
Reference Curves for Central Appalachian Red Spruce


Abstract
:1. Introduction
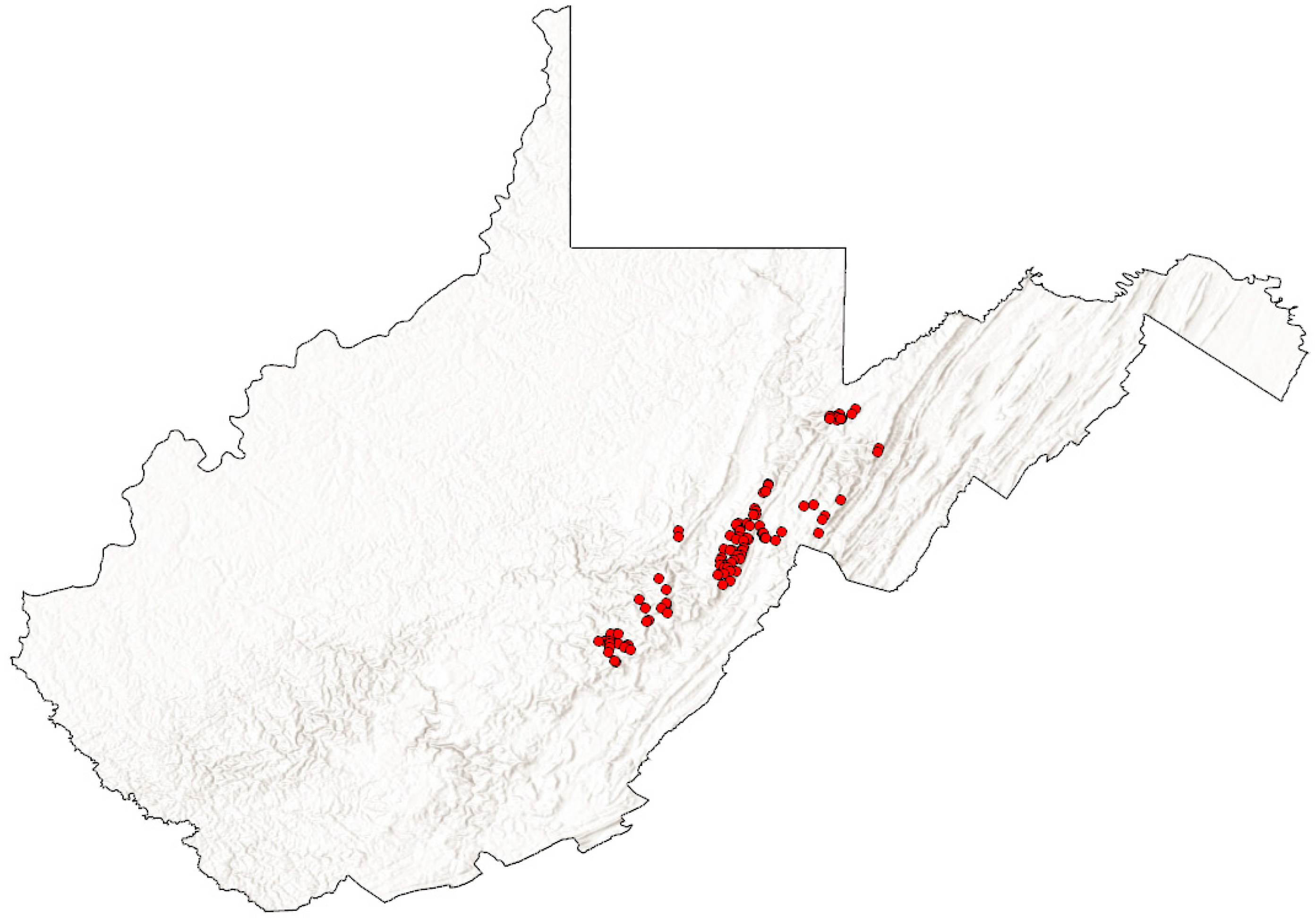
2. Materials and Methods
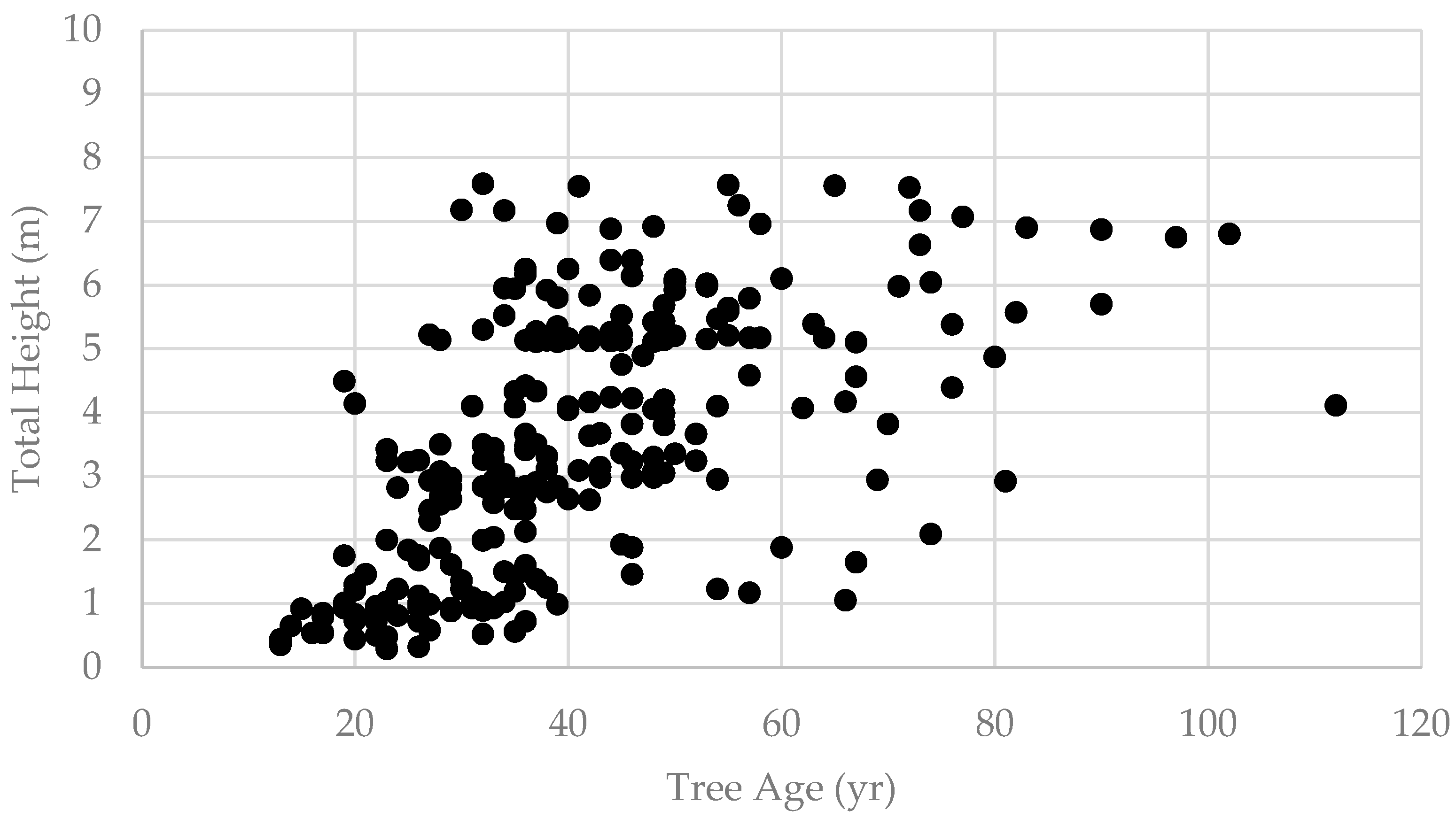
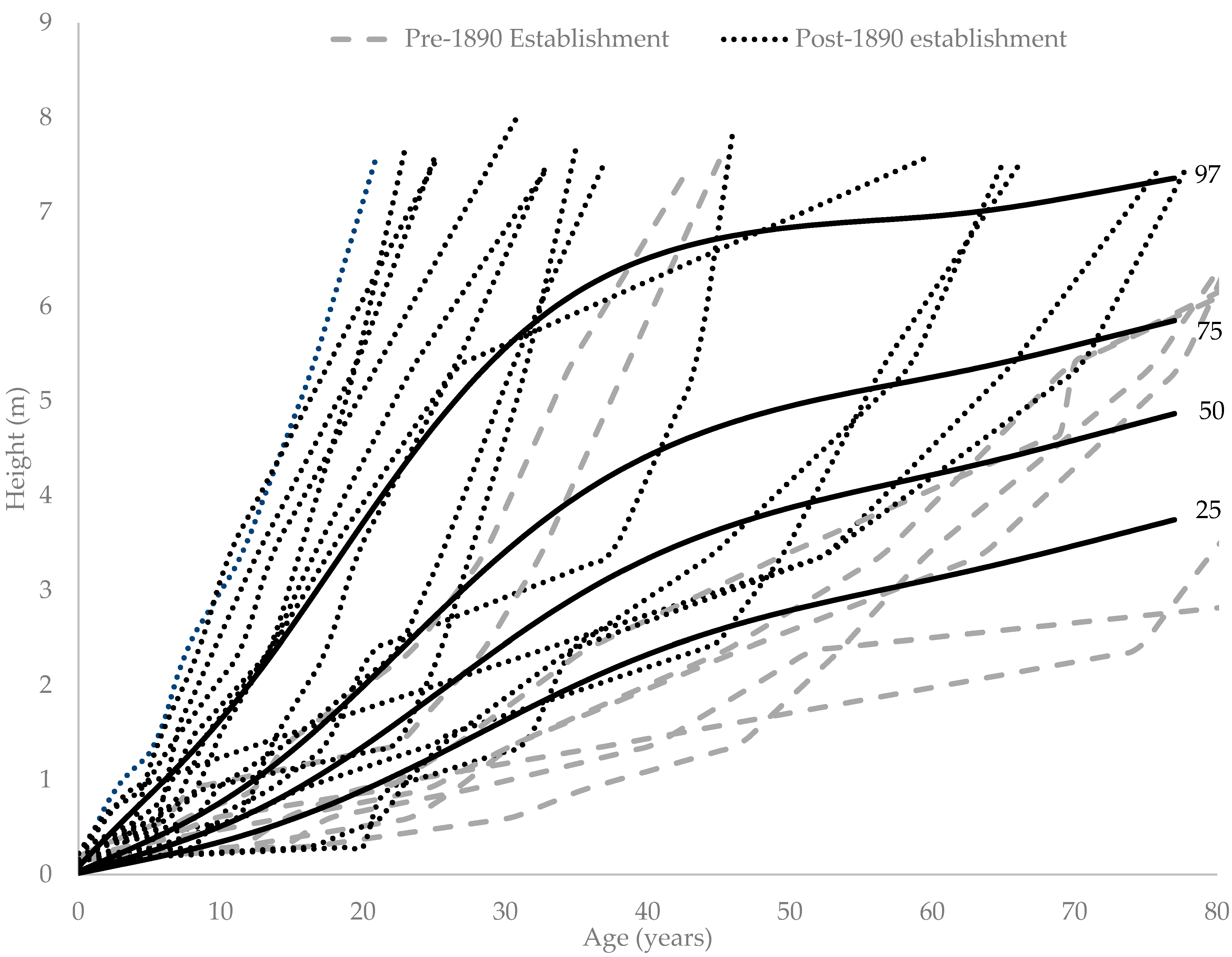
3. Results
4. Discussion
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Hopkins, A.D. Report on investigations to determine the cause of unhealthy conditions of the spruce and pine from 1880–1893. Part I, the spruce investigation. West Virginia Agricultural Experiment Station. Bulletin 1899, 56, 197–270. [Google Scholar]
- Strausbaugh, P.D.; Core, E.L. Flora of West Virginia (Part 1); West Virginia University Bulletin: Morgantown, WV, USA, 1952; p. 273. [Google Scholar]
- Lewis, R.L. Transforming the Appalachian Countryside: Railroads, Deforestation, and Social Change in West Virginia, 1890–1920; The University of North Carolina Press: Chapel Hill, NC, USA, 1998; 348p. [Google Scholar]
- Gundy, M.A.T.-V.; Sturtevant, B.R. Using scenario modeling for red spruce. J. For. 2014, 112, 457–466. [Google Scholar]
- Adams, H.S.; Stephenson, S.L. Old-growth red spruce communities in the mid-Appalachians. Vegetation 1989, 85, 45–56. [Google Scholar] [CrossRef]
- Griffith, D.M.; Widmann, R.H. Forest Statistics for West Virginia: 1989 and 2000; USDA Forest Service Resource Bulletin NE-157: Newtown Square, PA, USA, 2003. [Google Scholar]
- Schuler, T.M.; Ford, W.M.; Collins, R.J. Successional dynamics and restoration implications of a montane coniferous forest in the central Appalachians, USA. Nat. Areas J. 2002, 22, 88–98. [Google Scholar]
- Soulé, P.T. Changing Climate, Atmospheric Composition, and radial tree growth in a spruce-fir ecosystem on Grandfather Mountain, North Carolina. Nat. Areas J. 2011, 31, 65–74. [Google Scholar] [CrossRef]
- Rollins, A.W. Analysis of Red Spruce (Picea rubens) Regeneration in Pocahontas, Randolph, and Tucker Counties, West Virginia. Master’s Thesis, West Virginia University, Morgantown, WV, USA, 2005. [Google Scholar]
- Mayfield, A.E.; Hicks, J.R. Abundance of red spruce regeneration across spruce-hardwood ecotones at Gaudineer Knob, West Virginia. In Ecology and Management of High Elevation Forests in the Central and Southern Appalachian Mountains; Rentch, J.S., Schuler, T.M., Eds.; U.S. Department of Agriculture, Forest Service, Northern Research Station: Slatyfork, WV, USA, 2010; pp. 113–125. [Google Scholar]
- Nowacki, G.; Carr, R.; Van Dyck, M. The current status of red spruce in the Eastern United States: Distribution, population trends, and environmental drivers. In Proceedings of the Ecology and Management of High-Elevation Forests in the Central and Southern Appalachian Mountains, Slatyfork, WV, USA, 14–15 2009 May; General Technical Report NRS-P-64. USDA, Forest Service, Northern Research Station: Slatyfork, WV, USA, 2010; pp. 140–162. [Google Scholar]
- Adams, H.S.; Stephenson, S.L.; Blasing, T.; Duvick, D. Growth-trend declines of spruce and fir in mid-Appalachian subalpine forests. Environ. Exp. Bot. 1985, 25, 315–325. [Google Scholar] [CrossRef]
- Beane, N.R.; Rentch, J.S.; Schuler, T.M. Using Maximum Entropy Modeling to Identify and Prioritize Red Spruce Forest Habitat in West Virginia; Research Paper NRS-23; U.S. Department of Agriculture, Forest Service, Northern Research Station: Newtown Square, PA, USA, 2015; 16p.
- Lutz, A. Characterizing Red Spruce (Picea rubens Sarg.) Advanced Reproduction in a High Elevation Stand in West Virginia; Graduate Theses, Dissertations, and Problem Reports; West Virginia University: Morgantown, WV, USA, 2018; p. 7209. [Google Scholar]
- Rentch, J.S.; Schuler, T.M.; Ford, W.M.; Nowacki, G.J. Red Spruce stand dynamics, simulations, and restoration opportunities in the central Appalachians. Restor. Ecol. 2007, 15, 440–452. [Google Scholar] [CrossRef]
- Yetter, E.; Chhin, S.; Brown, J.P. Dendroclimatic analysis of Central Appalachian red spruce in West Virginia. Can. J. For. Res. 2021, 51, 1607–1620. [Google Scholar] [CrossRef]
- Adams, M.B.; Eager, C. Effects of acidic deposition on high-elevation spruce-fir forests in the United States. In Acid Depsoition; Longhurst, J.W.S., Ed.; Springer: Berlin/Heidelberg, Germany, 1991. [Google Scholar]
- Rentch, J.S.; Ford, W.M.; Schuler, T.M.; Palmer, J.; Diggins, C.A. Release of suppressed red spruce using canopy gap creation- ecological restoration in the central Appalachians. Nat. Areas J. 2016, 36, 29–37. [Google Scholar] [CrossRef]
- Dumais, D.; Prévost, M. Germination and establishment of natural red spruce (Picea rubens) seedlings in silvicultural gaps of different sizes. For. Chron. 2016, 92, 90–100. [Google Scholar] [CrossRef]
- Seymour, R.S. The red spruce-balsam fir fores of Maine: Evolution of silvicultural practice in response to stand development patterns and disturbances. In The Ecology and Silviculture of Mixed-Species Forests; Kelty, M.J., Larson, B.C., Oliver, C.D., Eds.; Forestry Sciences; Springer: Dordrecht, The Netherlands, 1992; p. 40. [Google Scholar]
- Stehn, S.E.; Webster, C.R.; Jenkins, M.A.; Jose, S. High-elevation ground-layer plant community composition across environmental gradients in spruce-fir forests. Ecol. Res. 2011, 26, 1089–1101. [Google Scholar] [CrossRef]
- Rollins, A.W.; Adams, H.S.; Stephenson, S.L. Changes in forest composition and structure across the red spruce-hardwood ecotone in the Central Appalachians. Castanea 2010, 75, 303–314. [Google Scholar] [CrossRef]
- Rentch, J.S.; Schuler, T.M.; Nowacki, G.J.; Beane, N.R.; Ford, W.M. Canopy gap dynamics of second-growth red spruce-northern hardwood stands in West Virginia. For. Ecol. Manag. 2010, 260, 1921–1929. [Google Scholar] [CrossRef]
- Rhodes, B. Evaluating Restoration Outcomes: Red Spruce Reforestation in the West Virginia Highlands. Master’s Thesis, University of Kentucky, Lexington, Kentucky, 2022. [Google Scholar]
- Vickers, L.A.; Larsen, D.R.; Knapp, B.O.; Kabrick, J.M.; Dey, D.C. Reference charts for young stands—A quantitative methodology for assessing tree performance. Can. J. For. Res. 2017, 47, 1677–1686. [Google Scholar] [CrossRef]
- Theobald, D.M.; Stevens, D.L.; White, D.; Urquhart, N.S.; Olsen, A.R.; Norman, J.B. Norman. Using GIS to generate spatially balanced random survey designs for natural resources applications. Environ. Assess. 2007, 46, 134–146. [Google Scholar]
- West Virginia GIS Technical Center. Red Spruce (Picea rubens) Cover in West Virginia. 2013. Available online: https://wvgis.wvu.edu (accessed on 20 July 2020).
- National Climatic Data Center. Monthly Summaries, Pocahontas, Randolph, Tucker Counties, WV. 2020. Available online: www.ncdc.noaa.gov/cdo-web (accessed on 21 July 2020).
- PRISM Climate Group. Oregon State University. 30 Year Normals. 2020. Available online: http://prism.oregonstate.edu (accessed on 20 July 2020).
- Jenkins, A.B. Organic Carbon and Fertility of Forest Soils on the Allegheny Plateau of West Virginia. Master’s Thesis, West Virginia University, Morgantown, WV, USA, 2002. [Google Scholar]
- Price, P.H. Geologic Map of West Virginia; WV Geologic and Economic Survey: Morgantown, WV, USA, 1968. [Google Scholar]
- Thomas-Van Gundy, M.A.; Nowacki, G.J.; Schuler, T.M. Rule-Based Mapping of Fire-Adapted Vegetation and Fire Regimes for the Monongahela National Forest; Gen. Tech. Rep. NRS-12; U.S. Department of Agriculture, Forest Service, Northern Research Station: Newtown Square, PA, USA, 2007; 24p.
- Yamaguchi, D.K. A simple method for cross-dating increment cores from living trees. Can. J. For. Res. 1991, 21, 414–416. [Google Scholar] [CrossRef]
- Grissino-Mayer, H.D. Evaluating crossdating accuracy: A manual and tutorial for the computer program Cofecha. Tree-Ring Res. 2001, 57, 205–221. [Google Scholar]
- Stokes, M.A.; Smiley, T.L. An Introduction to Tree-Ring Dating; University of Chicago Press: Chicago, IL, USA, 1968. [Google Scholar]
- Carmean, W.H. Site index curves for upland oaks in the Central States. For. Sci. 1972, 18, 109–120. [Google Scholar]
- Winkler; Linden, K.; Mayr, A.; Schultz, T.; Welchowski, T.; Breuer, J.; Herberg, U. RefCurv: A Software for the Construction of Pediatric Reference Curves. Softw. Impacts 2019, 6, 100040. [Google Scholar] [CrossRef]
- Cole, T.J. The LMS method for constructing normalized growth standards. Eur. J. Clin. Nutr. 1990, 44, 45–60. [Google Scholar]
- Stasinopoulos, D.M.; Rigby, R.A. Generalized additive models for location scale and shape (GAMLSS) in R. J. Stat. Softw. 2007, 23, 1–46. [Google Scholar] [CrossRef]
- Gray, J.M. Central Appalachian Understory Red Spruce (Picea rubens Sarg.) Growth Rates and Allometric Relationships. Master’s Thesis, West Virginia University, Morgantown, WV, USA, 2020. [Google Scholar]
- Newton, M.; Cole, E.C.; McCormack, M.L., Jr.; White, D.E. Young Spruce-Fir Forests Released by Herbicides II. Conifer Response to Residual Hardwoods and Overstocking. North. J. Appl. For. 1993, 9, 130–135. [Google Scholar] [CrossRef]
- Blum, B.M. Red spruce. In Silvics of North America: 1. Conifers; 2. Hardwoods; Agriculture Handbook 654; Burns, R.M., Barbara, H., Honkala, U.S., Eds.; Department of Agriculture, Forest Service: Washington, DC, USA, 1990; Volume 2, 877p. [Google Scholar]
- McCormick, F.J.; Busing, R.T. Growth pattern of Picea rubens prior to canopy recruitment. Plant Ecol. 1999, 140, 245–253. [Google Scholar]
- Morin, R.S.; Widmann, R.H. A comparison of the status of spruce in high-elevation forests on public and private lands in the southern and central Appalachian Mountains. In Proceedings of the Ecology and Management of High-Elevation Forests in the Central and Southern Appalachian Mountains, Slatyfork, MV, USA, 14–15 May 2009; Rentch, J.S., Schuler, T.M., Eds.; Gen. Tech. Rep. NRS-P-64. U.S. Department of Agriculture, Forest Service, Northern Research Station: Newtown Square, PA, USA, 2010; pp. 132–139. [Google Scholar]
- Mielke, M.E.; Soctomah, D.C.; Marsden, M.A.; Ciesla, W.M. Decline and Mortality of Red Spruce in West Virginia; USDA Forest Service Report 86-4; U.S. Department of Agriculture, Forest Service, Northern Research Station: Newtown Square, PA, USA, 1986.
- Johnson, A.H.; Cook, E.R.; Siccama, T.G. Climate and red spruce growth and decline in the northern Appalachians. Proc. Natl. Acad. Sci. USA 1988, 85, 5369–5373. [Google Scholar] [CrossRef] [PubMed]
- McLaughlin, S.B.; Downing, D.J.; Blasing, T.J.; Cook, E.R.; Adams, H.S. An analysis of climate and competition as contributors to decline of red spruce in high elevation Appalachian forests of the eastern United States. Oecologia 1987, 72, 487–501. [Google Scholar] [CrossRef] [PubMed]
- Hamburg, S.P.; Cogbill, C.V. Historical decline of red spruce populations and climatic warming. Nature 1988, 331, 428–431. [Google Scholar] [CrossRef]
- Allard, L.E. The Canaan and the Stony River Valleys of West Virginia, their former magnificent spruce forests, their vegetation and floristics today. Castanea 1952, 17, 1–60. [Google Scholar]
- Moores, A.R.; Seymour, R.S.; Kenefic, L.S. Height development of shade-tolerant conifer saplings in multiaged Acadian forest stands. Can. J. For. Res. 2007, 37, 2715–2723. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Height Class | N | Height (m) | Live Crown Height (m) | DBH (cm) | Crown Width (m) | Age (years) | Annual Height Growth (m) |
|---|---|---|---|---|---|---|---|
| 1 (0.1–2.6 m) | 83 | 1.22 (0.59) | 0.53 (0.42) | 1.17 (1.00) | 0.68 (0.40) | 29.8 (12.0) | 0.04 (0.02) |
| 2 (2.6–5.1 m) | 85 | 3.47 (0.62) | 1.50 (0.94) | 3.84 (1.28) | 1.27 (0.48) | 42.1 (15.0) | 0.09 (0.03) |
| 3 (5.1–7.6 m) | 84 | 5.87 (0.78) | 2.74 (1.35) | 7.08 (1.74) | 1.83 (0.61) | 50.7 (16.2) | 0.13 (0.04) |
| All Classes | 252 | 3.53 | 1.59 | 4.86 | 1.26 | 40.94 | 0.09 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gray, J.M.; Schuler, J.L.; Thomas-Van Gundy, M.; Chhin, S. Reference Curves for Central Appalachian Red Spruce. Forests 2023, 14, 2260. https://doi.org/10.3390/f14112260
Gray JM, Schuler JL, Thomas-Van Gundy M, Chhin S. Reference Curves for Central Appalachian Red Spruce. Forests. 2023; 14(11):2260. https://doi.org/10.3390/f14112260
Chicago/Turabian StyleGray, Joseph M., Jamie L. Schuler, Melissa Thomas-Van Gundy, and Sophan Chhin. 2023. "Reference Curves for Central Appalachian Red Spruce" Forests 14, no. 11: 2260. https://doi.org/10.3390/f14112260
APA StyleGray, J. M., Schuler, J. L., Thomas-Van Gundy, M., & Chhin, S. (2023). Reference Curves for Central Appalachian Red Spruce. Forests, 14(11), 2260. https://doi.org/10.3390/f14112260