Abstract
Changes in the microenvironment induced by forest gaps may affect litter decomposition, yet it is unclear how the gap effects respond to altitudinal and seasonal differences. Here, a four-year litterbag decomposition experiment along an elevation gradient (3000, 3300, 3600 m) was conducted in an Abies faxoniana Rehd. subalpine forest of southwestern China, to assess the potential seasonal effects of forest gaps (large: ≈250 m2, middle: ≈125 m2, small: ≈40 m2 vs. closed canopy) on litter mass loss and carbon release at different elevations. We found that the A. faxoniana litter mass loss and carbon release reached 50~53 and 58~64% after four years of decomposition, respectively. Non-growing seasons (November to April) had a greater decline than the growing seasons (May to October). Litter in the forest gaps exhibited significantly higher mass loss than that under the closed canopy, and the decomposition constant (k) exhibited a gradually declining trend from large gaps, middle gaps, small gaps to closed canopy. Moreover, more significant differences of gap on both carbon content and release were observed at the 3600 m site than the other two elevations. Our findings indicate that (i) a rather high mass loss and carbon release during the decomposition of A. faxoniana litter was observed at high elevations of the subalpine forest subjected to low temperatures in the non-growing seasons and (ii) there were stimulative effects of forest gaps on litter mass loss and carbon release in early decomposition, especially in the non-growing seasons, driven by fewer freeze–thaw cycles when compared to the closed canopy, which diminished at the end of the experiment. The results will provide crucial ecological data for further understanding how opening gaps as a main regeneration method would induce changes in carbon cycling in subalpine forest ecosystems.
1. Introduction
Litter decomposition is a crucial component of carbon and nutrient turnover, and determines the carbon balance in forest ecosystems [1,2]. In the past decades, research into this complex ecological process has grown steadily since the development of the litterbag technique [3,4], which reveals that ambient temperature and moisture influence litter decomposition rate in the early stage [5]. Forest gaps are a main natural regeneration method, caused by pests, wildfires, natural stem breakage, human deforestation and other disturbances, and are widely distributed in forest ecosystems [6], which may induce changes in the microenvironment such as: temperature, precipitation, sunlight exposure, snow coverage and further affecting litter mass loss and carbon release during the decomposition processes [7]. Previous studies suggested forest gaps affected decomposition rates in a different way, which were documented to be greater, weaker or the same compared to that under closed canopies [8,9,10]. Furthermore, litter decomposition rates and nutrient release were reported to be stimulated, inhibited and insignificantly related to increasing gap sizes across multiple forests ecosystems [11,12,13]. Such uncertainties may limit our understanding of how gaps affect decomposition-induced carbon cycling in forest ecosystems.
The altitudinal gradient across a relatively smaller spatial scale could mirror the large-scale environmental conditions along climatic gradients, leading to changes in soil microbial biomass, enzyme activity and other driving factors for decomposition [14,15,16,17]. In general, the atmospheric and soil temperatures gradually decrease with increasing elevation, accompanied by deeper seasonal snowpack as well as a longer coverage period in higher elevations [18,19]. However, there were divergent results of the variations in soil biological indicators with an increasing elevation, such as promoted or inhibited enzyme activities and soil microbial quotient [20,21,22], which may directly affect litter decomposition, implying that the decomposition pattern with increasing elevation is still uncertain. Theoretically, getting away from the canopy interception would help the floor in forest gap obtain more precipitation and sunlight [7,23,24], further favoring the abundant biological community [25,26]. Therefore, the effects of forest gap size on litter decomposition along an elevation gradient need to be further assessed to strengthen our uncertain knowledge of this critical carbon-cycling ecological process.
Litter decomposition has obvious seasonal dynamics, which are more remarkable in high-elevation regions with distinct growing and non-growing seasons driven mainly by soil temperature and moisture [27]. Consequently, the strength of forest gap effects on litter mass loss and carbon release in response to an increasing elevation might be highly season-dependent. The non-growing season tends to be a critical period for the decomposition of newly shed litter [28,29]. Snowpack accumulated in forest gaps offers insulated protection against the outer extremely low temperature and frequent freeze–thaw cycles to maintain preferable microenvironmental conditions for decomposers [30], and this induces intensive leaching losses attributed to the strong snowmelt in early spring. Both functions would promote litter decomposition in winter [31,32]. Moreover, enhanced photodegradation and eluviation from sunlight exposure and heavy rain wash would also stimulate the litter decomposition within gaps when compared to that under closed canopy [33,34]. In subalpine regions, relevant information is scarce, especially when discussed in conjunction with different elevation effects.
Subalpine forests in the eastern Qinghai–Tibetan Plateau contain shallow soil thus making litter decomposition a crucial role in ecological processes, such as maintaining carbon balance in the area [35]. In elevations from 3000 to 3700 m, Abies faxoniana Rehd. is the dominant tree species and 84% of forest gaps are less than 240 m2 in these forests according to our previous investigation [36]. A four-year litterbag decomposition experiment was conducted in an A. faxoniana subalpine forest of southwestern China, to assess the potential effects of forest gaps (large: ≈250 m2, middle: ≈125 m2, small: ≈40 m2 vs. closed canopy) on litter mass loss and carbon release in decomposing A. faxoniana litter. More specifically, we evaluated the responses of decomposition for three elevations (3000, 3300, 3600 m) to forest gaps at different critical stages for four years. We hypothesized that (i) the A. faxoniana leaf litter mass loss and carbon release were lower under closed canopies than in forest gaps; (ii) the gap effects on the mass loss and carbon release would vary with gap size and elevation, and diminish gradually along with the decomposition; (iii) temperature was the key driver of the gap and elevation induced changes in litter decomposition. The objective of this study was to understand the seasonal effects of forest gap on litter decomposition along elevational differences, which could provide crucial ecological data for further understanding how the opening of gaps would induce changes in carbon cycling in subalpine forest ecosystems.
2. Materials and Methods
2.1. Site Description and Experimental Design
The study was conducted at the Long-term Research Station of Alpine Forest Ecosystems (31°14′–31°19′ N, 102°53′–102°57′ E, 2458~4619 m a.s.l.), located in Li County, southwestern China. It is a transitional area between the Tibetan Plateau and Sichuan Basin with remarkable changes in climate, vegetation and soil types along the elevation gradient [37,38]. The mean annual temperature and precipitation are 2~4 °C and 850 mm, respectively [39]. Winter generally starts from late October to the next April, with seasonal snowpack accumulated on the forest floor of up to approximately 50 cm and frequent soil freeze–thaw cycles [9]. Abies faxoniana Rehd., Betula albo-sinensis and Sabina saltuaria are the dominant trees species, and Salix paraplesia, Rhododendron Lapponicum and Fargesia spathacea are the dominant shrubs. The forest soil is classified as Cambisols [40]. The concentrations of total C, N, and P in soil (0~20 cm) were 363, 6.6 and 1.6 g·kg−1, respectively [33].
Based on the previous investigation, forest gaps caused by human deforestation and natural stem breakage were widely distributed in the study area [36]. In September 2011, we selected A. faxoniana forests with similar slopes (24°~34°) and aspects (NE 38°~45°) at 3000 m (31°19′ N, 102°56′ E), 3300 m (31°17′ N, 102°56′ E) and 3600 m (31°15′ N, 102°53′ E)as our three study sites. For the 3000 m site, the dominant species are Betula albosinensi, A. faxoniana, Berberis diaphana, Fragaria orientalis, Thalictrum aquilegifolium, Polygonum viviparum etc., the annual precipitation and temperature are 825 mm and 3.6 °C, the forest soil is classified as Cambisols, and the organic layer is 15 ± 2 cm. For the 3300 m site, the dominant species are A. faxoniana, Betula albosinensi, Sorbus, Hippophae rhamnoides, Parasenecio forrestii, Fragaria orientalis etc. the annual precipitation and temperature are 825 mm and 2.8 °C, the forest soil is classified as Cambisols, and the organic layer is 12 ± 2 cm. For the 3600 m site, the dominant species are A. faxoniana, Prunus tatsienensis, Rhododendron lapponicum, Salix paraplesia, Fargesia spathacea, Fargesia spathacea etc., the annual precipitation and temperature are 825 mm and 1.6 °C, the forest soil is classified as Cambisols, and the organic layer is 12 ± 2 cm. At each site, three plots of each gap size were established: three gap sizes were large gaps (≈250 m2), middle gaps (≈125 m2) and small gaps (≈40 m2), with 9 gap plots in total; another three plots (10 m×10 m) under the ambient closed canopy were set up as the control. These forests gaps have all existed for about 25 years, and all the 36 plots were located apart with over 50 m apart from each other. Other detailed information is shown in Table S1.
2.2. Litterbag Experiment
Foliar litter of A. faxoniana in this study area was collected using a litter trap method in late autumn (October 2011). Only freshly fallen leaves (without signs of galls, herbivory or atypical coloration) were collected and air-dried for two weeks. Nylon litterbags (20 cm × 20 cm, mesh size: 1 mm top vs. 0.5 mm bottom) were used to fill 10 g of dried litter. Litterbags were placed on the top of existing litter layer in each plot on November 21, 2011. In total, we applied 3 elevations × 4 gap sizes × 3 replicates × 3 bags × 16 samplings = 1728 litterbags. The snow depth in each plot was obtained by averaging the direct measurements from 9 random locations, and the measurements were performed immediately after each sampling event using a ruler. The temperatures of the litter layers were recorded every 2 h by iButton DS1923-F5 recorders (Maxim Inc., Sunnyvale, CA, USA), which were placed in the litterbags.
Based on previous field investigations, the growing season was from April to October every year [31]. In order to explore litter decay processes comprehensively, the bags were retrieved at the end of each period: these were in December (OF, onset of freezing stage), February (DP, deep freezing stage), April (TS, thawing stage), June (EGS, early snow-free stage), August (MGS, middle snow-free stage) and October (LGS, late snow-free stage) of the initial two decomposing years (2011–2013). For the latter two years (2014–2015), bags were only retrieved in April (NGS, non-growing season) and October (GS, growing season), which were defined according to the phenological characteristics of the vegetation in the study area. After carefully removing the roots, mosses and foreign materials from the litter, each sample was oven-dried at 65 °C for 48 h to a constant weight to determine the litter’s remaining mass. Litter carbon content was determined using the dichromate oxidation–ferrous sulfate titration method [41].
2.3. Data Analysis
The mean temperature and frequency of the freeze–thaw cycles of each forest gap were calculated in each decomposition stage using the every 2 h temperature data in situ. Freezing and thawing in the non-growing season are important physical processes that impact below-ground ecological functions in alpine forests. We calculated the freeze–thaw cycles per day during each decomposition period; one freeze–thaw cycle was defined as the temperature increasing above 0 °C or decreasing below 0 °C for 3 h or more, followed by a decrease below 0 °C or an increase above 0 °C for at least 3 h [42].
The litter mass loss (ML, %) and carbon release (CR, %) were calculated as follows:
where M0 (g) and Mt (g) are the litter’s remaining mass in the litterbags at the initial and sampling times, respectively; C0 (g·kg−1), Ct (g·kg−1) Ct-1 (g·kg−1) are the litter carbon content at the initial, sampling and previous sampling times, respectively. All analyses were conducted in triplicate.
MLt = (M0 − Mt)/M0 × 100%
CRt = (Ct-1 × Mt-1 − Ct × Mt)/(C0 × M0) × 100%
The Olson exponential decay model [43] was used to fit the relationship of litter remaining mass and decomposition year:
where M0 is the litter initial dry mass; e is a natural constant; k is the litter decomposition constant; t is the decomposition year; Mt is the litter’s remaining mass at t.
Mt = M0·e−kt
In this study, gap size (with three sizes and closed canopy) was a main-plot factor, elevation (with three elevations) was a sub-plot factor, and decomposition time (with sixteen stages) was a sub-subplot factor.
A two-way ANOVA (with the LSD post hoc test) was used to examine the differences of litter decomposition constant (k) generated from the Olson exponential decay model, mass loss, carbon content and release both among the gap sizes and elevations. Linear mixed models were conducted to analyze the effects of gap size, elevation, decomposition time and their interaction on litter mass loss, carbon content and release; the bags within gap sizes were set as random effects. All data were subjected to a normality test and homogeneity of variance test prior to the analysis of variance. Multiple non-linear regressions were conducted to evaluate the links of the litter’s remaining mass and carbon content from the scales of gap size and elevation. Spearman correlation analysis was performed to describe the relationship between mean temperature, freeze–thaw cycles and litter mass loss, carbon release in the non-growing season, growing season and the whole four decomposition years. All statistical analyses were performed using SPSS 27.0 (SPSS Inc., Chicago, IL, USA) and Prism GraphPad 8.0 (GraphPad Software Inc. San Diego, CA, USA).
3. Results
3.1. Temperature Characteristics and Snow Depth
Temperature characteristics of litter layer fluctuated consistently in different forest gaps and elevations are shown in Table 1. For the non-growing and growing seasons, a higher mean temperature was observed in forest gaps when compared to that under the closed canopy in all the three elevations. Furthermore, the highest mean temperatures were shown in the large gap in the 3000 m, while it was found in the middle or small gaps in the 3300 m and 3600 m plots. Freeze–thaw cycles’ frequency was lower in large and middle gaps at the 3600 m and 3300 m sites, while a higher cycle frequency was found at the 3000 m site. During the non-growing season (November to April), the snow depth on the forest soil was consistently ranked in the following two orders: large gap > middle gap > small gap > closed canopy, 3600 m > 3300 m > 3000 m. At the OF and DF of the first two decomposing years, the accumulated snow was deeper than for other stages.

Table 1.
Seasonal environmental factors of in the study sites.
3.2. Litter Mass Loss and Decomposition Constant k
After the four decomposition years, the A. faxoniana litter mass loss reached 50~53%, and it was significantly affected by gap size, elevation, and decomposition time, as well as their interactions (Table 2). Specifically, the litter in the forest gaps exhibited significantly higher mass loss at the 3300 m and 3600 m sites than that under the closed canopy, especially in the late decomposition period (Figure 1b,c). However, litter at the 3000 m site showed significantly higher mass loss under the closed canopy when compared to those in the forest gaps (Figure 1a). Furthermore, there were higher litter mass loss rates during the non-growing season than during the growing season (Figure 2a,b). Although there were no significant differences, the mass loss was ranked in the order: large gap > middle gap > small gap in the non-growing season, but this showed a opposite trend in the growing season. For the four years, the mass loss was significantly lower at the 3000 m site than that at the 3300 m site. Besides, the decomposition k exhibited a gradually increasing trend from the large gap, middle gap, small gap to closed canopy plots at the 3000 m site, while that at 3300 m and 3600 m sites showed a totally opposite decreasing trend (Figure 1d).
Table 2.
Results (F-values) of linear mixed effects models testing the effects of elevation (3000, 3300, 3600 m), gap size (large, middle, small), decomposition time (sampled times) and their interactions on mass loss and carbon content and release in in decomposing A. faxoniana litter. ** p < 0.001.
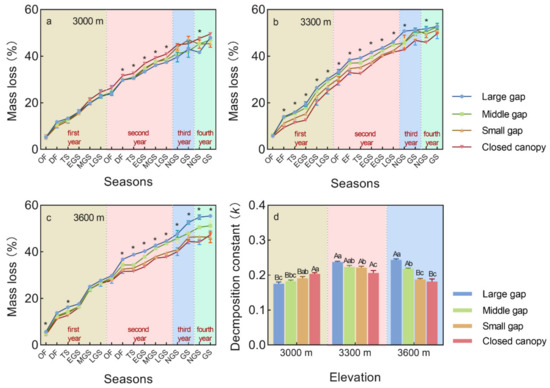
Figure 1.
Mass loss and decomposition constant (k) of A. faxoniana litter in forest gaps (large gap, middle gap, small gap, closed canopy) along an elevation gradient (3000, 3300, 3600 m) at different decomposition stages in a subalpine forest of southwestern China. (a) Mass loss at 3000 m; (b) Mass loss at 3300 m; (c) Mass loss at 3600 m; (d) Decomposition constant k. Mean ± SE, n = 3. OF, onset of freezing stage; DF, deep freezing stage; TS, thawing stage; EGS, early snow-free stage; MGS, middle snow-free stage; LGS, late snow-free stage; NGS, non-growing season; GS, growing season. Asterisk above lines indicates significant differences among forest gaps (p < 0.05), lowercase letters above columns indicate significant differences among forest gaps (p < 0.05), capital letters above columns indicate significant differences among elevations (p < 0.05).
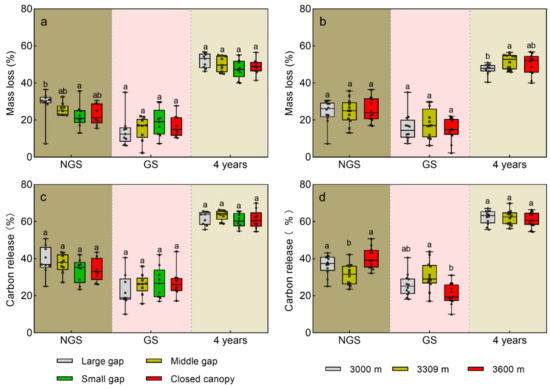
Figure 2.
Effects of forest size and elevation on mass loss and carbon release in decomposing A. faxoniana litter. (a) Mass loss on forest gap scale; (b) Mass loss on elevation scale; (c) Carbon release on forest gap scale; (d) Carbon release on elevation scale. Mean ± SE, n = 3. NGS, the non-growing seasons in the four decomposing years; GS, the growing seasons in the four decomposing years. Lowercase letters indicate significant differences among forest gaps or elevations (p < 0.05).
3.3. Litter Carbon Content
During the four decomposition years, the A. faxoniana litter content gradually decreased by 18~29%, and it was significantly affected by gap size, elevation, decomposition time as well as their interactions (Figure 3, Table 2). Specifically, obvious declines were observed in the first non-growing season and the second growing season during the first two years. When compared with the closed canopy, higher carbon contents were shown in forest gaps at the 3000 m and 3300 m sites. However, at the 3000 m site, the highest carbon content was measured under the closed canopy in the first decomposition year, and then exhibited the following order: large gap > closed canopy > middle/small gap. Additionally, the non-linear regression indicates that the carbon content significantly declined with decomposition processing, meanwhile the decomposed litter with less remaining mass had a higher carbon content at the 3300 and 3600 m sites than that at 3000 m site (Figure 4a,b).
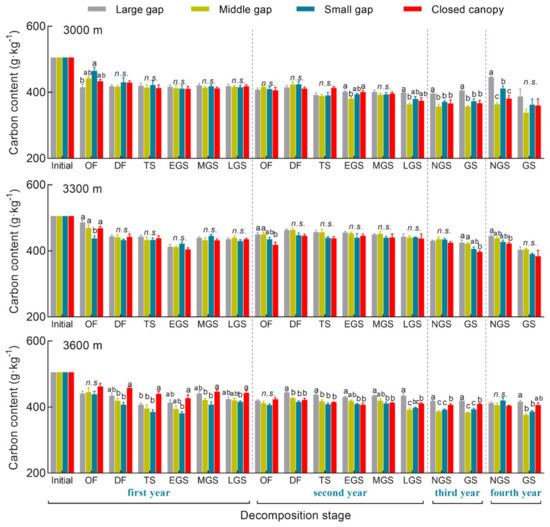
Figure 3.
Carbon content of A. faxoniana litter in forest gaps (large gap, middle gap, small gap, closed canopy) along an elevation gradient (3000, 3300, 3600 m) at different decomposition stages in a subalpine forest of southwestern China. Mean ± SE, n = 3. OF, onset of freezing stage; DF, deep freezing stage; TS, thawing stage; EGS, early snow-free stage; MGS, middle snow-free stage; LGS, late snow-free stage; NGS, non-growing season; GS, growing season. Lowercase letters indicate significant differences among forest gaps (p < 0.05), n.s. indicates no significant differences (p > 0.05).
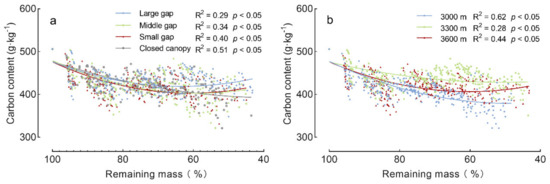
Figure 4.
Carbon content versus remaining mass in decomposing A. faxoniana litter. (a) Forest gap scale; (b) Elevation scale. R2 and p values from linear regression are shown in each panel.
3.4. Litter Carbon Release
After the four decomposition years, 58~64% of carbon was released from the A. faxoniana litter; carbon release was not affected by the gap size (Figure 5), but within the time period, it was significantly affected by gap size, elevation, decomposition time as well as their interactions (Table 2). When compared to the 3000 m and 3300 m sites, the carbon release variation was greater at the 3600 m site. When compared with closed canopy, more carbon was released from decomposing litter in forest gaps at the 3300 m and 3600 m sites. However, less carbon was released from decomposing litter in the large/small gasp at the 3000 m site. At the end of decomposition, there were no significant differences among forest gaps and the closed canopy at all sites, and significant differences were only observed in the first two years. Under the closed canopy, less carbon was released from decomposing litter at the 3300 and 3600 m sites in the first year but more carbon was released at the 3000 and 3300 m sites in the second year. Carbon released in the first two years accounted for 81~86% of the total four-year release, while that of the combined non-growing seasons’ release across the four years accounted for 45~63%. There were significant differences in litter carbon release in the combined non-growing seasons across the four years among the gap sizes at the 3600 m site, but there were no significant differences in the combined growing seasons at all the sites. Further, there was a higher litter carbon release during the non-growing season than during the growing season (Figure 2c,d). The effects of gap size on carbon release were not significant, but at 3300 m they were significant higher and lower in the non-growing season and growing season, respectively.
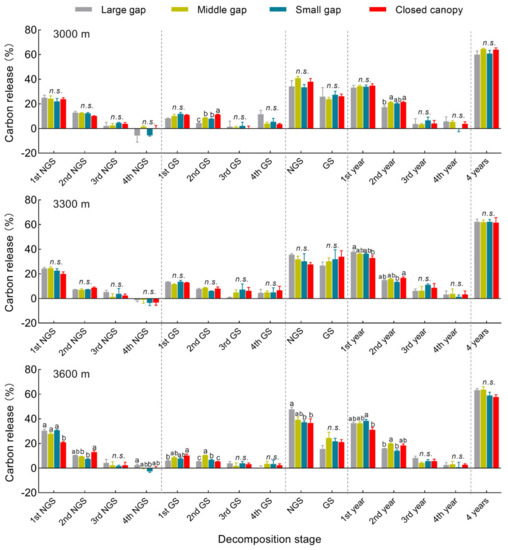
Figure 5.
Carbon release percentage of A. faxoniana litter in forest gaps (large gap, middle gap, small gap, closed canopy) along an elevation gradient (3000, 3300, 3600 m) at different decomposition stages in a subalpine forest of southwestern China. Mean ± SE, n = 3. 1st NGS, non-growing season in the first decomposing year; 2nd NGS, non-growing season in the second decomposing year; 3rd NGS, non-growing season in the third decomposing year; 4th NGS, non-growing season in the fourth decomposing year; 1st GS, growing season in the first decomposing year; 2nd GS, growing season in the second decomposing year; 3rd GS, growing season in the third decomposing year; 4th GS, growing season in the fourth decomposing year; NGS, the non-growing seasons in the four decomposing years; GS, the growing seasons in the four decomposing years. Lowercase letters indicate significant differences among forest gaps (p < 0.05), n.s. indicates no significant differences (p > 0.05).
3.5. Key Drivers of Litter Mass Loss and Carbon Release
Based on the Spearman correlation analysis, litter mass loss and carbon release were both significantly negative correlated with freeze–thaw cycles in the non-growing season, and carbon release was significantly negative correlated with freeze–thaw cycles for the whole four decomposition years. Besides, the litter carbon release was also significantly negative correlated with the positive accumulated temperature and the mean temperature in the non-growing season (Table 3).
Table 3.
Spearman correlation coefficient between mass loss, carbon release and environmental factors in the non-growing season, growing season and the 4 total decomposition years. Significant correlations are indicated in a bold font, with * p < 0.05, ** p < 0.001.
4. Discussion
After four years of decomposition, 50~53 and 58~64% of mass and carbon were lost (Figure 1 and Figure 3), which were similar to other pine tree species in other temperate forests [44,45], and more than 50% of the losses were observed in the 3600 m sites during the non-growing seasons (Figure 2), indicating rather high mass loss and carbon release in decomposition. A. faxoniana litter was present at higher elevations of the subalpine forest subjected to low temperatures in the non-growing seasons. Forest gaps formed by natural stem breakage were commonly distributed in high-latitude and high-altitude ecosystems, which would modify the temperature, precipitation, snow coverage and radiation conditions within the gap areas when compared to the closed canopies [10,46]. Our results partially support the first hypothesis that forest gaps increased the mass loss and release in the decomposing A. faxoniana leaf litter, but only in the 3300 m and 3600 m sites. For the decomposition constant (k), we found that higher decomposition rates were in the gaps when compared to the closed canopy in both the 3300 and 3600 m sites. A previous study has demonstrated that there was more accumulated snow coverage within gaps, helping microbes maintain high activities through the isolated protection in winter, as well as the more intense precipitation and strong radiation attributed to the opening canopy increasing the leaching loss and photodegradation in the growing season, thereby promoting the litter mass loss [8,47], which was consist with our results. However, the gap-stimulated effects were not observed in the 3000 m site, exhibiting a higher decomposition rate under the closed canopy. According to the environmental factors presented in Table 1, we recorded relatively small differences in the snow depth among different gaps and the closed canopy; besides, there was a lower frequency of freeze–thaw cycles under closed canopy than in gaps in the non-growing seasons, which was the opposite to the other two elevations. Therefore, a likely explanation was that the lower variables of environmental conditions among the gaps and closed canopy compared to the two high elevations resulted in different effects of gaps on the decomposition rate. Additionally, the Spearman correlation analysis showed that both mass loss and carbon release were significantly negatively related to the freeze–thaw cycles (Table 3), implying that the effects of gaps on litter decomposition were more correlated to the freeze–thaw cycles in winter. It has been reported that soil freezing would modify carbon release from plant litter in the subalpine forests of this region [9,48], and our study addressed that freeze–thaw cycle variations were a key driver changing the effects of gaps on litter decomposition at different elevations of high-altitude forest ecosystems, which also supported our third hypothesis.
In this study, the effects of forest gaps were not always in accordance with the gap size gradient. Previous studies have revealed that nitrogen mineralization in forest gaps increased in the first decomposition year and then decreased later [49,50]; thus, the soil microorganisms contributing to the degradation of carbon-associated compounds at the early decomposition stage would benefit from increasing the nitrogen availability [46,51]. However, carbon release did not consistently present a general pattern along with the gap size gradient (Figure 2c). In general, although deep accumulated snowpack on the forest floor within gaps decreased the contribution of soil freezing to the physical breakdown of litter plant-derived materials [52,53], higher biological activities were still maintained under the snow cover [8,54]. Moreover, intense hydrological leaching during the snowmelt period also contributes to the labile carbon releasing from newly shed litter [55,56]. These parts of carbon, in turn, provide available carbon and nutrients for microbial utilization and thus affect the decomposition processes [50,57]. When compared with the lower elevation sites, the effect of forest gaps was greater in the high elevation site (3600 m) for both carbon content and release from A. faxoniana litter. Previous studies revealed that cold regions are proven to be more sensitive to environmental disturbances, and the relatively lower temperatures in the high elevation site are at much higher risk of promoting soil freezing, which would physically favor the decomposition of the A. faxoniana litter [9]. Meanwhile, snow coverage in forest gaps has also been documented to accelerate carbon-associated compound (both labile and recalcitrant substrate) degradation [58]. Therefore, the stimulative effects of forest gaps on mass loss and carbon release were more responsive to higher elevation in subalpine forests.
The stimulative effects of forest gap on litter decomposition were more remarkable in the non-growing seasons when compared to the growing seasons. Carbon content decreased over time at the three elevations as decomposition proceeded, particularly for the non-growing seasons (Figure 2). Moreover, a rather higher mass loss and carbon release in A. faxoniana litter was observed in subalpine forests subjected to low temperatures in the non-growing seasons. These results suggest that carbon release greatly varies between non-growing and growing seasons in the subalpine forest. When carbon release in the non-growing seasons throughout the four decomposition years was combined, we observed that the large gaps significantly enhanced the carbon release at the 3600 m site, suggesting that deep snow coverage at the high-elevation forest promoted carbon release from the A. faxoniana litter [59]. Forest gaps only exhibited significant effects at the early litter decomposition stage (the first two decomposition years), but such a gap influence was dissimilar, with the increasing carbon release in the first decomposition year and decreasing in the second year. Previous studies indicated that the difference between forest gaps was greater at the late stage of decomposition [23], which is attributed to the disturbances of the original environment resulting from the formation of forest gaps. However, in this study, the gap influence on carbon release disappeared by the end of four decomposition years, which supported our second hypothesis. Another likely explanation was that 56, 50 and 49% of the initial carbon was lost in the first two decomposition years at all the three sites, and the remaining carbon compounds were primarily recalcitrant components with a resistant structure, which decay quite slowly and are not able to be affected by environmental alteration easily [45].
5. Conclusions
Collectively, our study found that (i) rather high mass loss and carbon release in decomposition A. faxoniana litter was observed in high elevations of the subalpine forest subjected to low temperatures in the non-growing seasons and (ii) there were stimulative effects of forest gaps on litter decomposition in early decomposition, especially in the non-growing seasons, driven by fewer freeze–thaw cycles when compared to the closed canopy, which diminished at the end of the experiment though. Overall, these findings provide new insights into how opening gaps as a main regeneration method at different elevations would induce changes in carbon cycling in subalpine forests. We therefore suggest that future models of litter decomposition may benefit from including forest gaps and emphasizing non-growing seasons to make more accurate predictions. However, the biological mechanisms involved in the decomposition processes may need further investigation, such as considering the microbial and faunal strategy reactions to the forest gap formation.
Supplementary Materials
The following supporting information can be downloaded at: https://www.mdpi.com/article/10.3390/f13081201/s1, Table S1: Geographic information of forest gaps.
Author Contributions
Conceptualization, H.L. (Han Li), Q.W. and L.Z.; software, T.D., Y.C. and J.Y.; validation, T.D., Y.C., Y.Z. and Q.D.; data curation, H.L. (Han Li); writing—original draft preparation, H.L. (Han Li) and Q.W.; writing—review and editing, H.L. (Han Li), L.Z. and Q.W.; visualization, H.L. (Han Li), Y.Y. and L.Z.; funding acquisition, H.L. (Han Li), L.Z. and Q.W. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by the National Natural Science Foundation of China (31901295, 32071747 and 32001165), the Program of Sichuan Applied Basic Research Foundation (2022NSFSC0997, 2022NSFSC0083, 2022NSFSC0087 and 2022NSFSC1173), the Open Fund of Ecological Security and Protection Key Laboratory of Sichuan Province Mianyang Normal University (ESP1902), and Research Fund of Mianyang Normal University (QD2020A18).
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Written informed consent has been obtained from the patient to publish this paper.
Data Availability Statement
Not applicable.
Acknowledgments
We are grateful to the Long-term Research Station of Alpine Forest Ecosystems and the Collaborative Innovation Center of Ecological Security in the Upper Reaches of the Yangtze River.
Conflicts of Interest
The authors declare no conflict of interest. The founding sponsors had no role in the design of the study; in the collection, analyses, or interpretation of data; in the writing of the manuscript, and in the decision to publish the results.
References
- Ssmaneh, T.; Shamsollah, A.; Jahangir, K.; Shaban, S. Effects of Tree Species Composition on Soil Properties and Invertebrates in A Deciduous Forest. Arab. J. Geosci. 2019, 12, 368. [Google Scholar]
- Prescott, C.E.; Grayston, S.J.; Helmisaari, H.S.; Kaštovská, E.; Körner, C.; Lambers, H.; Meier, I.C.; Millard, P.; Ostonen, I. Surplus Carbon Drives Allocation and Plant–Soil Interactions. Trends Ecol. Evol. 2020, 35, 1110–1118. [Google Scholar] [CrossRef] [PubMed]
- Prescott, C.E. Litter Decomposition: What Controls It and How Can We Alter It to Sequester More Carbon in Forest Soils? Biogeochemistry 2010, 101, 133–149. [Google Scholar] [CrossRef]
- Berg, B. Decomposition Patterns for Foliar Litter—A Theory for Influencing Factors. Soil Biol. Biochem. 2014, 78, 222–232. [Google Scholar] [CrossRef]
- Aerts, R. The Freezer Defrosting: Global Warming and Litter Decomposition Rates in Cold Biomes. J. Ecol. 2006, 94, 713–724. [Google Scholar] [CrossRef]
- Schliemann, S.A.; Bockheim, J.G. Methods for Studying Treefall Gaps: A Review. For. Ecol. Manag. 2011, 261, 1143–1151. [Google Scholar] [CrossRef]
- Ritter, E. Litter Decomposition and Nitrogen Mineralization in Newly Formed Gaps in a Danish Beech (Fagus sylvatica) Forest. Soil Biol. Biochem. 2005, 37, 1237–1247. [Google Scholar] [CrossRef]
- González, G.; Lodge, D.J.; Richardson, B.A.; Richardson, M.J. A Canopy Trimming Experiment in Puerto Rico: The Response of Litter Decomposition and Nutrient Release to Canopy Opening and Debris Deposition in a Subtropical Wet Forest. For. Ecol. Manag. 2014, 332, 32–46. [Google Scholar] [CrossRef]
- Gliksman, D.; Haenel, S.; Osem, Y.; Yakir, D.; Zangy, E.; Preisler, Y.; Grünzweig, J.M. Litter decomposition in Mediterranean pine forests is enhanced by reduced canopy cover. Plant Soil 2018, 422, 317–329. [Google Scholar] [CrossRef]
- Tan, B.; Zhang, J.; Yang, W.Q.; Yin, R.; Xu, Z.F.; Liu, Y.; Zhang, L.; Li, H.; You, C.M. Forest gaps retard carbon and nutrient release from twig litter in alpine forest ecosystems. Eur. J. For. Res. 2020, 139, 53–65. [Google Scholar] [CrossRef]
- Prescott, C.E.; Blevins, L.L.; Staley, C.L. Effects of clear-cutting on decomposition rates of litter and forest floor in forests of British Columbia. Can. J. For. Res. 2020, 30, 1751–1757. [Google Scholar] [CrossRef]
- Sariyildiz, T. Effects of Gap-Size Classes on Long-Term Litter Decomposition Rates of Beech, Oak and Chestnut Species at High Elevations in Northeast Turkey. Ecosystems 2008, 11, 841–853. [Google Scholar] [CrossRef]
- Obojes, N.; Meurer, A.; Newesely, C.; Tasser, E.; Oberhuber, W.; Mayr, S.; Tappeiner, U. Water Stress Limits Transpiration and Growth of European Larch up to the Lower Subalpine Belt in an Inner-Alpine Dry Valley. New Phytol. 2018, 220, 460. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cao, R.; Yang, W.; Chang, C.; Wang, Z.; Wang, Q.; Jiang, Y.; Li, H.; Tan, B. Soil Microbial Biomass Carbon and Freeze-Thaw Cycles Drive Seasonal Changes in Soil Microbial Quotient Along a Steep Altitudinal Gradient. J. Geophys. Res. Biogeosci. 2021, 126, e2021JG006325. [Google Scholar] [CrossRef]
- Cao, R.; Yang, W.; Chang, C.; Wang, Z.; Wang, Q.; Li, H.; Tan, B. Differential Seasonal Changes in Soil Enzyme Activity along an Altitudinal Gradient in an Alpine-Gorge Region. Appl. Soil Ecol. 2021, 166, 104078. [Google Scholar] [CrossRef]
- Withington, C.L.; Sanford, R.L. Decomposition Rates of Buried Substrates Increase with Altitude in the Forest-Alpine Tundra Ecotone. Soil Biol. Biochem. 2007, 39, 68–75. [Google Scholar] [CrossRef]
- Fierer, N.; Mccain, C.M.; Meir, P.; Zimmermann, M.; Rapp, J.M.; Silman, M.R.; Knight, R. Microbes Do Not Follow the Elevational Diversity Patterns of Plants and Animals. Ecology 2011, 92, 797–804. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Körner, C. The Use of “altitude” in Ecological Research. Trends Ecol. Evol. 2007, 22, 569–574. [Google Scholar] [CrossRef] [PubMed]
- Gutiérrez-Girón, A.; Díaz-Pinés, E.; Rubio, A.; Gavilán, R.G. Both Altitude and Vegetation Affect Temperature Sensitivity of Soil Organic Matter Decomposition in Mediterranean High Mountain Soils. Geoderma 2015, 237–238, 1–8. [Google Scholar] [CrossRef]
- Ma, H.P.; Yang, X.L.; Guo, Q.Q.; Zhang, X.J.; Zhou, C.N. Soil Organic Carbon Pool along Different Altitudinal Level in the Sygera Mountains, Tibetan Plateau. J. Mt. Sci. 2016, 13, 476–483. [Google Scholar] [CrossRef]
- He, X.; Hou, E.; Liu, Y.; Wen, D. Altitudinal Patterns and Controls of Plant and Soil Nutrient Concentrations and Stoichiometry in Subtropical China. Sci. Rep. 2016, 6, 24261. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Denslow, J.S.; Ellison, A.M.; Sanford, R.E. Treefall Gap Size Effects on Above- and below-Ground Processes in a Tropical Wet Forest. J. Ecol. 1998, 86, 597–609. [Google Scholar] [CrossRef]
- Zhang, Q.; Liang, Y. Effects of Gap Size on Nutrient Release from Plant Litter Decomposition in a Natural Forest Ecosystem. Can. J. For. Res. 1995, 25, 1627–1638. [Google Scholar] [CrossRef]
- Zhang, Q.; Zak, J.C. Potential Physiological Activities of Fungi and Bacteria in Relation to Plant Litter Decomposition along a Gap Size Gradient in a Natural Subtropical Forest. Microb. Ecol. 1998, 35, 172–179. [Google Scholar] [CrossRef] [PubMed]
- Yin, R.; Qin, W.; Zhao, H.; Wang, X.; Cao, G.; Zhu, B. Climate Warming in an Alpine Meadow: Differential Responses of Soil Faunal vs. Microbial Effects on Litter Decomposition. Biol. Fertil. Soils 2022, 58, 509–514. [Google Scholar] [CrossRef]
- Tan, B.; Yin, R.; Zhang, J.; Xu, Z.; Liu, Y.; He, S.; Zhang, L.; Li, H.; Wang, L.; Liu, S.; et al. Temperature and Moisture Modulate the Contribution of Soil Fauna to Litter Decomposition via Different Pathways. Ecosystems 2021, 24, 1142–1156. [Google Scholar] [CrossRef]
- Li, H.; Wu, F.; Yang, W.; Xu, L.; Ni, X.; He, J.; Tan, B.; Hu, Y. Effects of Forest Gaps on Litter Lignin and Cellulose Dynamics Vary Seasonally in an Alpine Forest. Forests 2016, 7, 27. [Google Scholar] [CrossRef] [Green Version]
- Blok, D.; Elberling, B.; Michelsen, A. Initial stages of tundra shrub litter decomposition may be accelerated by deeper winter snow but slowed down by spring warming. Ecosystems 2016, 19, 155–169. [Google Scholar] [CrossRef] [Green Version]
- Gong, L.; Chen, X.; Zhang, X.N.; Yang, X.D.; Cai, Y.J. Schrenk spruce leaf litter decomposition varies with snow depth in the Tianshan Mountains. Sci. Rep. 2020, 10, 19556. [Google Scholar] [CrossRef] [PubMed]
- Baptist, F.; Yoccoz, N.G.; Choler, P. Direct and Indirect Control by Snow Cover over Decomposition in Alpine Tundra along a Snowmelt Gradient. Plant Soil 2010, 328, 397–410. [Google Scholar] [CrossRef]
- He, W.; Wu, F.; Yang, W.; Tan, B.; Zhao, Y.; Wu, Q.; He, M. Lignin Degradation in Foliar Litter of Two Shrub Species from the Gap Center to the Closed Canopy in an Alpine Fir Forest. Ecosystems 2016, 19, 115–128. [Google Scholar] [CrossRef]
- Wu, Q. Short- and Long-Term Effects of Snow-Depth on Korean Pine and Mongolian Oak Litter Decomposition in Northeastern China. Ecosystems 2020, 23, 662–674. [Google Scholar] [CrossRef]
- Ni, X.; Yang, W.; Liao, S.; Li, H.; Tan, B.; Yue, K.; Xu, Z.; Zhang, L.; Wu, F. Rapid Release of Labile Components Limits the Accumulation of Humic Substances in Decomposing Litter in an Alpine Forest. Ecosphere 2018, 9, e02434. [Google Scholar] [CrossRef]
- Wang, Q.W.; Pieristè, M.; Liu, C.; Kenta, T.; Robson, T.M.; Kurokawa, H. The Contribution of Photodegradation to Litter Decomposition in a Temperate Forest Gap and Understorey. New Phytol. 2021, 229, 2625–2636. [Google Scholar] [CrossRef] [PubMed]
- Wu, F.; Yang, W.; Zhang, J.; Deng, R. Litter Decomposition in Two Subalpine Forests during the Freezeethaw Season. Acta Oecologica 2010, 36, 135–140. [Google Scholar] [CrossRef]
- Wu, Q.; Wu, F.; Yang, W.; Tan, B.; Yang, Y.; Ni, X.; He, J. Characteristics of Gaps and Disturbance Regimes of the Alpine Fir Forest in Western Sichuan. Chin. J. Appl. Environ. Biol. 2013, 19, 922–928. [Google Scholar] [CrossRef]
- Zhu, J.; Yang, W.; He, X. Temporal Dynamics of Abiotic and Biotic Factors on Leaf Litter of Three Plant Species in Relation to Decomposition Rate along a Subalpine Elevation Gradient. PLoS ONE 2013, 8, e62073. [Google Scholar] [CrossRef] [Green Version]
- Xu, Z.; Zhu, J.; Wu, F.; Liu, Y.; Tan, B.; Yang, W. Effects of Litter Quality and Climate Change along an Elevational Gradient on Litter Decomposition of Subalpine Forests, Eastern Tibetan Plateau, China. J. For. Res. 2016, 27, 505–511. [Google Scholar] [CrossRef]
- Zhu, J.; He, X.; Wu, F.; Yang, W.; Tan, B. Decomposition of Abies Faxoniana Litter Varies with Freeze-Thaw Stages and Altitudes in Subalpine/Alpine Forests of Southwest China. Scand. J. For. Res. 2012, 27, 586–596. [Google Scholar] [CrossRef]
- IUSS Working Group. WRB World Reference Base for Soil Resources 2014. International Soil Classification System for Naming Soils and Creating Legends for Soil Maps; IUSS Working Group: Vienna, Austria, 2014. [Google Scholar]
- Reed, S.; Martens, D. Methods of Soil Analysis, Part 3: Chemical Methods; John Wiley & Sons: Hoboken, NJ, USA, 1996. [Google Scholar]
- Konestabo, H.S.; Michelsen, A.; Holmstrup, M. Responses of Springtail and Mite Populations to Prolonged Periods of Soil Freeze-Thaw Cycles in a Sub-Arctic Ecosystem. Appl. Soil Ecol. 2007, 36, 136–146. [Google Scholar] [CrossRef]
- Olson, J.S. Energy Storage and the Balance of Producers and Decomposers in Ecological Systems. Ecology 1963, 44, 322–331. [Google Scholar] [CrossRef] [Green Version]
- Qualls, R.G. Long-Term (13 Years) Decomposition Rates of Forest Floor Organic Matter on Paired Coniferous and Deciduous watersheds with Contrasting Temperature Regimes. Forests 2016, 7, 231. [Google Scholar] [CrossRef] [Green Version]
- Rovira, P.; Vallejo, V.R. Labile and Recalcitrant Pools of Carbon and Nitrogen in Organic Matter Decomposing at Different Depths in Soil: An Acid Hydrolysis Approach. Geoderma 2002, 107, 109–141. [Google Scholar] [CrossRef]
- Bauhus, J.; Vor, T.; Bartsch, N.; Cowling, A. The Effects of Gaps and Liming on Forest Floor Decomposition and Soil C and N Dynamics in a Fagus Sylvatica Forest. Can. J. For. Res. 2004, 34, 509–518. [Google Scholar] [CrossRef]
- Bagnato, S.; Marziliano, P.A.; Sidari, M.; Mallamaci, C.; Marra, F.; Muscolo, A. Effects of gap size and cardinal directions on natural regeneration, growth dynamics of trees outside the gaps and soil properties in European beech forests of southern Italy. Forests 2021, 12, 1563. [Google Scholar] [CrossRef]
- Fuzhong, W.; Changhui, P.; Jianxiao, Z.; Jian, Z.; Bo, T.; Wanqin, Y. Impacts of Freezing and Thawing Dynamics on Foliar Litter Carbon Release in Alpine/Subalpine Forests along an Altitudinal Gradient in the Eastern Tibetan Plateau. Biogeosciences 2014, 11, 6471–6481. [Google Scholar] [CrossRef]
- Li, H.; Wu, F.; Yang, W.; Xu, L.; Ni, X.; He, J.; Tan, B.; Hu, Y.; Justin, M.F. The Losses of Condensed Tannins in Six Foliar Litters Vary with Gap Position and Season in an Alpine Forest. iForest 2016, 9, 910. [Google Scholar] [CrossRef] [Green Version]
- Tamura, M.; Tharayil, N. Plant Litter Chemistry and Microbial Priming Regulate the Accrual, Composition and Stability of Soil Carbon in Invaded Ecosystems. New Phytol. 2014, 203, 110–124. [Google Scholar] [CrossRef]
- Hobara, S.; Osono, T.; Hirose, D.; Noro, K.; Hirota, M.; Benner, R. The Roles of Microorganisms in Litter Decomposition and Soil Formation. Biogeochemistry 2014, 118, 471–486. [Google Scholar] [CrossRef]
- Scharenbroch, B.C.; Bockheim, J.G. Gaps and Soil C Dynamics in Old Growth Northern Hardwood-Hemlock Forests. Ecosystems 2008, 11, 426–441. [Google Scholar] [CrossRef]
- Ni, X.; Yang, W.; Li, H.; Xu, L.; He, J.; Tan, B.; Wu, F. The Responses of Early Foliar Litter Humification to Reduced Snow Cover during Winter in an Alpine Forest. Can. J. Soil Sci. 2014, 94, 453–461. [Google Scholar] [CrossRef]
- Groffman, P.M.; Hardy, J.P.; Fisk, M.C.; Fahey, T.J.; Driscoll, C.T. Climate Variation and Soil Carbon and Nitrogen Cycling Processes in a Northern Hardwood Forest. Ecosystems 2009, 12, 927–943. [Google Scholar] [CrossRef]
- Bokhorst, S.; Metcalfe, D.B.; Wardle, D.A. Reduction in Snow Depth Negatively Affects Decomposers but Impact on Decomposition Rates Is Substrate Dependent. Soil Biol. Biochem. 2013, 62, 157–164. [Google Scholar] [CrossRef]
- Zhao, Y.; Wu, F.; Yang, W.; Tan, B.; He, W. Variations in Bacterial Communities during Foliar Litter Decomposition in the Winter and Growing Seasons in an Alpine Forest of the Eastern Tibetan Plateau. Can. J. Microbiol. 2015, 62, 35–48. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Aerts, R.; Callaghan, T.V.; Dorrepaal, E.; van Logtestijn, R.S.P.; Cornelissen, J.H.C. Seasonal Climate Manipulations Have Only Minor Effects on Litter Decomposition Rates and N Dynamics but Strong Effects on Litter P Dynamics of Sub-Arctic Bog Species. Oecologia 2012, 170, 809–819. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guenet, B.; Danger, M.; Abbadie, L.; Lacroix, G. Priming Effect: Bridging the Gap between Terrestrial and Aquatic Ecology. Ecology 2010, 91, 2850–2861. [Google Scholar] [CrossRef] [PubMed]
- Kreyling, J.; Haei, M.; Laudon, H. Snow Removal Reduces Annual Cellulose Decomposition in a Riparian Boreal Forest. Can. J. Soil Sci. 2013, 93, 427–433. [Google Scholar] [CrossRef]
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).