
Salinity Differentially Affects Growth and Ecophysiology of Two Mastic Tree (Pistacia lentiscus L.) Accessions

 ,
,  ,
, 
 and
and 
Abstract
:
1. Introduction
2. Materials and Methods
3. Results
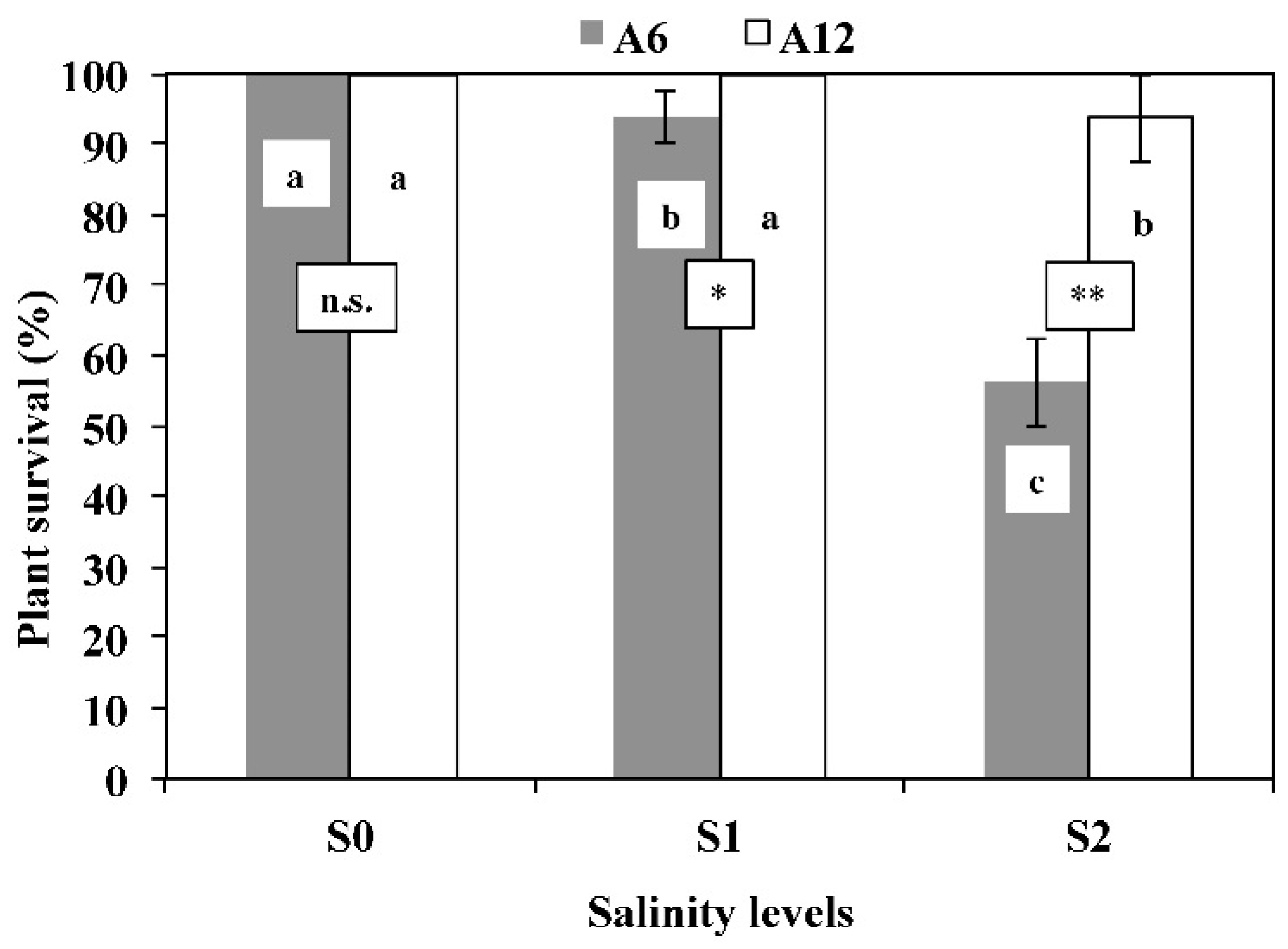
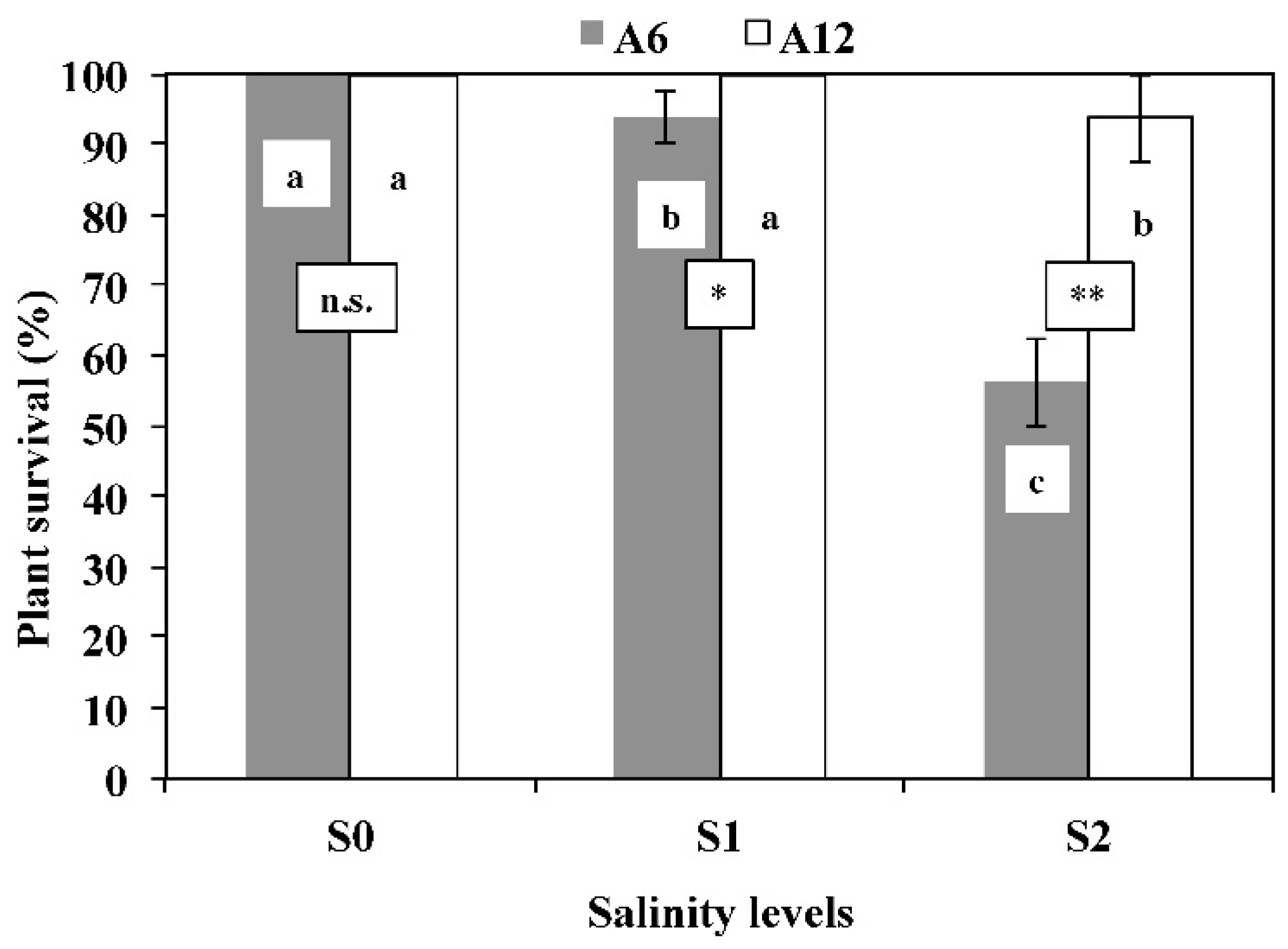
3.1. Plants Survival
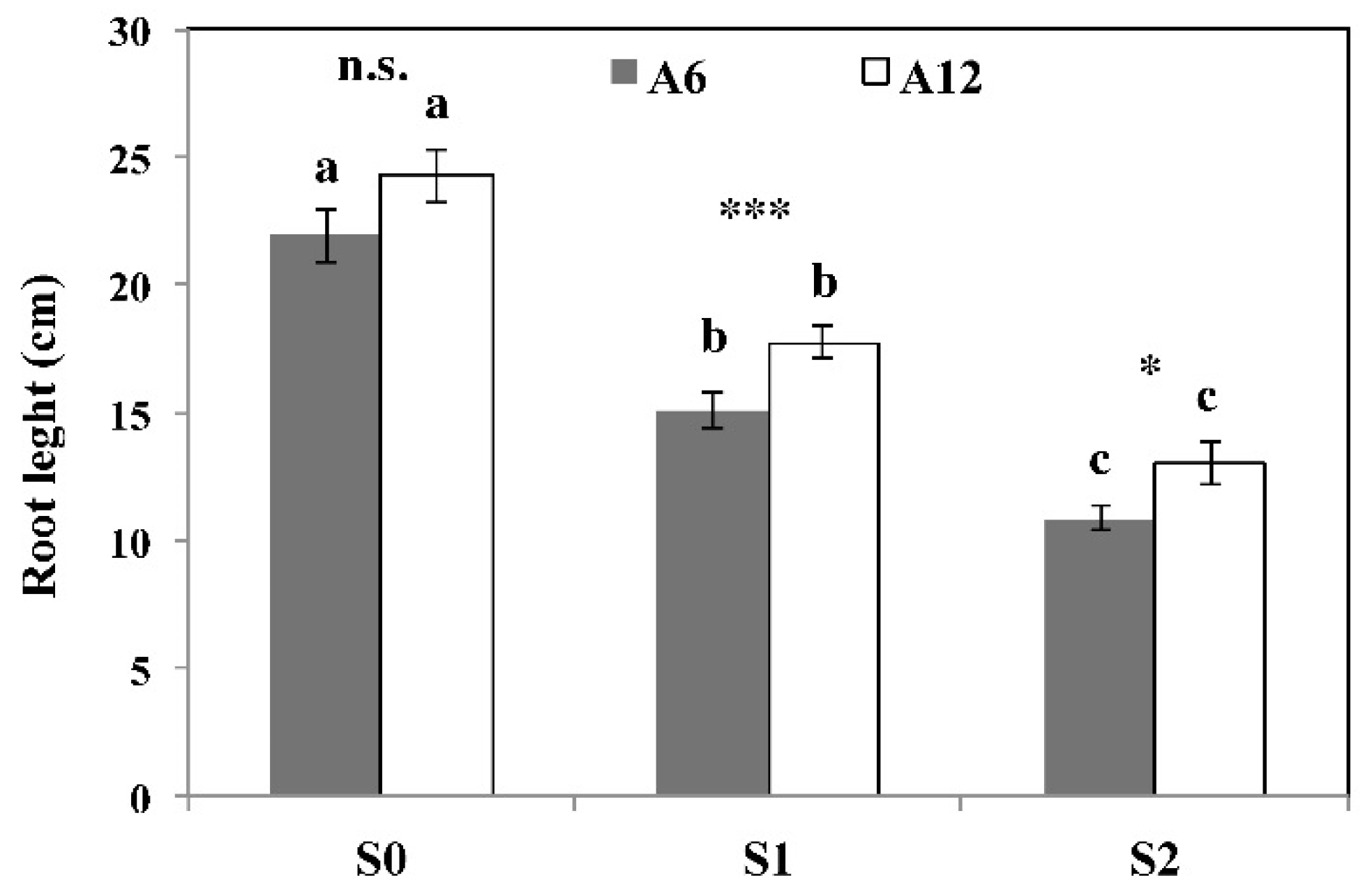
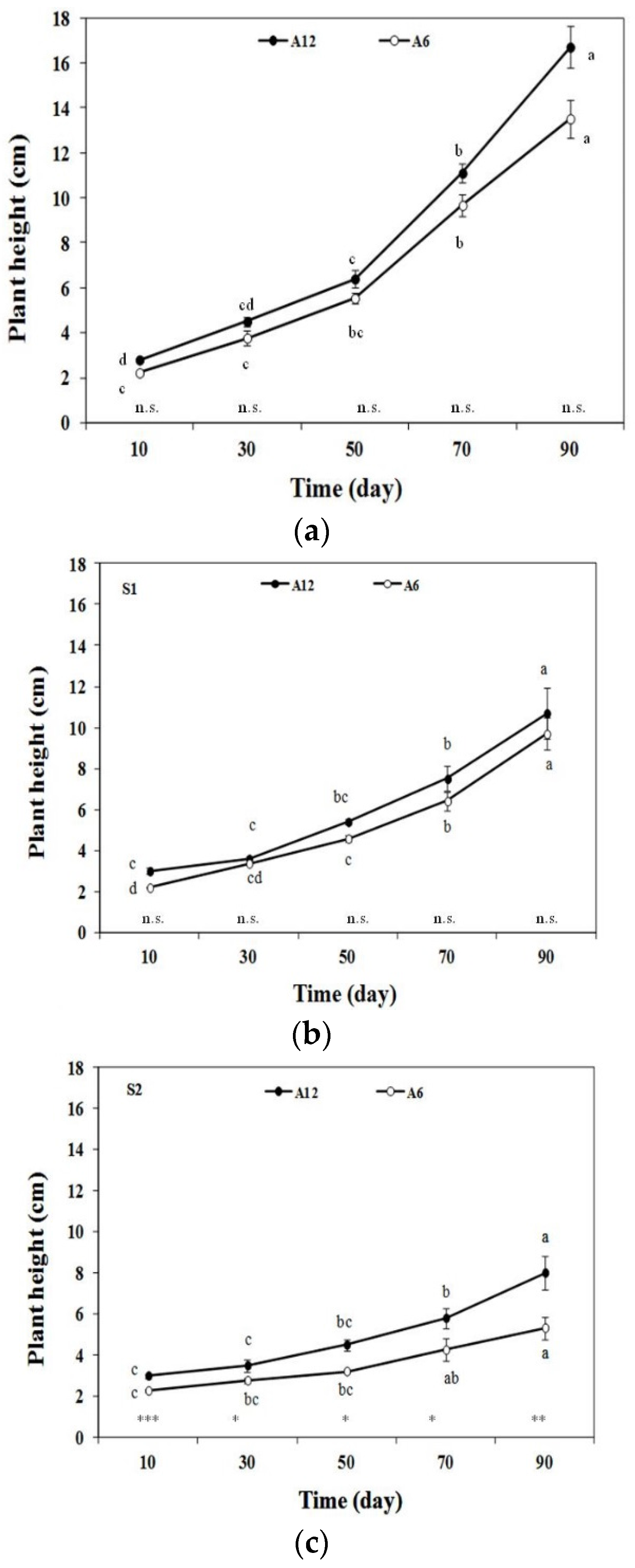
3.2. Plant Height, Root Length, and Total Dry Biomass
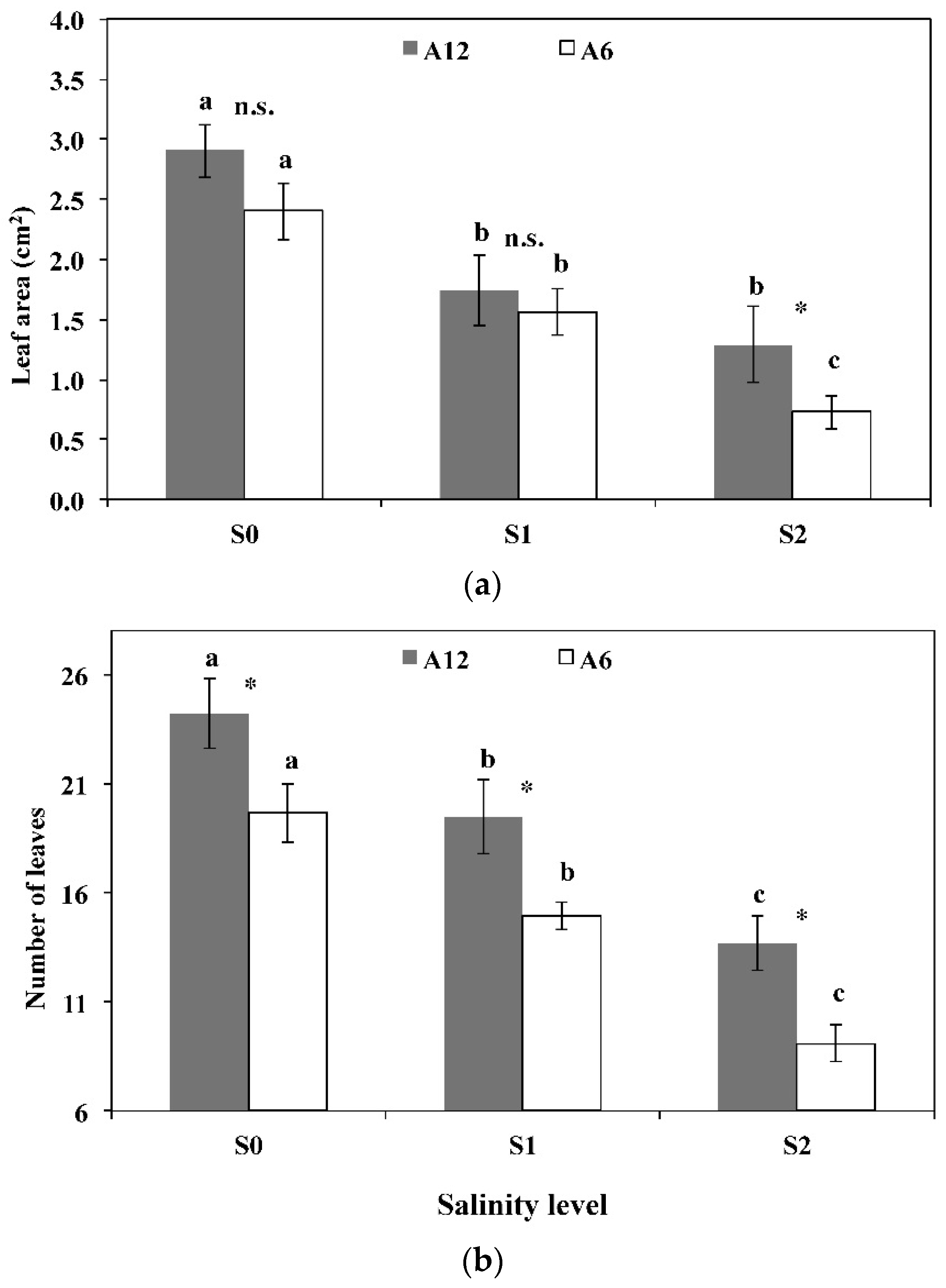
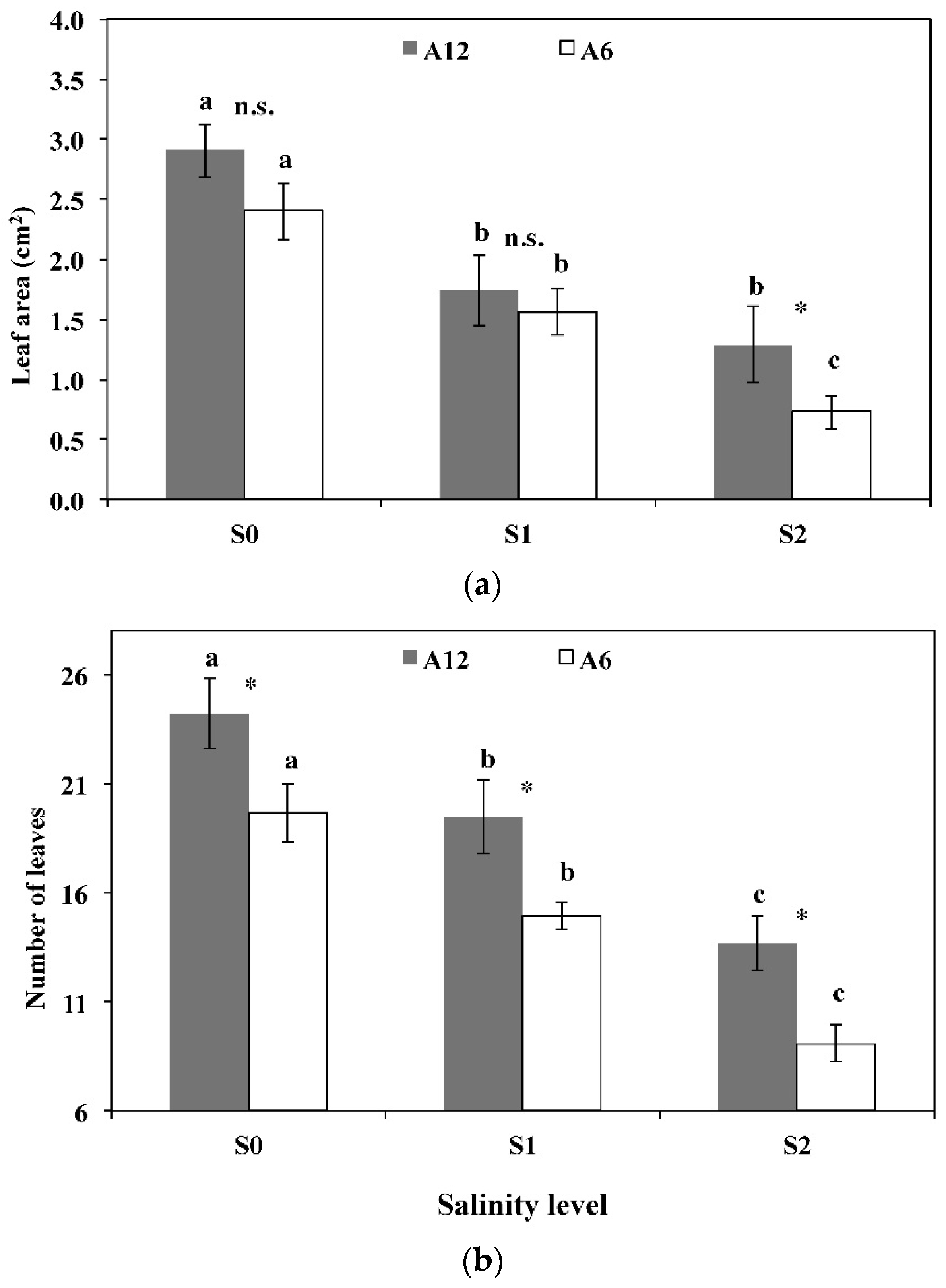
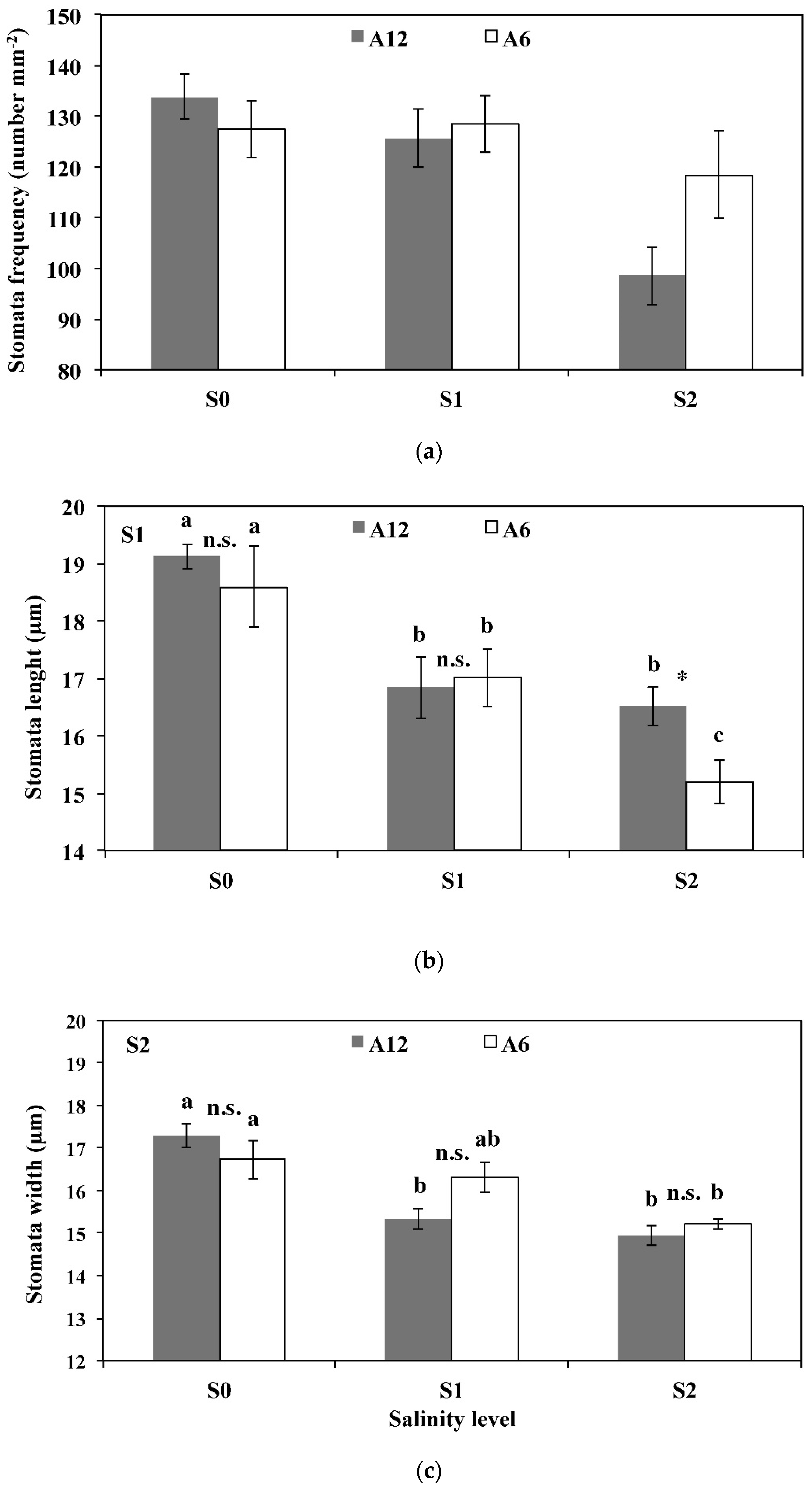
3.3. Leaf Area, Number of Leaves, Stomatal Frequency, and Size
3.4. Ecophysiological Parameters
3.5. Total N and Cations in Biomass
4. Discussion
5. Conclusions
Author Contributions
Conflicts of Interest
References
- Filella, I.; Llusià, J.; Piñol, J.; Peñuelas, J. Leaf gas exchange and fluorescence of Phillyrea latifolia, Pistacia lentiscus and Quercus ilex saplings in severe drought and high temperature conditions. Environ. Exp. Bot. 1998, 39, 213–220. [Google Scholar] [CrossRef]
- Tattini, M.; Remorini, D.; Pinelli, P.; Agati, G.; Saracini, E.; Traversi, M.L.; Massai, R. Morpho-anatomical, physiological and biochemical adjustments in response to root zone salinity stress and high solar radiation in two Mediterranean evergreen shrubs, Myrtus communis and Pistacia lentiscus. New Phytol. 2006, 170, 779–794. [Google Scholar] [CrossRef] [PubMed]
- Vasques, A.R.; Pinto, G.; Dias, M.C.; Correia, C.M.; Moutinho-Pereira, J.M.; Vallejo, V.R.; Santos, C.; Keizer, J.J. Physiological response to drought in seedlings of Pistacia lentiscus (mastic tree). New For. 2015, 1–12. [Google Scholar] [CrossRef]
- Margaris, N.S. Adaptive strategies in plants dominating Mediterranean-type ecosystems. In Ecosystems of the World; di Castri, F., Goodal, D.W., Specht, R.L., Eds.; Elsevier: New York, NY, USA, 1981; Volume 11, pp. 309–315. [Google Scholar]
- Gucci, R.; Aronne, G.; Lombardini, L.; Tattini, M. Salinity tolerance in Phillyrea species. New Phytol. 1997, 135, 227–234. [Google Scholar] [CrossRef]
- Browicz, K. Pistacia lentiscus cv. Chia (Anacardiaceae) on Chios Island. Plant Syst. Evol. 1987, 155, 189–195. [Google Scholar] [CrossRef]
- Zohary, D. The genus Pistacia L. In Taxonomy, Distribution, Conservation and Uses of Pistacia Genetic Resources; Padulosi, S., Caruso, T., Barone, E., Eds.; IPGRI: Rome, Italy, 1996; pp. 1–11. [Google Scholar]
- Murillo, J.M.; Madejón, E.; Madejón, P.; Cabrera, F. The response of wild olive to the addition of a fulvic acid-rich amendment to soils polluted by trace elements (SW Spain). J. Arid Environ. 2005, 63, 284–303. [Google Scholar] [CrossRef]
- Stellacci, A.M.; Cristiano, G.; Rubino, P.; de Lucia, B.; Cazzato, E. Nitrogen uptake, nitrogen partitioning and N-use efficiency of container-grown Holm oak (Quercus ilex L.) under different nitrogen levels and fertilizer sources. J. Food Agric. Environ. 2013, 11, 990–994. [Google Scholar]
- Rinaldi, S.; de Lucia, B.; Salvati, L.; Rea, E. Understanding complexity in the response of ornamental rosemary to different substrates: A multivariate analysis. Sci. Hortic. 2014, 176, 218–224. [Google Scholar] [CrossRef]
- De Lucia, B.; Cristiano, G. Composted amendment affects soil quality and hedges performance in the Mediterranean urban landscape. Compos. Sci. Utili. 2015, 23, 48–57. [Google Scholar] [CrossRef]
- Gebauer, J.; El-Siddig, K.; Salih, A.A.; Ebert, G. Tamarindus indica L. seedlings are moderately salt tolerant when exposed to NaCl-induced salinity. Sci. Hortic. 2004, 103, 1–8. [Google Scholar] [CrossRef]
- Barakat, N.; Laudadio, V.; Cazzato, E.; Tufarelli, V. Antioxidant potential and oxidative stress markers in wheat (Triticum aestivum) treated with phytohormones under salt-stress condition. Int. J. Agric. Biol. 2013, 15, 843–849. [Google Scholar]
- Barakat, N.A.; Cazzato, E.; Nedjimi, B.; Kabiel, H.F.; Laudadio, V.; Tufarelli, V. Ecophysiological and species-specific responses to seasonal variations in halophytic species of the chenopodiaceae in a Mediterranean salt marsh. Afr. J. Ecol. 2014, 52, 163–172. [Google Scholar] [CrossRef]
- USEPA, EPA. Oceans and Coastal Protection Division Report 842-B-92–010. In Plastic Pellets in the Aquatic Environment: Sources and Recommendations; EPA Publisher: Washington, DC, USA, 1992. [Google Scholar]
- Shaviv, I. Autecology of Pistacia lentiscus L. Ph.D. Thesis, Science to the Senate of the Technion-Israel Institute of Technology, Haifa, Israel, June 1978. [Google Scholar]
- Nahum, S.; Inbar, M.; Neeman, G.; Ben-Shlomo, R. Phenotypic plasticity and gene diversity in Pistacia lentiscus L. along environmental gradients in Israel. Tree Genet. Genomes 2008, 4, 777–785. [Google Scholar] [CrossRef]
- Camposeo, S.; Palasciano, M.; Vivaldi, G.A.; Godini, A. Effect of increasing climatic water deficit on some leaf and stomatal parameters of wild and cultivated almonds under Mediterranean conditions. Sci. Hortic. 2011, 127, 234–241. [Google Scholar] [CrossRef]
- Camposeo, S.; Vivaldi, G.A. Short-term effects of de-oiled olive pomace mulching application on a young super high-density olive orchard. Sci. Hortic. 2011, 129, 613–621. [Google Scholar] [CrossRef]
- Scholander, P.F.; Hammel, H.T.; Bradstreet, E.D.; Hemmingsen, E.A. Sap pressure in vascular plants. Science 1965, 148, 339–346. [Google Scholar] [CrossRef] [PubMed]
- Maas, E.V.; Hoffman, G.J. Crop salt tolerance\-current assessment. J. Irrig. Drain. 1977, 103, 115–134. [Google Scholar]
- Greenway, H.; Munns, R. Effect of salinity on phosphate accumulation and injury in soybeans II. Role of substrate Cl− and Na+. Plant Soil 1980, 109, 65–71. [Google Scholar]
- Ehret, D.L.; Plant, A.L. Salt tolerance in crop plants. In Environmental Stress in Crop Plants; Dhaliwal, G.S., Arora, R., Eds.; Commonwealth Publishers: New Delhi, India, 1999; pp. 69–120. [Google Scholar]
- Hasegawa, P.M.; Bressan, R.A.; Zhu, J.K.; Bohnert, H.J. Plant cellular and molecular responses to high salinity. Ann. Rev. Plant Physiol. Plant Mol. Biol. 2000, 51, 463–499. [Google Scholar] [CrossRef] [PubMed]
- Parida, A.K.; Das, A.B. Salt tolerance and salinity effects on plants: A review. Ecotoxicol. Environ. Saf. 2005, 60, 324–349. [Google Scholar] [CrossRef] [PubMed]
- Cassaniti, C.; Romano, D.; Flowers, T.J. The Response of Ornamental Plants to Saline Irrigation Water; Intech: Vienna, Austria, 2012. [Google Scholar]
- Beltrán-Morales, A.; Córdoba-Matson, M.V.; García-Hernández, J.L.; Troyo-Diéguez, E.; Azadi, H.; Ruiz-Espinoza, F.H.; Valdez-Cepeda, B.M.A. Salinity effects on germination and seedlings biomass of Pachycereus pecten-aboriginum: An endangered species. J. Prof. Assoc. Cactus Dev. 2015, 17, 107–122. [Google Scholar]
- Gratani, L.; Varone, L. Adaptive photosynthetic strategies of the Mediterranean maquis species according to their origin. Photosynthetica 2004, 42, 551–558. [Google Scholar] [CrossRef]
- Tattini, M.; Traversi, M.L. Responses to changes in Ca2+ supply in two Mediterranean evergreens, Phillyrea latifolia and Pistacia lentiscus, during salinity stress and subsequent relief. Ann. Bot. 2008, 102, 609–622. [Google Scholar] [CrossRef] [PubMed]
- Barazani, O.; Golan-Goldhirsh, A. Salt-driven interactions between Pistacia lentiscus and Salsola inermis. Environ. Sci. Pollut. Res. 2009, 16, 855–861. [Google Scholar] [CrossRef] [PubMed]
- Seema, M.; Habib-ur-Rehmam, A. Germination and growth of Panicum turgidum provenance under saline condition. Pak. J. Biol. Sci. 2003, 6, 164–166. [Google Scholar]
- Zidan, M.A.; Elewa, M.A. Effects of salinity on germination, seedling and some metabolic changes in four plant species (Umbellifere). Indian J. Plant Physiol. 1995, 38, 57–61. [Google Scholar]
- Ramoliya, P.J.; Pandey, A.N. Effect of increasing salt concentration on emergence, growth and survival of seedlings of Salvadora oleoides (Salvadoraceae). J. Arid Environ. 2002, 51, 121–132. [Google Scholar] [CrossRef]
- Aragüés, R.; Puy, J.; Royo, A.; Espada, J.L. Three-year field response of young trees (Olea europaea L., cv. Arbequina) to soil salinity: Trunk growth and leaf ion accumulation. Plant Soil 2005, 271, 265–273. [Google Scholar] [CrossRef]
- Ebert, G. Growth, ion uptake and gas exchange of two Annona species under salt stress. J. Appl. Bot. 1998, 72, 61–65. [Google Scholar]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Accession | Site | Altitude (m a.s.l.) | Latitude | Longitude | ECe (dS·m−1) | pH | N ‰ |
|---|---|---|---|---|---|---|---|
| A6 | Manfredonia (FG) | 5 | 41°35′ N | 15°49′ E | 0.60 | 7.99 | 2.77 |
| A12 | Specchia (LE) | 230 | 39°55′ N | 18°17′ E | 0.27 | 8.08 | 1.82 |
| Substrates/Salinity Levels | Salt Content (g 100 L−1) | ECe (dS·m−1) | pH |
|---|---|---|---|
| S0 | 0 | 1.7 | 7.95 |
| S1 | 400 | 12.5 | 7.35 |
| S2 | 800 | 21.5 | 7.27 |
| Accessions | Equation | ECe50 (dS·m−1) |
|---|---|---|
| A6 | y = 100 − 4.70(x − 12.38) | 23.0 |
| A12 | y = 100 − 1.56(x − 17.23) | 49.3 |
| Salinity Levels | A6 | A12 | t-test | Mean |
|---|---|---|---|---|
| S0 | 1059 ± 141 A | 1550 ± 124 A | ** | 1305 ± 127 A |
| S1 | 620 ± 128 B | 756 ± 68 B | * | 688 ± 72 B |
| S2 | 257 ± 60 C | 527 ± 73 C | ** | 392 ± 67 C |
| mean | 645 ± 115 | 944 ± 110 | * | 795 ± 105 |
| Salinity Levels | A6 | A12 | t-test |
|---|---|---|---|
| Leaf Water Potential (−MPa) | |||
| S0 | 2.2 ± 0.3 b | 2.3 ± 0.3 b | n.s. |
| S1 | 2.8 ± 0.4 a | 3.0 ± 0.3 a | * |
| S2 | 3.0 ± 0.3 a | 3.1 ± 0.2 a | * |
| Stomatal Conductance (mmol H2O m−2·s−1) | |||
| S0 | 41 ± 1.21 a | 42 ± 2.02 a | n.s. |
| S1 | 12 ± 0.90 b | 34 ± 1.17 b | * |
| S2 | 11 ± 0.79 b | 31 ± 1.01 b | * |
| Net Assimilation Rate (μmol CO2 m−2·s−1) | |||
| S0 | 6.5 ± 2.0 a | 6.7 ± 2.0 a | n.s. |
| S1 | 4.0 ± 1.7 b | 4.0 ± 0.7 b | n.s. |
| S2 | 1.7 ± 1.0 c | 2.3 ± 1.5 c | ** |
| Water Use Efficiency (μmol CO2 mmol·H2O−1) | |||
| S0 | 3.1 ± 0.3 a | 3.0 ± 0.3 a | n.s. |
| S1 | 3.0 ± 0.3 a | 2.7 ± 0.2 a | n.s. |
| S2 | 1.9 ± 0.1 b | 2.4 ± 0.2 b | ** |
| Salinity Levels | Total N | K+ | Na+ | Ca2+ | Mg2+ | K+/Na+ |
|---|---|---|---|---|---|---|
| Leaves | ||||||
| S0 | 16.1 b | 9.7 | 1.0 b | 4.3 b | 1.1 | 9.2 A |
| S1 | 17.8 ab | 9.0 | 1.6 b | 5.5 a | 1.1 | 5.7 B |
| S2 | 18.7 a | 9.0 | 3.2 a | 5.3 a | 1.1 | 2.9 C |
| SEM | 0.61 | 0.32 | 0.11 | 0.20 | 0.09 | 0.29 |
| Stems | ||||||
| S0 | 8.0 b | 11.7 | 1.1 b | 4.5 b | 0.6 | 10.7 a |
| S1 | 9.2 ab | 12.3 | 1.9 b | 5.4 ab | 0.6 | 6.6 ab |
| S2 | 11.1 a | 13.2 | 3.3 a | 7.3 a | 0.6 | 4.0 b |
| SEM | 0.35 | 0.44 | 0.13 | 0.21 | 0.02 | 0.30 |
| Roots | ||||||
| S0 | 8.7 a | 11.8 | 1.7 c | 2.0 b | 0.6 | 7.2 a |
| S1 | 10.2 a | 12.1 | 3.4 b | 2.8 b | 0.6 | 3.6 b |
| S2 | 11.1 a | 12.5 | 5.0 a | 4.8 a | 0.8 | 2.5 b |
| SEM | 0.38 | 0.45 | 0.20 | 0.17 | 0.03 | 0.36 |
| Whole plant | ||||||
| S0 | 10.9 b | 11.1 | 1.3 b | 3.6 b | 0.8 | 9.0 a |
| S1 | 12.4 ab | 11.1 | 2.3 b | 4.6 b | 0.8 | 5.3 b |
| S2 | 13.6 a | 11.5 | 3.8 a | 5.8 a | 0.8 | 3.1 c |
| SEM | 0.41 | 0.39 | 0.17 | 0.19 | 0.04 | 0.31 |
© 2016 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC-BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Cristiano, G.; Camposeo, S.; Fracchiolla, M.; Vivaldi, G.A.; De Lucia, B.; Cazzato, E. Salinity Differentially Affects Growth and Ecophysiology of Two Mastic Tree (Pistacia lentiscus L.) Accessions. Forests 2016, 7, 156. https://doi.org/10.3390/f7080156
Cristiano G, Camposeo S, Fracchiolla M, Vivaldi GA, De Lucia B, Cazzato E. Salinity Differentially Affects Growth and Ecophysiology of Two Mastic Tree (Pistacia lentiscus L.) Accessions. Forests. 2016; 7(8):156. https://doi.org/10.3390/f7080156
Chicago/Turabian StyleCristiano, Giuseppe, Salvatore Camposeo, Mariano Fracchiolla, Gaetano Alessandro Vivaldi, Barbara De Lucia, and Eugenio Cazzato. 2016. "Salinity Differentially Affects Growth and Ecophysiology of Two Mastic Tree (Pistacia lentiscus L.) Accessions" Forests 7, no. 8: 156. https://doi.org/10.3390/f7080156
APA StyleCristiano, G., Camposeo, S., Fracchiolla, M., Vivaldi, G. A., De Lucia, B., & Cazzato, E. (2016). Salinity Differentially Affects Growth and Ecophysiology of Two Mastic Tree (Pistacia lentiscus L.) Accessions. Forests, 7(8), 156. https://doi.org/10.3390/f7080156