
Drought Differently Modifies Tolerance and Metal Uptake in Zn- or Cu-Treated Male and Female Salix × fragilis L.


 ,
,  ,
,  ,
,  ,
,  and
and 
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Materials and Methods
2.1. Plant Material and Experiment Set-Up
2.2. Biomass Measurements and Handling
2.3. Relative Water Content
2.4. Metal Content and Uptake Ratios
2.5. Metabolic Investigations
2.5.1. Total Phenolic Content
2.5.2. Profiling of Phenolic Metabolites and Low-Molecular-Weight Organic Acids (LMWOAs)
2.6. Statistical Analysis
3. Results
3.1. Plant Growth and Toxicity Symptoms
3.1.1. Shoot Biomass
3.1.2. Root Biomass
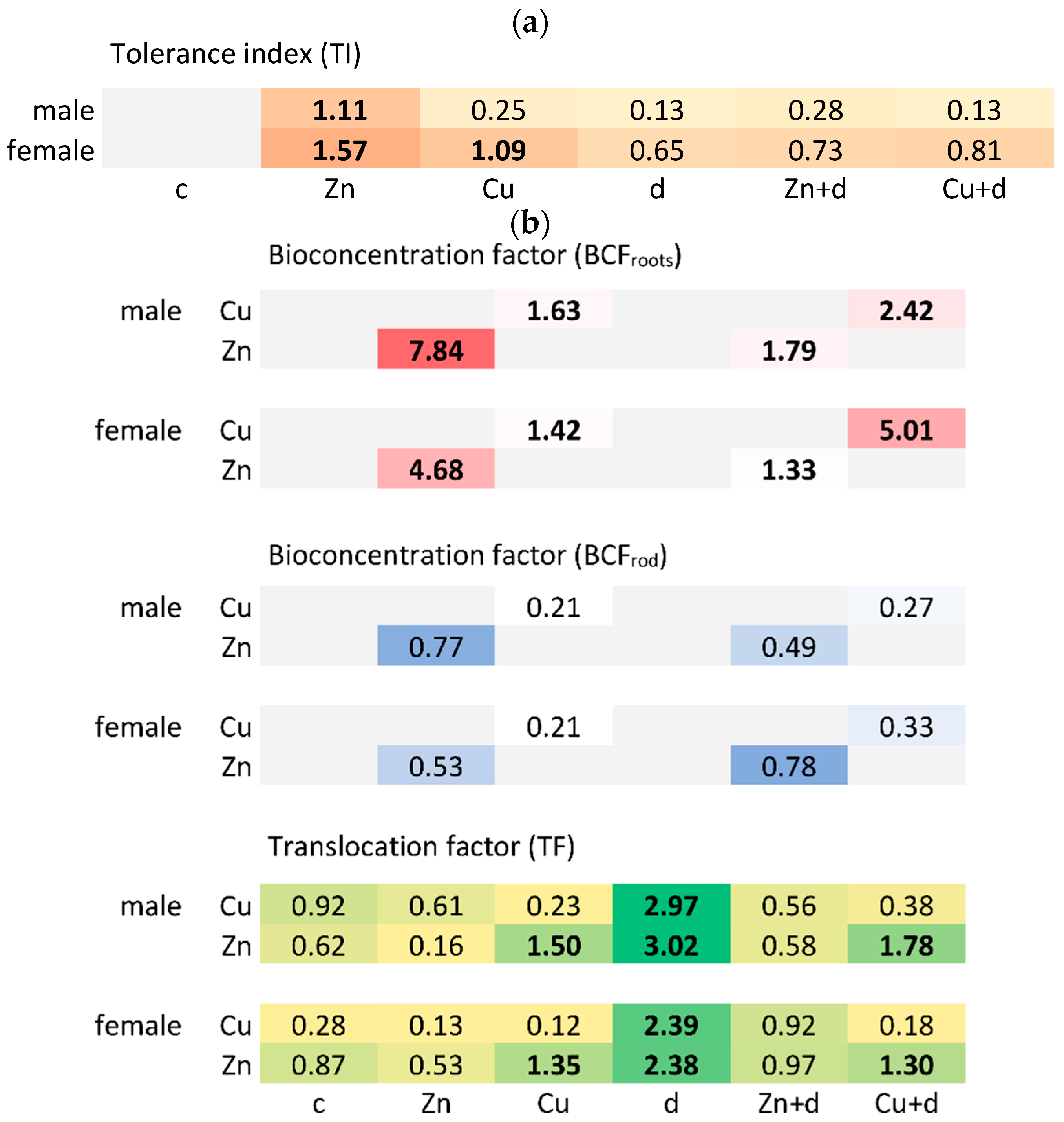
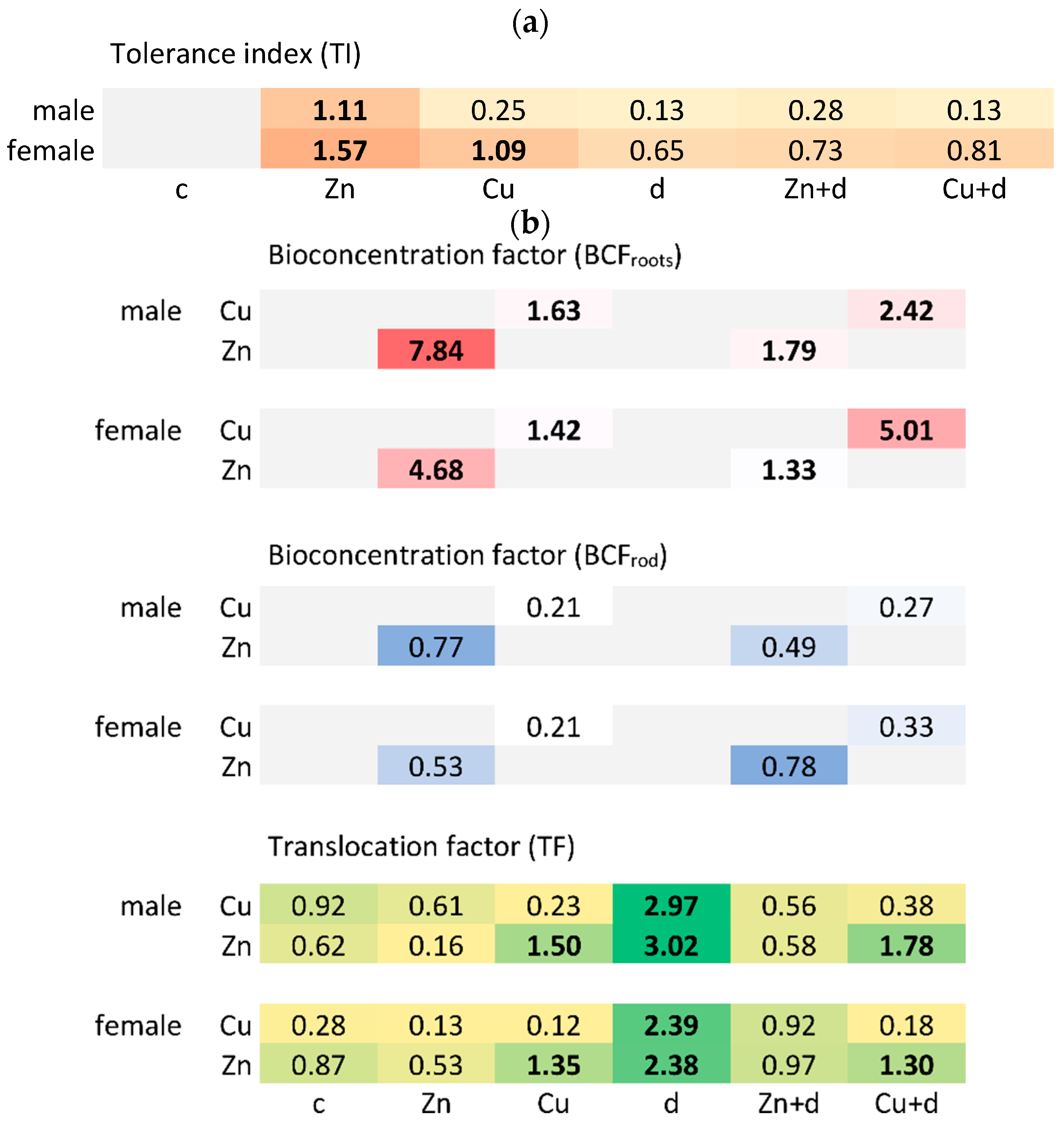
3.1.3. Tolerance Index and Toxicity Symptoms
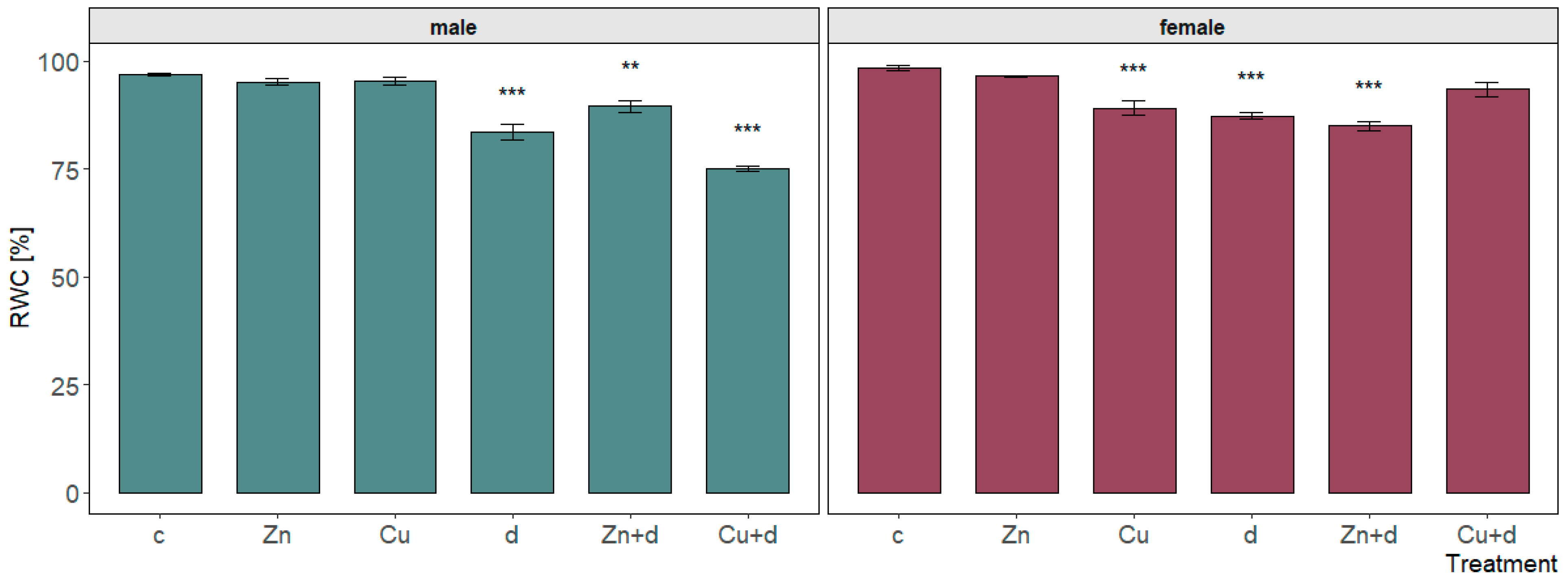
3.2. Leaf RWC
3.3. Cu and Zn Uptake and Translocation
3.3.1. Roots
3.3.2. Rod
3.3.3. Leaves
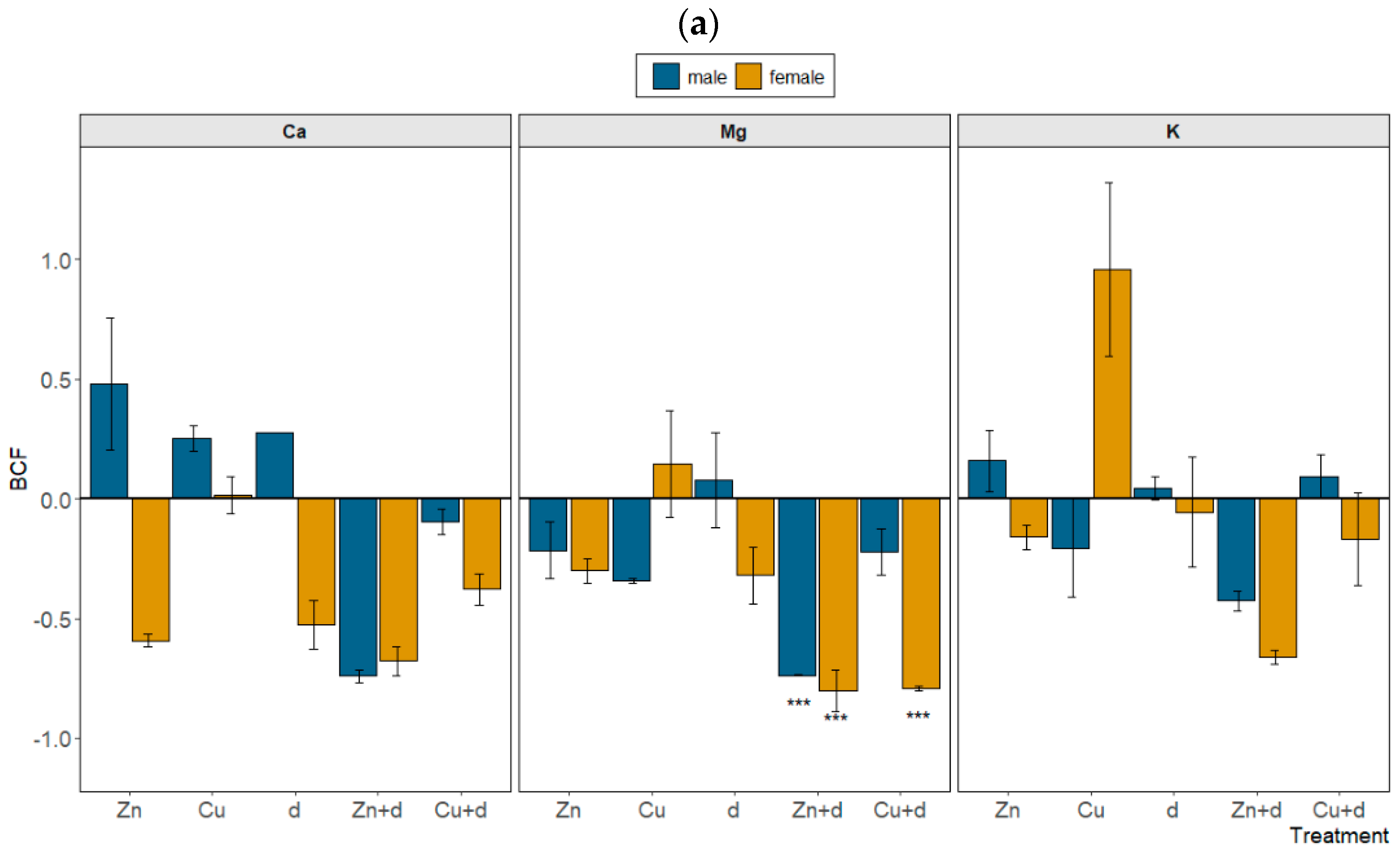
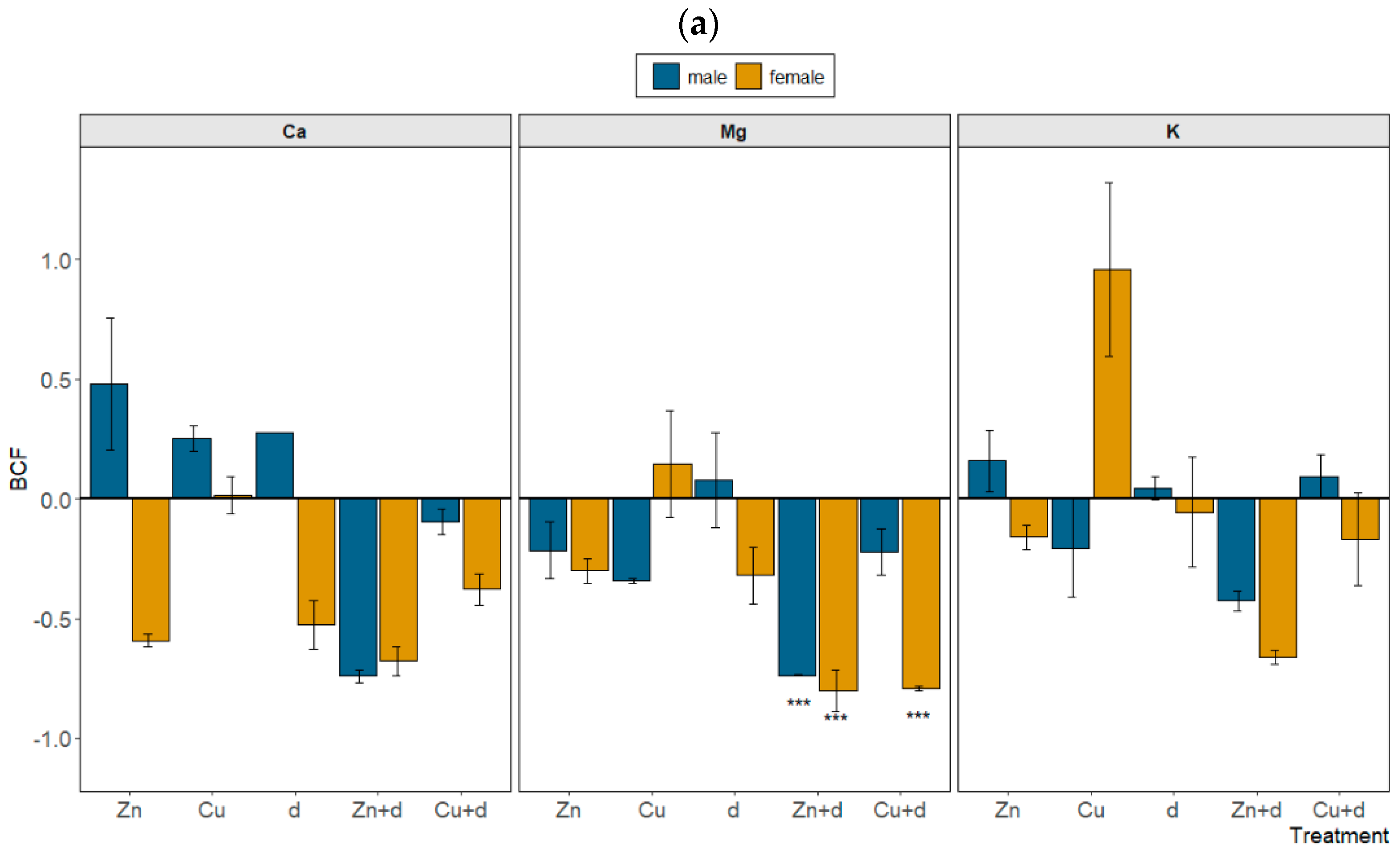
3.4. Mineral Nutrients Allocation
3.5. Metabolite Investigations
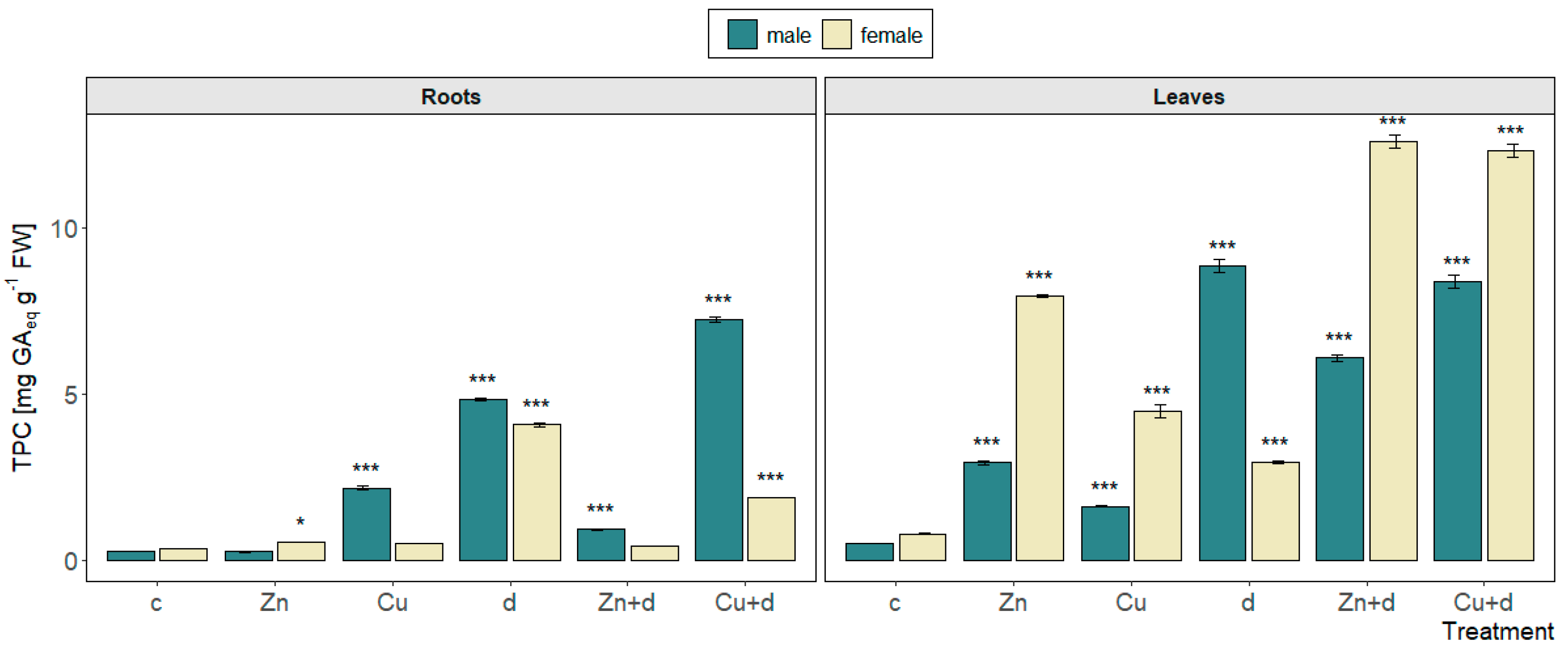
3.5.1. Total Phenolics Content
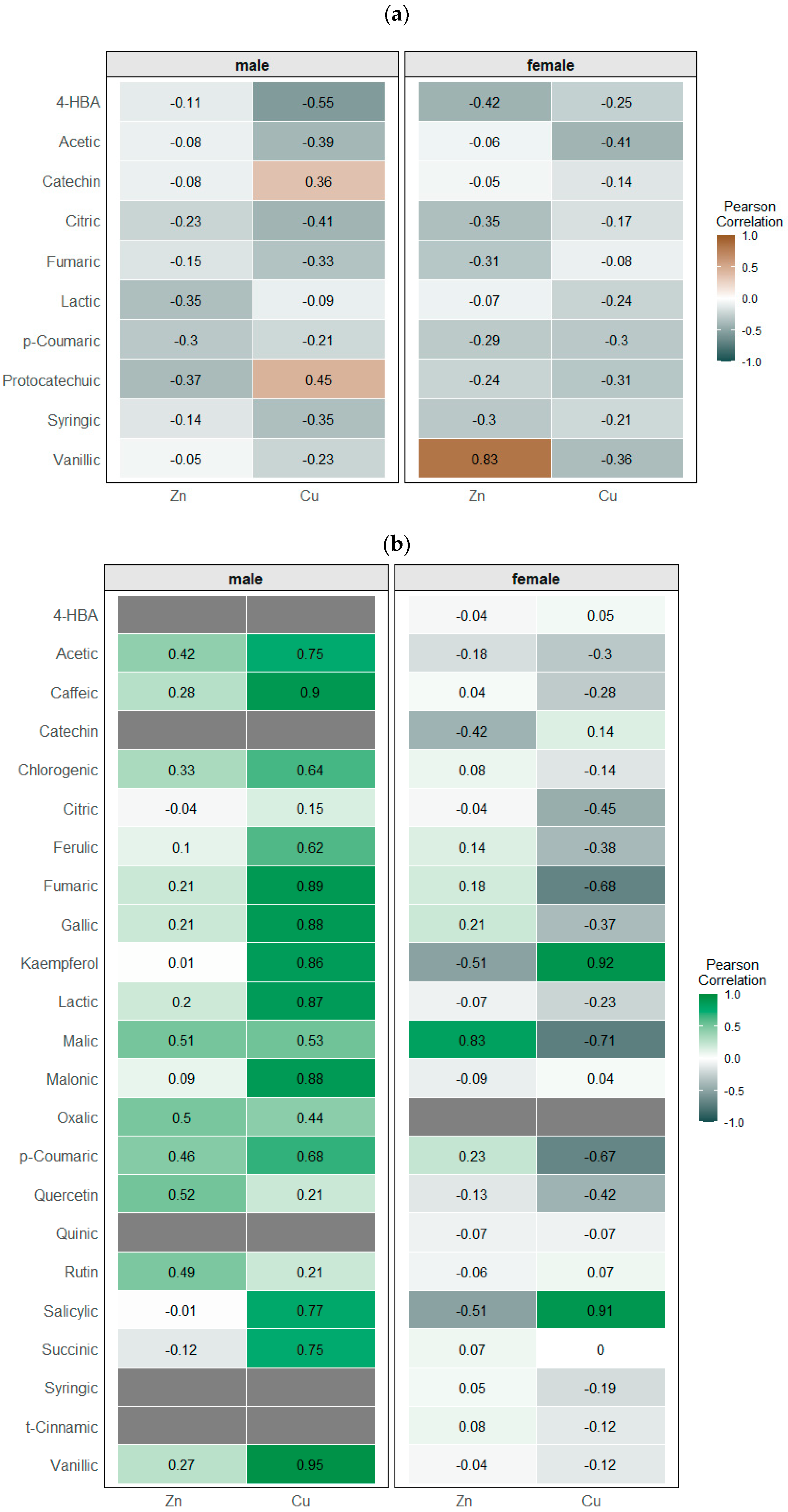
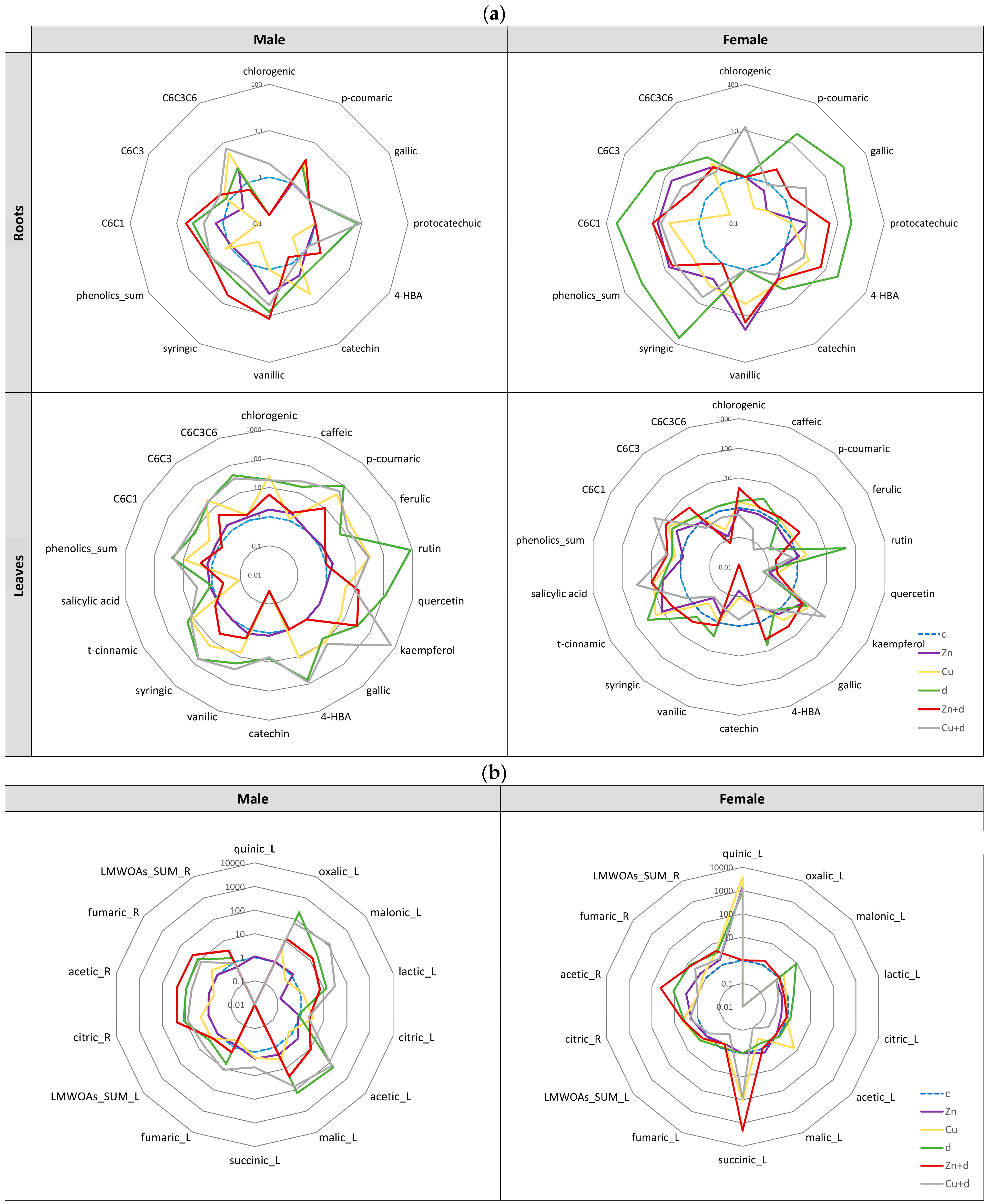
3.5.2. Profiling of Phenolic Metabolites
3.5.3. Profiling of Low-Molecular-Weight Organic Acids
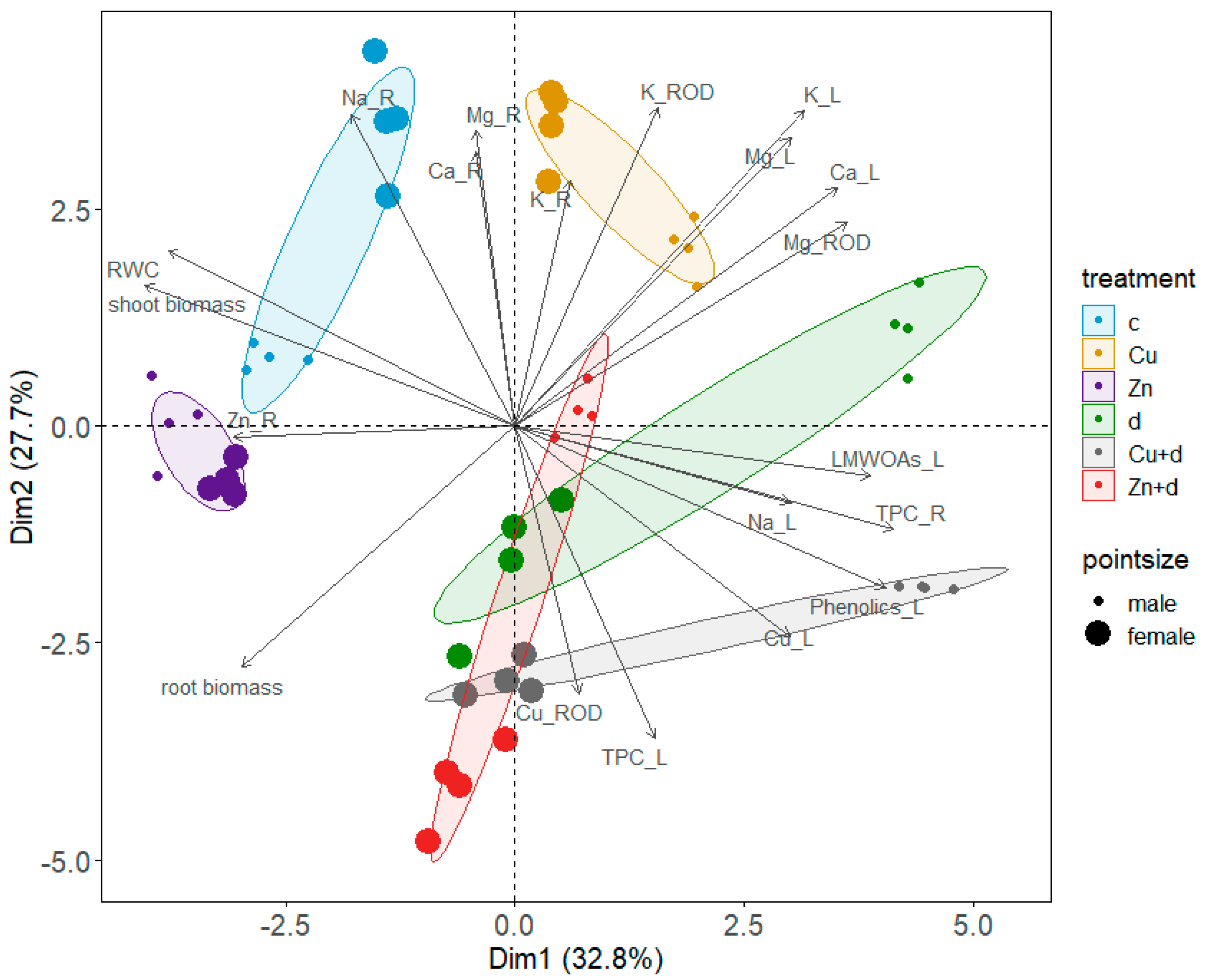
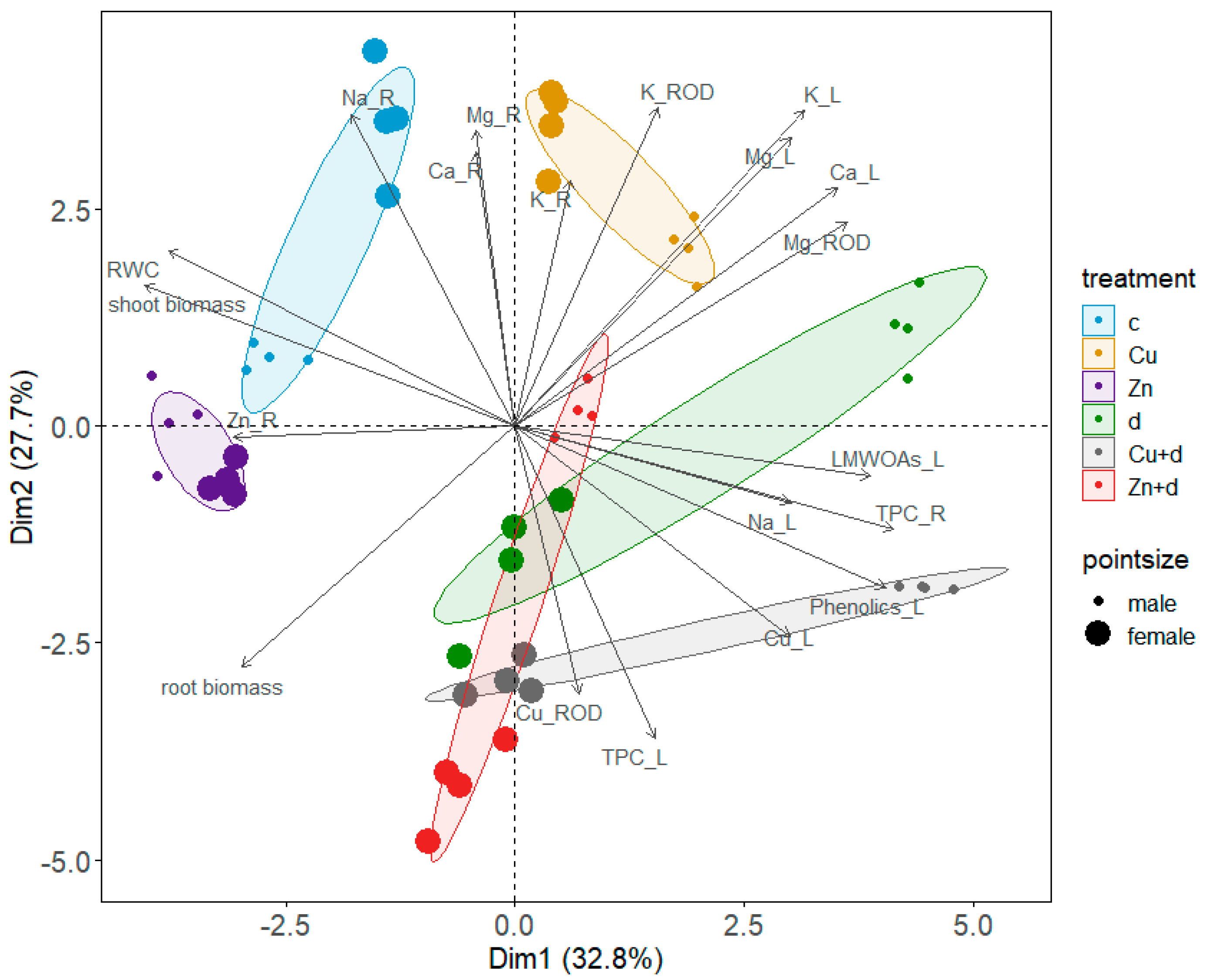
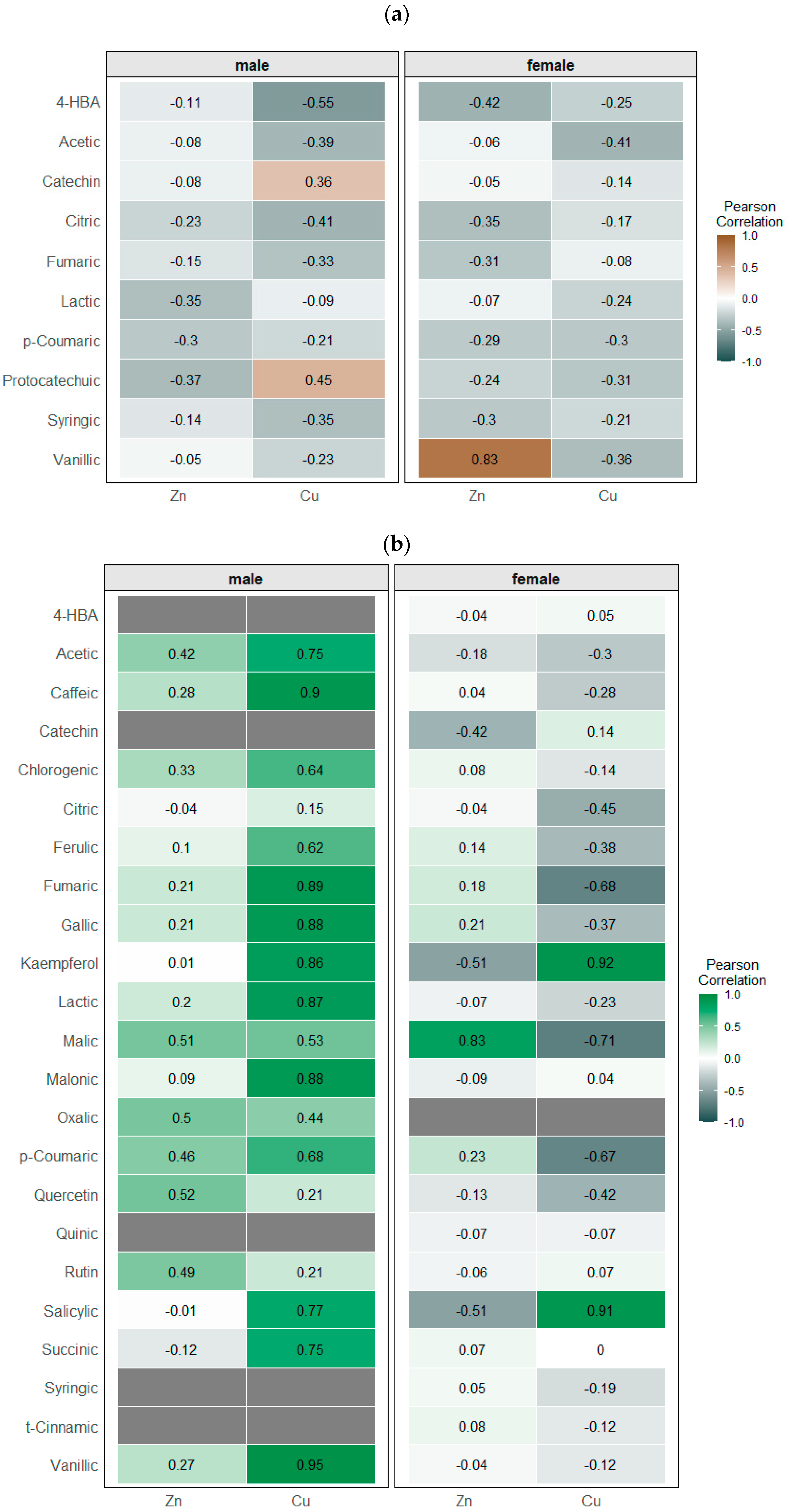
3.6. Principal Components Analysis (PCA) and Simple Linear Correlations
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Ali, H.; Khan, E.; Sajad, M.A. Phytoremediation of heavy metals—Concepts and applications. Chemosphere 2013, 91, 869–881. [Google Scholar] [CrossRef]
- Shen, X.; Dai, M.; Yang, J.; Sun, L.; Tan, X.; Peng, C.; Ali, I.; Naz, I. A critical review on the phytoremediation of heavy metals from environment: Performance and challenges. Chemosphere 2022, 291, 132979. [Google Scholar] [CrossRef]
- Oladoye, P.O.; Olowe, O.M.; Asemoloye, M.D. Phytoremediation technology and food security impacts of heavy metal contaminated soils: A review of literature. Chemosphere 2022, 288, 132555. [Google Scholar] [CrossRef]
- Sarwar, N.; Imran, M.; Shaheen, M.R.; Ishaque, W.; Kamran, M.A.; Matloob, A.; Rehim, A.; Hussain, S. Phytoremediation strategies for soils contaminated with heavy metals: Modifications and future perspectives. Chemosphere 2017, 171, 710–721. [Google Scholar] [CrossRef]
- Gavrilescu, M. Enhancing phytoremediation of soils polluted with heavy metals. Curr. Opin. Biotechnol. 2022, 74, 21–31. [Google Scholar] [CrossRef]
- Kafle, A.; Timilsina, A.; Gautam, A.; Adhikari, K.; Bhattarai, A.; Aryal, N. Phytoremediation: Mechanisms, plant selection and enhancement by natural and synthetic agents. Environ. Adv. 2022, 8, 100203. [Google Scholar] [CrossRef]
- Pouresmaieli, M.; Ataei, M.; Forouzandeh, P.; Azizollahi, P.; Mahmoudifard, M. Recent progress on sustainable phytoremediation of heavy metals from soil. J. Environ. Chem. Eng. 2022, 10, 108482. [Google Scholar] [CrossRef]
- Robinson, B.H.; Anderson, C.W.N.; Dickinson, N.M. Phytoextraction: Where’s the action? J. Geochem. Explor. 2015, 151, 34–40. [Google Scholar] [CrossRef]
- Müller, L.M.; Bahn, M. Drought legacies and ecosystem responses to subsequent drought. Glob. Chang. Biol. 2022, 28, 5086–5103. [Google Scholar] [CrossRef] [PubMed]
- Yao, Y.; Fu, B.; Liu, Y.; Li, Y.; Wang, S.; Zhan, T.; Wang, Y.; Gao, D. Evaluation of ecosystem resilience to drought based on drought intensity and recovery time. Agric. For. Meteorol. 2022, 314, 108809. [Google Scholar] [CrossRef]
- Hammond, W.M.; Williams, A.P.; Abatzoglou, J.T.; Adams, H.D.; Klein, T.; López, R.; Sáenz-Romero, C.; Hartmann, H.; Breshears, D.D.; Allen, C.D. Global field observations of tree die-off reveal hotter-drought fingerprint for Earth’s forests. Nat. Commun. 2022, 13, 1761. [Google Scholar] [CrossRef]
- Tian, K.; Wang, Y.; Chen, D.; Cao, M.; Luo, J. Influence of drought stress and post-drought rewatering on phytoremediation effect of Arabidopsis thaliana. Bull. Environ. Contam. Toxicol. 2022, 108, 594–599. [Google Scholar] [CrossRef]
- Lokhande, V.H.; Gor, B.K.; Desai, N.S.; Nikam, T.D.; Suprasanna, P. Sesuvium portulacastrum, a plant for drought, salt stress, sand fixation, food and phytoremediation. A review. Agron. Sustain. Dev. 2013, 33, 329–348. [Google Scholar] [CrossRef]
- Marchenko, A.M.; Kuzovkina, Y.A. Notes on the nomenclature and taxonomy of Salix fragilis (Salicaceae). Taxon 2022, 71, 721–732. [Google Scholar] [CrossRef]
- Clive, S. A S. × fragilis. In New Flora of the British Isles, 4th ed.; C & M Floristics: Stowmarket, UK, 2019; p. 348. [Google Scholar]
- Bita-Nicolae, C. Distribution of the riparian Salix communities in and around Romanian Carpathians. Diversity 2023, 15, 397. [Google Scholar] [CrossRef]
- Abdikaimova, T.; Dogan, I. Evaluation of heavy metal pollution along Alamedin River in Bishkek/Kyrgyzstan using woody species, Populus nigra (black poplar) and Salix fragilis (crack willow). 2022; Preprint. [Google Scholar] [CrossRef]
- Sharma, R.C.; Singh, N.B.; Tripathi, D.; Anil, S. Status and distribution of willow mortality in Lahaul and Spiti cold desert of Himachal Pradesh. Indian. For. 2011, 137, 196–204. [Google Scholar]
- Sharma, R.K.; Sharma, D. Drying willow (Salix fragilis L.) population under agroforestry system in cold desert region of Trans-Himalaya: A possible consequence of repeated vegetative propagation. Int. J. Ecol. Environ. Sci. 2022, 48, 119–125. [Google Scholar] [CrossRef]
- Henderson, L. Alien weeds and invasive plants. In Plant Protection Research Institute Handbook No 12; Agricultural Research Council: Pretoria, South Africa, 2001. [Google Scholar]
- Weber, E. Invasive Plant Species of the World: A Reference Guide to Environmental Weeds; CAB International: Wallingford, UK, 2003; p. 548. [Google Scholar]
- Magdziak, Z.; Mleczek, M.; Rutkowski, P.; Goliński, P. Diversity of low-molecular weight organic acids synthesized by Salix growing in soils characterized by different Cu, Pb and Zn concentrations. Acta Physiol. Plant 2017, 39, 137. [Google Scholar] [CrossRef]
- Drzewiecka, K.; Mleczek, M.; Gąsecka, M.; Magdziak, Z.; Budka, A.; Chadzinikolau, T.; Kaczmarek, Z.; Goliński, P. Copper and nickel co-treatment alters metal uptake and stress parameters of Salix purpurea × viminalis. J. Plant Physiol. 2017, 216, 125–134. [Google Scholar] [CrossRef]
- Osmolovskaya, N.; Shumilina, J.; Kim, A.; Didio, A.; Grishina, T.; Bilova, T.; Keltsieva, O.A.; Zhukov, V.; Tikhonovich, I.; Tarakhovskaya, E.; et al. Methodology of drought stress research: Experimental setup and physiological characterization. Int. J. Mol. Sci. 2018, 19, 4089. [Google Scholar] [CrossRef] [PubMed]
- Adeniji, B.A.; Budimir-Hussey, M.T.; Macfie, S.M. Production of organic acids and adsorption of Cd on roots of durum wheat (Triticum turgidum L. var. durum). Acta Physiol. Plant 2010, 32, 1063–1072. [Google Scholar] [CrossRef]
- Baker, A.J.M.; McGrath, S.P.; Sidoli, C.M.D.; Reeves, R.D. The possibility of in situ heavy metal decontamination of polluted soils using crops of metal-accumulating plants. Resour. Conserv. Recycl. 1994, 11, 41–49. [Google Scholar] [CrossRef]
- Weatherley, P.E.; Slatyer, R.O. Relationship between relative turgidity and diffusion pressure deficit in leaves. Nature 1957, 179, 1085–1086. [Google Scholar] [CrossRef]
- Baker, A.J. Accumulators and excluders-strategies in the response of plants to heavy metals. J. Plant Nutr. 1981, 3, 643–654. [Google Scholar] [CrossRef]
- Ma, L.Q.; Komar, K.M.; Tu, C.; Zhang, W.; Cai, Y.; Kennelley, E.D. A fern that hyperaccumulates arsenic. Nature 2001, 409, 579. [Google Scholar] [CrossRef]
- Dewanto, V.; Wu, X.; Adom, K.K.; Liu, R.H. Thermal processing enhances the nutritional value of tomatoes by increasing total antioxidant activity. J. Agric. Food Chem. 2002, 50, 3010–3014. [Google Scholar] [CrossRef]
- Magdziak, Z.; Gąsecka, M.; Budka, A.; Goliński, P.; Mleczek, M. Profile and concentration of the low molecular weight organic acids and phenolic compounds created by two-year-old Acer platanoides seedlings growing under different As forms. J. Hazard. Mater. 2020, 392, 122280. [Google Scholar] [CrossRef]
- Drzewiecka, K.; Gąsecka, M.; Magdziak, Z.; Budzyńska, S.; Szostek, M.; Niedzielski, P.; Budka, A.; Roszyk, E.; Doczekalska, B.; Górska, M.; et al. The possibility of using Paulownia elongata SY Hu × Paulownia fortunei hybrid for phytoextraction of toxic elements from post-industrial wastes with biochar. Plants 2021, 10, 2049. [Google Scholar] [CrossRef]
- Wickham, H. Elegant Graphics for Data Analysis: Ggplot2, Applied Spatial Data Analysis with R, Use R; Springer International Publishing: Cham, Switzerland, 2016. [Google Scholar]
- McElrone, A.J.; Choat, B.; Gambetta, G.A.; Brodersen, C.R. Water uptake and transport in vascular plants. Nat. Educ. Knowl. 2013, 4, 6. [Google Scholar]
- Seleiman, M.F.; Al-Suhaibani, N.; Ali, N.; Akmal, M.; Alotaibi, M.; Refay, Y.; Dindaroglu, T.; Abdul-Wajid, H.H.; Battaglia, M.L. Drought stress impacts on plants and different approaches to alleviate its adverse effects. Plants 2021, 10, 259. [Google Scholar] [CrossRef]
- Atta, K.; Pal, A.K.; Jana, K. Effects of salinity, drought and heavy metal stress during seed germination stage in ricebean [Vigna umbellata (Thunb.) Ohwi and Ohashi]. Plant Physiol. Rep. 2021, 26, 109–115. [Google Scholar] [CrossRef]
- Disante, K.B.; Fuentes, D.; Cortina, J. Response to drought of Zn-stressed Quercus suber L. seedlings. Environ. Exp. Bot. 2011, 70, 96–103. [Google Scholar] [CrossRef]
- Konatowska, M.; Rutkowski, P.; Budka, A.; Goliński, P.; Szentner, K.; Mleczek, M. The interactions between habitat, sex, biomass and leaf traits of different willow (Salix) genotypes. Int. J. Environ. Res. 2021, 15, 395–412. [Google Scholar] [CrossRef]
- Han, Y.; Wang, L.; Zhang, X.; Korpelainen, H.; Li, C. Sexual differences in photosynthetic activity, ultrastructure and phytoremediation potential of Populus cathayana exposed to lead and drought. Tree Physiol. 2013, 33, 1043–1060. [Google Scholar] [CrossRef] [PubMed]
- Qin, F.; Liu, G.; Huang, G.; Dong, T.; Liao, Y.; Xu, X. Zinc application alleviates the adverse effects of lead stress more in female Morus alba than in males. Environ. Exp. Bot. 2018, 146, 68–76. [Google Scholar] [CrossRef]
- Xu, X.; Yang, F.A.N.; Xiao, X.; Zhang, S.; Korpelainen, H.; Li, C. Sex-specific responses of Populus cathayana to drought and elevated temperatures. Plant Cell Environ. 2008, 31, 850–860. [Google Scholar] [CrossRef]
- Lin, T.; Tang, J.; He, F.; Chen, G.; Shi, Y.; Wang, X.; Han, S.; Li, S.; Zhu, T.; Chen, L. Sexual differences in above-and belowground herbivore resistance between male and female poplars as affected by soil cadmium stress. Sci. Total Environ. 2022, 803, 150081. [Google Scholar] [CrossRef] [PubMed]
- Utmazian, M.N.D.S.; Wieshammer, G.; Vega, R.; Wenzel, W.W. Hydroponic screening for metal resistance and accumulation of cadmium and zinc in twenty clones of willows and poplars. Environ. Pollut. 2007, 148, 155–165. [Google Scholar] [CrossRef]
- Chen, L.; Wang, L.; Chen, F.; Korpelainen, H.; Li, C. The effects of exogenous putrescine on sex-specific responses of Populus cathayana to copper stress. Ecotoxicol. Environ. Saf. 2013, 97, 94–102. [Google Scholar] [CrossRef]
- Jeyakumar, P.; Loganathan, P.; Sivakumaran, S.; Anderson, C.W.N.; McLaren, R.G. Bioavailability of copper and zinc to poplar and microorganisms in a biosolids-amended soil. Soil. Res. 2010, 48, 459–469. [Google Scholar] [CrossRef]
- Zimmer, D.; Kruse, J.; Baum, C.; Borca, C.; Laue, M.; Hause, G.; Meissner, R.; Leinweber, P. Spatial distribution of arsenic and heavy metals in willow roots from a contaminated floodplain soil measured by X-ray fluorescence spectroscopy. Sci. Total Environ. 2011, 409, 4094–4100. [Google Scholar] [CrossRef]
- Yang, W.D.; Wang, Y.Y.; Zhao, F.L.; Ding, Z.L.; Zhang, X.C.; Zhu, Z.Q.; Yang, X.E. Variation in copper and zinc tolerance and accumulation in 12 willow clones: Implications for phytoextraction. J. Zhejiang Univ. Sci. B 2014, 15, 788–800. [Google Scholar] [CrossRef]
- Santana, K.B.; de Almeida, A.-A.F.; Souza, V.L.; Mangabeira, P.A.O.; Silva, D.d.C.; Gomes, F.P.; Dutruch, L.; Loguercio, L.L. Physiological analyses of Genipa americana L. reveals a tree with ability as phytostabilizer and rhizofilterer of chromium ions for phytoremediation of polluted watersheds. Environ. Exp. Bot. 2012, 80, 35–42. [Google Scholar] [CrossRef]
- Umair Hassan, M.; Aamer, M.; Umer Chattha, M.; Haiying, T.; Shahzad, B.; Barbanti, L.; Nawaz, M.; Rasheed, A.; Afzal, A.; Liu, Y.; et al. The critical role of zinc in plants facing the drought stress. Agriculture 2020, 10, 396. [Google Scholar] [CrossRef]
- Waraich, E.A.; Ahmad, R.; Ashraf, M.Y. Role of mineral nutrition in alleviation of drought stress in plants. Aust. J. Crop Sci. 2011, 5, 764–777. [Google Scholar]
- Ahanger, M.A.; Morad-Talab, N.; Abd-Allah, E.F.; Ahmad, P.; Hajiboland, R. Plant growth under drought stress: Significance of mineral nutrients. In Water Stress and Crop Plants: A Sustainable Approach, 2 Volume Set; John Wiley & Sons, Inc.: Hoboken, NJ, USA, 2016; pp. 649–668. [Google Scholar] [CrossRef]
- Wang, S.; Shi, X.; Sun, H.; Chen, Y.; Pan, H.; Yang, X.; Rafiq, T. Variations in metal tolerance and accumulation in three hydroponically cultivated varieties of Salix integra treated with lead. PLoS ONE 2014, 9, e108568. [Google Scholar] [CrossRef] [PubMed]
- Nieves-Cordones, M.; Al Shiblawi, F.R.; Sentenac, H. Roles and transport of sodium and potassium in plants. The alkali metal ions: Their role for life. Met. Ions Life Sci. 2016, 16, 291–324. [Google Scholar] [PubMed]
- Wang, M.; Zheng, Q.; Shen, Q.; Guo, S. The Critical Role of Potassium in Plant Stress Response. Int. J. Mol. Sci. 2013, 14, 7370–7390. [Google Scholar] [CrossRef]
- Grabařová, S.; Martinková, M. Changes in mineral nutrition of Norway spruce (Picea abies [L.] Karst.) under the impact of drought. Ekologia 2001, 20, 46–60. [Google Scholar]
- Hu, Y.; Schmidhalter, U. Drought and salinity: A comparison of their effects on mineral nutrition of plants. J. Plant Nutr. Soil. Sci. 2005, 168, 541–549. [Google Scholar] [CrossRef]
- Sardans, J.; Penuelas, J.; Ogaya, R. Drought’s impact on Ca, Fe, Mg, Mo and S concentration and accumulation patters in the plants and soil a Mediterranean evergreen Quercus ilex forest. Biogeochemistry 2008, 87, 49–69. [Google Scholar] [CrossRef]
- Hafsi, C.; Debez, A.; Abdelly, C. Potassium deficiency in plants: Effects and signaling cascades. Acta Physiol. Plant 2014, 36, 1055–1070. [Google Scholar] [CrossRef]
- Ali, N.; Hadi, F. Phytoremediation of cadmium improved with the high production of endogenous phenolics and free proline contents in Parthenium hysterophorus plant treated exogenously with plant growth regulator and chelating agent. Environ. Sci. Pollut. Res. 2015, 22, 13305–13318. [Google Scholar] [CrossRef] [PubMed]
- Sakihama, Y. Plant phenolic antioxidant and prooxidant activities: Phenolics-induced oxidative damage mediated by metals in plants. Toxicology 2002, 177, 67–80. [Google Scholar] [CrossRef] [PubMed]
- Dehghanian, Z.; Habibi, K.; Dehghanian, M.; Aliyar, S.; Asgari Lajayer, B.; Astatkie, T.; Minkina, T.; Keswani, C. Reinforcing the bulwark: Unravelling the efficient applications of plant phenolics and tannins against environmental stresses. Heliyon 2022, 8, e09094. [Google Scholar] [CrossRef] [PubMed]
- Almughraby, E.; Kalimullin, M.I.; Timofeeva, O.A. Phytochemical composition and antioxidant activity in Brassica oleracea var. Sabellica under the effect of plant growth regulators. Eurasia J. Biosci. 2019, 13, 1037–1043. [Google Scholar]
- Schützendübel, A.; Schwanz, P.; Teichmann, T.; Gross, K.; Langenfeld-Heyser, R.; Godbold, D.L.; Andrea Polle, A. Cadmium-induced changes in antioxidative systems, hydrogen peroxide content, and differentiation in Scots pine roots. Plant Physiol. 2001, 127, 887–898. [Google Scholar] [CrossRef] [PubMed]
- Drzewiecka, K.; Piechalak, A.; Goliński, P.; Gąsecka, M.; Magdziak, Z.; Szostek, M.; Budzyńska, S.; Niedzielski, P.; Mleczek, M. Differences of Acer platanoides L. and Tilia cordata Mill. response patterns/survival strategies during cultivation in extremely polluted mining sludge—A Pot Trial. Chemosphere 2019, 229, 589–601. [Google Scholar] [CrossRef]
- Mleczek, M.; Budka, A.; Gąsecka, M.; Budzyńska, S.; Drzewiecka, K.; Magdziak, Z.; Rutkowski, P.; Goliński, P.; Niedzielski, P. Copper, lead and zinc interactions during phytoextraction using Acer platanoides L.—A pot trial. Environ. Sci. Pollut. Res. 2022, 30, 27191–27207. [Google Scholar] [CrossRef]
- Uraguchi, S.; Watanabe, I.; Yoshitomi, A.; Kiyono, M.; Kuno, K. Characteristics of cadmium accumulation and tolerance in novel Cd-accumulating crops, Avena strigosa and Crotalaria juncea. J. Exp. Bot. 2006, 57, 2955–2965. [Google Scholar] [CrossRef]
- Budny, M.; Zalewski, K.; Stolarski, M.J.; Wiczkowski, W.; Okorski, A.; Stryiński, R. The phenolic compounds in the young shoots of selected willow cultivars as a determinant of the plants’ attractiveness to cervids (Cervidae, Mammalia). Biology 2021, 10, 612. [Google Scholar] [CrossRef]
- Dixon, R.A.; Achnine, L.; Kota, P.; Liu, C.J.; Reddy, M.S.; Wang, L. The phenylpropanoid pathway and plant defence—A genomics perspective. Mol. Plant Pathol. 2002, 3, 371–390. [Google Scholar] [CrossRef]
- Facchini, P.J.; Hagel, J.; Zulak, K.G. Hydroxycinnamic acid amide metabolism: Physiology and biochemistry. Can. J. Bot. 2002, 80, 577–589. [Google Scholar] [CrossRef]
- Kováčik, J.; Klejdus, B.; Hedbavny, J.; Zoń, J. Significance of phenols in cadmium and nickel uptake. J. Plant Physiol. 2011, 168, 576–584. [Google Scholar] [CrossRef] [PubMed]
- Lavid, N.; Schwartz, A.; Lewinsohn, E.; Tel-Or, E. Phenols and phenol oxidases are involved in cadmium accumulation in the water plants Nymphoides peltata (Menyanthaceae) and Nymphaeae (Nymphaeaceae). Planta 2001, 214, 189–195. [Google Scholar] [CrossRef] [PubMed]
- Ray, S.; Mishra, S.; Bisen, K.; Singh, S.; Sarma, B.K.; Singh, H.B. Modulation in phenolic root exudate profile of Abelmoschus esculentus expressing activation of defense pathway. Microbiol. Res. 2018, 207, 100–107. [Google Scholar] [CrossRef]
- Hura, T.; Hura, K.; Ostrowska, A.; Grzesiak, M.; Dziurka, K. The cell wall-bound phenolics as a biochemical indicator of soil drought resistance in winter triticale. Plant Soil. Environ. 2013, 59, 189–195. [Google Scholar] [CrossRef]
- Ballizany, W.L.; Hofmann, R.W.; Jahufer, M.Z.; Barrett, B.A. Multivariate associations of flavonoid and biomass accumulation in white clover (Trifolium repens) under drought. Funct. Plant Biol. 2012, 39, 167–177. [Google Scholar] [CrossRef] [PubMed]
- Bautista, I.; Boscaiu, M.; Lidón, A.; Llinares, J.V.; Lull, C.; Donat, P.; Mayoral, O. Environmentally induced changes in antioxidant phenolic compounds levels in wild plants. Acta Physiol. Plant 2016, 38, 9. [Google Scholar] [CrossRef]
- Fini, A.; Brunetti, C.; Di Ferdinando, M.; Ferrini, F.; Tattini, M. Stress-induced flavonoid biosynthesis and the antioxidant machinery of plants. Plant Signal. Behav. 2011, 6, 709–711. [Google Scholar] [CrossRef] [PubMed]
- Köhler, A.; Förster, N.; Zander, M.; Ulrichs, C. Compound-specific responses of phenolic metabolites in the bark of drought-stressed Salix daphnoides and Salix purpurea. Plant Physiol. Biochem. 2020, 155, 311–320. [Google Scholar] [CrossRef]
- Turtola, S.; Rousi, M.; Pusenius, J.; Yamaji, K.; Heiska, S.; Tirkkonen, V.; Meier, B.H.; Julkunen-Tiitto, R. Clone-specific responses in leaf phenolics of willows exposed to enhanced UVB radiation and drought stress. Glob. Chang. Biol. 2005, 11, 1655–1663. [Google Scholar] [CrossRef]
- Bhatia, N.P.; Walsh, K.B.; Baker, A.J. Detection and quantification of ligands involved in nickel detoxification in a herbaceous Ni hyperaccumulator Stackhousia tryonii Bailey. J. Exp. Bot. 2005, 56, 1343–1349. [Google Scholar] [CrossRef]
- Callahan, D.L.; Baker, A.J.; Kolev, S.D.; Wedd, A.G. Metal ion ligands in hyperaccumulating plants. J. Biol. Inorg. Chem. 2006, 11, 2–12. [Google Scholar] [CrossRef] [PubMed]
- Zacchini, M.; Iori, V.; Scarascia Mugnozza, G.; Pietrini, F.; Massacci, A. Cadmium accumulation and tolerance in Populus nigra and Salix alba. Biol. Plant 2011, 55, 383–386. [Google Scholar] [CrossRef]
- Pietrini, F.; Iori, V.; Cheremisina, A.; Shevyakova, N.I.; Radyukina, N.; Kuznetsov, V.V.; Zacchini, M. Evaluation of nickel tolerance in Amaranthus paniculatus L. plants by measuring photosynthesis, oxidative status, antioxidative response and metal-binding molecule content. Environ. Sci. Pollut. Res. 2015, 22, 482–494. [Google Scholar] [CrossRef] [PubMed]
- Hou, X.; Han, H.; Cai, L.; Liu, A.; Ma, X.; Zhou, C.; Wang, G.; Meng, F. Pb stress effects on leaf chlorophyll fluorescence, antioxidative enzyme activities, and organic acid contents of Pogonatherum crinitum seedlings. Flora 2018, 240, 82–88. [Google Scholar] [CrossRef]
- Dastogeer, K.M.; Li, H.; Sivasithamparam, K.; Jones, M.G.; Du, X.; Ren, Y.; Wylie, S.J. Metabolic responses of endophytic Nicotiana benthamiana plants experiencing water stress. Environ. Exp. Bot. 2017, 143, 59–71. [Google Scholar] [CrossRef]
- Khan, N.; Ali, S.; Zandi, P.; Mehmood, A.; Ullah, S.; Ikram, M.; Ismail, I.; Shahid, M.; Babar, A. Role of sugars amino acids organic acids in improving plant abiotic stress tolerance Pak. J. Bot. 2020, 52, 355–363. [Google Scholar] [CrossRef]
- Shi, X.; Wang, S.; Wang, D.; Sun, H.; Chen, Y.; Liu, J.; Jiang, Z. Woody species Rhus chinensis Mill. seedlings tolerance to Pb: Physiological and biochemical response. J. Environ. Sci. 2019, 78, 63–73. [Google Scholar] [CrossRef] [PubMed]
- Naya, L.; Ladrera, R.; Ramos, J.; González, E.M.; Arrese-Igor, C.; Minchin, F.R.; Becana, M. The response of carbon metabolism and antioxidant defenses of alfalfa nodules to drought stress and to the subsequent recovery of plants. Plant Physiol. 2007, 144, 1104–1114. [Google Scholar] [CrossRef] [PubMed]
- Ullah, N.; Yüce, M.; Neslihan Öztürk Gökçe, Z.; Budak, H. Comparative metabolite profiling of drought stress in roots and leaves of seven Triticeae species. BMC Genom. 2017, 18, 969. [Google Scholar] [CrossRef] [PubMed]
- Gargallo-Garriga, A.; Preece, C.; Sardans, J.; Oravec, M.; Urban, O.; Peñuelas, J. Root exudate metabolomes change under drought and show limited capacity for recovery. Sci. Rep. 2018, 8, 12696. [Google Scholar] [CrossRef] [PubMed]
- Levi, A.; Paterson, A.H.; Cakmak, I.; Saranga, Y. Metabolite and mineral analyses of cotton near-isogenic lines introgressed with QTLs for productivity and drought-related traits. Physiol. Plant 2011, 141, 265–275. [Google Scholar] [CrossRef]
- Goliński, P.; Rutkowski, P.; Waliszewska, B.; Stolarski, M.; Czapiewski, G.; Szentner, K.; Gąsecka, M.; Magdziak, Z.; Mleczek, M. Poplars and Willows. In Cultivation, Applications and Environmental Benefits. Chapter 2: Salix: Properties and Practical Purposes; Nova Publishers: New York, NY, USA, 2016; pp. 29–73. [Google Scholar]


Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Drzewiecka, K.; Gąsecka, M.; Magdziak, Z.; Rybak, M.; Budzyńska, S.; Rutkowski, P.; Niedzielski, P.; Mleczek, M. Drought Differently Modifies Tolerance and Metal Uptake in Zn- or Cu-Treated Male and Female Salix × fragilis L. Forests 2024, 15, 562. https://doi.org/10.3390/f15030562
Drzewiecka K, Gąsecka M, Magdziak Z, Rybak M, Budzyńska S, Rutkowski P, Niedzielski P, Mleczek M. Drought Differently Modifies Tolerance and Metal Uptake in Zn- or Cu-Treated Male and Female Salix × fragilis L. Forests. 2024; 15(3):562. https://doi.org/10.3390/f15030562
Chicago/Turabian StyleDrzewiecka, Kinga, Monika Gąsecka, Zuzanna Magdziak, Michał Rybak, Sylwia Budzyńska, Paweł Rutkowski, Przemysław Niedzielski, and Mirosław Mleczek. 2024. "Drought Differently Modifies Tolerance and Metal Uptake in Zn- or Cu-Treated Male and Female Salix × fragilis L." Forests 15, no. 3: 562. https://doi.org/10.3390/f15030562
APA StyleDrzewiecka, K., Gąsecka, M., Magdziak, Z., Rybak, M., Budzyńska, S., Rutkowski, P., Niedzielski, P., & Mleczek, M. (2024). Drought Differently Modifies Tolerance and Metal Uptake in Zn- or Cu-Treated Male and Female Salix × fragilis L. Forests, 15(3), 562. https://doi.org/10.3390/f15030562