Growth and Needle Properties of Young Pinus koraiensis Sieb. et Zucc. Trees across an Elevational Gradient

Abstract
1. Introduction
2. Materials and Methods
2.1. Study Area
2.2. Plant Materials
2.3. Canopy Openness
2.4. Soil Properties
2.5. Needle Gas Exchange
2.6. Needle Chlorophyll Concentration
2.7. Needle Morphological Trait
2.8. Needle Water Band Index
2.9. Growth Properties
2.10. Needle Nutrients
2.11. Statistical Analyses
3. Results
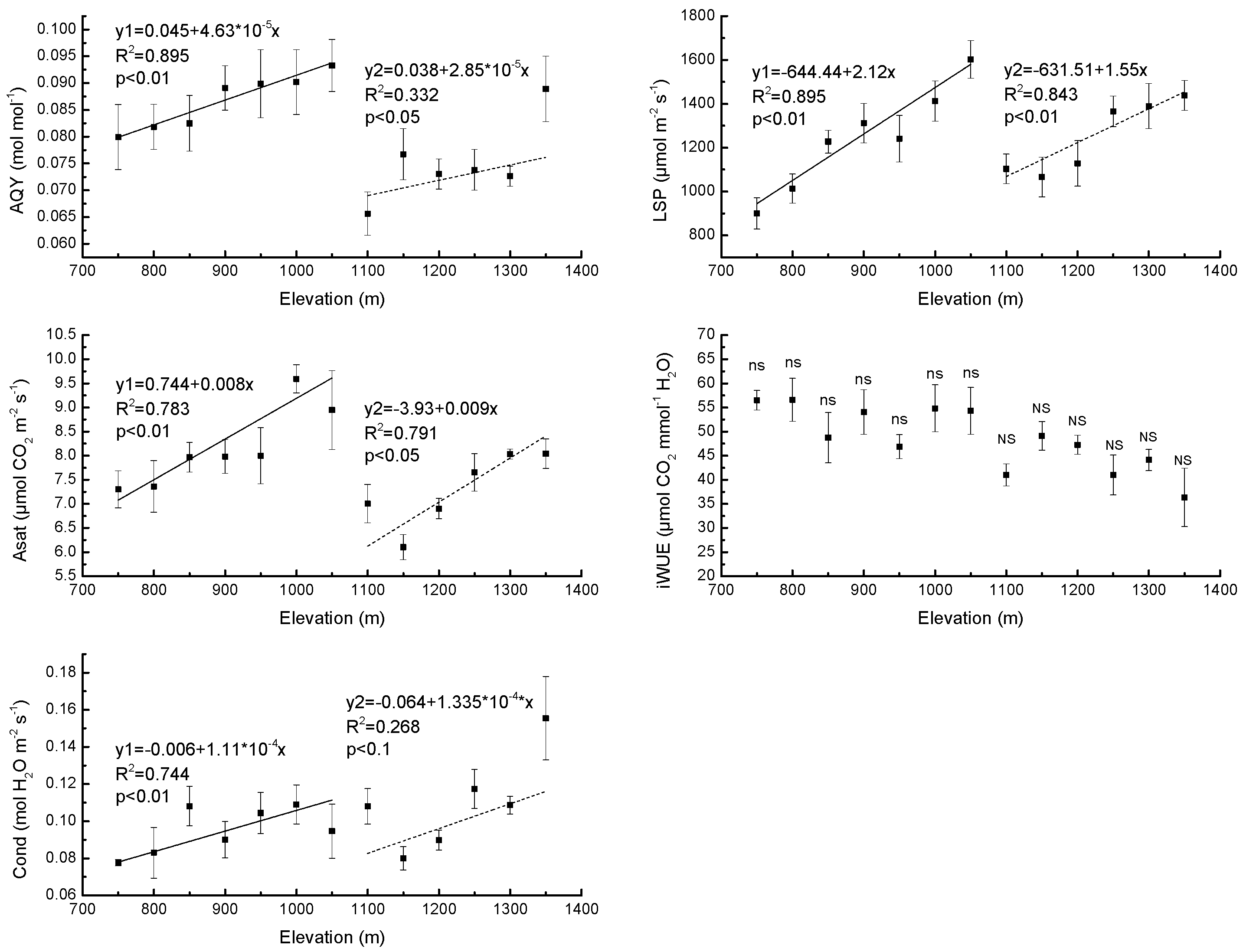
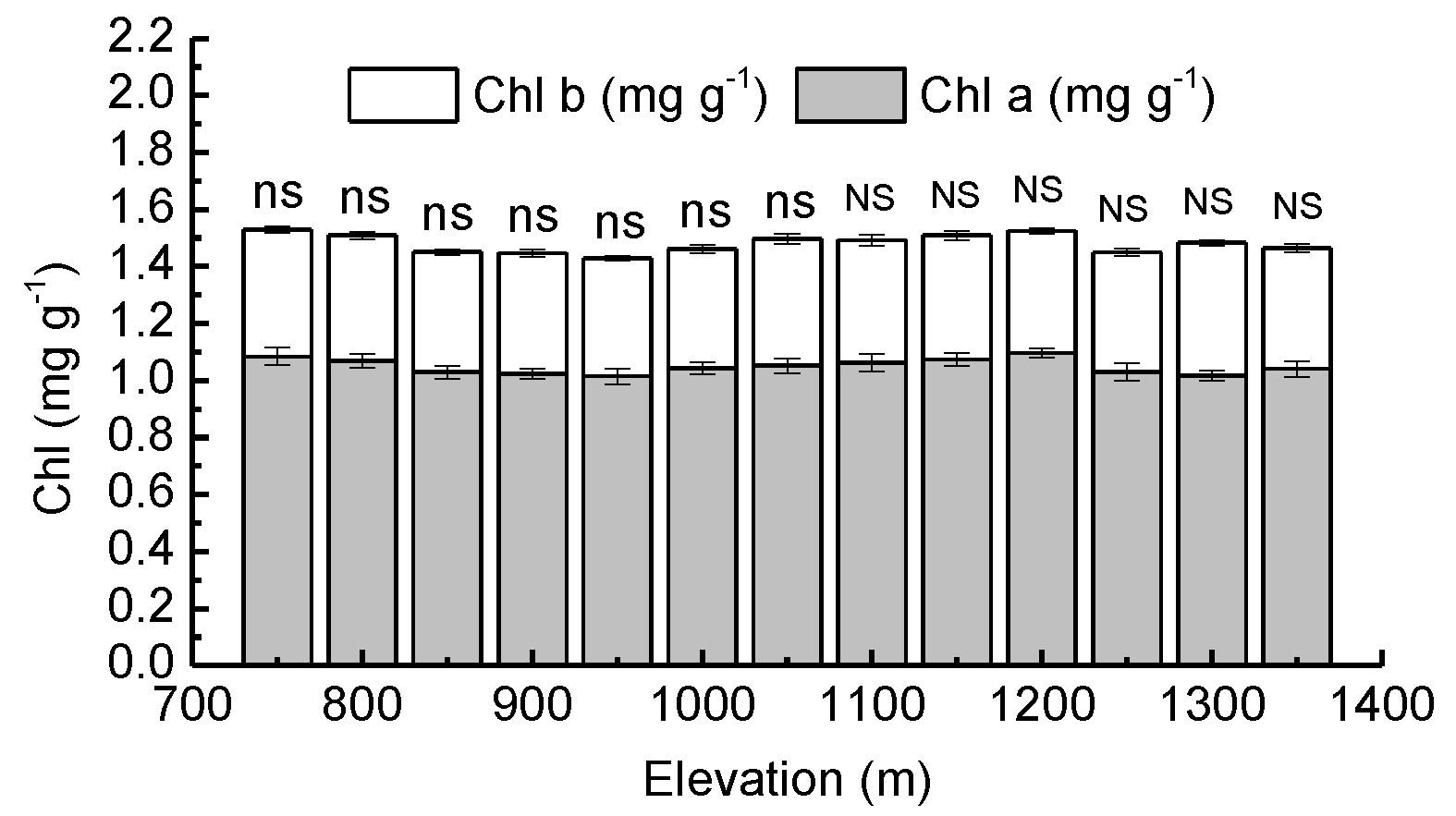
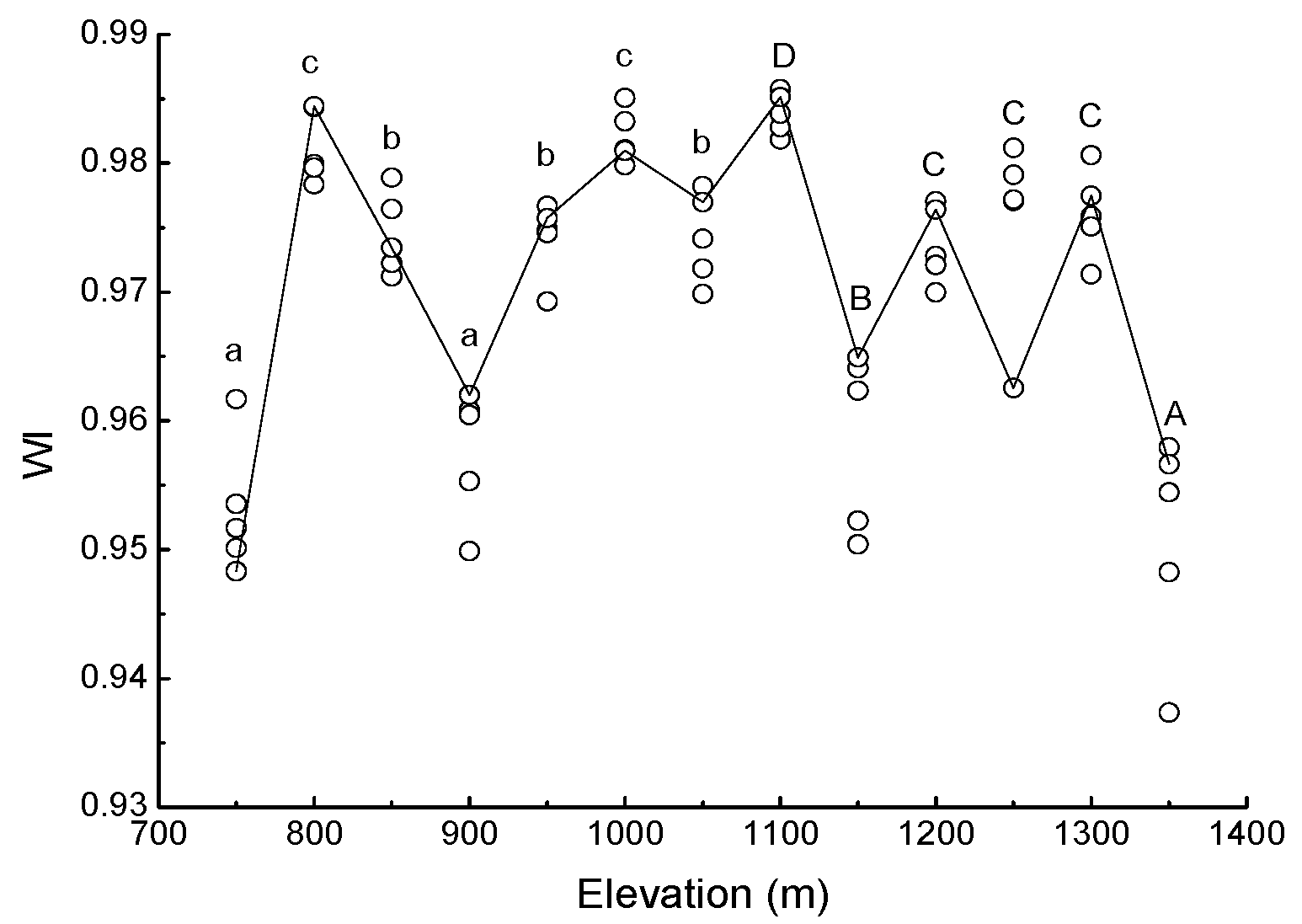
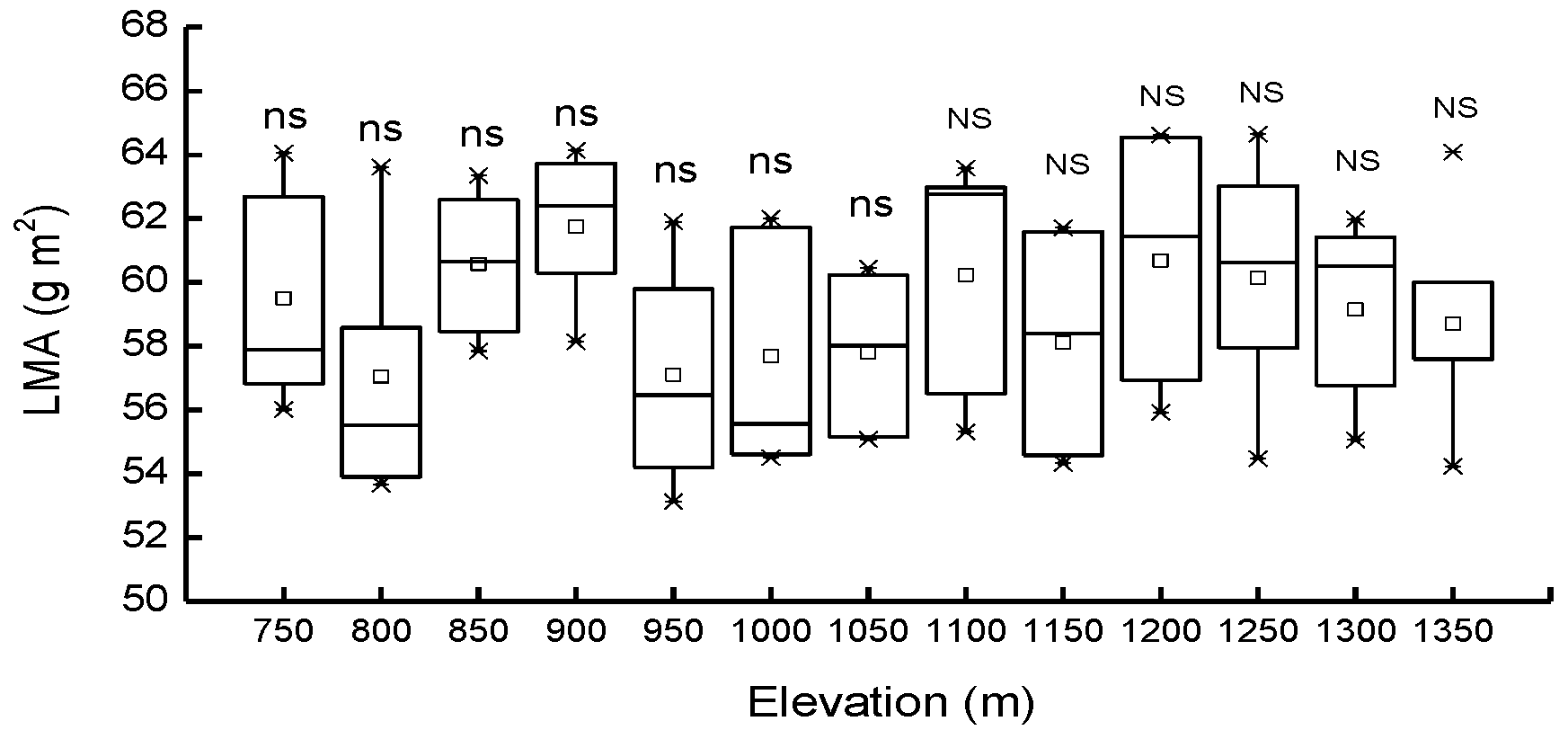
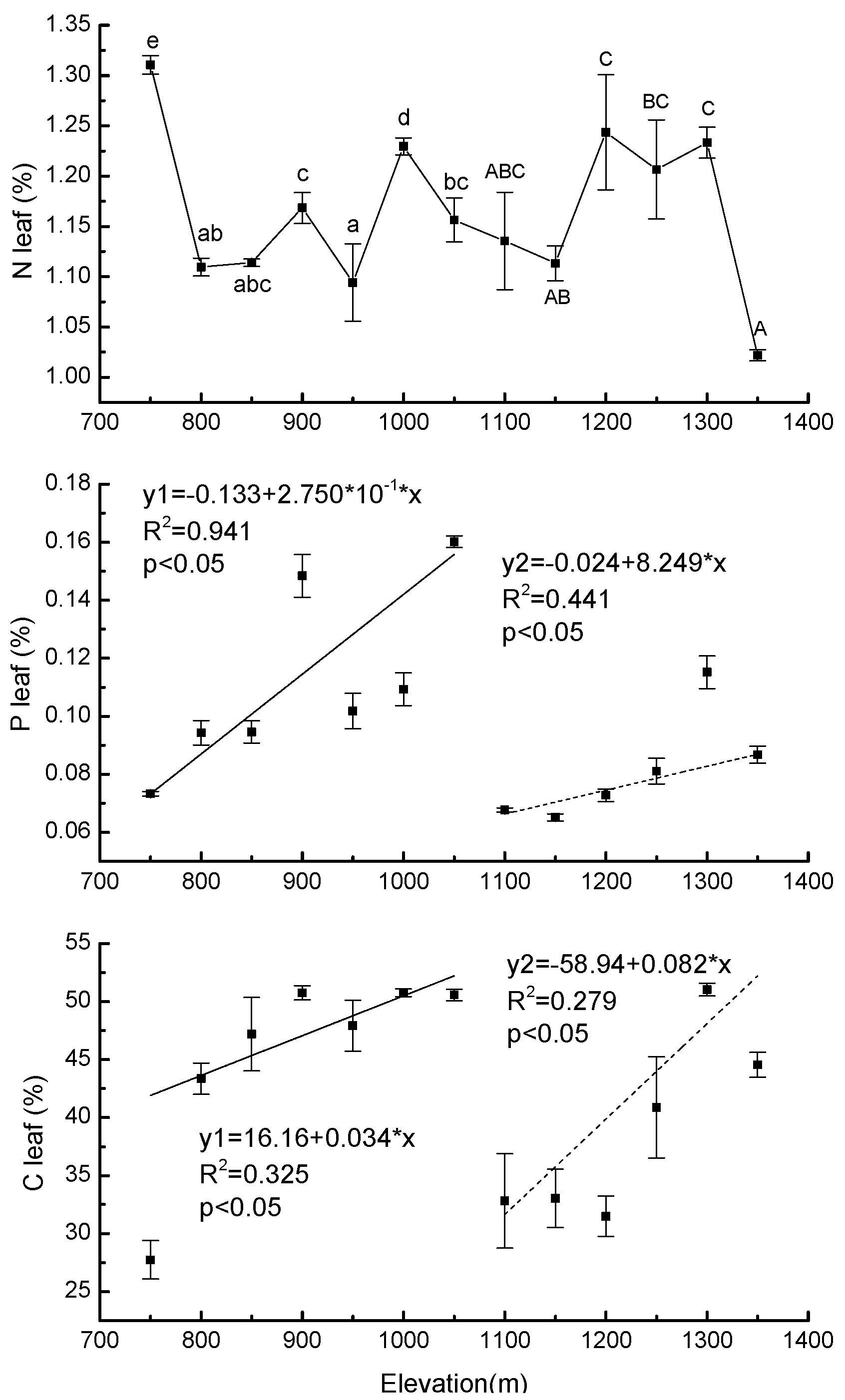
3.1. Needle Properties of Young Pinus koraiensis Trees along an Elevational Gradient in Two Different Forest Types
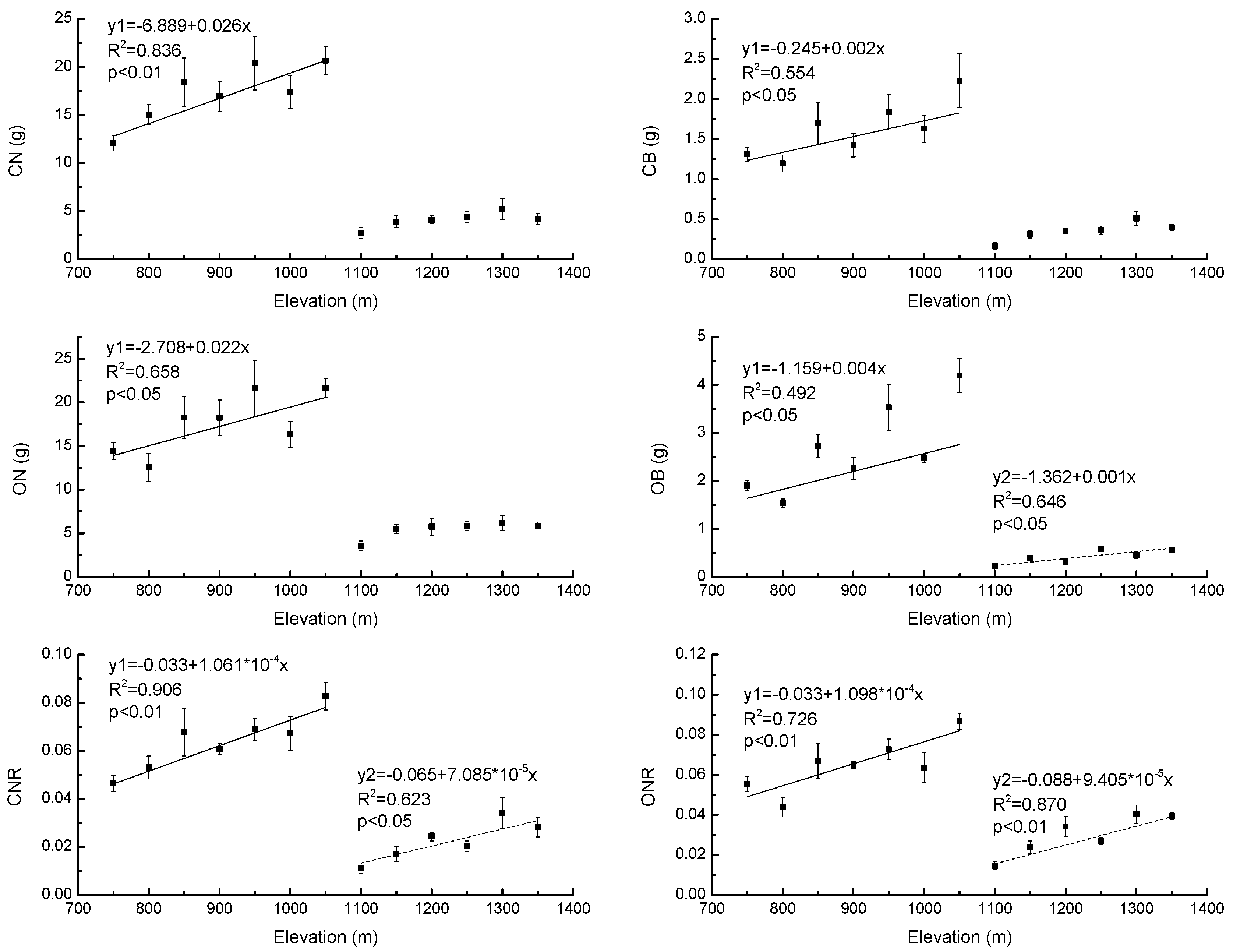
3.2. Growth Properties of Young Pinus Koraiensis Trees along an Elevational Gradient in Two Different Forest Types
4. Discussion
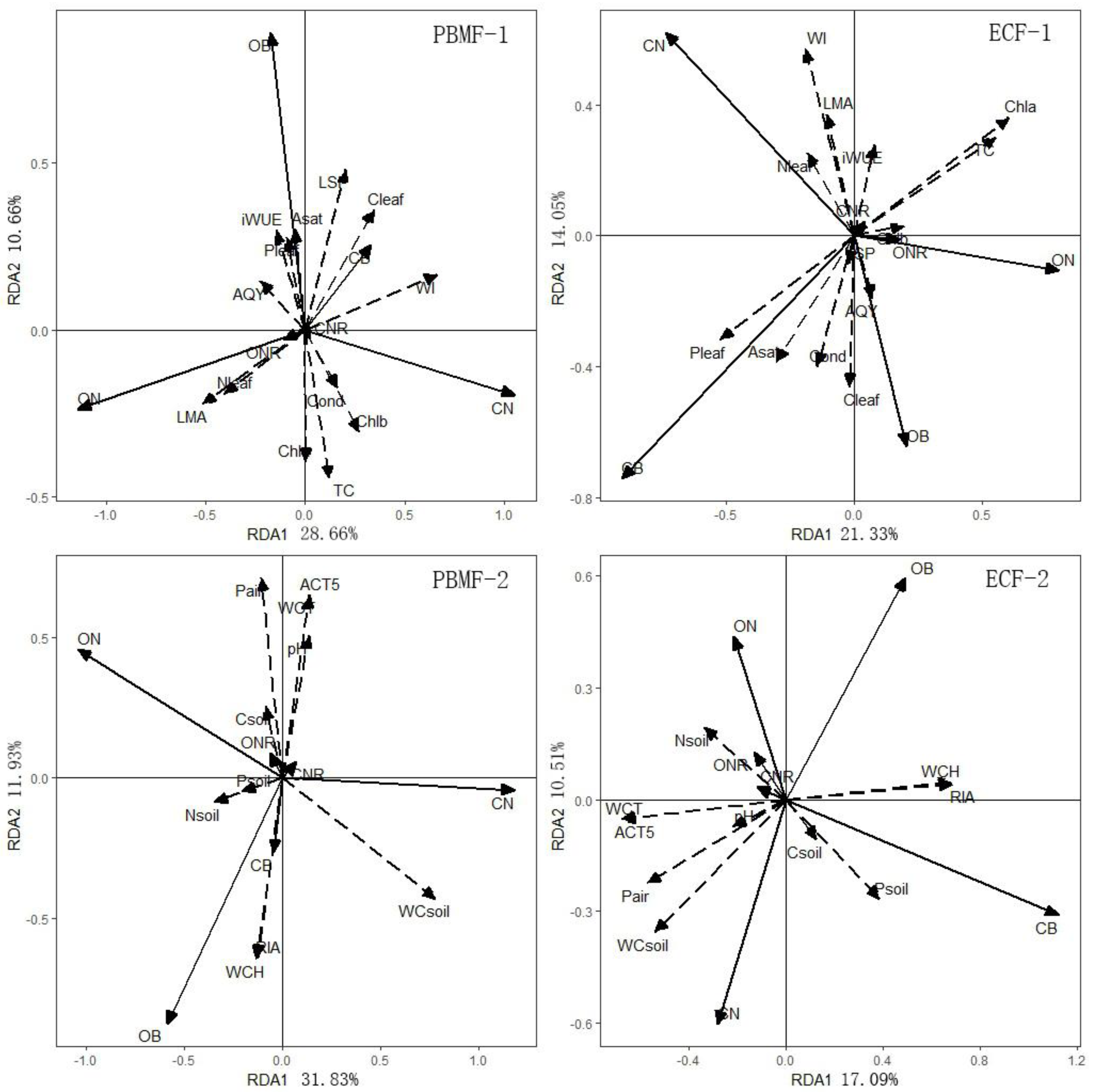
4.1. Response of Needle Properties to Elevational Gradient in Two Different Forest Types
4.2. Response of Growth Properties to Elevational Gradient in Two Different Forest Types
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Sundqvist, M.K.; Sanders, N.J.; Wardle, D.A. Community and ecosystem responses to elevational gradients: Processes, mechanisms, and insights for global change. Ann. Rev. Ecol. Evol. Syst. 2013, 44, 261–280. [Google Scholar] [CrossRef]
- Panek, J.A.; Waring, R.H. Stable carbon isotopes as indicators of limitations to forest growth imposed by climate stress. Ecol. Appl. 1997, 7, 854–863. [Google Scholar] [CrossRef]
- Miller, J.M.; Farquhar, G.D. Carbon isotope discrimination by a sequence of eucalyptus species along a subcontinental rainfall gradient in Australia. Funct. Ecol. 2001, 15, 222–232. [Google Scholar] [CrossRef]
- Kouwenberg, L.L.R.; Kürschner, W.M.; Mcelwain, J.C. Stomatal frequency change over altitudinal gradients: Prospects for paleoaltimetry. Rev. Mineral. Geochem. 2007, 66, 215–241. [Google Scholar] [CrossRef]
- Korner, C. Alpine Plant Life: Functional Plant Ecology of High Mountain Ecosystems; Springer: Berlin/Hamburg, Germany, 1999; p. 1501. [Google Scholar]
- Fukami, T.; Wardle, D.A. Long-term ecological dynamics: Reciprocal insights from natural and anthropogenic gradients. Proc. R. Soc. Lond. B Biol. Sci. 2005, 272, 2105–2115. [Google Scholar] [CrossRef] [PubMed]
- Walker, L.R.; Wardle, D.A.; Bardgett, R.D.; Clarkson, B.D. The use of chronosequences in studies of ecological succession and soil development. J. Ecol. 2010, 98, 725–736. [Google Scholar] [CrossRef]
- Malhi, Y.; Silman, M.; Salinas, N.; Bush, M.; Meir, P.; Saatchi, S. Introduction: Elevation gradients in the tropics: Laboratories for ecosystem ecology and global change research. Glob. Chang. Biol. 2010, 16, 3171–3175. [Google Scholar] [CrossRef]
- Malhi, Y.; Girardin, C.A.; Goldsmith, G.R.; Doughty, C.E.; Salinas, N.; Metcalfe, D.B.; Huaraca, H.W.; Silvaespejo, J.E.; Del, A.J.; Farfán, A.F. The variation of productivity and its allocation along a tropical elevation gradient: A whole carbon budget perspective. New Phytol. 2017, 214, 1019–1032. [Google Scholar] [CrossRef]
- Terashima, I.; Masuzawa, T.; Ohba, H.; Yokoi, Y. Is photosynthesis suppressed at higher elevations due to low CO2 pressure? Ecology 1995, 76, 2663–2668. [Google Scholar] [CrossRef]
- Wang, Q.; Iio, A.; Tenhunen, J.; Kakubari, Y. Annual and seasonal variations in photosynthetic capacity of fagus crenata along an elevation gradient in the naeba mountains, Japan. Tree Physiol. 2008, 28, 277–285. [Google Scholar] [CrossRef]
- Körner, C.; Cochrane, P.M. Stomatal responses and water relations of eucalyptus pauciflora in summer along an elevational gradient. Oecologia 1985, 66, 443–455. [Google Scholar] [CrossRef] [PubMed]
- Chen, T.; Zhao, Z.; Zhang, Y.; Qiang, W.; Feng, H.; An, L.; Li, Z. Physiological variations in chloroplasts of rhodiola coccinea along an altitudinal gradient in tianshan mountain. Acta Physiol. Plant. 2012, 34, 1007–1015. [Google Scholar] [CrossRef]
- Reinhardt, K.; Castanha, C.; Germino, M.J.; Kueppers, L.M. Ecophysiological variation in two provenances of pinus flexilis seedlings across an elevation gradient from forest to alpine. Tree Physiol. 2011, 31, 615–625. [Google Scholar] [CrossRef] [PubMed]
- Fisher, J.B.; Malhi, Y.; Cuba Torres, I.; Metcalfe, D.B.; van de Weg, M.J.; Meir, P.; Silva-Espejo, J.E.; Huaraca Huasco, W. Nutrient limitation in rainforests and cloud forests along a 3,000-m elevation gradient in the peruvian andes. Oecologia 2013, 172, 889–902. [Google Scholar] [CrossRef] [PubMed]
- Chapin, F.S.I.; Matson, P.A.I.; Mooney, H.A. Principles of terrestrial ecosystem eology. In Terrestrial Decomposition; Springer: Berlin/Hamburg, Germany, 2002. [Google Scholar]
- Kramer, P.J.; Kozlowski, T.T. 17–environmental and cultural factors affecting growth. In Physiology of Woody Plants; Academic Press: Cambridge, MA, USA, 1979; pp. 628–702. [Google Scholar]
- Jensen, A.M.; Gardiner, E.S.; Vaughn, K.C. High-light acclimation in quercus robur l. Seedlings upon over-topping a shaded environment. Environ. Exp. Bot. 2012, 78, 25–32. [Google Scholar] [CrossRef]
- Wang, Q.; Zhang, Q.; Fan, D.; Lu, C. Photosynthetic light and CO2 utilization and c4 traits of two novel super-rice hybrids. Plan. Physiol. 2006, 163, 529–537. [Google Scholar] [CrossRef] [PubMed]
- Kneeshaw, D.D.; Kobe, R.K.; Coates, K.D.; Messier, C. Sapling size influences shade tolerance ranking among southern boreal tree species. J. Ecol. 2010, 94, 471–480. [Google Scholar] [CrossRef]
- Pastur, G.M.; Lencinas, M.V.; Peri, P.L.; Arena, M. Photosynthetic plasticity of nothofagus pumilio seedlings to light intensity and soil moisture. For. Ecol. Manag. 2007, 243, 274–282. [Google Scholar] [CrossRef]
- Roberntz, P.; Stockfors, J. Effects of elevated cO2 concentration and nutrition on net photosynthesis, stomatal conductance and needle respiration of field-grown norway spruce trees. Tree Physiol. 1998, 18, 233–241. [Google Scholar] [CrossRef] [PubMed]
- Ziska, L.H.; Sullivan, J.H. Physiological sensitivity of plants along an elevational gradient to uv-b radiation. Am. J. Bot. 1992, 79, 863–871. [Google Scholar] [CrossRef]
- Huang, R.; Zhu, H.; Liu, X.; Liang, E.; Grießinger, J.; Wu, G.; Li, X.; Bräuning, A. Does increasing intrinsic water use efficiency (iwue) stimulate tree growth at natural alpine timberline on the southeastern tibetan plateau? Glob. Planet. Chang. 2017, 148, 217–226. [Google Scholar] [CrossRef]
- Peñuelas, J.; Filella, I.; Biel, C.; Serrano, L.; Savé, R. The reflectance at the 950–970 nm region as an indicator of plant water status. Int. J. Remote Sens. 1993, 14, 1887–1905. [Google Scholar] [CrossRef]
- Li, Y.; Yang, D.; Xiang, S.; Li, G. Different responses in leaf pigments and leaf mass per area to altitude between evergreen and deciduous woody species. Aust. J. Bot. 2013, 61, 424–435. [Google Scholar] [CrossRef]
- Wright, I.J.; Reich, P.B.; Westoby, M.; Ackerly, D.D.; Baruch, Z.; Bongers, F.; Cavenderbares, J.; Chapin, T.; Cornelissen, J.H.; Diemer, M. The worldwide leaf economics spectrum. Nature 2004, 428, 821–827. [Google Scholar] [CrossRef]
- Reich, P.B.; Walters, M.B.; Kloeppel, B.D.; Ellsworth, D.S. Different photosynthesis-nitrogen relations in deciduous hardwood and evergreen coniferous tree species. Oecologia 1995, 104, 24–30. [Google Scholar] [CrossRef]
- Fang, O.; Wang, Y.; Shao, X. The effect of climate on the net primary productivity (npp) of pinus koraiensis in the changbai mountains over the past 50 years. Trees 2016, 30, 281–294. [Google Scholar] [CrossRef]
- Wang, H.; Shao, X.; Jiang, Y.; Fang, X.; Wu, S. The impacts of climate change on the radial growth of pinus koraiensis along elevations of changbai mountain in northeastern china. For. Ecol. Manag. 2013, 289, 333–340. [Google Scholar] [CrossRef]
- Zhang, Y.; Drobyshev, I.; Gao, L.; Zhao, X.; Bergeron, Y. Disturbance and regeneration dynamics of a mixed korean pine dominated forest on changbai mountain, north-eastern China. Dendrochronologia 2014, 32, 21–31. [Google Scholar] [CrossRef]
- Yu, D.; Wang, Q.; Liu, J.; Zhou, W.; Qi, L.; Wang, X.; Zhou, L.; Dai, L. Formation mechanisms of the alpine erman’s birch (Betula ermanii) treeline on changbai mountain in northeast china. Trees 2014, 28, 935–947. [Google Scholar] [CrossRef]
- Hutchins, H.E.; Hutchins, S.A.; Liu, B.W. The role of birds and mammals in korean pine (Pinus koraiensis) regeneration dynamics. Oecologia 1996, 107, 120–130. [Google Scholar] [CrossRef]
- Kang, K.S.; Choi, W.Y.; Han, S.U.; Kim, C.S. Effective number and seed production in a clonal seed orchard of pznus korazenszs’. For. Genet. 2004, 11, 277–280. [Google Scholar]
- Kim, J.H.; Lee, H.J.; Jeong, S.J.; Lee, M.H.; Kim, S.H. Essential oil of pinus koraiensis leaves exerts antihyperlipidemic effects via up-regulation of low-density lipoprotein receptor and inhibition of acyl-coenzyme a: Cholesterol acyltransferase. Phytother. Res. 2012, 26, 1314–1319. [Google Scholar] [CrossRef] [PubMed]
- Cho, S.M.; Lee, E.O.; Kim, S.H.; Lee, H.J. Essential oil of pinus koraiensis inhibits cell proliferation and migration via inhibition of p21-activated kinase 1 pathway in hct116 colorectal cancer cells. BMC Complement. Altern. Med. 2014, 14, 275. [Google Scholar] [CrossRef] [PubMed]
- Li, W.H. Degradation and restoration of forest ecosystems in china. For. Ecol. Manag. 2004, 201, 33–41. [Google Scholar]
- Zhu, J.; Mao, Z.; Hu, L.; Zhang, J. Plant diversity of secondary forests in response to anthropogenic disturbance levels in montane regions of northeastern china. J. For. Res. 2007, 12, 403–416. [Google Scholar] [CrossRef]
- Sun, Y.; Zhu, J.; Sun, O.J.; Yan, Q. Photosynthetic and growth responses of pinus koraiensis seedlings to canopy openness: Implications for the restoration of mixed-broadleaved korean pine forests. Environ. Exp. Bot. 2016, 129, 118–126. [Google Scholar] [CrossRef]
- Royo, A.A.; Carson, W.P. On the formation of dense understory layers in forests worldwide: Consequences and implications for forest dynamics, biodiversity, and succession. Can. J. For. Res. 2006, 36, 1345–1362. [Google Scholar] [CrossRef]
- Fan, B.; Sang, W.; Axmacher, J.C. Forest vegetation responses to climate and environmental change: A case study from changbai mountain, ne china. For. Ecol. Manag. 2012, 262, 2052–2060. [Google Scholar]
- Jennings, S.B.; Brown, N.D.; Sheil, D. Assessing forest canopies and understorey illumination: Canopy closure, canopy cover and other measures. Forestry 1999, 72, 59–74. [Google Scholar] [CrossRef]
- Frazer, G.W.; Fournier, R.A.; Trofymow, J.; Hall, R.J. A comparison of digital and film fisheye photography for analysis of forest canopy structure and gap light transmission. Agric. For. Meteorol. 2001, 109, 249–263. [Google Scholar] [CrossRef]
- Schollenberger, C.J. A rapid approximate method for determining soil organic matter. Soil Sci. 1927, 24, 65–68. [Google Scholar] [CrossRef]
- Mitchell, A.K. Acclimation of pacific yew (Taxus brevifolia) foliage to sun and shade. Tree Physiol. 1998, 18, 749–757. [Google Scholar] [CrossRef] [PubMed]
- Isaac Berenblum, E.C. An improved method for the colorimetric determination of phosphate. Biochem. J. 1938, 32, 295–298. [Google Scholar] [CrossRef]
- Shi, P.; Körner, C.; Hoch, G. End of season carbon supply status of woody species near the treeline in western china. Basic Appl. Ecol. 2005, 7, 370–377. [Google Scholar] [CrossRef]
- Ye, Z.-P. A new model for relationship between irradiance and the rate of photosynthesis in oryza sativa. Photosynthetica 2007, 45, 637–640. [Google Scholar] [CrossRef]
- Arnon, D.I. Copper enzymes in isolated chloroplasts. Polyphenoloxidase in beta vulgaris. Plant Physiol. 1949, 24, 1. [Google Scholar] [CrossRef] [PubMed]
- Blackman, C.J.; Brodribb, T.J.; Jordan, G.J. Leaf hydraulics and drought stress: Response, recovery and survivorship in four woody temperate plant species. Plant Cell Environ. 2009, 32, 1584–1595. [Google Scholar] [CrossRef] [PubMed]
- Sims, D.A.; Gamon, J.A. Estimation of vegetation water content and photosynthetic tissue area from spectral reflectance: A comparison of indices based on liquid water and chlorophyll absorption features. Remote Sens. Environ. 2003, 84, 526–537. [Google Scholar] [CrossRef]
- Korner, C.; Diemer, M. In situ photosynthetic responses to light, temperature and carbon dioxide in herbaceous plants from low and high altitude. Funct. Ecol. 1987, 1, 179–194. [Google Scholar] [CrossRef]
- Saxe, H.; Cannell, M.G.R.; Johnsen, B.; Ryan, M.G.; Vourlitis, G. Tree and forest functioning in response to global warming. New Phytol. 2001, 149, 369–399. [Google Scholar] [CrossRef]
- Cabrera, H.; Rada, F.; Cavieres, L. Effects of temperature on photosynthesis of two morphologically contrasting plant species along an altitudinal gradient in the tropical high andes. Oecologia 1998, 114, 145–152. [Google Scholar] [CrossRef] [PubMed]
- Rada, F.; Azocar, A.; Gonzalez, J.; Briceño, B. Leaf gas exchange in espeletia schultzii wedd, a giant caulescent rosette species, along an altitudinal gradient in the venezuelan andes. Acta Oecol. 1998, 19, 73–79. [Google Scholar] [CrossRef]
- Morison, J.I.L. Stomatal response to increased CO2 concentration. J. Exp. Bot. 1998, 49, 443–452. [Google Scholar] [CrossRef]
- Heber, U.; Neimanis, S.; Lange, O.L. Stomatal aperture, photosythesis and water fluxes in mesophyll cells as affected by the abscission of leaves. Simultaneous measurements of gas exchange, light scattering and chlorphyll fluorescence. Planta 1986, 167, 554–562. [Google Scholar] [CrossRef] [PubMed]
- Bunce, J.A. Effects of boundary layer conductance on substomatal pressures of carbon dioxide. Plant Cell Environ. 1988, 11, 205–208. [Google Scholar] [CrossRef]
- And, G.D.F.; Sharkey, T.D. Stomatal conductance and photosynthesis. Ann. Rev. Plant Physiol. 1982, 33, 317–345. [Google Scholar]
- Kumar, N.; Kumar, S.; Ahuja, P.S. Photosynthetic characteristics of hordeum, triticum, rumex, and trifolium species at contrasting altitudes. Photosynthetica 2005, 43, 195–201. [Google Scholar] [CrossRef]
- Liu, W.; Fan, X.; Wang, J.; Zhang, C.; Lu, W.; Gadow, K.V. Spectral reflectance response of fraxinus mandshurica leaves to above-and belowground competition. Int. J. Remote Sens. 2012, 33, 5072–5086. [Google Scholar] [CrossRef]
- Curran, P.J. Exploring the relationship between reflectance red edge and chlorophyll content in slash pine. Tree Physiol. 1990, 7, 33–48. [Google Scholar] [CrossRef]
- Haldimann, P. Effects of changes in growth temperature on photosynthesis and carotenoid composition in zea mays leaves. Physiol. Plant. 1996, 97, 554–562. [Google Scholar] [CrossRef]
- Lefsrud, M.G.; Kopsell, D.A. Biomass production and pigment accumulation in kale grown under different radiation cycles in a controlled environment. HortScience 2006, 41, 1412–1415. [Google Scholar]
- Oleksyn, J.; Modrzýnski, J.; Tjoelker, M.; Reich, P.; Karolewski, P. Growth and physiology of picea abies populations from elevational transects: Common garden evidence for altitudinal ecotypes and cold adaptation. Funct. Ecol. 1998, 12, 573–590. [Google Scholar] [CrossRef]
- Ran, F.; Zhang, X.; Zhang, Y.; Korpelainen, H.; Li, C. Altitudinal variation in growth, photosynthetic capacity and water use efficiency of abies faxoniana rehd. Et wils. Seedlings as revealed by reciprocal transplantations. Trees 2013, 27, 1405–1416. [Google Scholar] [CrossRef]
- Lovisolo, C.; Perrone, I.; Carra, A.; Ferrandino, A.; Flexas, J.; Medrano, H.; Schubert, A. Drought-induced changes in development and function of grapevine (Vitis spp.) organs and in their hydraulic and non-hydraulic interactions at the whole-plant level: A physiological and molecular update. Funct. Plant Biol. 2010, 37, 98–116. [Google Scholar] [CrossRef]
- Chaves, M.M.; Zarrouk, O.; Francisco, R.; Costa, J.M.; Santos, T.; Regalado, A.P.; Rodrigues, M.L.; Lopes, C.M. Grapevine under deficit irrigation: Hints from physiological and molecular data. Ann. Bot. 2010, 105, 661–676. [Google Scholar] [CrossRef] [PubMed]
- Dzikiti, S.; Verreynne, J.S.; Stuckens, J.; Strever, A.; Verstraeten, W.W.; Swennen, R.; Coppin, P. Determining the water status of satsuma mandarin trees [citrus unshiu marcovitch] using spectral indices and by combining hyperspectral and physiological data. Agric. For. Meteorol. 2010, 150, 369–379. [Google Scholar] [CrossRef]
- Cibula, W.G.; Zetka, E.F.; Rickman, D.L. Response of thematic mapper bands to plant water stress. Int. J. Remote Sens. 1992, 13, 1869–1880. [Google Scholar] [CrossRef]
- Ustin, S.L.; Roberts, D.A.; Pinzón, J.; Jacquemoud, S.; Gardner, M.; Scheer, G.; Castañeda, C.M.; Palacios-Orueta, A. Estimating canopy water content of chaparral shrubs using optical methods. Remote Sens. Environ. 1998, 65, 280–291. [Google Scholar] [CrossRef]
- Korner, C.; Allison, A.; Hilscher, H. Altitudinal variation of leaf diffusive conductance and leaf anatomy in heliophytes of montane new guinea and their interrelation with microclimate. Flora 1983, 174, 91–135. [Google Scholar] [CrossRef]
- Hultine, K.; Marshall, J. Altitude trends in conifer leaf morphology and stable carbon isotope composition. Oecologia 2000, 123, 32–40. [Google Scholar] [CrossRef]
- Piper, F.I. Intraspecific trait variation and covariation in a widespread tree species (Nothofagus pumilio) in southern chile. New Phytol. 2011, 189, 259–271. [Google Scholar]
- Bresson, C.C.; Vitasse, Y.; Kremer, A.; Delzon, S. To what extent is altitudinal variation of functional traits driven by genetic adaptation in european oak and beech? Tree Physiol. 2013, 31, 1164–1174. [Google Scholar] [CrossRef]
- Smith, M. Alpine treelines: Functional ecology of the global high elevation tree limits. Mt. Res. Dev. 2013, 33, 357. [Google Scholar] [CrossRef]
- Sveinbjornsson, B.; Nordell, O.; Kauhanen, H. Nutrient relations of mountain birch growth at and below the elevational tree-line in swedish lapland. Funct. Ecol. 1992, 6, 213–220. [Google Scholar] [CrossRef]
- Kudo, G. Altitudinal effects on leaf traits and shoot growth of betulaplatyphyl. Can. J. For. Res. 2011, 25, 1881–1885. [Google Scholar] [CrossRef]
- Birmann, K.; Körner, C. Nitrogen status of conifer needles at the alpine treeline. Plant Ecol. Divers. 2009, 2, 233–241. [Google Scholar] [CrossRef]
- Schoettle, A.W.; Rochelle, S.G. Morphological variation of pinus flexilis (pinaceae), a bird-dispersed pine, across a range of elevations. Am. J. Bot. 2000, 87, 1797–1806. [Google Scholar] [CrossRef]
- Ellsworth, D.S.; Reich, P.B. Leaf mass per area, nitrogen content and photosynthetic carbon gain in acer saccharum seedlings in contrasting forest light environments. Funct. Ecol. 1992, 6, 423–435. [Google Scholar] [CrossRef]
- Le, R.X.; Walcroft, A.S.; Sinoquet, H.; Chaves, M.M.; Rodrigues, A.; Osorio, L. Photosynthetic light acclimation in peach leaves: Importance of changes in mass: Area ratio, nitrogen concentration, and leaf nitrogen partitioning. Tree Physiol. 2001, 21, 377–386. [Google Scholar]
- Pollastrini, M.; Stefano, V.D.; Ferretti, M.; Agati, G.; Grifoni, D.; Zipoli, G.; Orlandini, S.; Bussotti, F. Influence of different light intensity regimes on leaf features of vitis vinifera l. In ultraviolet radiation filtered condition. Environ. Exp. Bot. 2011, 73, 108–115. [Google Scholar] [CrossRef]
- Luomala, E.M.; Laitinen, K.; Sutinen, S.; Kellomäki, S.; Vapaavuori, E. Stomatal density, anatomy and nutrient concentrations of scots pine needles are affected by elevated co 2 and temperature. Plant Cell Environ. 2005, 28, 733–749. [Google Scholar] [CrossRef]
- Björkman, O. Responses to Different Quantum Flux Densities; Springer: Berlin/Heidelberg, Germany, 1981; pp. 57–107. [Google Scholar]
- Reich, P.B.; Ellsworth, D.S.; Walters, M.B. Leaf structure (specific leaf area) modulates photosynthesis-nitrogen relations: Evidence from within and across species and functional groups. Funct. Ecol. 1998, 12, 948–958. [Google Scholar] [CrossRef]
- Park, B.-B.; Byun, J.-K.; Park, P.-S.; Lee, S.-W.; Kim, W.-S. Growth and tissue nutrient responses of fraxinus rhynchophylla, fraxinus mandshurica, pinus koraiensis, and abies holophylla seedlings fertilized with nitrogen, phosphorus, and potassium. J. Korean Soc. For. Sci. 2010, 99, 186–196. [Google Scholar]
- Cai-Feng, Y.; Shi-Jie, H.; Yu-Mei, Z.; Cun-Guo, W.; Guan-Hua, D.; Wen-Fa, X.; Mai-He, L. Needle-age related variability in nitrogen, mobile carbohydrates, and δ13c within pinus koraiensis tree crowns. PLoS ONE 2012, 7, e35076. [Google Scholar]
- Luo, J.; Zang, R.; Li, C. Physiological and morphological variations of picea asperata populations originating from different altitudes in the mountains of southwestern china. For. Ecol. Manag. 2006, 221, 285–290. [Google Scholar] [CrossRef]
- Hedin, L.O.; Brookshire, E.J.; Menge, D.N.; Barron, A.R. The nitrogen paradox in tropical forest ecosystems. Ann. Rev. Ecol. Evol. Syst. 2009, 40, 613–635. [Google Scholar] [CrossRef]
- Vitousek, P.M.; Porder, S.; Houlton, B.Z.; Chadwick, O.A. Terrestrial phosphorus limitation: Mechanisms, implications, and nitrogen-phosphorus interactions. Ecol. Appl. 2010, 20, 5–15. [Google Scholar] [CrossRef]
- Lichtenthaler, H.K.; Ač, A.; Marek, M.V.; Kalina, J.; Urban, O. Differences in pigment composition, photosynthetic rates and chlorophyll fluorescence images of sun and shade leaves of four tree species. Plant Physiol. Biochem. 2007, 45, 577–588. [Google Scholar] [CrossRef]
- Hendrik, P.; Ulo, N.; Lourens, P.; Wright, I.J.; Rafael, V. Causes and consequences of variation in leaf mass per area (lma): A meta-analysis. New Phytol. 2010, 182, 565–588. [Google Scholar]
- Guiying Ben, C.B.O.; Sharkey, T.D. Comparisons of photosynthetic responses of xanthium strumarium and helianthus annuus to chronic and acute water stress in sun and shade. Plant Physiol. 1987, 84, 476–482. [Google Scholar]
- Rosati, A.; Esparza, G.; Dejong, T.M.; Pearcy, R.W. Influence of canopy light environment and nitrogen availability on leaf photosynthetic characteristics and photosynthetic nitrogen-use efficiency of field-grown nectarine trees. Tree Physiol. 1999, 19, 173–180. [Google Scholar] [CrossRef]
- Girardin, C.A.J.; Malhi, Y.; Aragão, L.E.O.C.; Mamani, M.; Huasco, W.H.; Durand, L.; Feeley, K.J.; Rapp, J.; Silva-Espejo, J.E.; Silman, M. Net primary productivity allocation and cycling of carbon along a tropical forest elevational transect in the peruvian andes. Glob. Chang. Biol. 2010, 16, 3176–3192. [Google Scholar] [CrossRef]
- Rapp, J.M.; Silman, M.R.; Clark, J.S.; Girardin, C.A.; Galiano, D.; Tito, R. Intra- and interspecific tree growth across a long altitudinal gradient in the peruvian andes. Ecology 2012, 93, 2061–2072. [Google Scholar] [CrossRef]
- Yu, D.; Wang, Q.; Wang, Y.; Zhou, W.; Ding, H.; Fang, X.; Jiang, S.; Dai, L. Climatic effects on radial growth of major tree species on changbai mountain. Ann. For. Sci. 2011, 68, 921–933. [Google Scholar] [CrossRef]
- Gillooly, J.F.; Charnov, E.L. Effects of size and temperature on metabolic rate. Science 2001, 293, 2248–2251. [Google Scholar] [CrossRef]
- Reiners, W.A.; Hollinger, D.Y.; Lang, G.E. Temperature and evapotranspiration gradients of the white mountains, new hampshire, USA. Arct. Alp. Res. 1984, 16, 31–36. [Google Scholar] [CrossRef]
- Dittmar, C.; Fricke, W.; Elling, W. Impact of late frost events on radial growth of common beech (Fagus sylvatica L.) in southern germany. Eur. J. For. Res. 2006, 125, 249–259. [Google Scholar] [CrossRef]
- Yan, C.; Han, S.; Zhou, Y.; Zheng, X.; Yu, D.; Zheng, J.; Dai, G.; Li, M.-H. Needle δ13c and mobile carbohydrates in pinus koraiensis in relation to decreased temperature and increased moisture along an elevational gradient in ne china. Trees 2012, 27, 389–399. [Google Scholar] [CrossRef]
- Salinas, N.; Malhi, Y.; Meir, P.; Silman, M.; Cuesta, R.R.; Huaman, J.; Salinas, D.; Huaman, V.; Gibaja, A.; Mamani, M.; et al. The sensitivity of tropical leaf litter decomposition to temperature: Results from a large-scale leaf translocation experiment along an elevation gradient in peruvian forests. New Phytol. 2011, 189, 967–977. [Google Scholar] [CrossRef]
- Surabhi, G.K.; Reddy, K.R.; Singh, S.K. Photosynthesis, fluorescence, shoot biomass and seed weight responses of three cowpea (Vigna unguiculata (L.) Walp.) cultivars with contrasting sensitivity to uv-b radiation. Environ. Exp. Bot. 2009, 66, 160–171. [Google Scholar] [CrossRef]
- Vitousek, P.M.; Sanford, R.L. Nutrient cycling in moist tropical forest. Ann. Rev. Ecol. Syst. 1986, 17, 137–167. [Google Scholar] [CrossRef]
- Asakawa, S. Further investigation on hastening the germination of pinus koraiensis seeds. J. Jpn. For. Soc. 1956, 38, 1–4. [Google Scholar]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Elevation (m) | Type of Forests | Slope Aspect | Slope (°) | WCT # (°C) | ACT5 ## (°C) | RIA ## (mm) | WCH # (%) | Pair (kPa) |
|---|---|---|---|---|---|---|---|---|
| 750 | PBMF | N | <10 | 15.55 | 2388.28 | 644.25 | 77.17 | 91.80 ± 0.07 |
| 800 | PBMF | N | <10 | 15.21 | 2329.44 | 658.97 | 77.63 | 90.60 ± 0.02 |
| 850 | PBMF | N | <10 | 14.87 | 2270.61 | 673.69 | 78.10 | 90.75 ± 0.11 |
| 900 | PBMF | N | <10 | 14.53 | 2211.77 | 688.41 | 78.56 | 90.88 ± 0.03 |
| 950 | PBMF | N | <10 | 14.19 | 2152.94 | 703.13 | 79.03 | 90.78 ± 0.02 |
| 1000 | PBMF | N | <10 | 13.85 | 2094.10 | 717.85 | 79.49 | 90.03 ± 0.06 |
| 1050 | PBMF | N | <10 | 13.51 | 2035.27 | 732.57 | 79.96 | 89.05 ± 0.02 |
| 1100 | ECF | N | <10 | 13.17 | 1976.43 | 747.29 | 80.42 | 88.49 ± 0.01 |
| 1150 | ECF | N | <10 | 12.83 | 1917.60 | 762.01 | 80.89 | 87.85 ± 0.04 |
| 1200 | ECF | N | <10 | 12.49 | 1858.76 | 776.73 | 81.35 | 88.02 ± 0.04 |
| 1250 | ECF | N | <10 | 12.15 | 1799.93 | 791.45 | 81.82 | 86.99 ± 0.09 |
| 1300 | ECF | N | <10 | 11.81 | 1741.09 | 806.17 | 82.28 | 86.94 ± 0.04 |
| 1350 | ECF | N | <10 | 11.47 | 1682.26 | 820.89 | 82.75 | 85.81 ± 0.01 |
| Elevation (m) | N Soil (%) | C Soil (%) | P Soil (%) | pH | WC Soil (%) |
|---|---|---|---|---|---|
| 750 | 0.2148 ± 0.0389 de | 5.8475 ± 0.4568 ab | 0.3492 ± 0.0258 a | 5.4492 ± 0.1319 c | 27.7338 ± 1.6653 a |
| 800 | 0.0674 ± 0.0017 a | 4.8923 ± 0.4933 a | 0.3008 ± 0.0114 a | 5.2173 ± 0.0715 bc | 43.3591 ± 1.3379 c |
| 850 | 0.1046 ± 0.0098 ab | 6.0790 ± 1.0523 ab | 0.3005 ± 0.0072 a | 5.5433 ± 0.2471 c | 47.1886 ± 3.1609 b |
| 900 | 0.1624 ± 0.0203 bcd | 4.1960 ± 0.9026 a | 0.3228 ± 0.0241 a | 5.1678 ± 0.0570 abc | 50.7534 ± 0.6221 a |
| 950 | 0.2551 ± 0.0271 e | 8.1750 ± 0.5909 b | 0.3264 ± 0.0181 a | 5.0633 ± 0.0840 ab | 47.9172 ± 2.2047 b |
| 1000 | 0.1691 ± 0.0115 cd | 4.7742 ± 0.2660 a | 0.2957 ± 0.0109 a | 5.2323 ± 0.0293 bc | 50.7662 ± 0.3353 c |
| 1050 | 0.1154 ± 0.0047 abc | 3.6204 ± 1.2684 a | 0.3159 ± 0.0104 a | 4.8427 ± 0.0378 a | 50.5732 ± 0.4940 b |
| 1100 | 0.1343 ± 0.0063 CD | 2.4571 ± 0.1983 AB | 0.2519 ± 0.0103 BC | 5.1591 ± 0.0367 D | 32.8224 ± 4.0644 D |
| 1150 | 0.0919 ± 0.0027 A | 2.2594 ± 0.6608 AB | 0.2327 ± 0.0041 AB | 4.9967 ± 0.0194 C | 33.0481 ± 2.5072 AB |
| 1200 | 0.1107 ± 0.0038 B | 3.9636 ± 0.1945 CD | 0.2419 ± 0.0044 BC | 4.8632 ± 0.0620 B | 31.4864 ± 1.7371 BC |
| 1250 | 0.1199 ± 0.0024 BC | 4.8995 ± 0.1732 D | 0.1951 ± 0.0261 A | 4.5588 ± 0.0385 A | 40.8705 ± 4.3609 C |
| 1300 | 0.0868 ± 0.0067 A | 1.5724 ± 0.2830 A | 0.2733 ± 0.0130 BC | 4.9167 ± 0.0254 BC | 51.0298 ± 0.5400 C |
| 1350 | 0.1500 ± 0.0113 D | 3.3030 ± 0.5744 BC | 0.2794 ± 0.0118 C | 4.9364 ± 0.0037 BC | 44.5513 ± 1.0680 A |
| Elevation (m) | Height (cm) | BSD (cm) | Height/BSD | Crown Length (cm) | Crown Width (cm) | Canopy Openness (%) |
|---|---|---|---|---|---|---|
| 750 | 99.21 ± 4.79 | 1.79 ± 0.12 | 56.03 ± 2.90 ns | 95.00 ± 8.34 | 86.40 ± 10.33 | 77.93 ± 1.05 ns |
| 800 | 97.82 ± 4.69 | 1.84 ± 0.10 | 53.28 ± 1.74 ns | 95.60 ± 9.57 | 85.20 ± 8.06 | 76.93 ± 1.09 ns |
| 850 | 97.72 ± 5.13 | 1.85 ± 0.11 | 52.82 ± 0.84 ns | 95.00 ± 9.99 | 84.60 ± 10.08 | 78.77 ± 0.57 ns |
| 900 | 95.73 ± 3.20 | 1.83 ± 0.14 | 53.08 ± 2.51 ns | 93.60 ± 9.24 | 83.60 ± 7.54 | 76.82 ± 0.68 ns |
| 950 | 99.36 ± 4.35 | 1.80 ± 0.11 | 55.72 ± 2.43 ns | 97.00 ± 9.32 | 86.60 ± 9.63 | 78.83 ± 0.80 ns |
| 1000 | 99.89 ± 3.43 | 1.85 ± 0.13 | 54.82 ± 3.03 ns | 98.00 ± 10.26 | 85.80 ± 9.65 | 77.86 ± 0.51 ns |
| 1050 | 98.46 ± 4.85 | 1.79 ± 0.15 | 55.98 ± 3.30 ns | 108.20 ± 9.48 | 95.20 ± 10.28 | 78.28 ± 1.08 NS |
| 1100 | 97.96 ± 4.48 | 1.83 ± 0.13 | 54.12 ± 2.21 ns | 84.40 ± 4.55 | 73.80 ± 4.85 | 79.36 ± 0.72 NS |
| 1150 | 98.36 ± 3.62 | 1.82 ± 0.14 | 54.82 ± 2.79 ns | 94.20 ± 9.93 | 86.40 ± 9.56 | 78.28 ± 0.87 NS |
| 1200 | 99.50 ± 4.55 | 1.82 ± 0.13 | 55.13 ± 2.16 ns | 97.40 ± 9.35 | 85.20 ± 10.51 | 77.56 ± 0.57 NS |
| 1250 | 97.30 ± 5.17 | 1.82 ± 0.14 | 54.04 ± 2.02 ns | 97.40 ± 9.52 | 84.40 ± 10.37 | 78.49 ± 1.08 NS |
| 1300 | 94.04 ± 4.10 | 1.90 ± 0.13 | 50.11 ± 2.36 ns | 94.80 ± 9.56 | 85.60 ± 9.14 | 78.71 ± 0.98 NS |
| 1350 | 97.85 ± 5.37 | 1.79 ± 0.12 | 54.77 ± 1.22 ns | 96.00 ± 9.36 | 83.40 ± 8.13 | 77.56 ± 1.18 ns |
| Total | CN | CB | ON | OB | CNR | ONR | ||
|---|---|---|---|---|---|---|---|---|
| Forest Type | Pr(>F) | <2.2×10−16 *** | 3.2×10−15 *** | 7.533×10−13 *** | 1.598×10−13 *** | 2.516×10−14 *** | <2.2×10−16 *** | 6.209×10−14 *** |
| F | 39.208 | 107.150 | 80.318 | 87.467 | 96.466 | 140.900 | 92.001 | |
| Elevation | Pr(>F) | 1.805×10−9 *** | 4.699×10−15 *** | 1.895×10−14 *** | 3.697×10−16 *** | <2.2×10−16 *** | <2.2×10−16 *** | 5.336×10−16 *** |
| F | 2.691 | 19.262 | 17.976 | 21.788 | 120.520 | 22.425 | 21.408 |
| Chl a | Chl b | TC | N Leaf | P Leaf | C Leaf | LMA | AQY | LSP | Asat | Cond | iWUE | WI | |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| PBMF | |||||||||||||
| CN | −0.031 | 0.187 | 0.056 | −0.232 | 0.482 ** | 0.396 * | −0.234 | 0.228 | 0.522 ** | 0.175 | 0.310 | −0.285 | 0.337 * |
| CB | −0.111 | 0.249 | 0.019 | −0.153 | 0.440 ** | 0.325 | −0.258 | 0.171 | 0.513 ** | 0.121 | 0.153 | −0.134 | 0.214 |
| ON | −0.020 | 0.115 | 0.034 | −0.114 | 0.530 ** | 0.305 | −0.090 | 0.296 | 0.491 ** | 0.203 | 0.259 | −0.250 | 0.127 |
| OB | −0.141 | 0.038 | −0.098 | −0.212 | 0.594 ** | 0.416 * | −0.201 | 0.291 | 0.607 ** | 0.266 | 0.106 | −0.081 | 0.230 |
| CNR | −0.019 | 0.173 | 0.060 | −0.252 | 0.452 ** | 0.384 * | −0.202 | 0.191 | 0.518 ** | 0.206 | 0.356 * | −0.312 | 0.337 * |
| ONR | 0.004 | 0.067 | 0.033 | −0.061 | 0.488 ** | 0.224 | 0.012 | 0.274 | 0.434 ** | 0.231 | 0.251 | −0.228 | 0.022 |
| ECF | |||||||||||||
| CN | −0.130 | 0.244 | 0.004 | 0.155 | 0.646 ** | 0.377 * | −0.005 | 0.268 | 0.512 ** | 0.300 | 0.174 | −0.052 | −0.112 |
| CB | −0.322 | 0.169 | −0.179 | 0.131 | 0.713 ** | 0.450 * | −0.100 | 0.326 | 0.458 * | 0.384 * | 0.343 | −0.197 | −0.227 |
| ON | 0.001 | 0.289 | 0.127 | 0.132 | 0.611 ** | 0.377 * | −0.027 | 0.324 | 0.538 ** | 0.288 | 0.176 | −0.006 | −0.230 |
| OB | −0.112 | 0.182 | −0.008 | −0.108 | 0.593 ** | 0.535 ** | 0.007 | 0.492 ** | 0.441 * | 0.441 * | 0.357 | −0.133 | −0.422 |
| CNR | −0.119 | 0.242 | 0.012 | 0.147 | 0.644 ** | 0.388 * | 0.024 | 0.262 | 0.521 ** | 0.306 | 0.179 | −0.054 | −0.102 |
| ONR | 0.042 | 0.303 | 0.166 | 0.106 | 0.602 ** | 0.412 * | −0.023 | 0.327 | 0.565 ** | 0.295 | 0.197 | −0.015 | −0.260 |
| Mean ± S.E. | WCT | WCH | ACT5 | RIA | Pair | N Soil | C Soil | P Soil | pH Soil | WC Soil | |
|---|---|---|---|---|---|---|---|---|---|---|---|
| PBMF | |||||||||||
| CN | 18.15 ± 6.64 a | −0.647 ** | 0.648 ** | −0.647 ** | 0.647 ** | −0.611 ** | −0.040 | −0.185 | −0.198 | −0.185 | 0.226 |
| CB | 1.71 ± 0.77 a | −0.629 ** | 0.630 ** | −0.629 ** | 0.629 ** | −0.631 ** | −0.032 | −0.287 | −0.126 | −0.292 | 0.087 |
| ON | 18.33 ± 6.58 a | −0.665 ** | 0.666 ** | −0.665 ** | 0.665 ** | −0.555 ** | 0.039 | −0.125 | −0.133 | −0.233 | −0.058 |
| OB | 2.79 ± 1.28 a | −0.795 ** | 0.796 ** | −0.795 ** | 0.795 ** | −0.747 ** | 0.045 | −0.175 | −0.060 | −0.450 ** | 0.035 |
| CNR | 0.06 ± 0.02 a | −0.616 ** | 0.616 ** | −0.616 ** | 0.616 ** | −0.563 ** | −0.033 | −0.139 | −0.209 | −0.132 | 0.238 |
| ONR | 0.06 ± 0.02 a | −0.599 ** | 0.599 ** | −0.599 ** | 0.599 ** | −0.446 ** | 0.074 | −0.059 | −0.114 | −0.182 | −0.170 |
| ECF | |||||||||||
| CN | 4.62 ± 2.89 b | −0.605 ** | 0.604 ** | −0.605 ** | 0.605 ** | −0.497 ** | −0.116 | −0.022 | 0.215 | −0.214 | −0.257 |
| CB | 0.40 ± 0.24 b | −0.734 ** | 0.734 ** | −0.734 ** | 0.734 ** | −0.594 ** | −0.100 | 0.047 | 0.423 * | −0.217 | −0.397 * |
| ON | 6.02 ± 3.16 b | −0.683 ** | 0.682 ** | −0.683 ** | 0.683 ** | −0.569 ** | −0.048 | 0.029 | 0.285 | −0.231 | −0.401 * |
| OB | 0.46 ± 0.25 b | −0.874 ** | 0.874 ** | −0.874 ** | 0.874 ** | −0.845 ** | 0.112 | 0.072 | 0.348 | −0.304 | −0.555 ** |
| CNR | 0.02 ± 0.01 b | −0.617 ** | 0.617 ** | −0.617 ** | 0.617 ** | −0.509 ** | −0.105 | 0.025 | 0.201 | −0.244 | −0.254 |
| ONR | 0.03 ± 0.01 b | −0.724 ** | 0.723 ** | −0.724 ** | 0.724 ** | −0.609 ** | −0.031 | 0.083 | 0.281 | −0.274 | −0.439 * |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Fan, Y.; Moser, W.K.; Cheng, Y. Growth and Needle Properties of Young Pinus koraiensis Sieb. et Zucc. Trees across an Elevational Gradient. Forests 2019, 10, 54. https://doi.org/10.3390/f10010054
Fan Y, Moser WK, Cheng Y. Growth and Needle Properties of Young Pinus koraiensis Sieb. et Zucc. Trees across an Elevational Gradient. Forests. 2019; 10(1):54. https://doi.org/10.3390/f10010054
Chicago/Turabian StyleFan, Ying, W. Keith Moser, and Yanxia Cheng. 2019. "Growth and Needle Properties of Young Pinus koraiensis Sieb. et Zucc. Trees across an Elevational Gradient" Forests 10, no. 1: 54. https://doi.org/10.3390/f10010054
APA StyleFan, Y., Moser, W. K., & Cheng, Y. (2019). Growth and Needle Properties of Young Pinus koraiensis Sieb. et Zucc. Trees across an Elevational Gradient. Forests, 10(1), 54. https://doi.org/10.3390/f10010054