
Response of Understory Avifauna to Annual Flooding of Amazonian Floodplain Forests

 , ,
, ,
Abstract
:1. Introduction
2. Methods
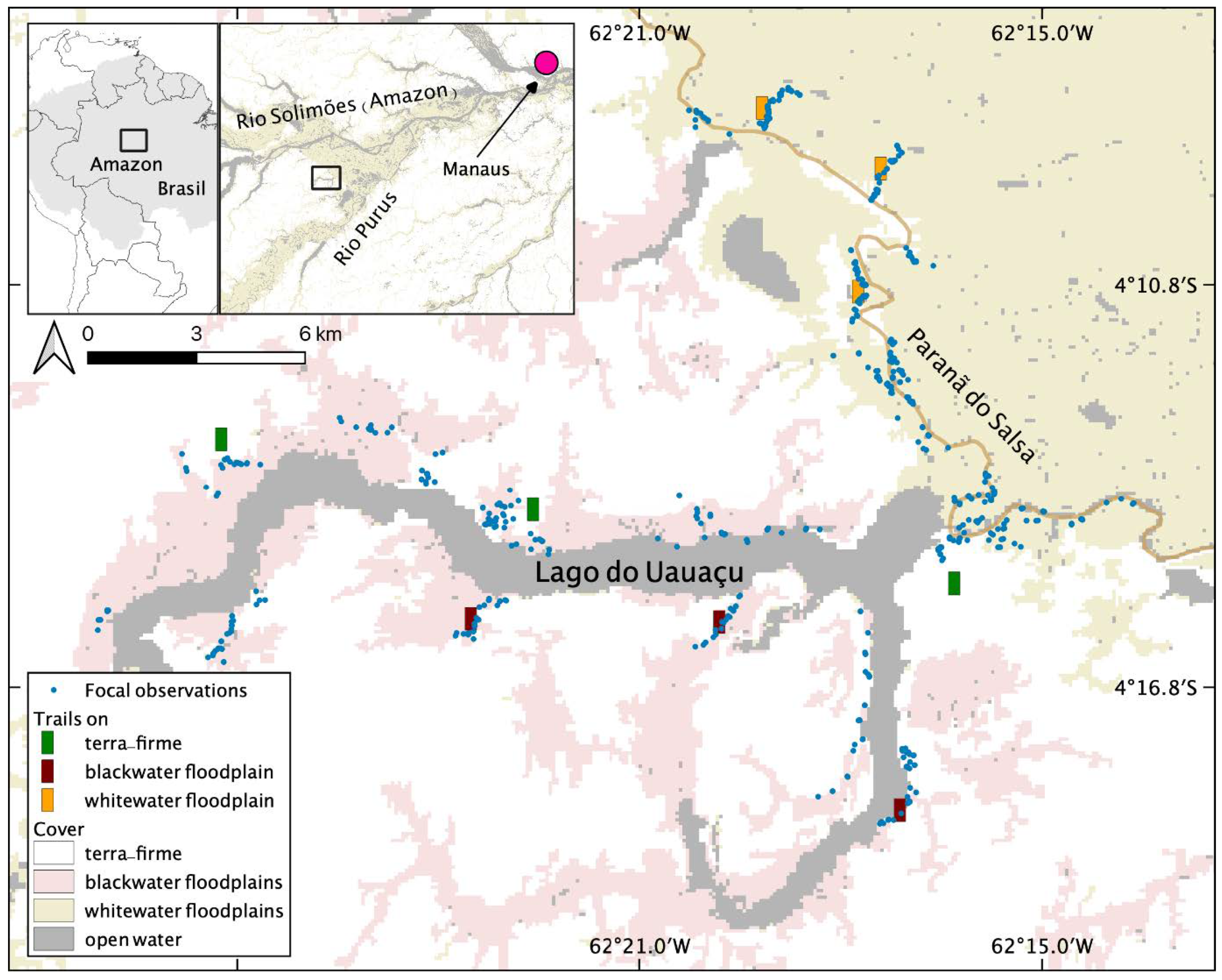
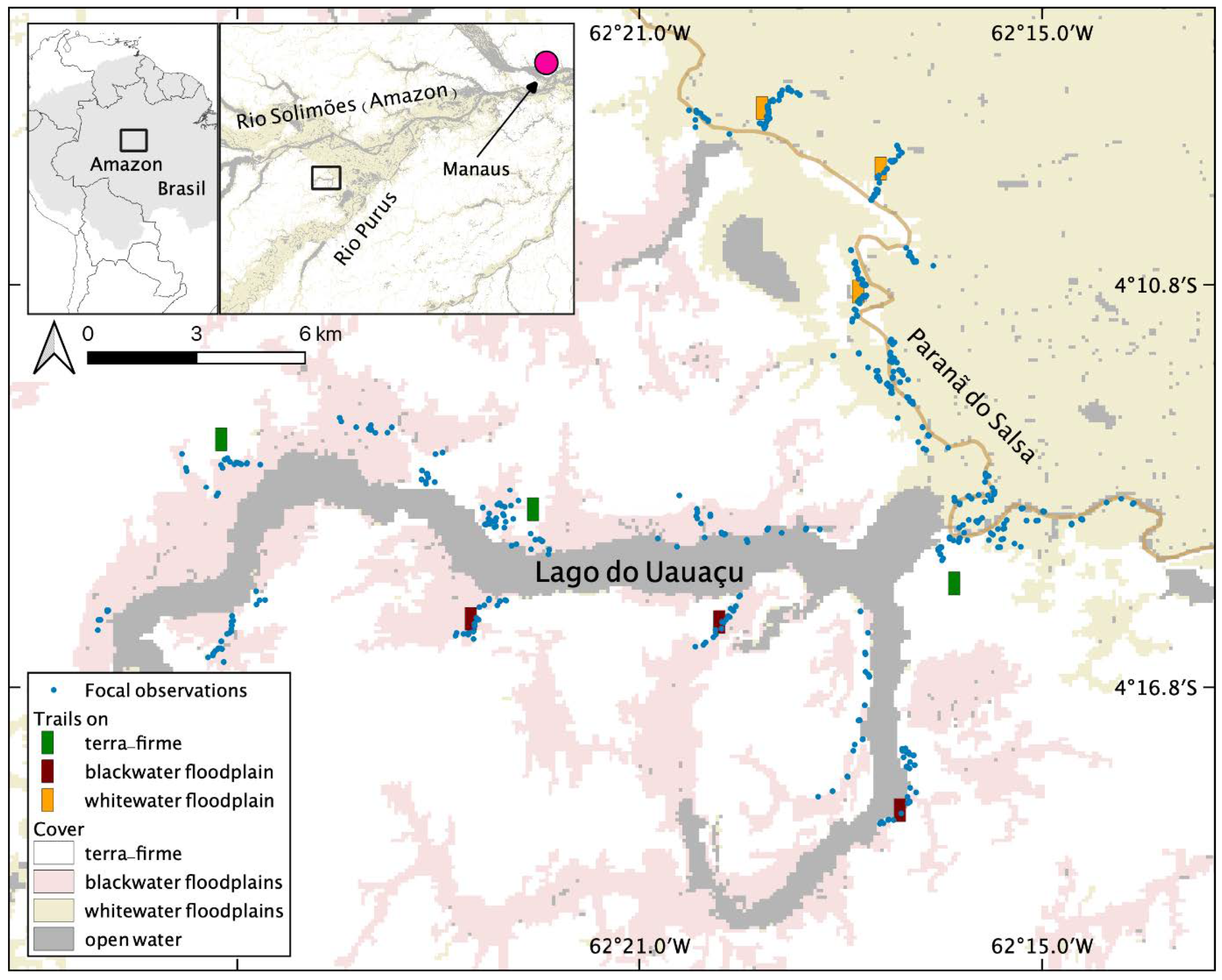
2.1. Study Area
2.2. Focal Bird Species
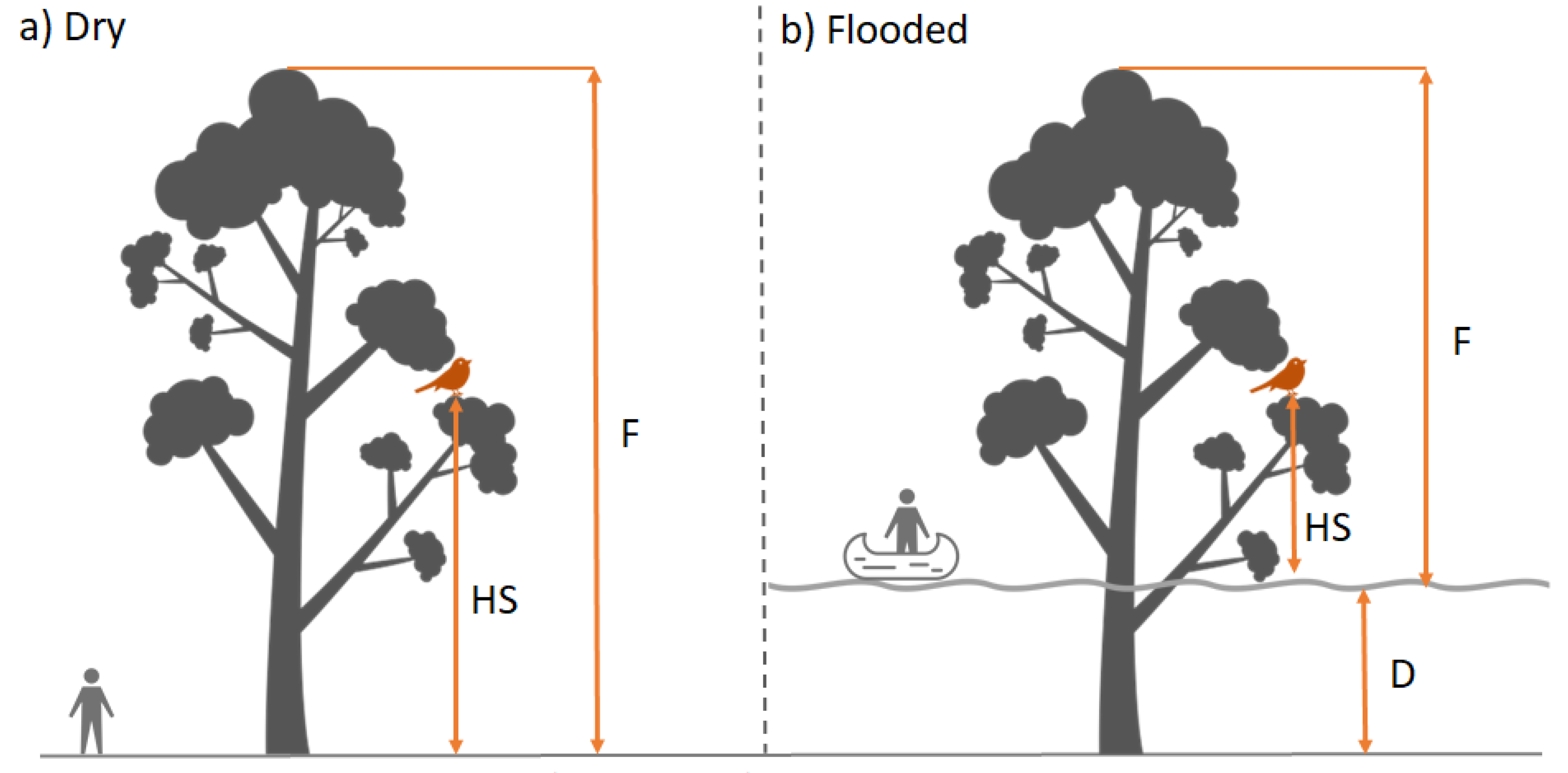
2.3. Sampling Design and Field Measurements
2.4. Data Analyses
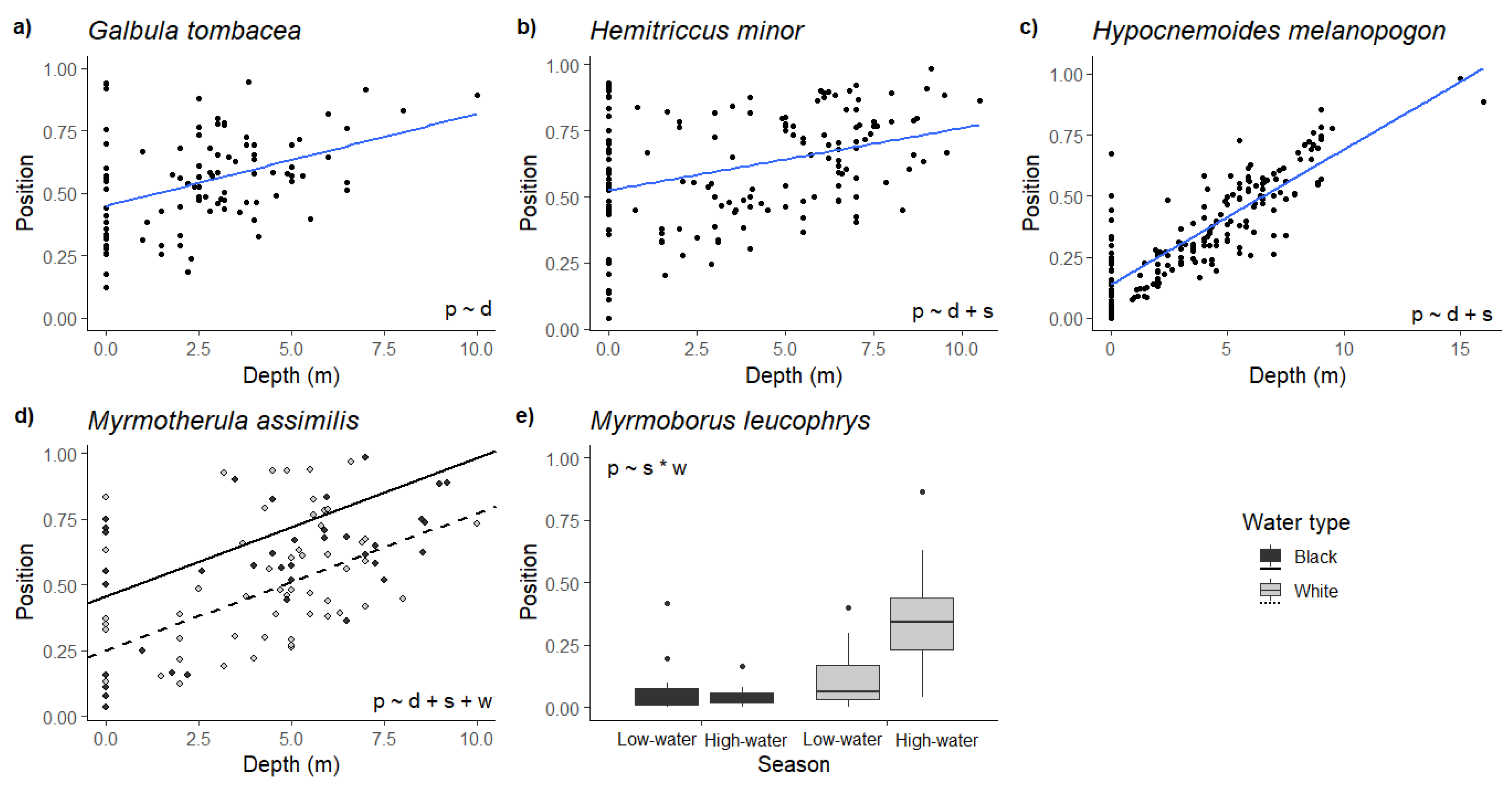
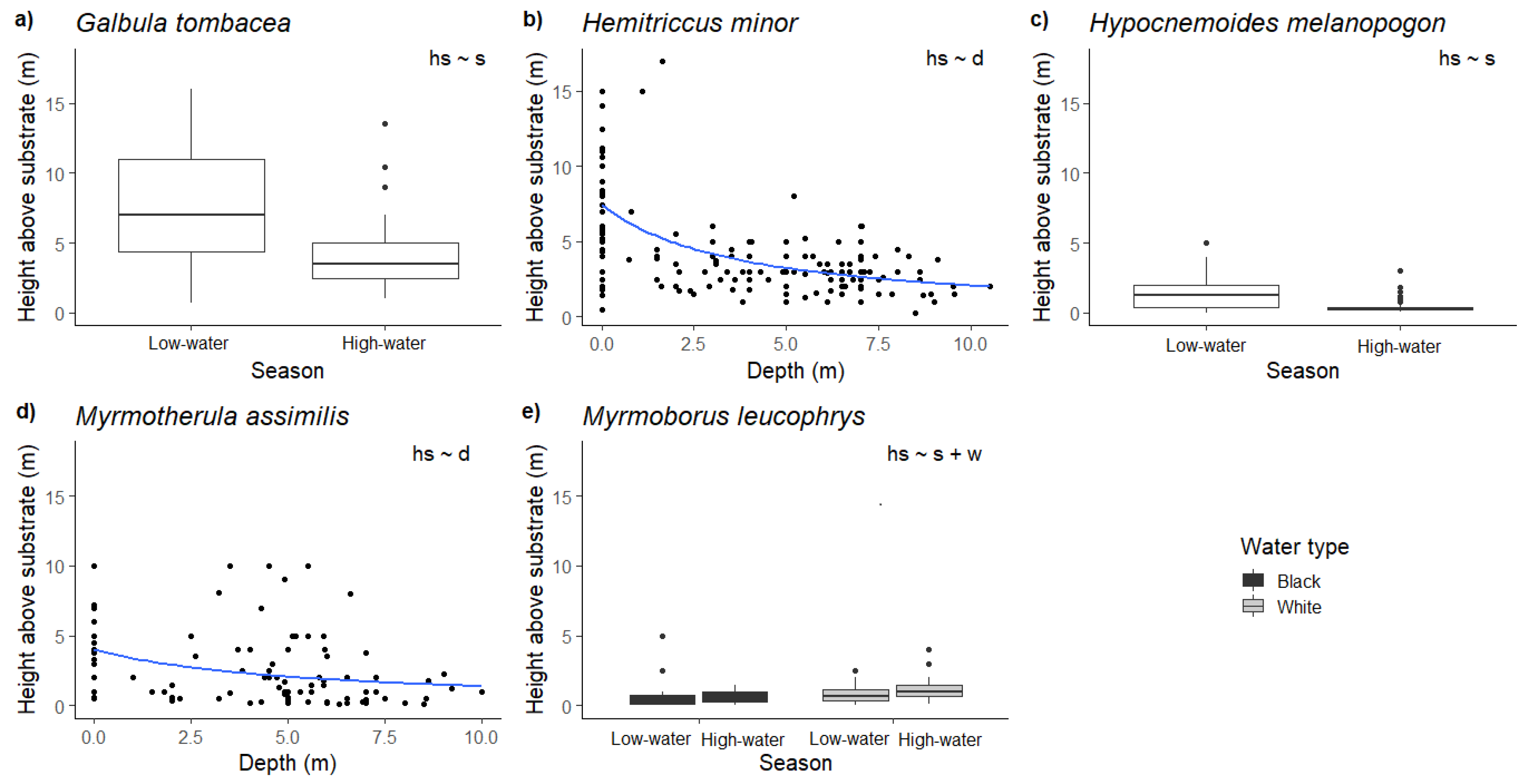
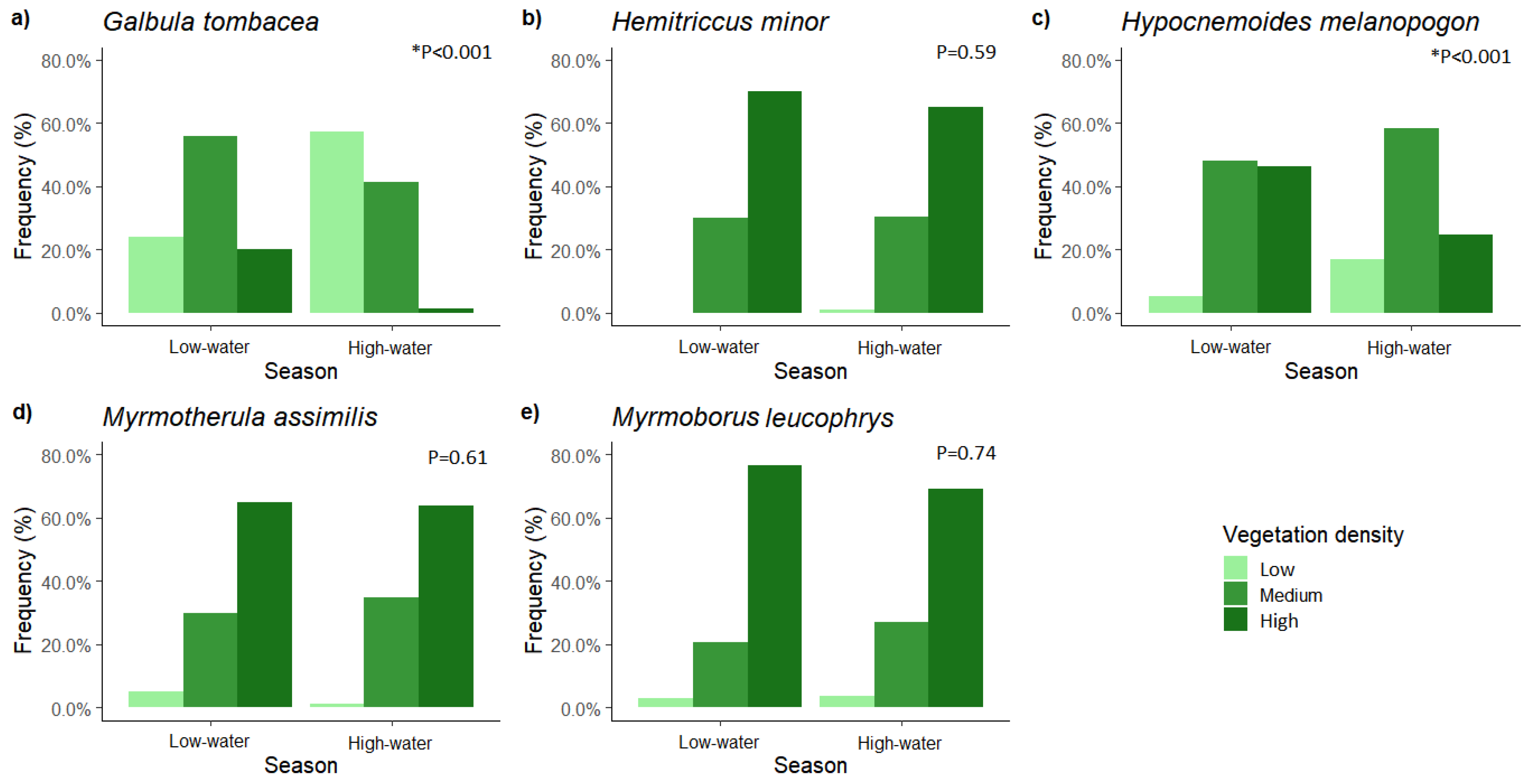
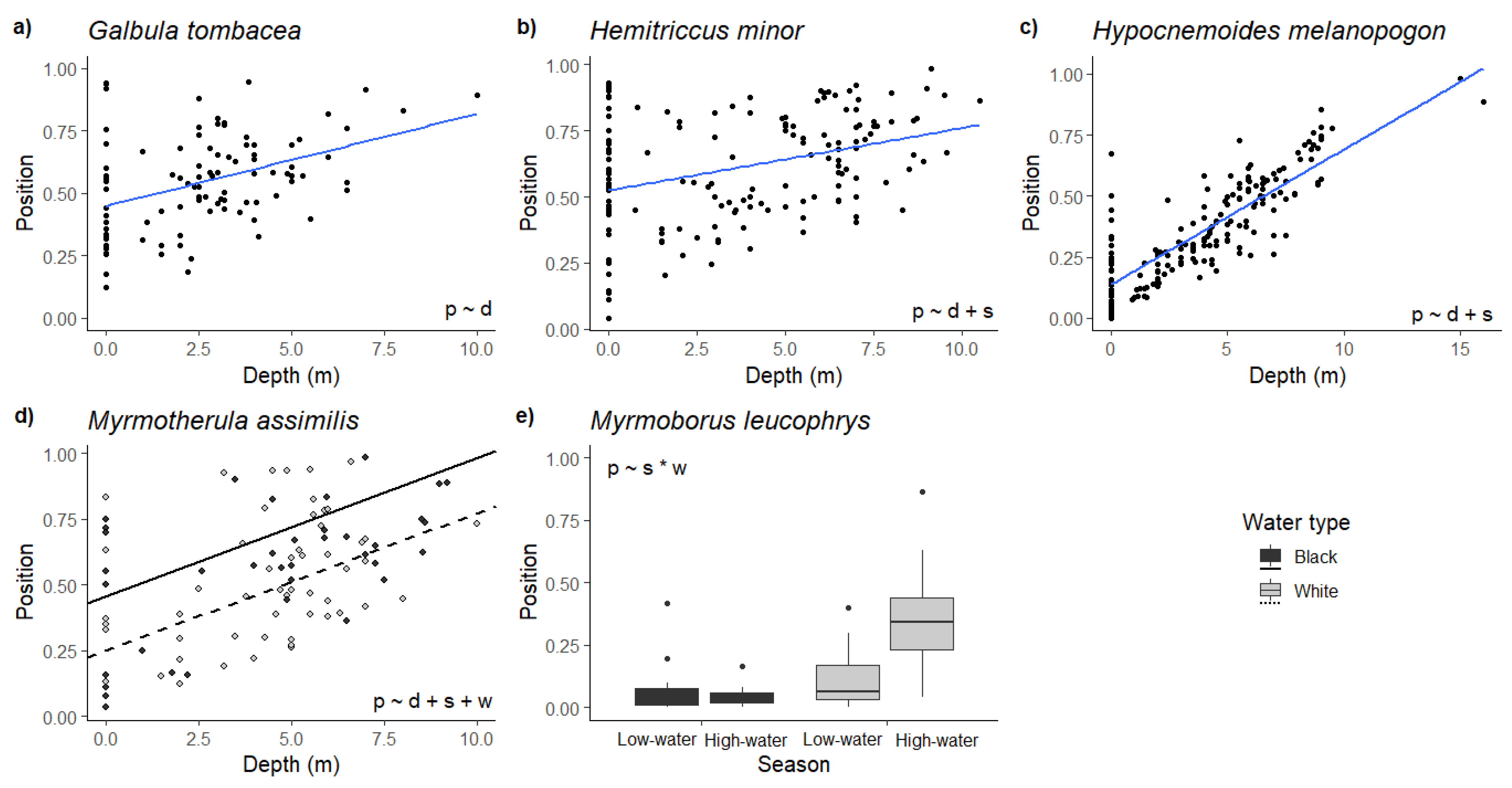
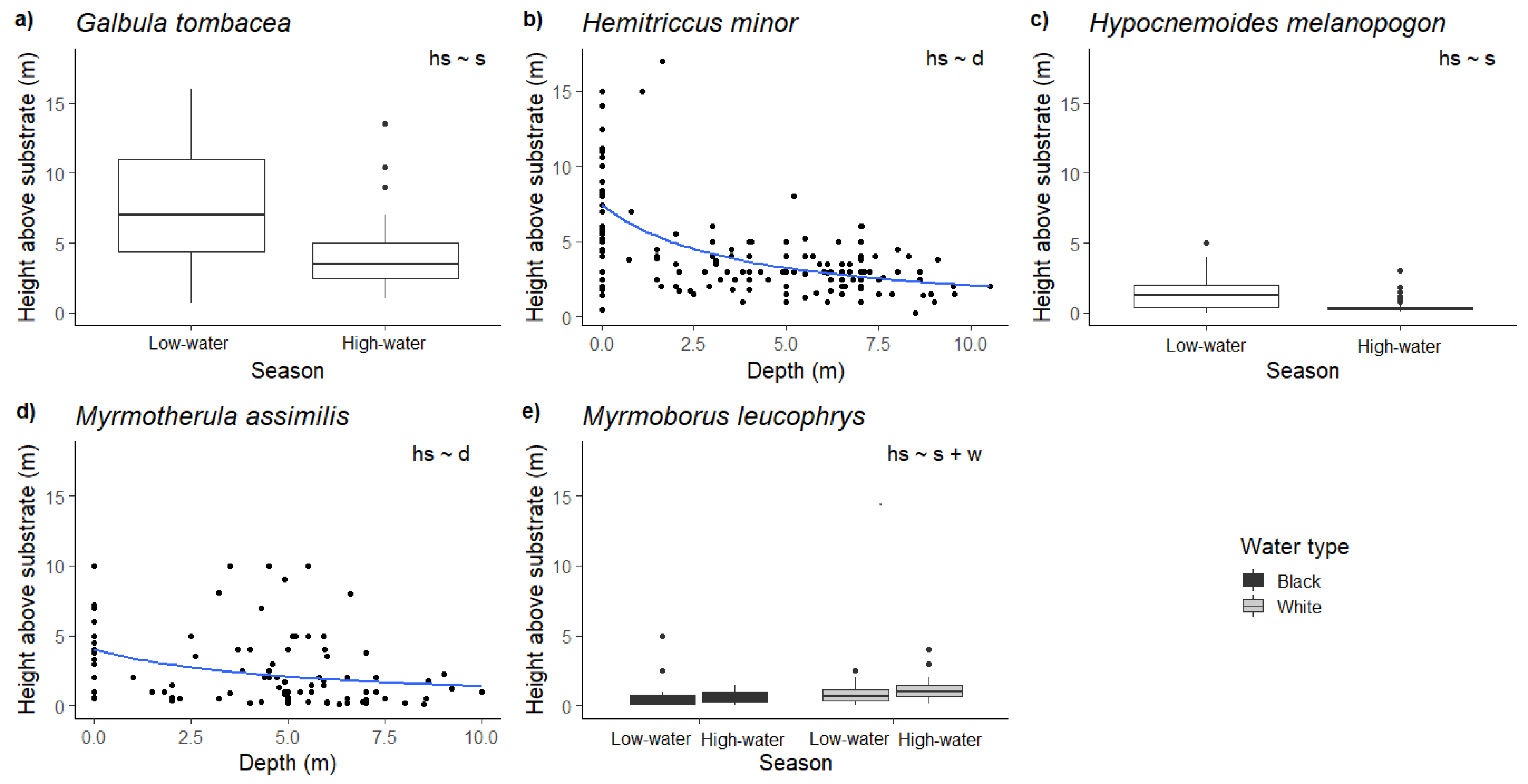
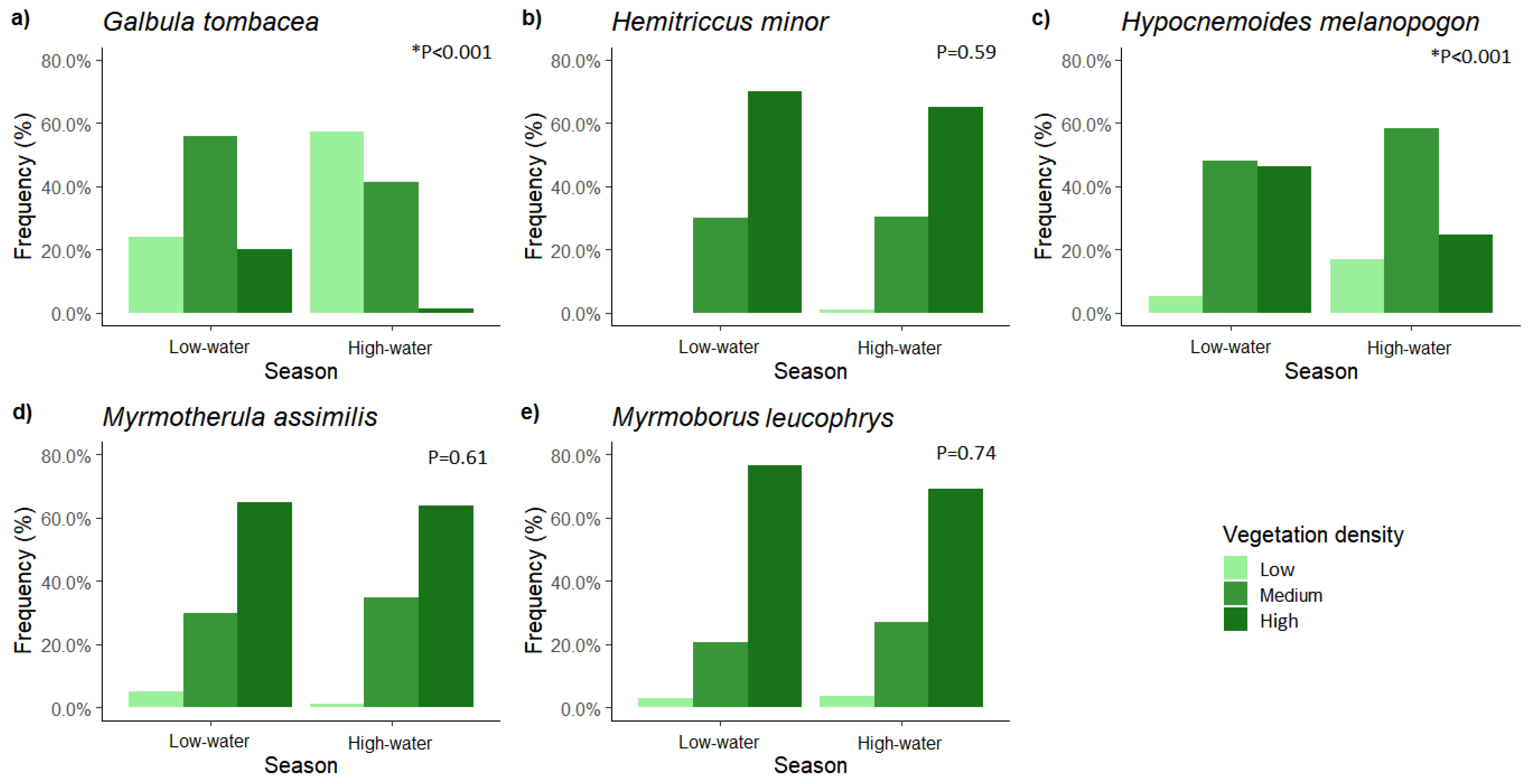
3. Results
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Junk, W.J.; Piedade, M.T.F.; Schöngart, J.; Cohn-Haft, M.; Adeney, J.M.; Wittmann, F. A classification of major naturally-occurring Amazonian lowland wetlands. Wetl. Ecol. Manag. 2011, 31, 623–640. [Google Scholar] [CrossRef]
- Orians, G.H. The number of bird species in some tropical forests. Ecology 1969, 50, 783–801. [Google Scholar] [CrossRef]
- Goulding, M.; Barthem, R.; Ferreira, E.J.G. The Smithsonian Atlas of the Amazon; Smithsonian Institution Press: Washington, DC, USA, 2003. [Google Scholar]
- Adis, J.; Mahnert, V. On the natural history and ecology of Pseudoscorpiones (Arachnida) from an Amazonian blackwater inundation forest. Amaz. Limnol. Oecologia Reg. Syst. Fluminis Amaz. 1985, 9, 297–314. [Google Scholar]
- Adis, J. Estratégias de sobrevivência de invertebrados terrestres em florestas inundáveis da amazônia central: Uma resposta à inundação de longo período. Acta Amaz. 1997, 27, 43–54. [Google Scholar] [CrossRef]
- Haugaasen, T.; Peres, C.A. Vertebrate responses to fruit production in Amazonian flooded and unflooded forests. Biodivers. Conserv. 2007, 16, 4165. [Google Scholar] [CrossRef]
- Costa, H.C.M.; Peres, C.A.; Abrahams, I.M. Seasonal dynamics of terrestrial vertebrate abundance between Amazonian flooded and unflooded forests. PeerJ 2018, 6, e5058. [Google Scholar] [CrossRef] [PubMed]
- Adis, J.; Mahnert, V.; Morais, J.W.; Rodrigues, J.M.G. Adaptation of an Amazonian Pseudoscorpion (Arachnida) from dryland forests to inundation forests. Ecol. Soc. Am. Esa 1988, 69, 287–291. [Google Scholar] [CrossRef]
- Platnick, N.I.; Höfer, H. Systematics and ecology of ground spiders (Araneae, Gnaphosidae) from central Amazonian inundation forests. Am. Mus. Novit. 1990, 2971, 1–16. [Google Scholar]
- Davenport, L.C.; Goodenough, K.S.; Haugaasen, T. Birds of two oceans? Trans-Andean and divergent migration of Black Skimmers (Rynchops niger cinerascens) from the Peruvian Amazon. PLoS ONE 2016, 11, e0144994. [Google Scholar] [CrossRef] [PubMed]
- Remsen, J.V.; Parker, T.A. Contribution of river-created habitats to bird species richness in Amazonia. Biotropica 1983, 15, 223–231. [Google Scholar] [CrossRef]
- Stotz, D.F.; Fitzpatrick, J.W.; Parker, T.A.; Moskovitz, D.K. Neotropical Birds: Ecology and Conservation; University of Chicago Press: Chicago, IL, USA, 1996. [Google Scholar]
- Terborgh, J. Habitat selection in Amazonian birds. In Habitat Selection in Birds; Cody, M.L., Ed.; Academic Press: Orlando, FL, USA, 1985; pp. 311–338. [Google Scholar]
- Pires, J.M.; Prance, G.T. The vegetation types of the Brazilian Amazon. In Key Enviroments: Amazonia; Pergamon Press: Oxford, UK, 1985; pp. 109–145. [Google Scholar]
- Haugaasen, T.; Peres, C.A. Floristic, edaphic and structural characteristics of flooded and unflooded forests in the lower Rio Purús region of central Amazonia, Brasil. Acta Amaz. 2006, 36, 25–36. [Google Scholar] [CrossRef] [Green Version]
- Haugaasen, T.; Peres, C.A. Tree phenology in adjacent Amazonian flooded and unflooded forests. Biotropica 2005, 37, 620–630. [Google Scholar] [CrossRef]
- Laranjeiras, T.O.; Naka, L.N.; Cohn-Haft, M. Using river color to predict Amazonian floodplain forest avifaunas in the world’s largest blackwater river basin. Biotropica 2019, 51, 330–341. [Google Scholar] [CrossRef]
- Gilmore, B.M. Species Richness and Composition of Avifaunal Communities in a Complex Amazonian Landscape. Master’s Thesis, University of Salford, Manchester, UK, 2020; 43p. [Google Scholar]
- Borghesio, L.; Laiolo, P. Seasonal foraging ecology in a forest avifauna of northern Kenya. J. Trop. Ecol. 2004, 20, 145–155. [Google Scholar] [CrossRef]
- Junk, W.J. General aspects of floodplain ecology with special reference to Amazonian floodplains. In The Central Amazon Floodplain; Springer: Berlin/Heidelberg, Germany, 1997; pp. 3–20. [Google Scholar]
- Burnham, K.P.; Anderson, D.R. Model selection and multimodel inference. In A practical Information-Theoretic Approach, 2nd ed.; Springer: Berlin/Heidelberg, Germany, 2002. [Google Scholar]
- R Core Team. Nlme: Linear and Nonlinear Mixed Effects Models. R Package Version 3.1-131. 2017. Available online: https://CRAN.R-project.org/package=nlme (accessed on 25 September 2020).
- Brooks, M.E.; Kristensen, K.; van Benthem, K.J.; Magnusson, A.; Berg, C.W.; Nielsen, A.; Bolker, B.M. Modeling zero-inflated count data with glmmTMB. BioRxiv 2017, 132753. [Google Scholar] [CrossRef] [Green Version]
- Bolker, B.; Bolker, M.B. bbmle: Tools for General Maximum Likelihood Estimation. R Package Version 1.0.20. 2020. Available online: https://rdrr.io/cran/bbmle/ (accessed on 25 September 2020).
- Wickham, H.; Chang, W.; Wickham, M.H. Package ‘ggplot2’. Creat. Elegant Data Vis. Using Gramm. Graph. 2016, 2, 1–189. [Google Scholar]
- Zimmer, K.; Isler, M.L. White-browed Antbird (Myrmoborus leucophrys), version 1.0. In Birds of the World; del Hoyo, J., Elliott, A., Sargatal, J., Christie, D.A., de Juana, E., Eds.; Cornell Lab of Ornithology: Ithaca, NY, USA, 2020. [Google Scholar] [CrossRef]
- Walther, B.A. Grounded ground birds and surfing canopy birds: Variation of foraging stratum breadth observed in Neotropical forest birds and tested with simulation models using boundary constraints. Auk 2002, 119, 658–675. [Google Scholar] [CrossRef]
- Vale, M.M.; Marques, T.L.; Cohn-Haft, M.; Vieira, M.V. Misuse of bird digital distribution maps creates reversed spatial diversity patterns in the Amazon. Biotropica 2017, 49, 636–642. [Google Scholar] [CrossRef]
- Ferreira, L.V. Effects of flooding duration on species richness, floristic composition and forest structure in river margin habitat in Amazonian blackwater floodplain forests: Implications for future design of protected areas. Biodivers. Conserv. 2000, 9, 1–14. [Google Scholar] [CrossRef]
- Powell, L.L.; Cordeiro, N.J.; Stratford, J.A. Ecology and conservation of avian insectivores of the rainforest understory: A pantropical perspective. Biol. Conserv. 2015, 188, 1–10. [Google Scholar] [CrossRef]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Species | Low-Water | High-Water | |||
|---|---|---|---|---|---|
| Blackwater | Whitewater | Blackwater | Whitewater | Total | |
| Galbula tombacea | 0 | 25 | 0 | 77 | 102 |
| Hemitriccus minor | 27 | 13 | 73 | 45 | 158 |
| Hypocnemoides melanopogon | 44 | 12 | 96 | 53 | 205 |
| Myrmotherula assimilis | 10 | 10 | 35 | 54 | 109 |
| Myrmoborus leucophrys | 18 | 16 | 17 | 35 | 86 |
| Total | 99 | 76 | 221 | 264 | 660 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Rowedder, A.R.P.; Laranjeiras, T.O.; Haugaasen, T.; Gilmore, B.; Cohn-Haft, M. Response of Understory Avifauna to Annual Flooding of Amazonian Floodplain Forests. Forests 2021, 12, 1004. https://doi.org/10.3390/f12081004
Rowedder ARP, Laranjeiras TO, Haugaasen T, Gilmore B, Cohn-Haft M. Response of Understory Avifauna to Annual Flooding of Amazonian Floodplain Forests. Forests. 2021; 12(8):1004. https://doi.org/10.3390/f12081004
Chicago/Turabian StyleRowedder, Anaís Rebeca Prestes, Thiago Orsi Laranjeiras, Torbjørn Haugaasen, Benjamin Gilmore, and Mario Cohn-Haft. 2021. "Response of Understory Avifauna to Annual Flooding of Amazonian Floodplain Forests" Forests 12, no. 8: 1004. https://doi.org/10.3390/f12081004
APA StyleRowedder, A. R. P., Laranjeiras, T. O., Haugaasen, T., Gilmore, B., & Cohn-Haft, M. (2021). Response of Understory Avifauna to Annual Flooding of Amazonian Floodplain Forests. Forests, 12(8), 1004. https://doi.org/10.3390/f12081004