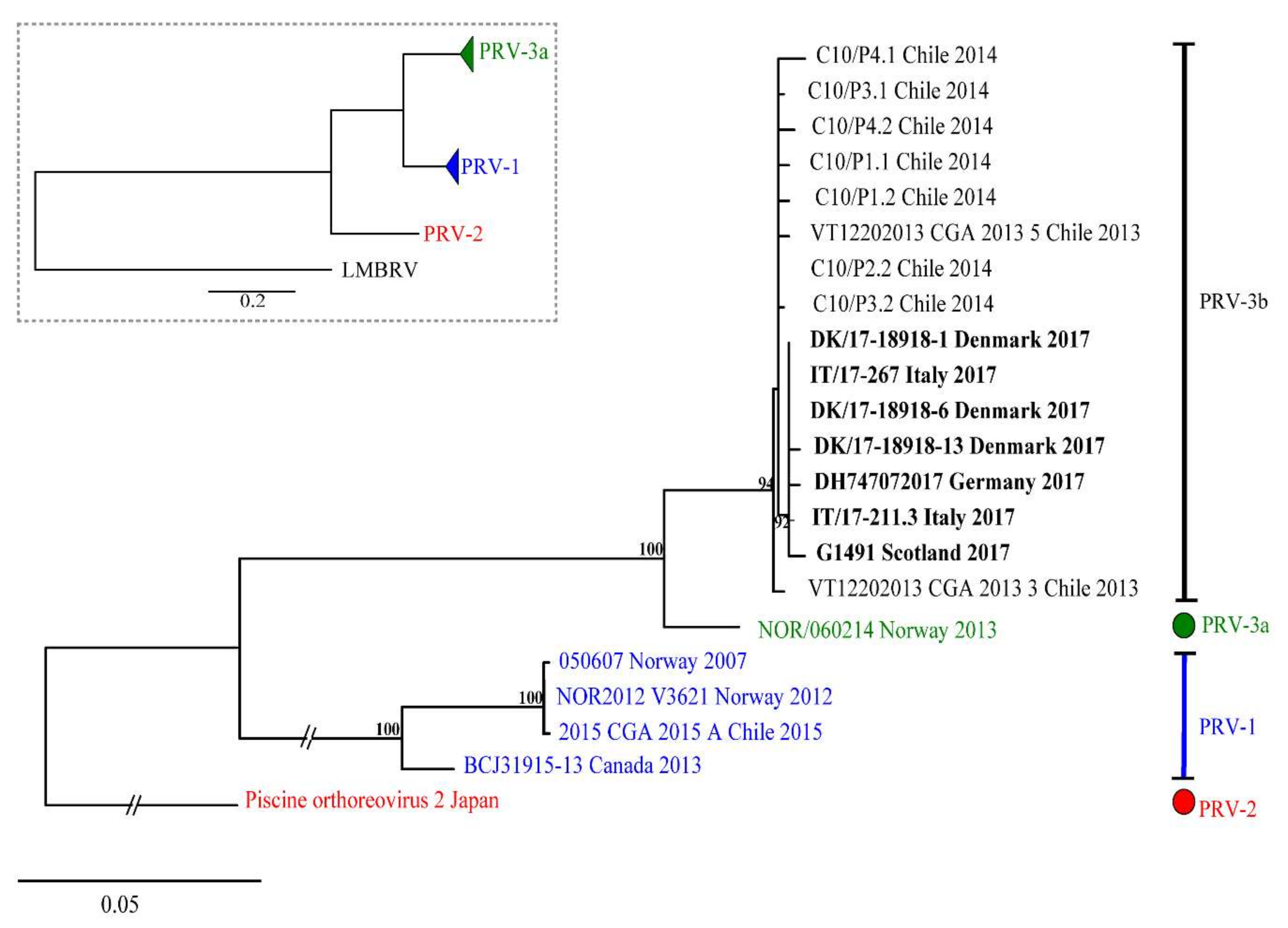
3.4. Phylogenetic Analyses
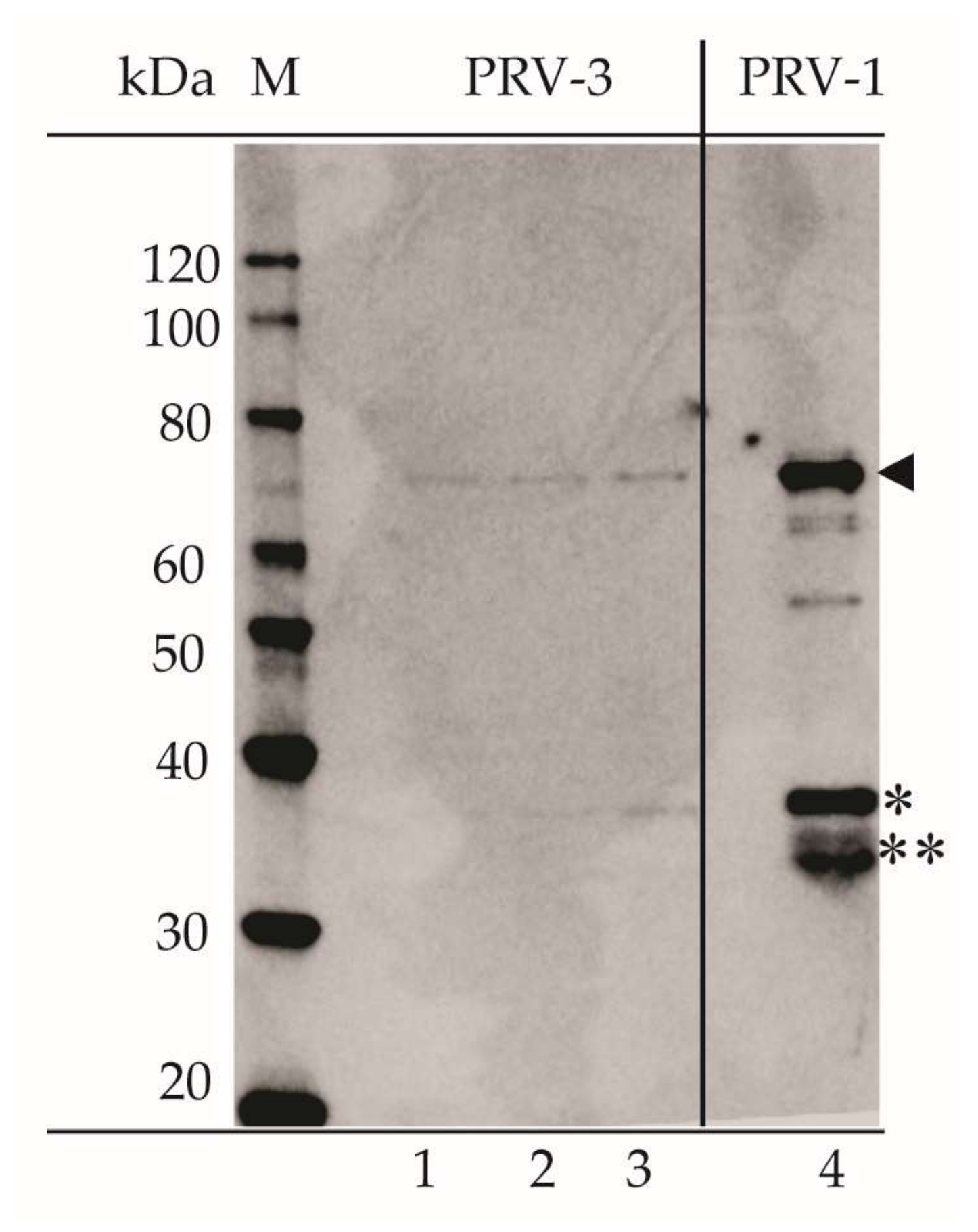
3.5. Serological Cross-Reaction between PRV-1 and PRV-3
4. Discussion
5. Conclusions
Supplementary Materials
Acknowledgments
Author Contributions
Conflicts of Interest
References
- FAO. Fao Yearbook of Fishery and Aquaculture Statistics—2014; Food and Agriculture Organization of the United Nations: Rome, Italy, 2016. [Google Scholar]
- Hjeltnes, B.; Bornø, G.; Jansen, M.D.; Haukaas, A.; Walde, C. The Fish Health Report 2015; Norwegian Veterinary Institute: Oslo, Norway, 2016. [Google Scholar]
- Olsen, A.B.; Hjortaas, M.; Tengs, T.; Hellberg, H.; Johansen, R. First description of a new disease in rainbow trout (Oncorhynchus mykiss (Walbaum)) similar to heart and skeletal muscle inflammation (HSMI) and detection of a gene sequence related to piscine orthoreovirus (PRV). PLoS ONE 2015, 10, e0131638. [Google Scholar] [CrossRef] [PubMed]
- Palacios, G.; Lovoll, M.; Tengs, T.; Hornig, M.; Hutchison, S.; Hui, J.; Kongtorp, R.T.; Savji, N.; Bussetti, A.V.; Solovyov, A.; et al. Heart and skeletal muscle inflammation of farmed salmon is associated with infection with a novel reovirus. PLoS ONE 2010, 5, e11487. [Google Scholar] [CrossRef] [PubMed]
- Wessel, O.; Braaen, S.; Alarcon, M.; Haatveit, H.; Roos, N.; Markussen, T.; Tengs, T.; Dahle, M.K.; Rimstad, E. Infection with purified piscine orthoreovirus demonstrates a causal relationship with heart and skeletal muscle inflammation in atlantic salmon. PLoS ONE 2017, 12, e0183781. [Google Scholar] [CrossRef] [PubMed]
- Hauge, H.; Vendramin, N.; Taksdal, T.; Olsen, A.B.; Wessel, O.; Mikkelsen, S.S.; Alencar, A.L.F.; Olesen, N.J.; Dahle, M.K. Infection experiments with novel piscine orthoreovirus from rainbow trout (Oncorhynchus mykiss) in salmonids. PLoS ONE 2017, 12, e0180293. [Google Scholar] [CrossRef] [PubMed]
- Godoy, M.G.; Kibenge, M.J.; Wang, Y.; Suarez, R.; Leiva, C.; Vallejos, F.; Kibenge, F.S. First description of clinical presentation of piscine orthoreovirus (PRV) infections in salmonid aquaculture in chile and identification of a second genotype (Genotype II) of PRV. Virol. J. 2016, 13, 98. [Google Scholar] [CrossRef] [PubMed]
- Kongtorp, R.T.; Kjerstad, A.; Taksdal, T.; Guttvik, A.; Falk, K. Heart and skeletal muscle inflammation in atlantic salmon, Salmo salar L.: A new infectious disease. J. Fish Dis. 2004, 27, 351–358. [Google Scholar] [CrossRef] [PubMed]
- Kongtorp, R.T.; Taksdal, T.; Lyngoy, A. Pathology of heart and skeletal muscle inflammation (HSMI) in farmed atlantic salmon Salmo salar. Dis. Aquat. Organ. 2004, 59, 217–224. [Google Scholar] [CrossRef] [PubMed]
- Kongtorp, R.T.; Taksdal, T. Studies with experimental transmission of heart and skeletal muscle inflammation in atlantic salmon, Salmo salar L. J. Fish Dis. 2009, 32, 253–262. [Google Scholar] [CrossRef] [PubMed]
- Markussen, T.; Dahle, M.K.; Tengs, T.; Lovoll, M.; Finstad, O.W.; Wiik-Nielsen, C.R.; Grove, S.; Lauksund, S.; Robertsen, B.; Rimstad, E. Sequence analysis of the genome of piscine orthoreovirus (PRV) associated with heart and skeletal muscle inflammation (HSMI) in atlantic salmon (Salmo salar). PLoS ONE 2013, 8, e70075. [Google Scholar] [CrossRef]
- Garseth, A.H.; Fritsvold, C.; Opheim, M.; Skjerve, E.; Biering, E. Piscine reovirus (PRV) in wild atlantic salmon, Salmo salar L., and sea-trout, Salmo trutta L., in Norway. J. Fish Dis. 2013, 36, 483–493. [Google Scholar] [CrossRef] [PubMed]
- Garver, K.A.; Johnson, S.C.; Polinski, M.P.; Bradshaw, J.C.; Marty, G.D.; Snyman, H.N.; Morrison, D.B.; Richard, J. Piscine orthoreovirus from western north america is transmissible to atlantic salmon and sockeye salmon but fails to cause heart and skeletal muscle inflammation. PLoS ONE 2016, 11, e0146229. [Google Scholar] [CrossRef] [PubMed]
- Lovoll, M.; Alarcon, M.; Bang Jensen, B.; Taksdal, T.; Kristoffersen, A.B.; Tengs, T. Quantification of piscine reovirus (PRV) at different stages of atlantic salmon Salmo salar production. Dis. Aquat. Organ. 2012, 99, 7–12. [Google Scholar] [CrossRef] [PubMed]
- Di Cicco, E.; Ferguson, H.W.; Schulze, A.D.; Kaukinen, K.H.; Li, S.; Vanderstichel, R.; Wessel, O.; Rimstad, E.; Gardner, I.A.; Hammell, K.L.; et al. Heart and skeletal muscle inflammation (HSMI) disease diagnosed on a British Columbia salmon farm through a longitudinal farm study. PLoS ONE 2017, 12, e0171471. [Google Scholar] [CrossRef] [PubMed]
- Ferguson, H.W.; Kongtorp, R.T.; Taksdal, T.; Graham, D.; Falk, K. An outbreak of disease resembling heart and skeletal muscle inflammation in Scottish farmed salmon, Salmo salar L., with observations on myocardial regeneration. J. Fish Dis. 2005, 28, 119–123. [Google Scholar] [CrossRef] [PubMed]
- Marty, G.D.; Morrison, D.B.; Bidulka, J.; Joseph, T.; Siah, A. Piscine reovirus in wild and farmed salmonids in British Columbia, Canada: 1974–2013. J. Fish Dis. 2015, 38, 713–728. [Google Scholar] [CrossRef] [PubMed]
- Takano, T.; Nawata, A.; Sakai, T.; Matsuyama, T.; Ito, T.; Kurita, J.; Terashima, S.; Yasuike, M.; Nakamura, Y.; Fujiwara, A.; et al. Full-genome sequencing and confirmation of the causative agent of erythrocytic inclusion body syndrome in coho salmon identifies a new type of piscine orthoreovirus. PLoS ONE 2016, 11, e0165424. [Google Scholar] [CrossRef] [PubMed]
- Sibley, S.D.; Finley, M.A.; Baker, B.B.; Puzach, C.; Armien, A.G.; Giehtbrock, D.; Goldberg, T.L. Novel reovirus associated with epidemic mortality in wild Largemouth Bass (Micropterus salmoides). J. Gen. Virol. 2016, 97, 2482–2487. [Google Scholar] [CrossRef] [PubMed]
- Mendez, I.I.; Hermann, L.L.; Hazelton, P.R.; Coombs, K.M. A comparative analysis of freon substitutes in the purification of reovirus and calicivirus. J. Virol. Methods 2000, 90, 59–67. [Google Scholar] [CrossRef]
- Bruner, R.; Vinograd, J. The evaluation of standard sedimentation coefficients of sodium RNA and sodium DNA from sedimentation velocity data in concentrated NaCl and CsCl solutions. Biochim. Biophys. Acta 1965, 108, 18–29. [Google Scholar] [CrossRef]
- Bankevich, A.; Nurk, S.; Antipov, D.; Gurevich, A.A.; Dvorkin, M.; Kulikov, A.S.; Lesin, V.M.; Nikolenko, S.I.; Pham, S.; Prjibelski, A.D.; et al. Spades: A new genome assembly algorithm and its applications to single-cell sequencing. J. Comput. Biol. 2012, 19, 455–477. [Google Scholar] [CrossRef] [PubMed]
- Kumar, S.; Stecher, G.; Tamura, K. Mega7: Molecular evolutionary genetics analysis version 7.0 for bigger datasets. Mol. Biol. Evol. 2016, 33, 1870–1874. [Google Scholar] [CrossRef] [PubMed]
- Jones, D.T. Protein secondary structure prediction based on position-specific scoring matrices. J. Mol. Biol. 1999, 292, 195–202. [Google Scholar] [CrossRef] [PubMed]
- Frazer, K.A.; Pachter, L.; Poliakov, A.; Rubin, E.M.; Dubchak, I. Vista: Computational tools for comparative genomics. Nucleic Acids Res. 2004, 32, W273–W279. [Google Scholar] [CrossRef] [PubMed]
- Tamura, K. Estimation of the number of nucleotide substitutions when there are strong transition-transversion and G+C-content biases. Mol. Biol. Evol. 1992, 9, 678–687. [Google Scholar] [PubMed]
- Hungnes, O.; Jonassen, T.O.; Jonassen, C.M.; Grinde, B. Molecular epidemiology of viral infections—How sequence information helps us understand the evolution and dissemination of viruses. Apmis 2000, 108, 81–97. [Google Scholar] [CrossRef] [PubMed]
- Tamura, K.; Stecher, G.; Peterson, D.; Filipski, A.; Kumar, S. Mega6: Molecular evolutionary genetics analysis version 6.0. Mol. Biol. Evol. 2013, 30, 2725–2729. [Google Scholar] [CrossRef] [PubMed]
- Edgar, R.C. Muscle: A multiple sequence alignment method with reduced time and space complexity. BMC Bioinform. 2004, 5, 113. [Google Scholar] [CrossRef] [PubMed]
- Stamatakis, A. Raxml version 8: A tool for phylogenetic analysis and post-analysis of large phylogenies. Bioinformatics (Oxford, England) 2014, 30, 1312–1313. [Google Scholar] [CrossRef] [PubMed]
- Lund, M.; Rosaeg, M.V.; Krasnov, A.; Timmerhaus, G.; Nyman, I.B.; Aspehaug, V.; Rimstad, E.; Dahle, M.K. Experimental piscine orthoreovirus infection mediates protection against pancreas disease in atlantic salmon (Salmo salar). Vet. Res. 2016, 47, 107. [Google Scholar] [CrossRef] [PubMed]
- Finstad, O.W.; Falk, K.; Lovoll, M.; Evensen, O.; Rimstad, E. Immunohistochemical detection of piscine reovirus (PRV) in hearts of atlantic salmon coincide with the course of heart and skeletal muscle inflammation (HSMI). Vet. Res. 2012, 43, 27. [Google Scholar] [CrossRef] [PubMed]
- Haatveit, H.M.; Wessel, O.; Markussen, T.; Lund, M.; Thiede, B.; Nyman, I.B.; Braaen, S.; Dahle, M.K.; Rimstad, E. Viral protein kinetics of piscine orthoreovirus infection in atlantic salmon blood cells. Viruses 2017, 9, 49. [Google Scholar] [CrossRef] [PubMed]
- Attoui, H.; Fang, Q.; Mohd Jaafar, F.; Cantaloube, J.F.; Biagini, P.; de Micco, P.; de Lamballerie, X. Common evolutionary origin of aquareoviruses and orthoreoviruses revealed by genome characterization of golden shiner reovirus, grass carp reovirus, striped bass reovirus and golden ide reovirus (genus Aquareovirus, family Reoviridae). J. Gen. Virol. 2002, 83, 1941–1951. [Google Scholar] [CrossRef] [PubMed]
- Duncan, R. Extensive sequence divergence and phylogenetic relationships between the fusogenic and nonfusogenic orthoreoviruses: A species proposal. Virology 1999, 260, 316–328. [Google Scholar] [CrossRef] [PubMed]
- Attoui, H.; Mertens, P.P.C.; Becnel, J.; Belaganahalli, S.; Bergoin, M.; Brussaard, C.P.; Chappell, J.D.; Ciarlet, M.; del Vas, M.; Dermody, T.S.; et al. Family—Reoviridae. In Virus Taxonomy: Ninth Report of the International Committee on Taxonomy of Viruses; Elsevier: San Diego, CA, USA, 2012; pp. 541–637. [Google Scholar]
- Xu, W.; Coombs, K.M. Conserved structure/function of the orthoreovirus major core proteins. Virus Res. 2009, 144, 44–57. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Nibert, M.L.; Fields, B.N. A carboxy-terminal fragment of protein mu 1/mu 1C is present in infectious subvirion particles of mammalian reoviruses and is proposed to have a role in penetration. J. Virol. 1992, 66, 6408–6418. [Google Scholar] [PubMed]
- Nibert, M.L.; Odegard, A.L.; Agosto, M.A.; Chandran, K.; Schiff, L.A. Putative autocleavage of reovirus μ1 protein in concert with outer-capsid disassembly and activation for membrane permeabilization. J. Mol. Biol. 2005, 345, 461–474. [Google Scholar] [CrossRef] [PubMed]
- Snyder, A.J.; Danthi, P. Cleavage of the c-terminal fragment of reovirus μ1 is required for optimal infectivity. J. Virol. 2018, 92. [Google Scholar] [CrossRef] [PubMed]
- Liemann, S.; Chandran, K.; Baker, T.S.; Nibert, M.L.; Harrison, S.C. Structure of the reovirus membrane-penetration protein, μ1, in a complex with is protector protein, σ3. Cell 2002, 108, 283–295. [Google Scholar] [CrossRef]
- Sarkar, P.; Danthi, P. The μ1 72–96 loop controls conformational transitions during reovirus cell entry. J. Virol. 2013, 87, 13532–13542. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.; Jin, L.; Fang, Q.; Hui, W.H.; Zhou, Z.H. 3.3 A Cryo-EM structure of a nonenveloped virus reveals a priming mechanism for cell entry. Cell 2010, 141, 472–482. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.; Tang, J.; Walker, S.B.; O’Hara, D.; Nibert, M.L.; Duncan, R.; Baker, T.S. Structure of avian orthoreovirus virion by electron cryomicroscopy and image reconstruction. Virology 2005, 343, 25–35. [Google Scholar] [CrossRef] [PubMed]
- Haatveit, H.M.; Nyman, I.B.; Markussen, T.; Wessel, O.; Dahle, M.K.; Rimstad, E. The non-structural protein muns of piscine orthoreovirus (PRV) forms viral factory-like structures. Vet. Res. 2016, 47, 5. [Google Scholar] [CrossRef] [PubMed]
- Goral, M.I.; Mochow-Grundy, M.; Dermody, T.S. Sequence diversity within the reovirus S3 gene: Reoviruses evolve independently of host species, geographic locale, and date of isolation. Virology 1996, 216, 265–271. [Google Scholar] [CrossRef] [PubMed]
- Tang, Y.; Lin, L.; Sebastian, A.; Lu, H. Detection and characterization of two co-infection variant strains of avian orthoreovirus (ARV) in young layer chickens using next-generation sequencing (NGS). Sci. Rep. 2016, 6, 24519. [Google Scholar] [CrossRef] [PubMed]
- Fan, Y.; Rao, S.; Zeng, L.; Ma, J.; Zhou, Y.; Xu, J.; Zhang, H. Identification and genomic characterization of a novel fish reovirus, hubei grass carp disease reovirus, isolated in 2009 in China. J. Gen. Virol. 2013, 94, 2266–2277. [Google Scholar] [CrossRef] [PubMed]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Segment | Protein Name a | Predicted Function | PRV-3 & PRV-1 | PRV-3 & PRV-2 | PRV-1 &PRV-2 | |||
|---|---|---|---|---|---|---|---|---|
| nt | aa | nt | aa | nt | aa | |||
| L1 | λ3 (Core RdRp) | RNA-dependent RNA polymerase | 80.9 | 95.2 | 76.1 | 88.4 | 77.3 | 89.0 |
| L2 | λ2 (Core turret) | Guanylyltransferase, methyltransferase | 77.8 | 90.0 | 70.9 | 77.1 | 71.1 | 76.9 |
| L3 | λ1 (Core shell) | Helicase, NTPase, RNA triphosphatase | 80.3 | 96.7 | 78.3 | 93.0 | 77.5 | 92.7 |
| M1 | µ2 (Core NTPase) | NTPase, RNA triphos-phatase, RNA binding | 78.4 | 88.7 | 72.0 | 78.3 | 72.2 | 78.1 |
| M2 | µ1 (Outer shell) | Outer capsid protein, membrane penetration | 81.2 | 91.5 | 76.4 | 84.3 | 76.4 | 85.1 |
| M3 | µNS (NS-factory) | Non-structural protein | 76.5 | 82.2 | 62.6 | 59.7 | 62.3 | 59.3 |
| S1 | σ3 (Outer clamp) | σ3: outer capsid protein, zinc metalloprotein | 80.5 | 79.1 | 71.3 | 69.7 | 71.6 | 69.7 |
| p13 | p13: cytotoxic, integral membrane protein | 85.6 | 78.2 | 78.1 | 63.7 | 77.3 | 62.9 | |
| S2 | σ2 (Core clamp) | Inner capsid, RNA binding | 80.4 | 88.8 | 70.1 | 73.8 | 70.2 | 77.1 |
| S3 | σNS (NS-RNA) | Non-structural protein | 87.9 | 94.6 | 77.8 | 85.3 | 76.6 | 84.7 |
| S4 | σ1 (Outer fibre) | Cell attachment protein | 80.0 | 81.6 | 64.5 | 64.4 | 65.5 | 66.7 |
| Concatenated coding sequences | 80.1 | 90.5 | 72.9 | 80.0 | 73.4 | 80.3 | ||
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Dhamotharan, K.; Vendramin, N.; Markussen, T.; Wessel, Ø.; Cuenca, A.; Nyman, I.B.; Olsen, A.B.; Tengs, T.; Krudtaa Dahle, M.; Rimstad, E. Molecular and Antigenic Characterization of Piscine orthoreovirus (PRV) from Rainbow Trout (Oncorhynchus mykiss). Viruses 2018, 10, 170. https://doi.org/10.3390/v10040170
Dhamotharan K, Vendramin N, Markussen T, Wessel Ø, Cuenca A, Nyman IB, Olsen AB, Tengs T, Krudtaa Dahle M, Rimstad E. Molecular and Antigenic Characterization of Piscine orthoreovirus (PRV) from Rainbow Trout (Oncorhynchus mykiss). Viruses. 2018; 10(4):170. https://doi.org/10.3390/v10040170
Chicago/Turabian StyleDhamotharan, Kannimuthu, Niccolò Vendramin, Turhan Markussen, Øystein Wessel, Argelia Cuenca, Ingvild B. Nyman, Anne Berit Olsen, Torstein Tengs, Maria Krudtaa Dahle, and Espen Rimstad. 2018. "Molecular and Antigenic Characterization of Piscine orthoreovirus (PRV) from Rainbow Trout (Oncorhynchus mykiss)" Viruses 10, no. 4: 170. https://doi.org/10.3390/v10040170
APA StyleDhamotharan, K., Vendramin, N., Markussen, T., Wessel, Ø., Cuenca, A., Nyman, I. B., Olsen, A. B., Tengs, T., Krudtaa Dahle, M., & Rimstad, E. (2018). Molecular and Antigenic Characterization of Piscine orthoreovirus (PRV) from Rainbow Trout (Oncorhynchus mykiss). Viruses, 10(4), 170. https://doi.org/10.3390/v10040170