Understanding Community Assembly Based on Functional Traits, Ontogenetic Stages, Habitat Types and Spatial Scales in a Subtropical Forest

 ,
,
Abstract

1. Introduction
2. Materials and Methods
2.1. Study Site
2.2. Functional Traits, Topography and Soil Data Measurements
2.3. Habitat Types, Ontogenetic Stages and Spatial Scales Classification
2.4. Trait-Based Analysis of Community Structure
2.5. Variance Partitioning Analyses
3. Results
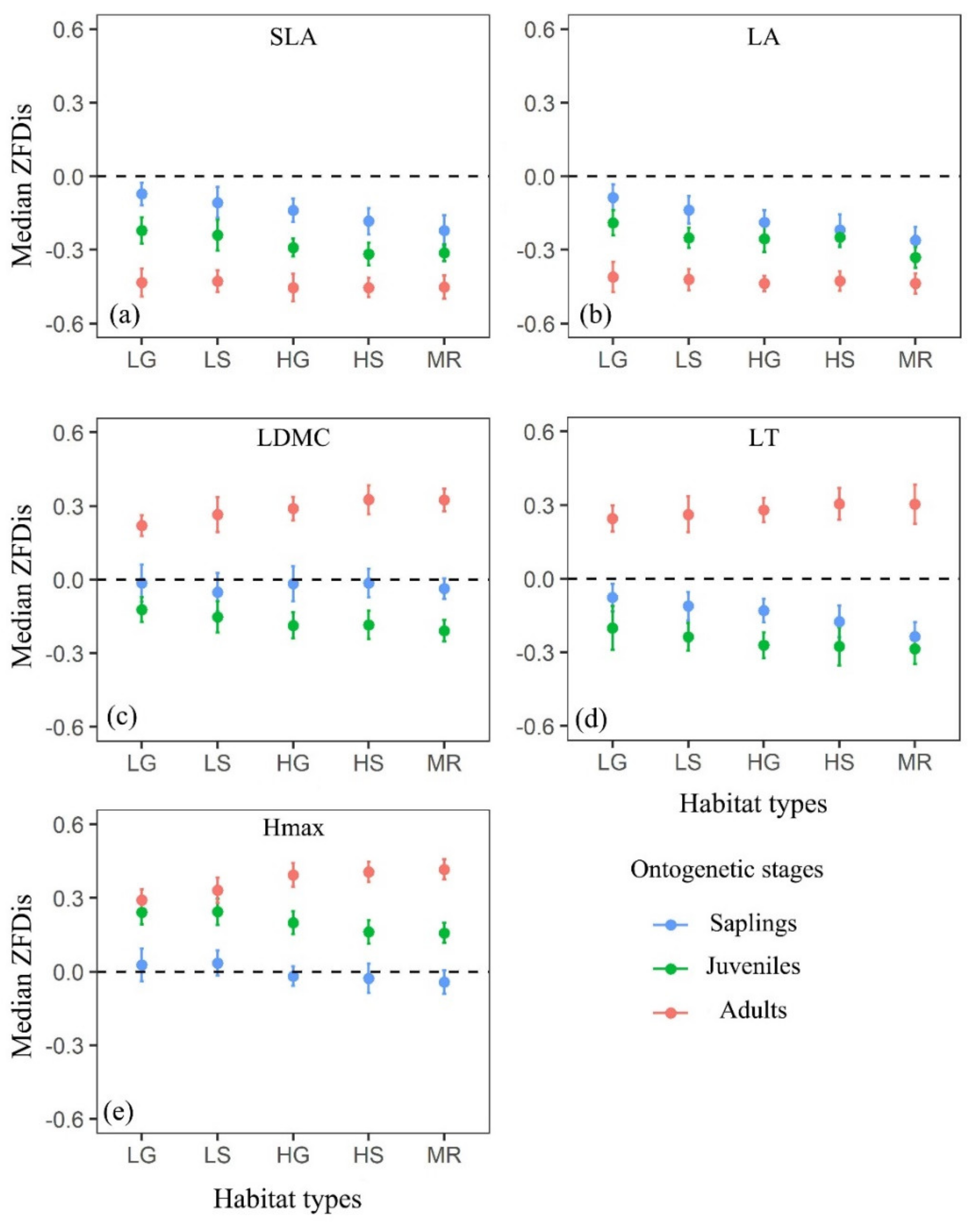
3.1. Trait Dispersions Across Different Functional Traits, Ontogenetic Stages and Habitat Types
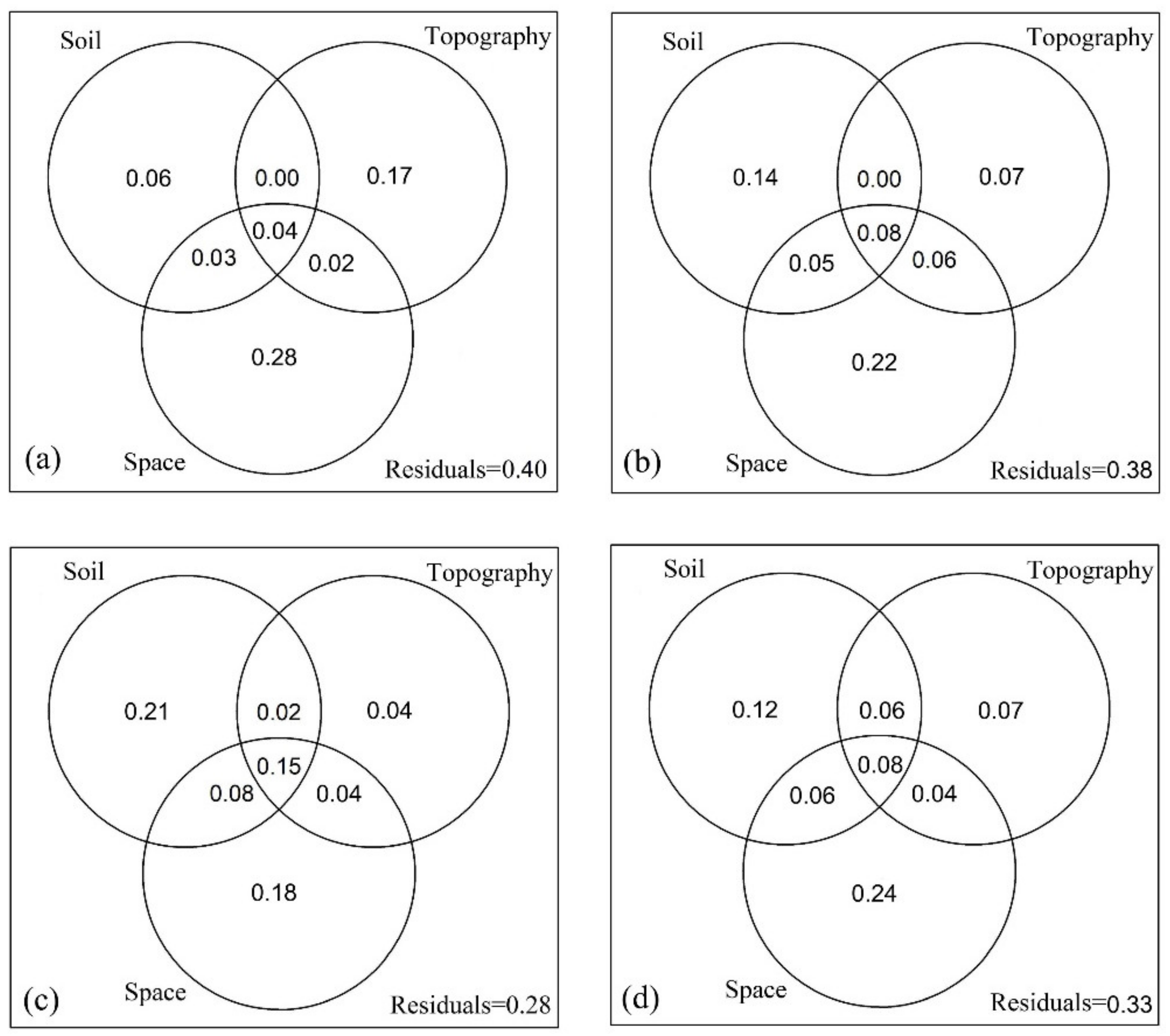
3.2. Effects of Soil, Topography and Space on Trait Dispersions Across Ontogenetic Stages
3.3. Trait Dispersions of the Multivariate Trait (AT) For All Trees Across Spatial Scales
4. Discussion
4.1. The Importance of Trait Selection, Ontogenetic Stages and Habitat Types on Community Assembly
4.2. The Variation of Trait Dispersion Explained by Soil, Topography and Space Changed with Ontogenetic Stages
4.3. The Effect of Spatial Scale on Community Assembly
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Chiang, J.M.; Spasojevic, M.J.; Muller-Landau, H.C.; Sun, I.F.; Lin, Y.; Su, S.H.; Chen, Z.S.; Chen, C.T.; Swenson, N.G.; McEwan, R.W. Functional composition drives ecosystem function through multiple mechanisms in a broadleaved subtropical forest. Oecologia 2016, 182, 829–840. [Google Scholar] [CrossRef]
- Gravel, D.; Canham, C.D.; Beaudet, M.; Messier, C. Reconciling niche and neutrality: The continuum hypothesis. Ecol. Lett. 2006, 9, 399–409. [Google Scholar] [CrossRef]
- Hubbell, S.P. The Unified Neutral Theory of Biodiversity and Biogeography; Princeton University Press: Princeton, NJ, USA, 2001; pp. 1–375. [Google Scholar]
- MacArthur, R.; Levins, R. The limiting similarity, convergence, and divergence of coexisting species. Am. Nat. 1967, 101, 377–385. [Google Scholar] [CrossRef]
- Chauvet, M.; Kunstler, G.; Roy, J.; Morin, X. Using a forest dynamics model to link community assembly processes and traits structure. Funct. Ecol. 2017, 31, 1452–1461. [Google Scholar] [CrossRef]
- Kunstler, G.; Falster, D.; Coomes, D.A.; Hui, F.; Kooyman, R.M.; Laughlin, D.C.; Poorter, L.; Vanderwel, M.; Vieilledent, G.; Wright, S.J.; et al. Plant functional traits have globally consistent effects on competition. Nature 2016, 529, 204–207. [Google Scholar] [CrossRef]
- Paine, C.E.T.; Baraloto, C.; Chave, J.; Herault, B. Functional traits of individual trees reveal ecological constraints on community assembly in tropical rain forests. Oikos 2011, 120, 720–727. [Google Scholar] [CrossRef]
- Wang, R.; Wang, Q.; Liu, C.; Kou, L.; Zhao, N.; Xu, Z.; Zhang, S.; Yu, G.; He, N. Changes in trait and phylogenetic diversity of leaves and absorptive roots from tropical to boreal forests. Plant Soil 2018, 432, 389–401. [Google Scholar] [CrossRef]
- Cornwell, W.K.; Schwilk, D.W.; Ackerly, D.D. A trait-based test for habitat filtering: Convex hull volume. Ecology 2006, 87, 1465–1471. [Google Scholar] [CrossRef]
- Swenson, N.G.; Enquist, B.J. Opposing assembly mechanisms in a Neotropical dry forest: Implications for phylogenetic and functional community ecology. Ecology 2009, 90, 2161–2170. [Google Scholar] [CrossRef] [PubMed]
- Díaz, S.; Cabido, M.; Casanoves, F. Plant functional traits and environmental filters at a regional scale. J. Veg. Sci. 2009, 9, 113–122. [Google Scholar]
- Götzenberger, L.; Botta-Dukat, Z.; Leps, J.; Partel, M.; Zobel, M.; de Bello, F. Which randomizations detect convergence and divergence in trait-based community assembly? A test of commonly used null models. J. Veg. Sci. 2016, 27, 1275–1287. [Google Scholar] [CrossRef]
- Grime, J.P. Trait convergence and trait divergence in herbaceous plant communities: Mechanisms and consequences. J. Veg. Sci. 2006, 17, 255–260. [Google Scholar] [CrossRef]
- Weiher, E.; Keddy, P.A. Assembly rules, null models, and trait dispersion: New questions from old patterns. Oikos 1995, 74, 159–164. [Google Scholar] [CrossRef]
- Lhotsky, B.; Kovacs, B.; Onodi, G.; Csecserits, A.; Redei, T.; Lengyel, A.; Kertesz, M.; Botta-Dukat, Z. Changes in assembly rules along a stress gradient from open dry grasslands to wetlands. J. Ecol. 2016, 104, 507–517. [Google Scholar] [CrossRef]
- Begon, M.; Mortimer, M.; Thompson, D.J. Population Ecology: A Unified Study of Animals and Plants, 3rd ed.; Blackwell Scientific Publications: Oxford, UK, 1996; pp. 246–247. [Google Scholar]
- Thompson, K.; Petchey, O.L.; Askew, A.P.; Dunnett, N.P.; Beckerman, A.P.; Willis, A.J. Little evidence for limiting similarity in a long-term study of a roadside plant community. J. Ecol. 2010, 98, 480–487. [Google Scholar] [CrossRef]
- Cornwell, W.K.; Ackerly, D.D. Community assembly and shifts in plant trait distributions across an environmental gradient in coastal California. Ecol. Monogr. 2009, 79, 109–126. [Google Scholar] [CrossRef]
- Price, J.; Tamme, R.; Gazol, A.; de Bello, F.; Takkis, K.; Uria-Diez, J.; Kasari, L.; Partel, M. Within-community environmental variability drives trait variability in species-rich grasslands. J. Veg. Sci. 2017, 28, 303–312. [Google Scholar] [CrossRef]
- John, R.; Dalling, J.W.; Harms, K.E.; Yavitt, J.B.; Stallard, R.F.; Mirabello, M.; Hubbell, S.P.; Valencia, R.; Navarrete, H.; Vallejo, M.; et al. Soil nutrients influence spatial distributions of tropical tree species. Proc. Natl. Acad. Sci. USA 2007, 104, 864–869. [Google Scholar] [CrossRef]
- Larsen, D.R.; Speckman, P.L. Multivariate regression trees for analysis of abundance data. Biometrics 2004, 60, 543–549. [Google Scholar] [CrossRef]
- Chapman, J.I.; McEwan, R.W. The Role of Environmental Filtering in Structuring Appalachian Tree Communities: Topographic Influences on Functional Diversity Are Mediated through Soil Characteristics. Forests 2018, 9, 19. [Google Scholar] [CrossRef]
- Kuczynski, L.; Grenouillet, G. Community disassembly under global change: Evidence in favor of the stress-dominance hypothesis. Glob. Chang. Biol. 2018, 24, 4417–4427. [Google Scholar] [CrossRef] [PubMed]
- Ramm, T.; Cantalapiedra, J.L.; Wagner, P.; Penner, J.; Roedel, M.-O.; Mueller, J. Divergent trends in functional and phylogenetic structure in reptile communities across Africa. Nat. Commun. 2018, 9, 4697. [Google Scholar] [CrossRef] [PubMed]
- Costa, D.S.; Gerschlauer, F.; Pabst, H.; Kuehnel, A.; Huwe, B.; Kiese, R.; Kuzyakov, Y.; Kleyer, M. Community-weighted means and functional dispersion of plant functional traits along environmental gradients on Mount Kilimanjaro. J. Veg. Sci. 2018, 28, 684–695. [Google Scholar] [CrossRef]
- Spasojevic, M.J.; Suding, K.N. Inferring community assembly mechanisms from functional diversity patterns: The importance of multiple assembly processes. J. Ecol. 2012, 100, 652–661. [Google Scholar] [CrossRef]
- Bernard-Verdier, M.; Navas, M.-L.; Vellend, M.; Violle, C.; Fayolle, A.; Garnier, E. Community assembly along a soil depth gradient: Contrasting patterns of plant trait convergence and divergence in a Mediterranean rangeland. J. Ecol. 2012, 100, 1422–1433. [Google Scholar] [CrossRef]
- Spasojevic, M.J.; Yablon, E.A.; Oberle, B.; Myers, J.A. Ontogenetic trait variation influences tree community assembly across environmental gradients. Ecosphere 2014, 5. [Google Scholar] [CrossRef]
- Yang, Q.; Shen, G.; Liu, H.; Wang, Z.; Ma, Z.; Fang, X.; Zhang, J.; Wang, X. Detangling the Effects of Environmental Filtering and Dispersal Limitation on Aggregated Distributions of Tree and Shrub Species: Life Stage Matters. PLoS ONE 2016, 11, e0156326. [Google Scholar] [CrossRef]
- Grubb, P.J. The maintenance of species-richness in plant communities: The importance of the regeneration niche. Biol. Rev. 1977, 52, 107–145. [Google Scholar] [CrossRef]
- Kanagaraj, R.; Wiegand, T.; Comita, L.S.; Huth, A. Tropical tree species assemblages in topographical habitats change in time and with life stage. J. Ecol. 2011, 99, 1441–1452. [Google Scholar] [CrossRef]
- Dong, T.; Zhang, Y.; Zhang, Y.; Zhang, S. Continuous planting under a high density enhances the competition for nutrients among young Cunninghamia lanceolata saplings. Ann. For. Sci. 2016, 73, 331–339. [Google Scholar] [CrossRef]
- Shi, H.; Xie, F.; Zhou, Q.; Shu, X.; Zhang, K.; Dang, C.; Feng, S.; Zhang, Q.; Dang, H. Effects of Topography on Tree Community Structure in a Deciduous Broad-Leaved Forest in North-Central China. Forests 2019, 10, 53. [Google Scholar] [CrossRef]
- Zhang, H.; Chen, H.Y.H.; Lian, J.; John, R.; Li, R.; Liu, H.; Ye, W.; Berninger, F.; Ye, Q. Using functional trait diversity patterns to disentangle the scale-dependent ecological processes in a subtropical forest. Funct. Ecol. 2018, 32, 1379–1389. [Google Scholar] [CrossRef]
- Legendre, P.; Mi, X.; Ren, H.; Ma, K.; Yu, M.; Sun, I.F.; He, F. Partitioning beta diversity in a subtropical broad-leaved forest of China. Ecology 2009, 90, 663–674. [Google Scholar] [CrossRef] [PubMed]
- Chase, J.M. Spatial scale resolves the niche versus neutral theory debate. J. Veg. Sci. 2014, 25, 319–322. [Google Scholar] [CrossRef]
- Siefert, A.; Ravenscroft, C.; Weiser, M.D.; Swenson, N.G. Functional beta-diversity patterns reveal deterministic community assembly processes in eastern North American trees. Glob. Ecol. Biogeogr. 2013, 22, 682–691. [Google Scholar] [CrossRef]
- Uriarte, M.; Condit, R.; Canham, C.D.; Hubbell, S.P. A spatially explicit model of sapling growth in a tropical forest: Does the identity of neighbours matter? J. Ecol. 2004, 92, 348–360. [Google Scholar] [CrossRef]
- Shen, Y.; Yu, S.X.; Lian, J.; Shen, H.; Cao, H.; Lu, H.; Ye, W. Inferring community assembly processes from trait diversity across environmental gradients. J. Trop. Ecol. 2016, 32, 290–299. [Google Scholar] [CrossRef]
- Li, L.; Wei, S.; Huang, Z.; Ye, W.; Cao, H. Spatial Patterns and Interspecific Associations of Three Canopy Species at Different Life Stages in a Subtropical Forest, China. J. Integr. Plant Biol. 2008, 50, 1140–1150. [Google Scholar] [CrossRef]
- Cornelissen, J.H.C.; Lavorel, S.; Garnier, E.; Diaz, S.; Buchmann, N.; Gurvich, D.E.; Reich, P.B.; ter Steege, H.; Morgan, H.D.; van der Heijden, M.G.A.; et al. A handbook of protocols for standardised and easy measurement of plant functional traits worldwide. Aust. J. Bot. 2003, 51, 335–380. [Google Scholar] [CrossRef]
- Wright, I.J.; Reich, P.B.; Westoby, M.; Ackerly, D.D.; Baruch, Z.; Bongers, F.; Cavender-Bares, J.; Chapin, T.; Cornelissen, J.H.C.; Diemer, M.; et al. The worldwide leaf economics spectrum. Nature 2004, 428, 821–827. [Google Scholar] [CrossRef]
- Vaieretti, M.V.; Diaz, S.; Vile, D.; Garnier, E. Two measurement methods of leaf dry matter content produce similar results in a broad range of species. Ann. Bot. 2007, 99, 955–958. [Google Scholar] [CrossRef] [PubMed]
- Onoda, Y.; Westoby, M.; Adler, P.B.; Choong, A.M.F.; Clissold, F.J.; Cornelissen, J.H.C.; Diaz, S.; Dominy, N.J.; Elgart, A.; Enrico, L.; et al. Global patterns of leaf mechanical properties. Ecol. Lett. 2011, 14, 301–312. [Google Scholar] [CrossRef] [PubMed]
- Kohyama, T.; Suzuki, E.; Partomihardjo, T.; Yamada, T.; Kubo, T. Tree species differentiation in growth, recruitment and allometry in relation to maximum height in a Bornean mixed dipterocarp forest. J. Ecol. 2003, 91, 797–806. [Google Scholar] [CrossRef]
- Lin, G.; Stralberg, D.; Gong, G.; Huang, Z.; Ye, W.; Wu, L. Separating the Effects of Environment and Space on Tree Species Distribution: From Population to Community. PLoS ONE 2013, 8, e56171. [Google Scholar] [CrossRef]
- Pei, N.; Lian, J.; Erickson, D.L.; Swenson, N.G.; Kress, W.J.; Ye, W.; Ge, X. Exploring Tree-Habitat Associations in a Chinese Subtropical Forest Plot Using a Molecular Phylogeny Generated from DNA Barcode Loci. PLoS ONE 2011, 6, e21273. [Google Scholar] [CrossRef]
- Comita, L.S.; Condit, R.; Hubbell, S.P. Developmental changes in habitat associations of tropical trees. J. Ecol. 2007, 95, 482–492. [Google Scholar] [CrossRef]
- Harms, K.E.; Condit, R.; Hubbell, S.P.; Foster, R.B. Habitat associations of trees and shrubs in a 50-ha neotropical forest plot. J. Ecol. 2001, 89, 947–959. [Google Scholar] [CrossRef]
- Lai, J.; Mi, X.; Ren, H.; Ma, K. Species-habitat associations change in a subtropical forest of China. J. Veg. Sci. 2009, 20, 415–423. [Google Scholar] [CrossRef]
- Valencia, R.; Foster, R.B.; Villa, G.; Condit, R.; Svenning, J.C.; Hernandez, C.; Romoleroux, K.; Losos, E.; Magard, E.; Balslev, H. Tree species distributions and local habitat variation in the Amazon: Large forest plot in eastern Ecuador. J. Ecol. 2004, 92, 214–229. [Google Scholar] [CrossRef]
- Condit, R. Tropical Forest Census Plots: Methods and Results from Barro Colorado Island, Panama and a Comparison with Other Plots; Springer: Belin, Germany, 1998. [Google Scholar]
- Condit, R.; Ashton, P.S.; Baker, P.; Bunyavejchewin, S.; Gunatilleke, S.; Gunatilleke, N.; Hubbell, S.P.; Foster, R.B.; Itoh, A.; LaFrankie, J.V.; et al. Spatial patterns in the distribution of tropical tree species. Science 2000, 288, 1414–1418. [Google Scholar] [CrossRef]
- Katabuchi, M.; Kurokawa, H.; Davies, S.J.; Tan, S.; Nakashizuka, T. Soil resource availability shapes community trait structure in a species-rich dipterocarp forest. J. Ecol. 2012, 100, 643–651. [Google Scholar] [CrossRef]
- Bivand, R.S. Spatial Econometric Functions. In R. Handbook of Applied Spatial Analysis; Springer: Berlin, Germany, 2010; pp. 53–71. [Google Scholar]
- Yang, J.; Zhang, G.; Ci, X.; Swenson, N.G.; Cao, M.; Sha, L.; Li, J.; Baskin, C.C.; Slik, J.W.F.; Lin, L. Functional and phylogenetic assembly in a Chinese tropical tree community across size classes, spatial scales and habitats. Funct. Ecol. 2014, 28, 520–529. [Google Scholar] [CrossRef]
- Ding, Y.; Zang, R.; Letcher, S.G.; Liu, S.; He, F. Disturbance regime changes the trait distribution, phylogenetic structure and community assembly of tropical rain forests. Oikos 2012, 121, 1263–1270. [Google Scholar] [CrossRef]
- Bagchi, R.; Henrys, P.A.; Brown, P.E.; Burslem, D.F.R.P.; Diggle, P.J.; Gunatilleke, C.V.S.; Gunatilleke, I.A.U.N.; Kassim, A.R.; Law, R.; Noor, S.; et al. Spatial patterns reveal negative density dependence and habitat associations in tropical trees. Ecology 2011, 92, 1723–1729. [Google Scholar] [CrossRef]
- Ribeiro, J.; Diggle, P. geoR: A package for geostatistical analysis. R News 2001, 1, 14–18. [Google Scholar]
- Kissling, W.D.; Carl, G. Spatial autocorrelation and the selection of simultaneous autoregressive models. Glob. Ecol. Biogeogr. 2008, 17, 59–71. [Google Scholar] [CrossRef]
- Gallardo, A. Spatial variability of soil properties in a floodplain forest in northwest Spain. Ecosystems 2003, 6, 564–576. [Google Scholar] [CrossRef]
- Laliberte, E.; Legendre, P. A distance-based framework for measuring functional diversity from multiple traits. Ecology 2010, 91, 299–305. [Google Scholar] [CrossRef]
- Hao, M.; Zhang, C.; Zhao, X.; von Gadow, K. Functional and phylogenetic diversity determine woody productivity in a temperate forest. Ecol. Evol. 2018, 8, 2395–2406. [Google Scholar] [CrossRef]
- Gurevitch, J.; Morrow, L.L.; Wallace, A.; Walsh, J.S. A meta-analysis of field experiments on competition. Am. Nat. 1992, 140, 539–572. [Google Scholar] [CrossRef]
- Kembel, S.W.; Hubbell, S.P. The phylogenetic structure of a neotropical forest tree community. Ecology 2006, 87, S86–S99. [Google Scholar] [CrossRef]
- Liu, X.; Swenson, N.G.; Zhang, J.; Ma, K. The environment and space, not phylogeny, determine trait dispersion in a subtropical forest. Funct. Ecol. 2013, 27, 264–272. [Google Scholar] [CrossRef]
- Oksanen, J.; Blanchet, F.G.; Friendly, M.; Kindt, R.; Legendre, P.; McGlinn, D.; Minchin, P.R.; O’Hara, R.B.; Simpson, G.L.; Solymos, P.; et al. Package ‘vegan’: Community Ecology Package. 2019. Available online: https://cran.r-project.org/web/packages/vegan/vegan.pdf (accessed on 25 April 2019).
- Yuan, Z.; Gazol, A.; Wang, X.; Lin, F.; Ye, J.; Bai, X.; Li, B.; Hao, Z. Scale specific determinants of tree diversity in an old growth temperate forest in China. Basic Appl. Ecol. 2011, 12, 488–495. [Google Scholar] [CrossRef]
- Borcard, D.; Legendre, P. All-scale spatial analysis of ecological data by means of principal coordinates of neighbour matrices. Ecol. Model. 2002, 153, 51–68. [Google Scholar] [CrossRef]
- Team, R.C. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2018. [Google Scholar]
- Kunstler, G.; Lavergne, S.; Courbaud, B.; Thuiller, W.; Vieilledent, G.; Zimmermann, N.E.; Kattge, J.; Coomes, D.A. Competitive interactions between forest trees are driven by species trait hierarchy, not phylogenetic or functional similarity: Implications for forest community assembly. Ecol. Lett. 2012, 15, 831–840. [Google Scholar] [CrossRef] [PubMed]
- Burns, J.H.; Strauss, S.Y. More closely related species are more ecologically similar in an experimental test. Proc. Natl. Acad. Sci. USA 2012, 109, 10605. [Google Scholar] [CrossRef]
- Liu, Z.; Fu, B.; Zheng, X.; Liu, G. Plant biomass, soil water content and soil N:P ratio regulating soil microbial functional diversity in a temperate steppe: A regional scale study. Soil Biol. Biochem. 2010, 42, 445–450. [Google Scholar] [CrossRef]
- Gross, N.; Liancourt, P.; Choler, P.; Suding, K.N.; Lavorel, S. Strain and vegetation effects on local limiting resources explain the outcomes of biotic interactions. Perspect. Plant Ecol. Evol. Syst. 2010, 12, 9–19. [Google Scholar] [CrossRef]
- Grime, J.P. Evidence for the existence of three primary strategies in plants and its relevance to ecological and evolutionary theory. Am. Nat. 1977, 111, 1169–1194. [Google Scholar] [CrossRef]
- Michalet, R. Is facilitation in arid environments the result of direct or complex interactions? Commentary. New Phytol. 2006, 169, 3–6. [Google Scholar] [CrossRef]
- Poorter, L.; Bongers, L.; Bongers, F. Architecture of 54 moist-forest tree species: Traits, trade-offs, and functional groups. Ecology 2006, 87, 1289–1301. [Google Scholar] [CrossRef]
- Petter, G.; Wagner, K.; Wanek, W.; Delgado, E.J.S.; Zotz, G.; Cabral, J.S.; Kreft, H. Functional leaf traits of vascular epiphytes: Vertical trends within the forest, intra- and interspecific trait variability, and taxonomic signals. Funct. Ecol. 2016, 30, 188–198. [Google Scholar] [CrossRef]
- Norden, N.; Letcher, S.G.; Boukili, V.; Swenson, N.G.; Chazdon, R. Demographic drivers of successional changes in phylogenetic structure across life-history stages in plant communities. Ecology 2012, 93, S70–S82. [Google Scholar] [CrossRef]
- Paine, C.E.T.; Beck, H. Seed predation by neotropical rain forest mammals increases diversity in seedling recruitment. Ecology 2007, 88, 3076–3087. [Google Scholar] [CrossRef] [PubMed]
- Dent, D.H.; DeWalt, S.J.; Denslow, J.S. Secondary forests of central Panama increase in similarity to old-growth forest over time in shade tolerance but not species composition. J. Veg. Sci. 2013, 24, 530–542. [Google Scholar] [CrossRef]
- Ali, A.; Lin, S.L.; He, J.K.; Kong, F.M.; Yu, J.H.; Jiang, H.S. Tree crown complementarity links positive functional diversity and aboveground biomass along large-scale ecological gradients in tropical forests. Sci. Total Environ. 2019, 656, 45–54. [Google Scholar] [CrossRef] [PubMed]
- Shipley, B. Comparative plant ecology as a tool for integrating across scales–Preface. Ann. Bot. 2007, 99, 965–966. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| LG | LS | HG | HS | MR | |
|---|---|---|---|---|---|
| Area (ha) | 6.92 | 4.6 | 3.08 | 2.92 | 2.48 |
| Elevation (m) | <326.30 | <326.30 | ≥326.30 | ≥326.30 | ≥326.30 |
| (297.50 ± 1.61) | (311.21 ± 1.64) | (382.64 ± 3.66) | (393.83 ± 4.39) | (389.45 ± 4.79) | |
| Convexity (degree) | - | - | <0.00 | >0.00 | >0.00 |
| (−4.37 ± 0.26) | (3.59 ± 0.32) | (−3.59 ± 0.38) | (5.00 ± 0.48) | (5.45 ± 0.46) | |
| Slope (degree) | <33.00 | ≥33.00 | ≥33.00 | ≥33.00 | <33.00 |
| (31.93 ± 0.76) | (34.45 ± 1.16) | (35.08 ± 1.06) | (33.43 ± 1.10) | (31.27 ± 0.98) | |
| pH | 3.77 ± 0.01a | 3.78 ± 0.01a | 3.71 ± 0.01b | 3.68 ± 0.01c | 3.74 ± 0.01b |
| SM (%) | 19.17 ± 0.23a | 19.19 ± 0.21a | 17.91 ± 0.34b | 17.80 ± 0.31b | 17.85 ± 0.28b |
| OM (g kg−1) | 58.62 ± 0.70c | 59.61 ± 0.73c | 62.92 ± 1.76b | 69.55 ± 1.52a | 57.64 ± 0.92c |
| TN (g kg−1) | 1.11 ± 0.04b | 1.52 ± 0.06a | 1.30 ± 0.08a | 1.06 ± 0.03b | 1.04 ± 0.04b |
| AN (mg kg−1) | 200.66 ± 2.86b | 219.55 ± 4.29a | 207.22 ± 5.12b | 196.47 ± 2.62bc | 187.18 ± 2.88bc |
| TK (g kg−1) | 19.18 ± 0.25a | 18.73 ± 0.36a | 16.93 ± 0.27bc | 17.59 ± 0.30b | 16.39 ± 0.43c |
| AK (mg kg−1) | 63.72 ± 1.66a | 51.76 ± 1.08b | 54.18 ± 2.61b | 53.02 ± 2.09b | 39.90 ± 0.76c |
| TP (g kg−1) | 0.30 ± 0.00a | 0.28 ± 0.00b | 0.28 ± 0.01b | 0.29 ± 0.01ab | 0.24 ± 0.00c |
| AP (mg kg−1) | 1.82 ± 0.09a | 2.07 ± 0.10a | 1.76 ± 0.09a | 1.20 ± 0.07b | 1.99 ± 0.11a |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Li, Y.; Bin, Y.; Xu, H.; Ni, Y.; Zhang, R.; Ye, W.; Lian, J. Understanding Community Assembly Based on Functional Traits, Ontogenetic Stages, Habitat Types and Spatial Scales in a Subtropical Forest. Forests 2019, 10, 1055. https://doi.org/10.3390/f10121055
Li Y, Bin Y, Xu H, Ni Y, Zhang R, Ye W, Lian J. Understanding Community Assembly Based on Functional Traits, Ontogenetic Stages, Habitat Types and Spatial Scales in a Subtropical Forest. Forests. 2019; 10(12):1055. https://doi.org/10.3390/f10121055
Chicago/Turabian StyleLi, Yanpeng, Yue Bin, Han Xu, Yunlong Ni, Ruyun Zhang, Wanhui Ye, and Juyu Lian. 2019. "Understanding Community Assembly Based on Functional Traits, Ontogenetic Stages, Habitat Types and Spatial Scales in a Subtropical Forest" Forests 10, no. 12: 1055. https://doi.org/10.3390/f10121055
APA StyleLi, Y., Bin, Y., Xu, H., Ni, Y., Zhang, R., Ye, W., & Lian, J. (2019). Understanding Community Assembly Based on Functional Traits, Ontogenetic Stages, Habitat Types and Spatial Scales in a Subtropical Forest. Forests, 10(12), 1055. https://doi.org/10.3390/f10121055