
Diversity and Structure of Soil Microbial Communities in Chinese Fir Plantations and Cunninghamia lanceolata–Phoebe bournei Mixed Forests at Different Successional Stages


Abstract
:1. Introduction
2. Material and Methods
2.1. Description of the Field Trial and Soil Sample Collection
2.2. Soil Properties and Soil Enzyme Activities Analysis
2.3. DNA Extraction, PCR Amplification, and Illumina MiSeq Sequencing of Soil Microorganisms
2.4. Statistical Analysis
3. Results
3.1. Soil Chemical Properties and Enzyme Activities
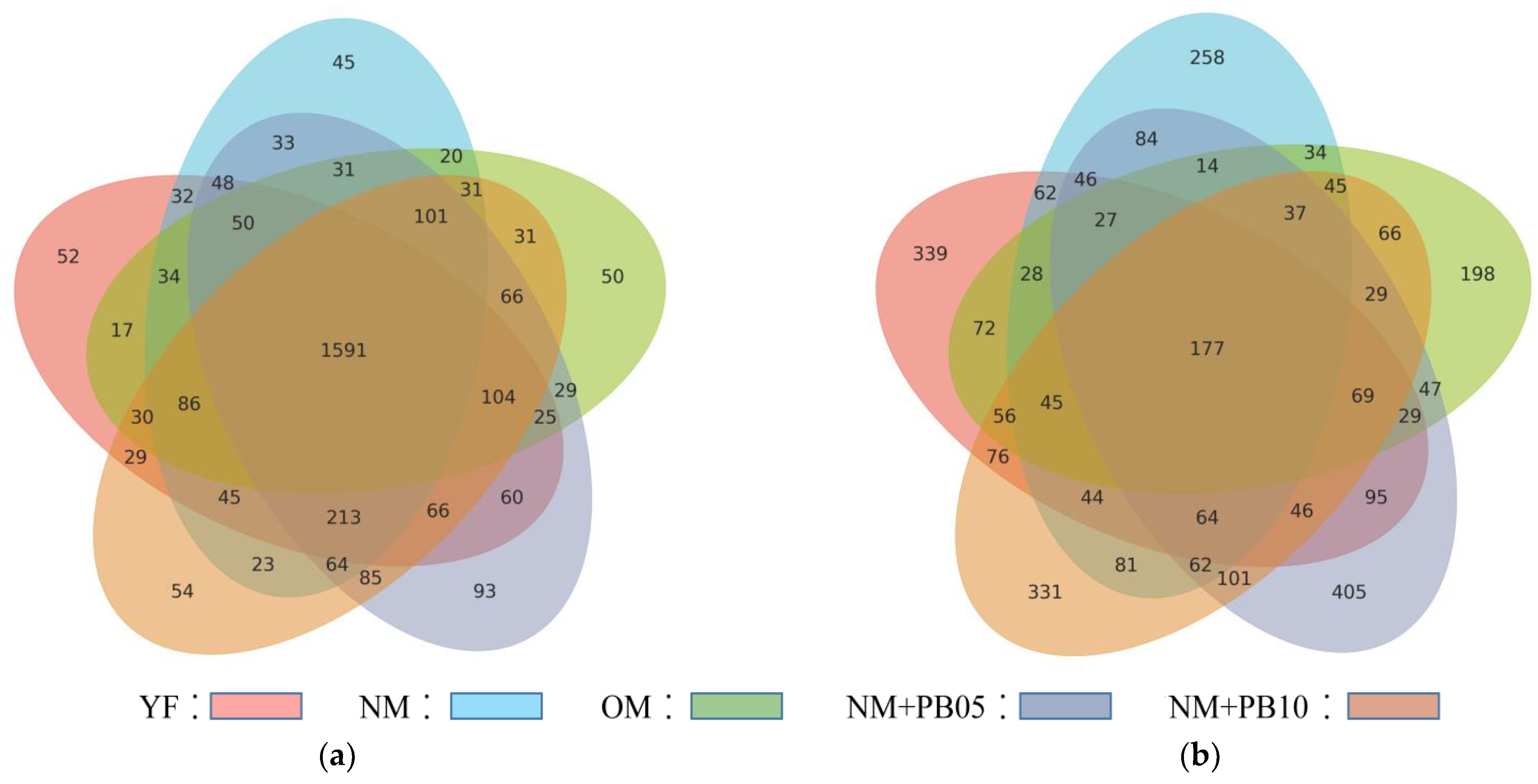
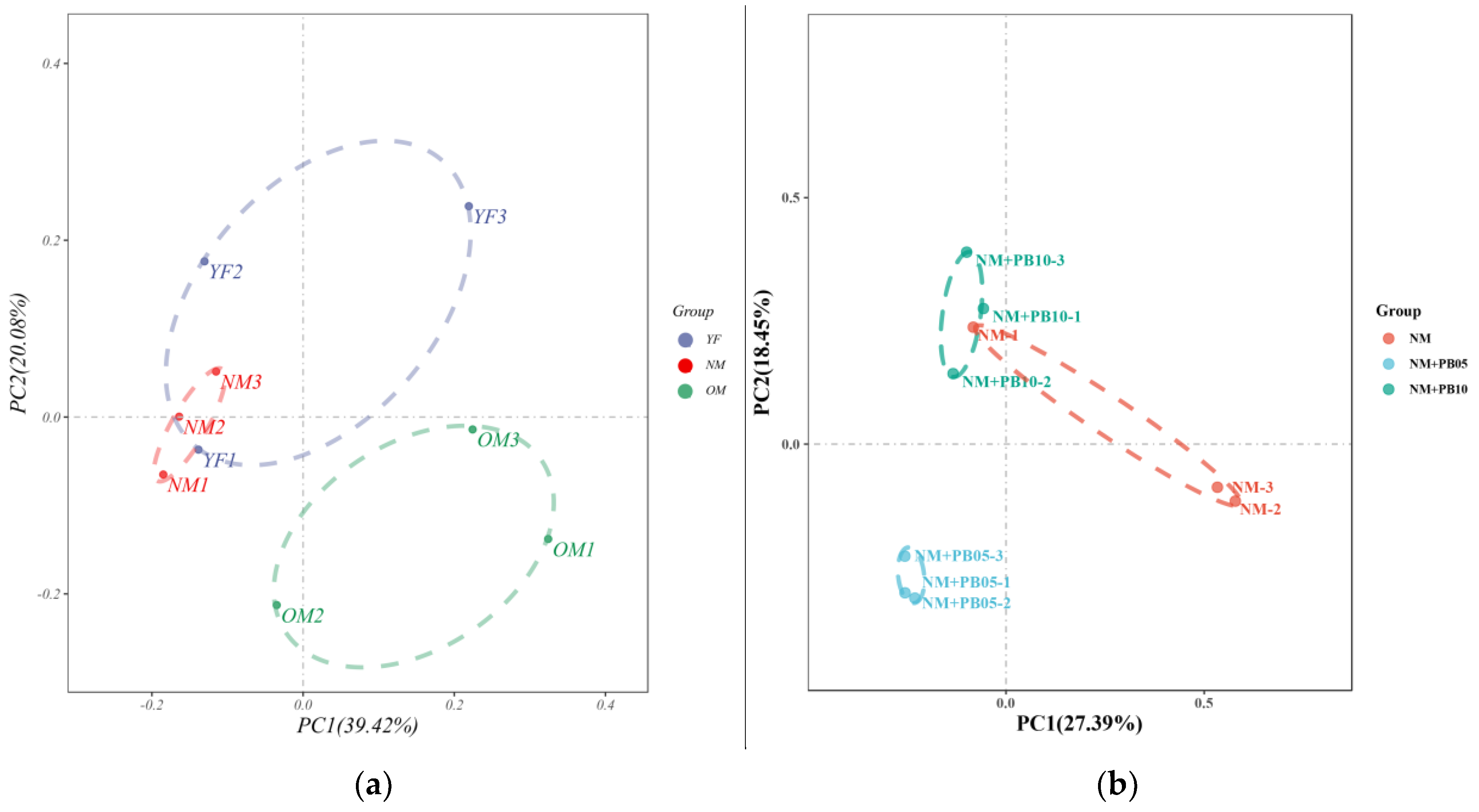
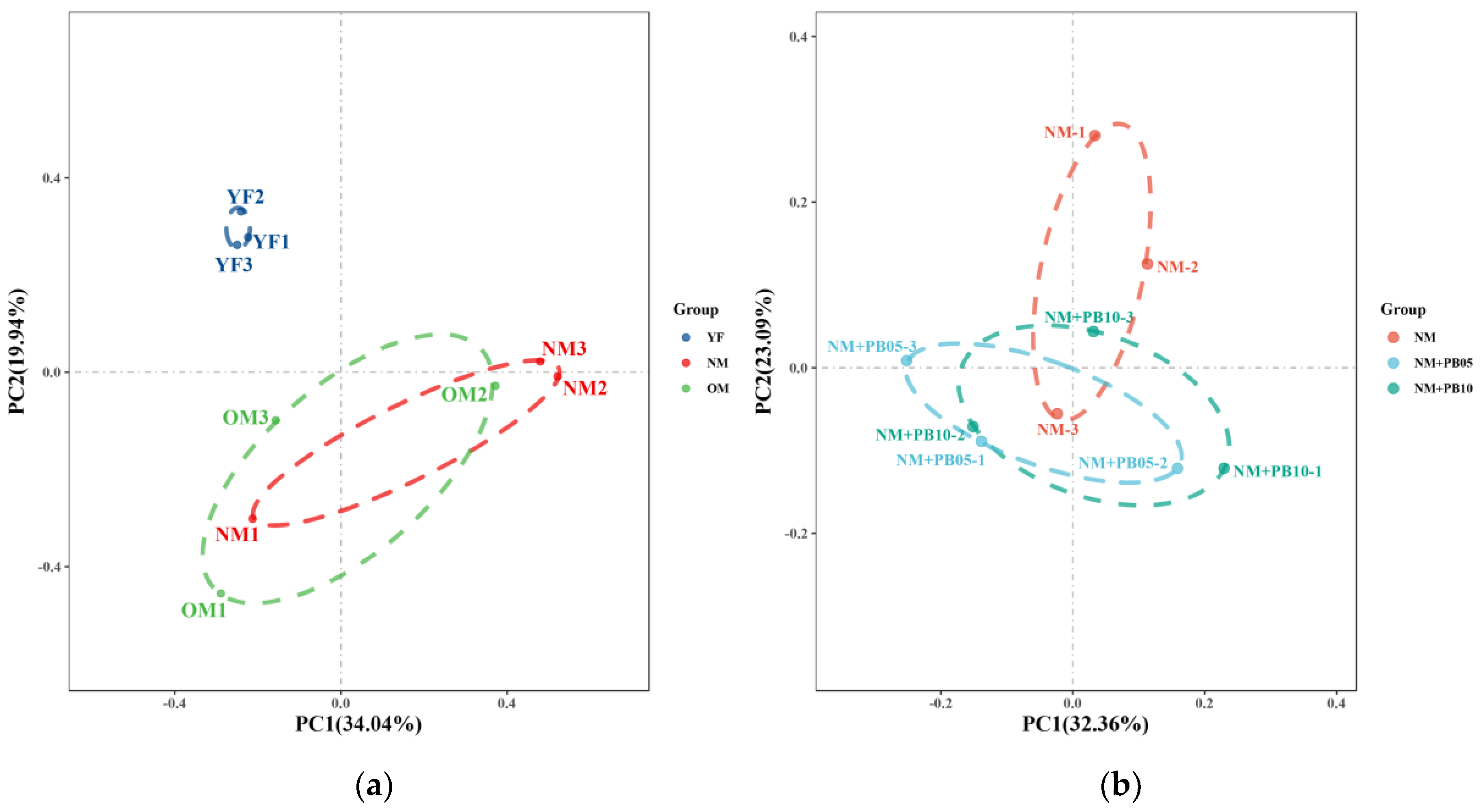
3.2. Characterization of Diversity of Soil Microbial Community
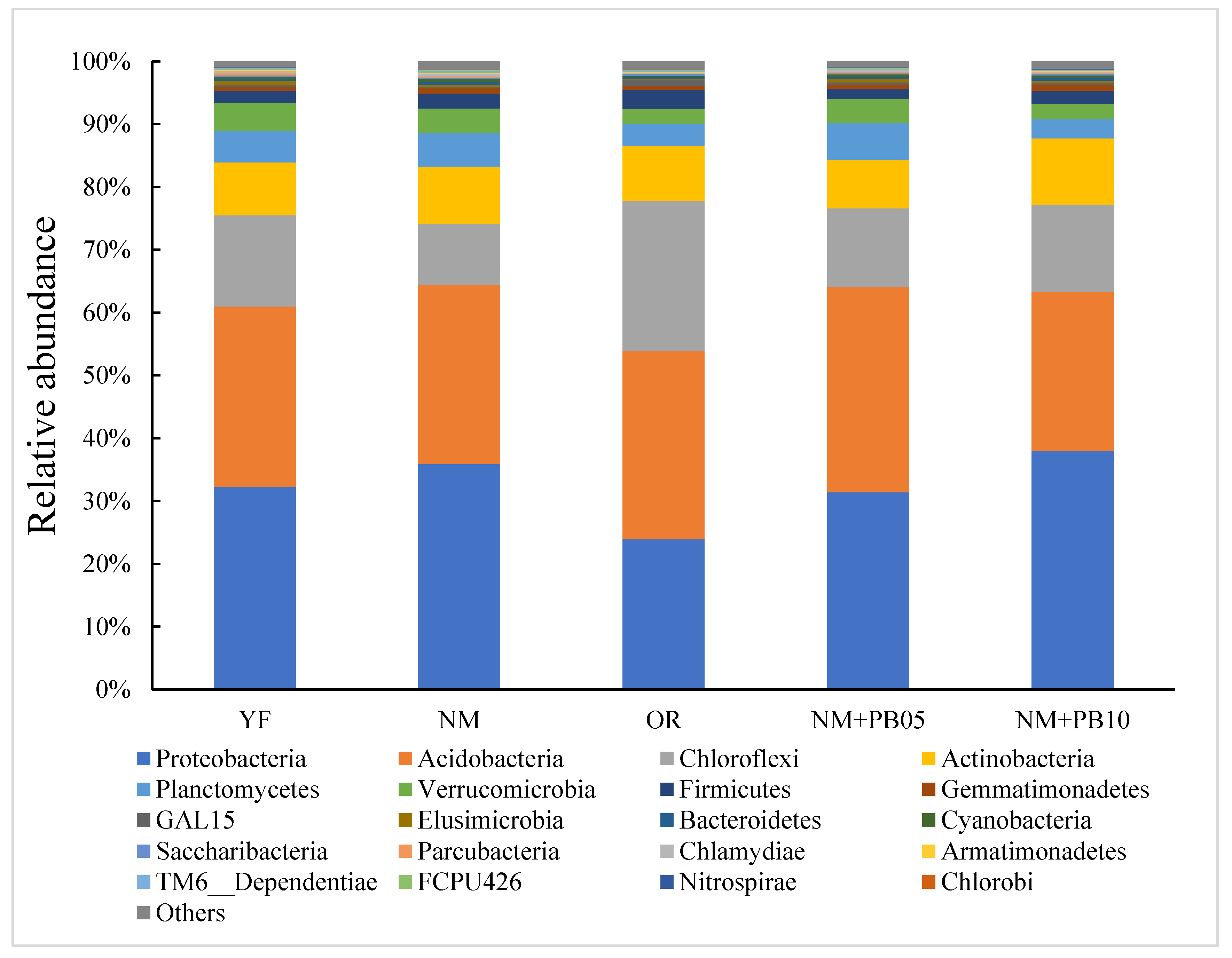
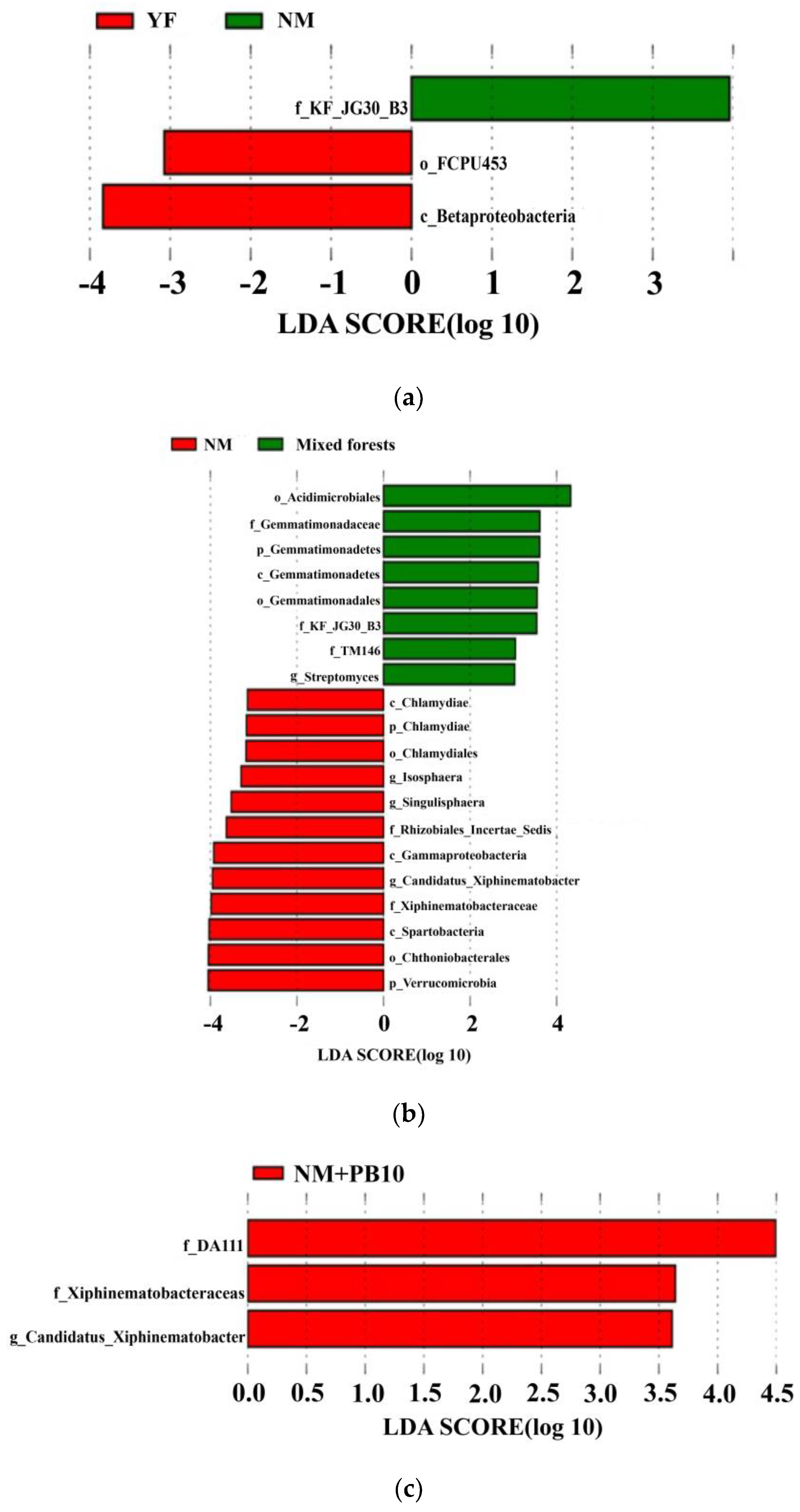
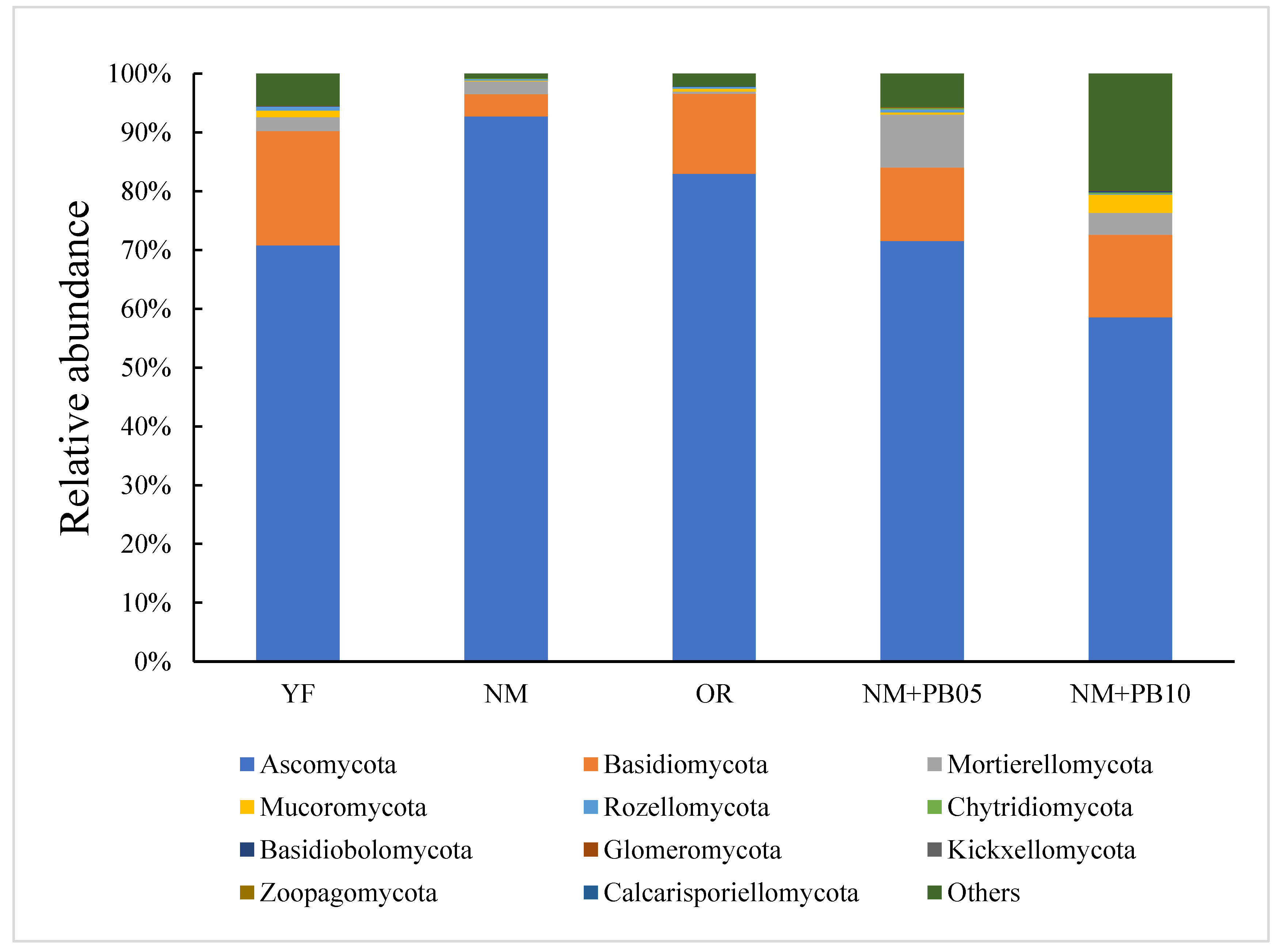
3.3. Differences in the Taxonomic Level Characteristics of Soil Microbial Communities
3.4. Structure of Microbial Community at OUT Level
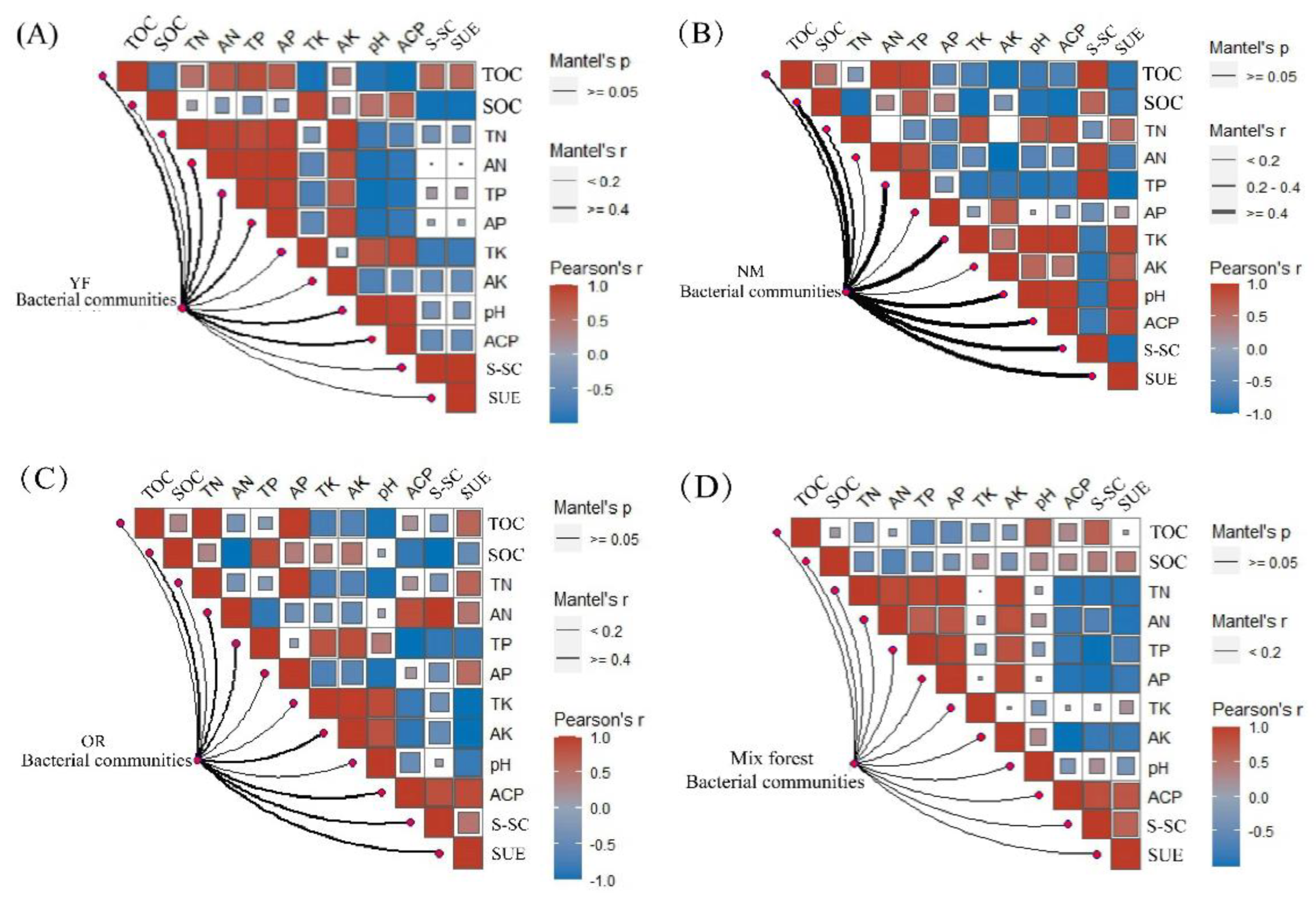
3.5. Identification of Key Drivers Affecting Microbial Community Structure
4. Discussion
4.1. Analysis of the Effects of Forest Age and Stand Type on Rhizosphere Soil Chemical Properties and Enzyme Activity
4.2. Rhizosphere Soil Microbial Diversity and Abundance Influenced by Tree Age and Stand Type
4.3. Effect of Stand Age and Stand Type on the Keystone Taxa
4.4. Different Stand Age and Forestry Management Practices Might Influence Soil Microbial Communities via Soil Properties
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Wang, Z.C.; He, G.X.; Hou, Z.H.; Luo, Z.; Chen, S.X.; Lu, J.; Zhao, J. Soil C:N:P stoichiometry of typical coniferous (Cunninghamia lanceolata) and/or evergreen broadleaved (Phoebe bournei) plantations in south China. For. Ecol. Manag. 2021, 486, 118974. [Google Scholar] [CrossRef]
- Bi, J.; Blanco, J.A.; Seely, B.; Kimmins, J.P.; Ding, Y.; Welham, C. Yield decline in Chinese-fir plantations: A simulation investigation with implications for model complexity. Can. J. For. Res. 2007, 9, 1615–1630. [Google Scholar] [CrossRef]
- Yu, Y.C.; Yang, J.Y.; Zeng, S.C.; Wu, D.M.; Douglass, F.J.; Sloan, L. Soil pH, organic matter, and nutrient content change with the continuous cropping of Cunninghamia lanceolata plantations in South China. J. Soils Sediments 2016, 9, 2230–2238. [Google Scholar] [CrossRef]
- Zhang, H.; Yu, J.D.; Li, H.Y.; Zhang, Y.L.; Pan, F.; Zhou, C.F.; Liu, A.Q. Characteristics of Soil Phosphorus in Cunninghamia lanceolata Plantations with Different Planting Rotations. For. Res. 2021, 1, 10–18. [Google Scholar]
- Kaarakka, L.; Cornett, M.; Domke, G.; Ontl, T.; Dee, L.E. Improved forest management as a natural climate solution: A review. Ecol. Solut. Evid. 2021, 2, e12090. [Google Scholar] [CrossRef]
- Ding, K.; Zhang, Y.; Wang, L.; Ge, S.; Zhang, Y.; Yang, Q.; Huang, H.H.; Tong, Z.K.; Zhang, J.H. Forest conversion from pure to mixed Cunninghamia lanceolata plantations enhances soil multifunctionality, stochastic processes, and stability of bacterial networks in subtropical southern China. Plant Soil. 2023, 488, 411–429. [Google Scholar] [CrossRef]
- Wang, Q.K.; Wang, S.L.; Huang, Y. Comparisons of litterfall, litter decomposition and nutrient return in a monoculture Cunninghamia lanceolata and a mixed stand in southern China. For. Ecol. Manag. 2008, 3–4, 1210–1218. [Google Scholar] [CrossRef]
- Zhao, J.Z.; Xie, D.M.; Wang, D.Y.; Deng, H.B. Current status and problems in certification of sustainable forest management in China. Env. Manag. 2011, 6, 1086–1094. [Google Scholar] [CrossRef]
- Wang, W.F.; Wei, X.H.; Liao, W.M.; Blanco, J.A.; Liu, Y.Q.; Liu, S.R.; Liu, G.H.; Zhang, L.; Guo, X.M.; Guo, S.M. Evaluation of the effects of forest management strategies on carbon sequestration in evergreen broad-leaved (Phoebe bournei) plantation forests using FORECAST ecosystem model. For. Ecol. Manag. 2013, 300, 21–32. [Google Scholar] [CrossRef]
- Khlifa, R.; Paquette, A.; Messier, C.; Reich, P.B.; Munson, A.D. Do temperate tree species diversity and identity influence soil microbial community function and composition? Ecol. Evol. 2017, 19, 7965–7974. [Google Scholar] [CrossRef]
- Wardle, D.A.; Bardgett, R.D.; Klironomos, J.N.; Setälä, H.; van-der Putten, W.H.; Wall, D.H. Ecological Linkages Between Aboveground and Belowground Biota. Science 2004, 5677, 1629–1633. [Google Scholar] [CrossRef] [PubMed]
- Shi, S.J.; Herman, D.J.; He, Z.J.; Pett-Ridge, J.; Wu, L.Y.; Zhou, J.Z.; Firestone, M.K. Plant roots alter microbial functional genes supporting root litter decomposition. Soil Biol. Biochem. 2018, 127, 90–99. [Google Scholar] [CrossRef]
- de Vries, F.T.; Griffiths, R.I.; Knight, C.G.; Nicolitch, O.; Williams, A. Harnessing rhizosphere microbiomes for drought-resilient crop production. Science 2020, 6488, 270–274. [Google Scholar] [CrossRef] [PubMed]
- Bastida, F.; Eldridge, D.J.; García, C.; Png, G.K.; Bardgett, R.D.; Delgado-Baquerizo, M. Soil microbial diversity–biomass relationships are driven by soil carbon content across global biomes. ISME J. 2021, 15, 2081–2091. [Google Scholar] [CrossRef]
- Hartmann, M.; Frey, B.; Mayer, J.; Mäde, P.; Widmer, F. Distinct soil microbial diversity under long-term organic and conventional farming. ISME J. 2015, 5, 1177–1194. [Google Scholar] [CrossRef]
- Tian, W.; Wang, L.; Li, Y.; Zhuang, K.M.; Li, G.; Zhang, J.B.; Xiao, X.J.; Xi, Y.G. Responses of microbial activity, abundance, and community in wheat soil after three years of heavy fertilization with manure-based compost and inorganic nitrogen. Agric. Ecosyst. Environ. 2015, 213, 219–227. [Google Scholar] [CrossRef]
- Zhang, P.; Guan, P.; Hao, C.; Yang, J.; Xie, Z.; Wu, D. Changes in assembly processes of soil microbial communities in forest-to-cropland conversion in Changbai Mountains, northeastern China. Sci. Total Environ. 2022, 818, 151738. [Google Scholar] [CrossRef]
- Meng, M.J.; Lin, J.; Guo, X.P.; Liu, X.; Wu, J.S.; Zhao, Y.P.; Zhang, J.C. Impacts of forest conversion on soil bacterial community composition and diversity in subtropical forests. CATENA 2019, 175, 167–173. [Google Scholar] [CrossRef]
- Dai, Z.M.; Xiong, X.Q.; Zhu, H.; Xu, H.J.; Li, X.H.; Tang, C.; Xu, J.M. Association of biochar properties with changes in soil bacterial, fungal and fauna communities and nutrient cycling processes. Biochar 2021, 3, 239–254. [Google Scholar] [CrossRef]
- Das, S.; Deb, S.; Sahoo, S.S.; Sahoo, U.K. Soil microbial biomass carbon stock and its relation with climatic and other environmental factors in forest ecosystems: A review. Acta Ecol. Sin. 2023, in press. [Google Scholar] [CrossRef]
- Xu, Z.Y.; Hu, Z.H.; Jiao, S.; Bell, S.M.; Xu, Q.; Ma, L.L.; Chen, J. Depth-dependent effects of tree species identity on soil microbial community characteristics and multifunctionality. Sci. Total Environ. 2023, 878, 162972. [Google Scholar] [CrossRef] [PubMed]
- Bach, L.H.; Grytnes, J.A.; Halvorsen, R.; Ohlson, M. Tree influence on soil microbial community structure. Soil Biol. Biochem. 2010, 11, 1934–1943. [Google Scholar] [CrossRef]
- Liu, Y.Y.; Dong, L.Z.; Zhang, H.J.; Deng, Y.Y.; Hu, B.; Wang, W. Distinct roles of bacteria and fungi in mediating soil extracellular enzymes under long-term nitrogen deposition in temperate plantations. For. Ecol. Manag. 2023, 529, 12068. [Google Scholar] [CrossRef]
- Baldrian, P.; Kolarik, M.; Stursova, M.; Kopecky, J.; Valaskova, V.; Vetrovsky, T.; Žifčáková, L.; Šnajdr, J.; Rídl, J.; Vlček, Č.; et al. Active and total microbial communities in forest soil are largely different and highly stratified during decomposition. ISME J. 2012, 2, 248–258. [Google Scholar] [CrossRef]
- Colombo, F.; Macdonald, C.A.; Jeffries, T.C.; Powell, J.R.; Singh, B.K. Impact of forest management practices on soil bacterial diversity and consequences for soil processes. Soil Biol. Biochem. 2016, 94, 200–210. [Google Scholar] [CrossRef]
- Kjøller, A.; Struwe, S. Microfungi in ecosystems: Fungal occurrence and activity in litter and soil. Oikos 1982, 39, 391–422. [Google Scholar] [CrossRef]
- Hayat, R.; Ali, S.; Amara, U.; Khalid, R.; Ahmed, I. Soil beneficial bacteria and their role in plant growth promotion: A review. Ann. Microbiol. 2010, 4, 579–598. [Google Scholar] [CrossRef]
- Zhou, L.; Sun, Y.J.; Saeed, S.; Zhang, B.; Luo, M. The difference of soil properties between pure and mixed Chinese fir (Cunninghamia lanceolata) plantations depends on tree species. Glob. Ecol. Conserv. 2020, 22, e01009. [Google Scholar] [CrossRef]
- Guo, J.H.; Feng, H.L.; Mcnie, P.; Liu, Q.Y.; Xu, X.; Pan, C.; Yan, K.; Feng, L.; Goitom, E.A.; Yu, Y.C. Species mixing improves soil properties and enzymatic activities in Chinese fir plantations: A meta-analysis. CATENA 2023, 220, 106723. [Google Scholar] [CrossRef]
- Ding, K.; Zhang, Y.; Liu, H.; Yang, X.; Zhang, J.; Tong, Z. Soil bacterial community structure and functions but not assembly processes are affected by the conversion from monospecific Cunninghamia lanceolata plantations to mixed plantations. Appl. Soil Ecol. 2023, 185, 104775. [Google Scholar] [CrossRef]
- Wang, Q.K.; Wang, S.L. Soil microbial properties and nutrients in pure and mixed Chinese fir plantations. J. For. Res. 2008, 2, 131–135. [Google Scholar] [CrossRef]
- Lei, J.; Wu, H.; Li, X.; Guo, W.; Duan, A.; Zhang, J. Response of rhizosphere bacterial communities to Near-Natural forest management and tree species within Chinese fir plantations. Microbiol. Spectr. 2023, 11, e02328-22. [Google Scholar] [CrossRef] [PubMed]
- Chu, H.Y.; Li, R.N.; Li, J.W.; Zhong, Z.Q.; Liu, X.F.; Li, Y.Q. Effects of forest conversion on soil microbial community structure. Chin. J. Appl. Environ. Biol. 2019, 1, 23–28. [Google Scholar]
- Li, J.; Cooper, J.M.; Lin, Z.A.; Li, Y.T.; Yang, X.D.; Zhao, B.Q. Soil microbial community structure and function are significantly affected by long-term organic and mineral fertilization regimes in the North China Plain. Appl. Soil Ecol. 2015, 96, 75–87. [Google Scholar] [CrossRef]
- Cui, Y.X.; Fang, L.C.; Deng, L.; Guo, X.B.; Han, F.; Ju, W.L.; Wang, X.; Chen, H.S.; Tan, W.F.; Zhang, X.C. Patterns of soil microbial nutrient limitations and their roles in the variation of soil organic carbon across a precipitation gradient in an arid and semi-arid region. Sci. Total Environ. 2019, 658, 1440–1451. [Google Scholar] [CrossRef] [PubMed]
- Sparks, D.L.; Helmke, P.A.; Page, A.L.; Loeppert, R.H.; Soltanpour, P.N.; Tabatabai, M.A.; Johnston, C.T.; Sumner, M.E. Methods of Soil Analysis: Part 3. Chemical Methods, 1st ed.; The Soil Science Society of America, American Society of Agronomy: Madison, WI, USA, 1996; pp. 65–90. [Google Scholar]
- Allen, S.E.; Grimshaw, H.M.; Parkinson, J.A.; Quarmby, C. Chemical Analysis of Ecological Materials, 1st ed; Institute Terrestrial Ecology, Merlewood Research Station: Cumbria, UK, 1977; pp. 494–495.
- Wu, J.; Joergensen, R.G.; Pommerening, B.; Chaussod, R.; Brookes, P.C. Measurement of soil microbial biomass C by fumigation-extraction—An automated procedure. Soil Biol. Biochem. 1990, 8, 1167–1169. [Google Scholar] [CrossRef]
- Zhou, L.K.; Zhang, Z.M. Determination method of soil enzyme activity. China J. Soil Sci. 1980, 5, 37–38. [Google Scholar]
- Zhang, J.J.; Kobert, K.; Flouri, T.; Stamatakis, A. PEAR: A fast and accurate Illumina Paired-End reAd mergeR. Bioinformatics 2014, 5, 614–620. [Google Scholar] [CrossRef]
- Ahn, J.; Sinha, R.; Pei, Z.H.; Dominianni, C.; Wu, J.; Shi, J.X.; Goedert, J.J.; Hayes, R.B.; Yang, L.Y. Human gut microbiome and risk for colorectalcancer. J. Natl. Cancer Inst. 2013, 24, 1907–1911. [Google Scholar] [CrossRef] [PubMed]
- Caporaso, J.G.; Kuczynski, J.; Stombaugh, J.; Bittinger, K.; Bushman, F.D.; Costello, E.K.; Fierer, N.; Peña, A.G.; Goodrich, J.K.; Gordon, J.I.; et al. QIIME allows analysis of high-throughput community sequencing data. Nat. Methods 2010, 5, 335–336. [Google Scholar] [CrossRef]
- Vasileiadis, S.; Puglisi, E.; Arena, M.; Cappa, F.; Cocconcelli, P.S.; Trevisan, M. Soil Bacterial Diversity Screening Using Single 16S rRNA Gene V Regions Coupled with Multi-Million Read Generating Sequencing Technologies. PLoS ONE 2012, 8, e42671. [Google Scholar]
- Heberle, H.; Meirelles, G.V.; da Silva, F.R.; Minghim, R. InteractiVenn: A web-based tool for the analysis of sets through Venn diagrams. BMC Bioinform. 2015, 1, 169. [Google Scholar] [CrossRef]
- Segata, N.; Izard, J.; Waldron, L.; Gevers, D.; Miropolsky, L.; Garrett, W.S.; Huttenhower, C. Metagenomic biomarker discovery and explanation. Genome Biol. 2011, 6, R60. [Google Scholar] [CrossRef]
- Deng, L.; Wang, K.B.; Chen, M.L.; Shangguan, Z.P.; Sweeney, S. Soil organic carbon stroage capacity positively related to forest succession on the loess plateau. CATENA 2013, 110, 1–7. [Google Scholar] [CrossRef]
- Wei, Z.C.; Huang, J.; Liu, Y.H.; Li, H.T.; Wu, P.F.; Liu, A.Q. Community characteristics of soil bacteria of Cunninghamia lanceolata plantations at different developmental stages. J. Southwest For. Univ. 2017, 5, 122–129. [Google Scholar]
- Chen, G.S.; Yang, Z.J.; Gao, R.; Xie, J.S.; Guo, J.F.; Huang, Z.Q.; Yang, Y.S. Carbon storage in a chronosequence of Chinese fir plantations in southern China. For. Ecol. Manag. 2013, 4, 68–76. [Google Scholar] [CrossRef]
- Liu, S.; Liu, X.S.; Zhu, X.Z.; Cheng, K.Y.; Guo, X.M.; Zhang, W.Y. Rhizosphere effects of nutrients and enzyme activities of Cunninghania lanceolata and soil fertility assessment. J. Plant Nutr. Fertil. 2017, 2, 492–501. [Google Scholar]
- Yang, Z.J.; Chen, S.D.; Liu, X.F.; Xiong, D.C.; Xu, C.; Arthur, M.A.; McCulley, R.L.; Yang, Y.S. Loss of soil organic carbon following natural forest conversion to Chinese fir plantation. For. Ecol. Manag. 2019, 449, 117476. [Google Scholar] [CrossRef]
- Wu, Z.Y.; Li, J.J.; Zheng, J.; Liu, J.F.; Liu, S.Y.; Lin, W.X.; Wu, C.Z. Soil microbial community structure and catabolic activity are significantly degenerated in successive rotations of Chinese fir plantations. Sci. Rep. 2017, 1, 6691. [Google Scholar] [CrossRef]
- Lan, S.A.; Du, H.; Zeng, F.P.; Song, T.Q.; Peng, W.X.; Han, C.; Chen, L.; Su, L. Carbon storage and allocation in Cunninghamia lanceolata plantations with different stand ages. China J. App. Ecol. 2016, 4, 1125–1134. [Google Scholar]
- Xu, L.; Shi, Y.J.; Fang, H.Y.; Zhou, G.M.; Xu, X.J.; Zhou, Y.F.; Tao, J.X.; Ji, B.Y.; Xu, J.; Li, C.; et al. Vegetation carbon stocks driven by canopy density and forest age in subtropical forest ecosystems. Sci. Total Environ. 2018, 631–632, 619–626. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Li, H.; Jiang, X.Y.; Dai, Q.S.; Zhang, H.C.; Song, H.L.; Zhang, J.Y. Soil factors influencing juvenile growth of Liriodendron chinense × tulipifera. J. Zhejiang Agric. For. Univ. 2016, 1, 94–101. [Google Scholar]
- Li, R.; Zhou, D.M.; Wu, Y.; Zhou, G.Y.; Huang, P.F.; Deng, X.J. Selection and characteristics of phosphate-solubilizing bacteria in rhizosphere of Cunninghaimia lanceolata. J. Cent. South Univ. 2012, 4, 95–99. [Google Scholar]
- Hao, Z.M.; Wu, S.R.; Qin, L.; Tan, L.; Guo, W.F. Soil physical and chemical properties of pure Pinus massoniana and its mixed forests in different ages in southern guangxi. Asian Agric. Res. 2018, 8, 53–57. [Google Scholar]
- Joshi, B.; Pant, S.C. Pedology of mixed deciduous and plantation forests in Tarai and Bhawar of Kumaun Himalaya, Uttarakhand. Indian For. 2014, 7, 679–684. [Google Scholar]
- Diakhate, S.; Gueye, M.; Chevallier, T.; Diallo, N.H.; Assigbetse, K.; Abadie, J.; Diouf, M.; Masse, D.; Sembène, M.; Ndour, Y.B.; et al. Soil microbial functional capacity and diversity in a millet-shrub intercropping system of semi-arid Senegal. J. Arid Environ. 2016, 129, 71–79. [Google Scholar] [CrossRef]
- Debenport, S.J.; Assigbetse, K.; Bayala, R.; Chapuis-Lardy, L.; Dick, R.P.; Gardener, B.B.M. Association of shifting populations in the root zone microbiome of millet with enhanced crop productivity in the Sahel Region (Africa). Appl. Environ. Microbiol. 2015, 8, 2841–2851. [Google Scholar] [CrossRef]
- Yarwood, S.A.; Hgberg, M.N. Soil bacteria and archaea change rapidly in the first century of Fennoscandian boreal forest development. Soil Biol Biochem. 2017, 114, 160–167. [Google Scholar] [CrossRef]
- Ren, C.J.; Liu, W.C.; Zhao, F.Z.; Zhong, Z.K.; Deng, J.; Han, X.H.; Yang, G.H.; Feng, Y.Z.; Ren, G.X. Soil bacterial and fungal diversity and compositions respond differently to forest development. CATENA 2019, 181, 104071. [Google Scholar] [CrossRef]
- Williams, M.A.; Jangid, K.; Shanmugam, S.G.; Whitman, W.B. Bacterial communities in soil mimic patterns of vegetative succession and ecosystem climax but are resilient to change between seasons. Soil Biol. Biochem. 2013, 57, 749–757. [Google Scholar] [CrossRef]
- Zhou, Z.H.; Wang, C.K.; Luo, Y.Q. Meta-analysis of the impacts of global change factors on soil microbial diversity and functionality. Nat. Commun. 2020, 1, 3072. [Google Scholar] [CrossRef] [PubMed]
- Li, Q. Effect of Neighboring Competition on Photosynthetic Characteristics and Biomass Allocation of Chinese Fir Seedlings under Low Phosphorus Stress. Master’s Thesis, Fujian Agriculture and Forestry University, Fuzhou, China, 2014. [Google Scholar]
- Zhuang, Z.; Li, Y.J.; Liu, Q.Q.; YU, Y.Y.; Liu, B.; Liu, A.Q. Sustained release of phosphorus by organic acid extraction from litter-derived red soil from Chinese fir plantation in southern China. J. Fujian Agric. For. Univ. 2017, 5, 569–575. [Google Scholar]
- Kang, H.Z.; Gao, H.H.; Yu, W.J.; Tang, Y.; Wang, Y.; Ning, M.L. Changes in soil microbial community structure and function after afforestation depend on species and age: Case study in a subtropical alluvial island. Sci. Total Environ. 2018, 625, 1423–1432. [Google Scholar] [CrossRef]
- Ogilvie, L.A.; Hirsch, P.R.; Johnston, A.W.B. Bacterial diversity of the broadbalk ’classical’ winter wheat experiment in relation to long-term fertilizer inputs. Microb. Ecol. 2008, 3, 525–537. [Google Scholar] [CrossRef] [PubMed]
- Kivlin, S.N.; Hawkes, C.V. Tree species, spatial heterogeneity, and seasonality drive soil fungal abundance, richness, and composition in neotropical rainforests. Environ. Microbiol. 2016, 12, 4662–4673. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.Z.; Chen, X.R.; Yang, C.D.; Xue, L. A study on the diversity of soil cultured fungi in the alpine grassland of eastern Qilian mountains. Acta Pratac. Sin. 2010, 2, 124–132. [Google Scholar]
- Eisenlord, S.D.; Zak, D.R. Simulated atmospheric nitrogen deposition alters actinobacterial community composition in forest soils. Soil Sci. Soc. Am. J. 2010, 4, 1157–1166. [Google Scholar] [CrossRef]
- Verzeaux, J.; Alahmad, A.; Habbib, H.; Nivelle, E.; Roger, D.; Lacoux, J.M.; Decocq, G.; Hirel, B.; Catterou, M.; Spicher, F.; et al. Cover crops prevent the deleterious effect of nitrogen fertilisation on bacterial diversity by maintaining the carbon content of ploughed soil. Geoderma 2016, 281, 49–57. [Google Scholar] [CrossRef]
- Debruyn, J.M.; Nixon, L.T.; Fawaz, M.N.; Johnson, A.M.; Radosevich, M. Global biogeography and quantitative seasonal dynamics of Gemmatimonadetes in Soil. Appl. Environ. Microbiol. 2011, 17, 6295–6300. [Google Scholar] [CrossRef]
- Su, J.Q.; Wei, B.; Ou-Yang, W.Y.; Huang, F.Y.; Zhao, Y.; Xu, H.J.; Zhu, Y.G. Antibiotic resistome and its association with bacterial communities during sewage sludge composting. Environ. Sci. Technol. 2015, 12, 7356–7363. [Google Scholar] [CrossRef]
- Yelle, D.J.; Ralph, J.; Lu, F.; Hammel, K.E. Evidence for cleavage of lignin by a brown rot basidiomycete. Environ. Microbiol. 2008, 7, 1844–1849. [Google Scholar] [CrossRef] [PubMed]
- Liang, Y.; Guo, L.D.; Mark, P. Role of mycorrhizal fungi in ecosystems. Chin. J. Plant Ecol. 2002, 6, 739–745. [Google Scholar]
- Nielsen, U.N.; Osler, G.H.R.; Campbell, C.D.; Burslem, D.F.R.P.; van der Wal, R. The influence of vegetation type, soil properties and precipitation on the composition of soil mite and microbial communities at the landscape scale. J. Biogeogr. 2010, 7, 1317–1328. [Google Scholar] [CrossRef]
- Lauber, C.L.; Hamady, M.; Knight, R.; Fierer, N. Pyrosequencing-Based assessment of soil pH as a predictor of soil bacterial community structure at the continental scale. Appl. Environ. Microbiol. 2009, 15, 5111–5120. [Google Scholar] [CrossRef] [PubMed]
- Griffiths, R.I.; Thomson, B.C.; James, P.; Bell, T.; Whiteley, A.S. The bacterial biogeography of British soils RID D-5237-2011 RID A-3395-2012. Environ. Microbiol. 2011, 6, 1642–1654. [Google Scholar] [CrossRef]
- Li, L.; Li, S.M.; Sun, J.H.; Zhou, L.L.; Bao, X.G.; Zhang, H.G.; Zhang, F.S. Diversity enhances agricultural productivity via rhizosphere phosphorus facilitation on phosphorus-deficient soils. Proc. Natl. Acad. Sci. USA 2007, 27, 11192–11193. [Google Scholar] [CrossRef]
- Liu, W.X.; Jiang, L.; Yang, S.; Wang, Z.; Tian, R.; Peng, Z.Y.; Chen, Y.L.; Zhang, X.X.; Kuang, J.L.; Ling, N.; et al. Critical transition of soil bacterial diversity and composition triggered by nitrogen enrichment. Ecology 2020, 101, e03053. [Google Scholar] [CrossRef]
- Maisto, G.; Marco, A.D.; Meola, A.; Sessa, L.; Santo, A.V.D. Nutrient dynamics in litter mixtures of four Mediterranean maquis species decomposing in situ. Soil Biol. Biochem. 2011, 3, 520–530. [Google Scholar] [CrossRef]
- Chen, Y.L.; Kang, L.H.; Malajczuk, N.; Dell, B. Selecting ectomycorrhizal fungi for inoculating plantations in south China: Effect of scleroderma on colonization and growth of exotic Eucalyptus globulus, E. urophylla, Pinus elliottii, and P. radiata. Mycorrhiza 2006, 4, 251–259. [Google Scholar] [CrossRef]
- Wang, J.Q.; Shi, X.Z.; Zheng, C.Z.; Suter, H.; Huang, Z. Different responses of soil bacterial and fungal communities to nitrogen deposition in a subtropical forest. Sci. Total Environ. 2020, 755, 142449. [Google Scholar] [CrossRef]
- Zhao, Y.G.; Zhang, F.H.; Yang, L.; Wang, D.; Wang, W.C. Response of soil bacterial community structure to different reclamation years of abandoned salinized farmland in arid China. Arch. Microbiol. 2019, 9, 1219–1232. [Google Scholar] [CrossRef] [PubMed]
- Fierer, N.; Jackson, R.B. The diversity and biogeography of soil bacterial communities. Proc. Natl. Acad. Sci. USA 2006, 3, 626–631. [Google Scholar] [CrossRef] [PubMed]
- Dennis, P.G.; Rushton, S.P.; Newsham, K.K.; Lauducina, V.A.; Ord, V.J.; Daniell, T.J.; O’Donnell, A.G.; Hopkins, D.W. Soil fungal community composition does not alter along a latitudinal gradient through the maritime and sub-Antarctic. Fungal Ecol. 2012, 4, 403–408. [Google Scholar] [CrossRef]
- Lin, K.M.; Zhang, Z.Q.; Cao, G.Q.; He, Z.M.; Ma, X.Q. Decomposition characteristics and its nutrient dynamics of leaf litter mixtures of both Chinese fir and Phoeba bournei. Acta Ecol. Sin. 2006, 8, 2732–2738. [Google Scholar]
- Xia, Z.C.; Yu, L.; He, Y.; Korpelainen, H.; Li, C.Y. Broadleaf trees mediate chemically the growth of Chinese fir through root exudates. Biol. Fertil. Soils 2019, 7, 737–749. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| TOC (g/kg) | SOC (mg/g) | TN (g/kg) | AN (mg/g) | TP (g/kg) | AP (mg/g) | TK (g/kg) | AK (mg/g) | pH | ACP (mg·g−1·h−1) | S-SC (mg·g−1·h−1) | SUE (mg·g−1·h−1) | |
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| YF | 9.06 ± 0.79 a | 0.94 ± 0.51 a | 1.29 ± 0.10 bc | 60.76 ± 8.08 bc | 0.22 ± 0.56 b | 0.667 ± 0.09 b | 7.47 ± 0.50 a | 22.69 ± 4.10 a | 4.67 ± 0.20 c | 0.46 ± 0.01 a | 3.27 ± 0.52 a | 28.38 ± 8.50 b |
| NM | 14 ± 0.72 b | 1.14 ± 0.87 ab | 0.68 ± 0.03 a | 33.32 ± 5.27 a | 0.07 ± 0.00 a | 0.37 ± 0.10 b | 12.68 ± 0.25 b | 33.41 ± 3.37 b | 4.19 ± 0.71 b | 0.52 ± 0.02 ab | 4.72 ± 0.80 a | 58.11 ± 9.60 d |
| OR | 14.7 ± 0.45 bc | 1.21 ± 0.05 ab | 1.13 ± 0.15 b | 56.04 ± 6.99 b | 0.264 ± 0.34 b | 0.56 ± 0.05 ab | 15.5 ± 0.95 c | 36.62 ± 6.70 b | 3.85 ± 0.07 a | 0.50 ± 0.01 a | 5.10 ± 0.37 a | 41.63 ± 0.09 c |
| NM + PB05 | 14.7 ± 1.4 bc | 1.18 ± 0.29 ab | 0.75 ± 0.15 a | 72.38 ± 10.72 c | 0.3719 ± 0.00 c | 4.04 ± 0.13 d | 14.9 ± 1.45 c | 49.32 ± 0.51 c | 4.62 ± 0.08 c | 0.70 ± 0.11 c | 11.81 ± 3.53 b | 13.18 ± 1.65 a |
| NM + PB10 | 16.1 ± 1.3 c | 1.34 ± 0.76 b | 1.35 ± 0.05 c | 51.36 ± 10.59 b | 0.226 ± 0.23 b | 2.73 ± 0.20 c | 15.13 ± 0.86 c | 34.23 ± 5.89 b | 4.63 ± 0.13 c | 0.56 ± 0.10 bc | 14.14 ± 1.23 b | 16.73 ± 1.79 a |
| Treatment | OTU | ACE | Chao1 | Shannon | Simpson |
|---|---|---|---|---|---|
| YF | 1671.67 ± 89.07 b | 1988.67 ± 60.43 c | 2010.67 ± 71.93 c | 5.81 ± 0.23 a | 0.88 ± 0.40 a |
| NM | 1944.67 ± 131.80 a | 2319.67 ± 111.79 a | 2342.00 ± 112.93 a | 6.02 ± 0.12 a | 0.64 ± 0.02 a |
| OR | 1761.00 ± 69.46 ab | 2071.00 ± 125.41 bc | 2101.67 ± 155.00 bc | 6.02 ± 0.17 a | 0.62 ± 0.13 a |
| NM + PB05 | 1831.00 ± 100.54 ab | 2172.33 ± 155.99 abc | 2210.33 ± 144.67 abc | 5.95 ± 0.17 a | 0.67 ± 0.17 a |
| NM + PB10 | 1889.33 ± 165.24 ab | 2279.67 ± 140.63 ab | 2311.00 ± 93.21 ab | 5.97 ± 0.15 a | 0.67 ± 0.08 a |
| Treatment | Chao1 | Observed Species | PD Whole Tree | Shannon | Simpson |
|---|---|---|---|---|---|
| YF | 779.93 ± 93.18 ab | 676.00 ± 96.02 a | 173.13 ± 20.75 a | 6.40 ± 0.60 a | 0.97 ± 0.02 a |
| NM | 681.17 ± 80.70 ab | 549.00 ± 131.53 a | 157.88 ± 23.00 a | 3.34 ± 1.95 b | 0.57 ± 0.33 b |
| OR | 584.43 ± 217.54 b | 487.33 ± 202.21 a | 145.97 ± 51.81 a | 3.93 ± 2.32 ab | 0.67 ± 0.33 ab |
| NM + PB05 | 768.49 ± 33.36 ab | 697.67 ± 47.60 a | 193.93 ± 8.98 a | 6.23 ± 0.71 a | 0.94 ± 0.05 ab |
| NM + PB10 | 826.32 ± 130.59 a | 687.00 ± 150.12 a | 189.86 ± 29.42 a | 5.10 ± 1.31 ab | 0.87 ± 0.13 ab |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Li, W.; Sun, H.; Cao, M.; Wang, L.; Fang, X.; Jiang, J. Diversity and Structure of Soil Microbial Communities in Chinese Fir Plantations and Cunninghamia lanceolata–Phoebe bournei Mixed Forests at Different Successional Stages. Forests 2023, 14, 1977. https://doi.org/10.3390/f14101977
Li W, Sun H, Cao M, Wang L, Fang X, Jiang J. Diversity and Structure of Soil Microbial Communities in Chinese Fir Plantations and Cunninghamia lanceolata–Phoebe bournei Mixed Forests at Different Successional Stages. Forests. 2023; 14(10):1977. https://doi.org/10.3390/f14101977
Chicago/Turabian StyleLi, Weiyang, Huimin Sun, Minmin Cao, Liyan Wang, Xianghua Fang, and Jiang Jiang. 2023. "Diversity and Structure of Soil Microbial Communities in Chinese Fir Plantations and Cunninghamia lanceolata–Phoebe bournei Mixed Forests at Different Successional Stages" Forests 14, no. 10: 1977. https://doi.org/10.3390/f14101977
APA StyleLi, W., Sun, H., Cao, M., Wang, L., Fang, X., & Jiang, J. (2023). Diversity and Structure of Soil Microbial Communities in Chinese Fir Plantations and Cunninghamia lanceolata–Phoebe bournei Mixed Forests at Different Successional Stages. Forests, 14(10), 1977. https://doi.org/10.3390/f14101977