Biodentine™ Boosts, WhiteProRoot®MTA Increases and Life® Suppresses Odontoblast Activity

 ,
,  ,
,  ,
,  ,
,  and
and 
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Materials and Methods
2.1. Materials
2.2. Sample Preparation
2.3. Cell Culture
2.4. Metabolic Activity
2.5. Cell Viability
2.6. Cell Proliferation
2.7. Cell Cycle
2.8. Types of Cell Death
2.9. Mitochondrial Membrane Potential
2.10. Cellular Morphology
2.11. Oxidative Stress
2.12. Alkaline Phosphatase—Gene Expression
2.13. Expression of Dentin Sialoprotein (DSP)
2.14. Mineralized Nodule Formation
2.15. Statistical Analysis
3. Results
3.1. Biomaterials Extracts pH
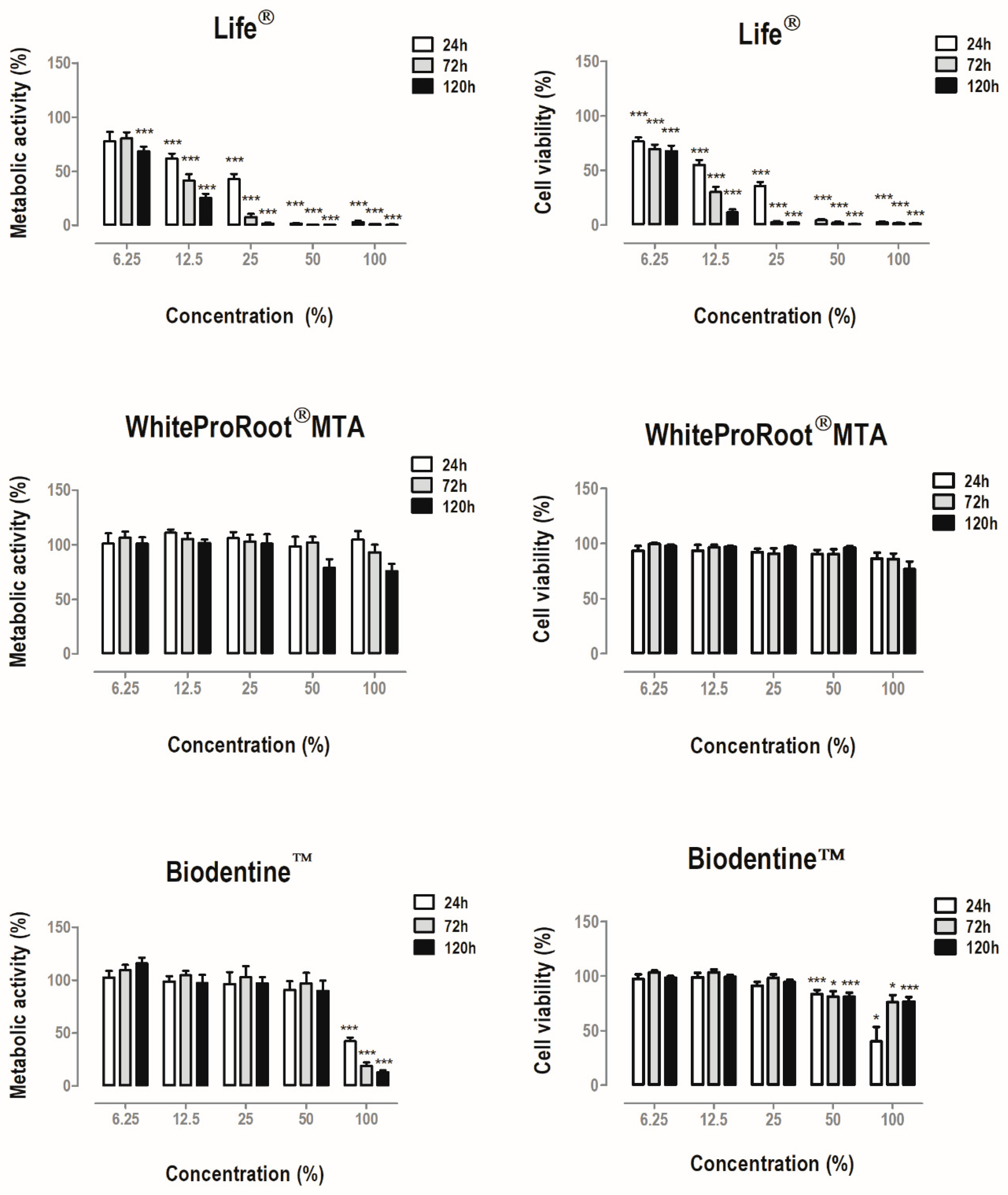
3.2. Metabolic Activity and Cell Viability
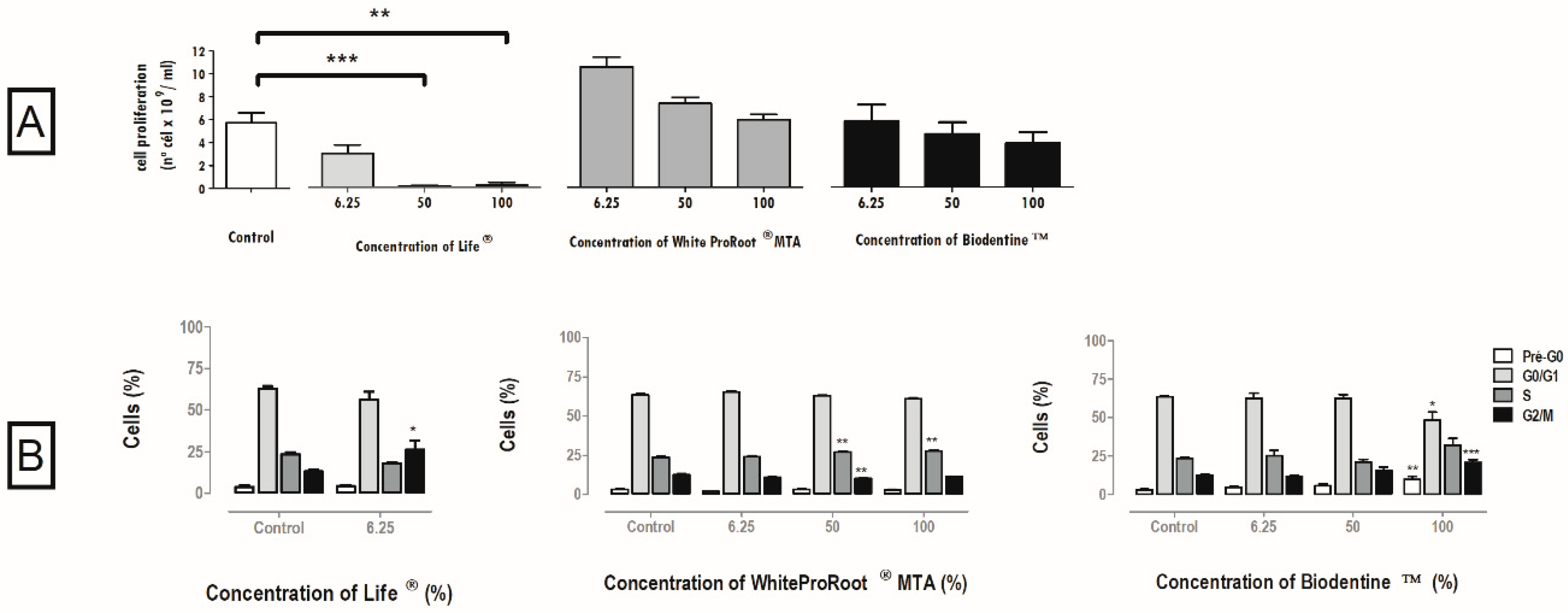
3.3. Cell Proliferation
3.4. Cell Cycle
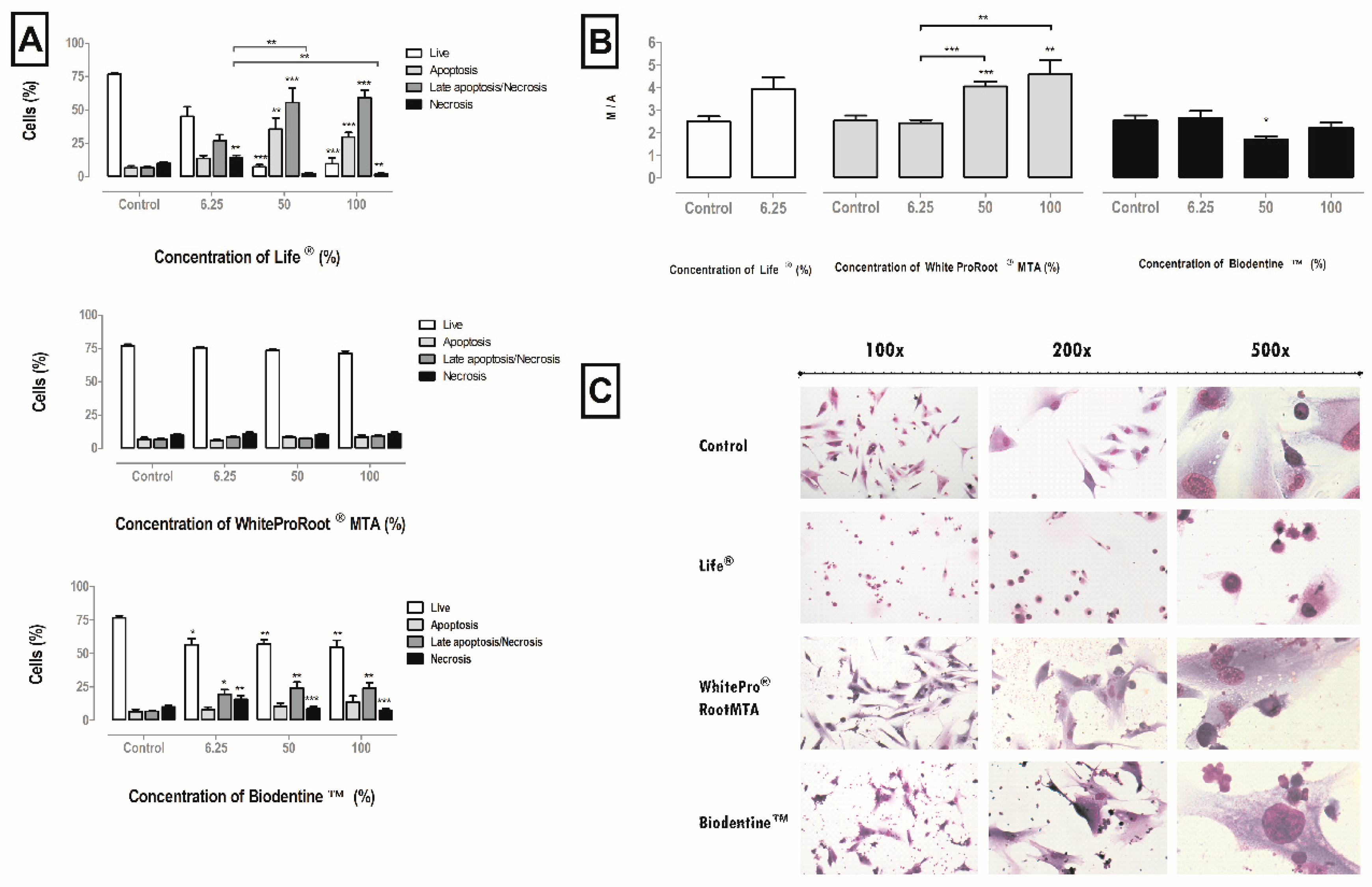
3.5. Cell Death Pathways
3.6. Mitochondrial Membrane Potential
3.7. Cellular Morphology
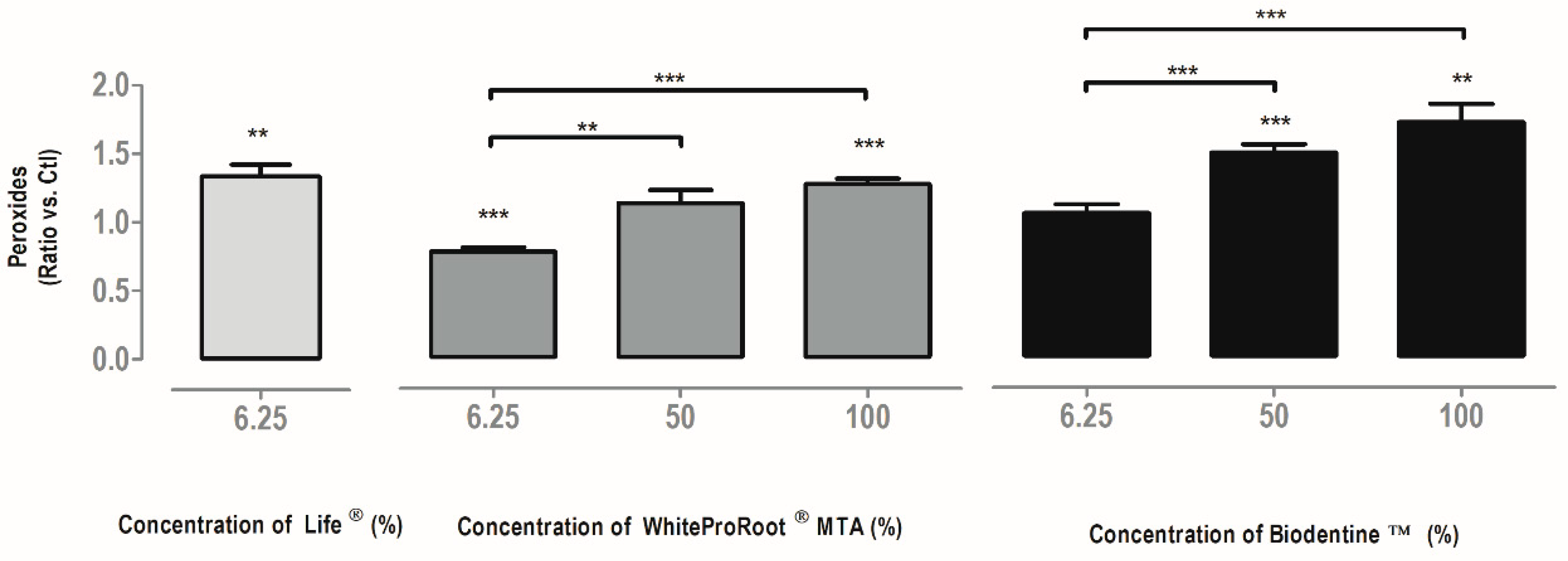
3.8. Oxidative Stress
3.9. Alkaline Phosphatase—Gene Expression
3.10. Immunocytochemistry Evaluation of Sialoprotein of Dentin (DSP)
3.11. Calcium Nodules Evaluation—Morphology and Quantification
4. Discussion
5. Conclusions
- Calcium hydroxide-based cements decreased metabolic activity and cellular viability, with a marked increase in cell death, considerable changes in the cell cycle, and no protein synthesis or calcium nodules formation.
- Mineral trioxide aggregates and tricalcium silicates materials increased metabolic activity and cell viability, with a high percentage of living cells and without interference in the cell cycle. Also, in the later stages of differentiation and mineralization, the tricalcium silicate cements had a better performance, with a marked increase of alkaline phosphatase expression, of dentin sialoprotein and calcium nodule formation compared to the mineral trioxide aggregate cements.
- This work supports tricalcium silicate, and mineral aggregate trioxide cements-based can be indicated for pulp capping therapeutics.
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Murray, P.E.; About, I.; Lumley, P.J.; Franquin, J.-C.; Windsor, L.J.; Smith, A.J. Odontoblast morphology and dental repair. J. Dent. 2003, 31, 75–82. [Google Scholar] [CrossRef]
- Sangwan, P.; Sangwan, A.; Duhan, J.; Rohilla, A. Tertiary dentinogenesis with calcium hydroxide: A review of proposed mechanisms. Int. Endod. J. 2013, 46, 3–19. [Google Scholar] [CrossRef] [PubMed]
- Ferracane, J.L.; Cooper, P.R.; Smith, A.J. Can interaction of materials with the dentin-pulp complex contribute to dentin regeneration? Odontology 2010, 98, 2–14. [Google Scholar] [CrossRef]
- Goldberg, M. The Dental Pulp; Goldberg, M., Ed.; Springer: Berlin/Heidelberg, Germany, 2014; ISBN 978-3-642-55159-8. [Google Scholar]
- Song, M.; Yu, B.; Kim, S.; Hayashi, M.; Smith, C.; Sohn, S.; Kim, E.; Lim, J.; Stevenson, R.G.; Kim, R.H. Clinical and Molecular Perspectives of Reparative Dentin Formation. Dent. Clin. North Am. 2017, 61, 93–110. [Google Scholar] [CrossRef] [PubMed]
- Linde, A.; Goldberg, M. Dentinogenesis. Crit. Rev. Oral Biol. Med. 1993, 4, 679–728. [Google Scholar] [CrossRef]
- Simon, S.; Smith, A.J.; Lumley, P.J.; Berdal, A.; Smith, G.; Finney, S.; Cooper, P.R. Molecular characterization of young and mature odontoblasts. Bone 2009, 45, 693–703. [Google Scholar] [CrossRef]
- Tziafas, D. Mechanisms controlling secondary initiation of dentinogenesis: A review. Int. Endod. J. 1994, 27, 61–74. [Google Scholar] [CrossRef]
- Tziafas, D.; Smith, A.J.; Lesot, H. Designing new treatment strategies in vital pulp therapy. J. Dent. 2000, 28, 77–92. [Google Scholar] [CrossRef]
- Farhad, A.; Mohammadi, Z. Calcium hydroxide: A review. Int. Dent. J. 2005, 55, 293–301. [Google Scholar] [CrossRef]
- Poggio, C.; Arciola, C.R.; Beltrami, R.; Monaco, A.; Dagna, A.; Lombardini, M.; Visai, L. Cytocompatibility and Antibacterial Properties of Capping Materials. Sci. World J. 2014, 2014, 1–10. [Google Scholar] [CrossRef]
- Marciano, M.A.; Duarte, M.A.H.; Camilleri, J. Dental discoloration caused by bismuth oxide in MTA in the presence of sodium hypochlorite. Clin. Oral Investig. 2015, 19, 2201–2209. [Google Scholar] [CrossRef]
- Rocha, A.C.R.; Padrón, G.H.; Garduño, M.V.G.; Aranda, R.L.G. Physicochemical analysis of MTA Angelus® and Biodentine® conducted with X ray difraction, dispersive energy spectrometry, X ray fluorescence, scanning electron microscope and infra red spectroscopy. Rev. Odontol. Mex. 2015, 19, e170–e176. [Google Scholar] [CrossRef]
- Miles, J.P.; Gluskin, A.H.; Chambers, D.; Peters, O.A. Pulp capping with mineral trioxide aggregate (MTA): A retrospective analysis of carious pulp exposures treated by undergraduate dental students. Oper. Dent. 2010, 35, 20–28. [Google Scholar] [CrossRef]
- Aeinehchi, M.; Eslami, B.; Ghanbariha, M.; Saffar, A.S. Mineral trioxide aggregate (MTA) and calcium hydroxide as pulp-capping agents in human teeth: A preliminary report. Int. Endod. J. 2003, 36, 225–231. [Google Scholar] [CrossRef]
- Akhlaghi, N.; Khademi, A. Outcomes of vital pulp therapy in permanent teeth with different medicaments based on review of the literature. Dent. Res. J. (Isfahan) 2015, 12, 406–417. [Google Scholar]
- Darvell, B.W.; Wu, R.C.T. “MTA”—An Hydraulic Silicate Cement: Review update and setting reaction. Dent. Mater. 2011, 27, 407–422. [Google Scholar] [CrossRef]
- Song, M.; Kang, M.; Kim, H.-C.; Kim, E. A Randomized Controlled Study of the Use of ProRoot Mineral Trioxide Aggregate and Endocem as Direct Pulp Capping Materials. J. Endod. 2015, 41, 11–15. [Google Scholar] [CrossRef]
- Rajasekharan, S.; Martens, L.C.; Cauwels, R.G.E.C.; Verbeeck, R.M.H. BiodentineTM material characteristics and clinical applications: A review of the literature. Eur. Arch. Paediatr. Dent. 2014, 15, 147–158. [Google Scholar] [CrossRef]
- Nuñez, C.M.; Bosomworth, H.J.; Field, C.; Whitworth, J.M.; Valentine, R.A. Biodentine and mineral trioxide aggregate induce similar cellular responses in a fibroblast cell line. J. Endod. 2014, 40, 406–411. [Google Scholar] [CrossRef]
- Malkondu, Ö.; Kazandag, M.K.; Kazazoglu, E. A review on biodentine, a contemporary dentine replacement and repair material. Biomed Res. Int. 2014, 2014, 10. [Google Scholar] [CrossRef]
- ISO 10993-5:2009 Biological Evaluation of Medical Devices—Part 5: Tests for In Vitro Cytotoxicity. 2009. Available online: https://www.iso.org/standard/36406.html (accessed on 10 April 2019).
- Zhou, H.; Shen, Y.; Wang, Z.; Li, L.; Zheng, Y.; Häkkinen, L.; Haapasalo, M. In Vitro Cytotoxicity Evaluation of a Novel Root Repair Material. J. Endod. 2013, 39, 478–483. [Google Scholar] [CrossRef]
- Wei, W.; Qi, Y.; Nikonov, S.Y.; Niu, L.; Messer, R.L.W.; Mao, J.; Primus, C.M.; Pashley, D.H.; Tay, F.R. Effects of an Experimental Calcium Aluminosilicate Cement on the Viability of Murine Odontoblast-like Cells. J. Endod. 2012, 38, 936–942. [Google Scholar] [CrossRef]
- Vichai, V.; Kirtikara, K. Sulforhodamine B colorimetric assay for cytotoxicity screening. Nat. Protoc. 2006, 1, 1112–1116. [Google Scholar] [CrossRef]
- Laranjo, M.; Serra, A.C.; Abrantes, M.; Piñeiro, M.; Gonçalves, A.C.; Casalta-Lopes, J.; Carvalho, L.; Sarmento-Ribeiro, A.B.; Rocha-Gonsalves, A.; Botelho, F. 2-Bromo-5-hydroxyphenylporphyrins for photodynamic therapy: Photosensitization efficiency, subcellular localization and in vivo studies. Photodiagnosis Photodyn. Ther. 2013, 10, 51–61. [Google Scholar] [CrossRef]
- Daltoé, M.O.; Paula-Silva, F.W.G.; Faccioli, L.H.; Gatón-Hernández, P.M.; De Rossi, A.; Bezerra Silva, L.A. Expression of Mineralization Markers during Pulp Response to Biodentine and Mineral Trioxide Aggregate. J. Endod. 2016, 42, 596–603. [Google Scholar] [CrossRef]
- Bae, W.-J.; Min, K.-S.; Kim, J.-J.; Kim, J.-J.; Kim, H.-W.; Kim, E.-C. Odontogenic responses of human dental pulp cells to collagen/nanobioactive glass nanocomposites. Dent. Mater. 2012, 28, 1271–1279. [Google Scholar] [CrossRef]
- Nozaki, T.; Takeyasu, M.; Hirao, A.; Shinohara, M.; Daito, M.; Ohura, K. Differentiation of Rat Dental Pulp-derived Cells into an Osteoblastic Lineage. Oral Sci. Int. 2005, 2, 118–125. [Google Scholar] [CrossRef]
- Osyczka, A.M.; Diefenderfer, D.L.; Bhargave, G.; Leboy, P.S. Different Effects of BMP-2 on Marrow Stromal Cells from Human and Rat Bone. Cells Tissues Organs 2004, 176, 109–119. [Google Scholar] [CrossRef]
- Atari, M.; Caballé-Serrano, J.; Gil-Recio, C.; Giner-Delgado, C.; Martínez-Sarrà, E.; García-Fernández, D.A.; Barajas, M.; Hernández-Alfaro, F.; Ferrés-Padró, E.; Giner-Tarrida, L. The enhancement of osteogenesis through the use of dental pulp pluripotent stem cells in 3D. Bone 2012, 50, 930–941. [Google Scholar] [CrossRef]
- Cai, S.; Zhang, W.; Chen, W. PDGFRβ+/c-kit+ pulp cells are odontoblastic progenitors capable of producing dentin-like structure in vitro and in vivo. BMC Oral Health 2016, 16, 113. [Google Scholar] [CrossRef]
- Salehi, S.; Cooper, P.; Smith, A.; Ferracane, J. Dentin matrix components extracted with phosphoric acid enhance cell proliferation and mineralization. Dent. Mater. 2016, 32, 334–342. [Google Scholar] [CrossRef]
- Tang, J.; Saito, T. Biocompatibility of Novel Type I Collagen Purified from Tilapia Fish Scale: An In Vitro Comparative Study. Biomed Res. Int. 2015, 2015, 1–8. [Google Scholar] [CrossRef]
- Yang, I.-S.; Lee, D.-S.; Park, J.-T.; Kim, H.-J.; Son, H.-H.; Park, J.-C. Tertiary Dentin Formation after Direct Pulp Capping with Odontogenic Ameloblast-associated Protein in Rat Teeth. J. Endod. 2010, 36, 1956–1962. [Google Scholar] [CrossRef]
- Lin, C.-Y.; Lin, H.-H.; Tsai, M.-H.; Lin, S.-P.; Chen, M.-H. Zinc Chloride for Odontogenesis of Dental Pulp Stem Cells via Metallothionein Up-regulation. J. Endod. 2011, 37, 211–216. [Google Scholar] [CrossRef]
- Man, J.; Shelton, R.M.; Cooper, P.R.; Scheven, B.A. Low-intensity Low-frequency Ultrasound Promotes Proliferation and Differentiation of Odontoblast-like Cells. J. Endod. 2012, 38, 608–613. [Google Scholar] [CrossRef]
- Wang, Y. Vascular Biology of the Placenta; Morgan & Claypool Life Sciences: San Rafael, CA, USA, 2010; Volume 2, pp. 1–98. [Google Scholar]
- Fujisawa, R.; Mizuno, M.; Tamura, M. Effect of dentin phosphoprotein on phosphate-induced apoptosis of odontoblast-like cells. Cells Tissues Organs 2009, 189, 60–64. [Google Scholar] [CrossRef]
- Eid, A.A.; Gosier, J.L.; Primus, C.M.; Hammond, B.D.; Susin, L.F.; Pashley, D.H.; Tay, F.R. In Vitro Biocompatibility and Oxidative Stress Profiles of Different Hydraulic Calcium Silicate Cements. J. Endod. 2014, 40, 255–260. [Google Scholar] [CrossRef]
- Vistica, D.T.; Skehan, P.; Scudiero, D.; Monks, A.; Pittman, A.; Boyd, M.R. Tetrazolium-based assays for cellular viability: A critical examination of selected parameters affecting formazan production. Cancer Res. 1991, 51, 2515–2520. [Google Scholar]
- Freshney, R.I. Database of misidentified cell lines. Int. J. cancer 2010, 126, 302. [Google Scholar] [CrossRef]
- Paranjpe, A.; Zhang, H.; Johnson, J.D. Effects of mineral trioxide aggregate on human dental pulp cells after pulp-capping procedures. J. Endod. 2010, 36, 1042–1047. [Google Scholar] [CrossRef]
- Yasuda, Y.; Ogawa, M.; Arakawa, T.; Kadowaki, T.; Saito, T. The Effect of Mineral Trioxide Aggregate on the Mineralization Ability of Rat Dental Pulp Cells: An In Vitro Study. J. Endod. 2008, 34, 1057–1060. [Google Scholar] [CrossRef]
- Luczaj-Cepowicz, E.; Marczuk-Kolada, G.; Pawinska, M.; Obidzinska, M.; Holownia, A. Evaluation of cytotoxicity and pH changes generated by various dental pulp capping materials—an in vitro study. Folia Histochem. Cytobiol. 2017, 55, 86–93. [Google Scholar] [CrossRef][Green Version]
- Poggio, C.; Ceci, M.; Beltrami, R.; Dagna, A.; Colombo, M.; Chiesa, M. Biocompatibility of a new pulp capping cement. Ann. Stomatol. (Roma) 2014, 5, 69–76. [Google Scholar] [CrossRef]
- Camargo, S.E.A.; Camargo, C.H.R.; Hiller, K.-A.; Rode, S.M.; Schweikl, H.; Schmalz, G. Cytotoxicity and genotoxicity of pulp capping materials in two cell lines. Int. Endod. J. 2009, 42, 227–237. [Google Scholar] [CrossRef]
- Pervaiz, S.; Clement, M.-V. Superoxide anion: Oncogenic reactive oxygen species? Int. J. Biochem. Cell Biol. 2007, 39, 1297–1304. [Google Scholar] [CrossRef]
- Zanini, M.; Sautier, J.M.; Berdal, A.; Simon, S. Biodentine induces immortalized murine pulp cell differentiation into odontoblast-like cells and stimulates biomineralization. J. Endod. 2012, 38, 1220–1226. [Google Scholar] [CrossRef]
- Pereira Paula, A.B.; Laranjo, M.; Marto, C.-M.; Paulo, S.; Abrantes, A.; Casalta-Lopes, J.; Marques-Ferreira, M.; Botelho, M.F.; Carrilho, E. Direct pulp capping: What is the most effective therapy?—Systematic review and meta-analysis. J. Evid. Based Dent. Pract. 2018, 4, 298–314. [Google Scholar] [CrossRef]
- Jalan, A.L.; Warhadpande, M.M.; Dakshindas, D.M. A comparison of human dental pulp response to calcium hydroxide and Biodentine as direct pulp-capping agents. J. Conserv. Dent. 2017, 20, 129–133. [Google Scholar] [CrossRef]
- Rathinam, E.; Rajasekharan, S.; Chitturi, R.T.; Martens, L.; De Coster, P. Gene Expression Profiling and Molecular Signaling of Dental Pulp Cells in Response to Tricalcium Silicate Cements: A Systematic Review. J. Endod. 2015, 41, 1805–1817. [Google Scholar] [CrossRef]
- Rathinam, E.; Rajasekharan, S.; Chitturi, R.T.; Declercq, H.; Martens, L.; De Coster, P. Gene Expression Profiling and Molecular Signaling of Various Cells in Response to Tricalcium Silicate Cements: A Systematic Review. J. Endod. 2016, 42, 1713–1725. [Google Scholar] [CrossRef]
- Loison-Robert, L.S.; Tassin, M.; Bonte, E.; Berbar, T.; Isaac, J.; Berdal, A.; Simon, S.; Fournier, B.P.J. In vitro effects of two silicate-based materials, Biodentine and BioRoot RCS, on dental pulp stem cells in models of reactionary and reparative dentinogenesis. PLoS ONE 2018, 13, e0190014. [Google Scholar] [CrossRef]
- Widbiller, M.; Lindner, S.R.; Buchalla, W.; Eidt, A.; Hiller, K.-A.; Schmalz, G.; Galler, K.M. Three-dimensional culture of dental pulp stem cells in direct contact to tricalcium silicate cements. Clin. Oral Investig. 2016, 20, 237–246. [Google Scholar] [CrossRef]





© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Paula, A.; Laranjo, M.; Marto, C.M.; Abrantes, A.M.; Casalta-Lopes, J.; Gonçalves, A.C.; Sarmento-Ribeiro, A.B.; Ferreira, M.M.; Botelho, M.F.; Carrilho, E. Biodentine™ Boosts, WhiteProRoot®MTA Increases and Life® Suppresses Odontoblast Activity. Materials 2019, 12, 1184. https://doi.org/10.3390/ma12071184
Paula A, Laranjo M, Marto CM, Abrantes AM, Casalta-Lopes J, Gonçalves AC, Sarmento-Ribeiro AB, Ferreira MM, Botelho MF, Carrilho E. Biodentine™ Boosts, WhiteProRoot®MTA Increases and Life® Suppresses Odontoblast Activity. Materials. 2019; 12(7):1184. https://doi.org/10.3390/ma12071184
Chicago/Turabian StylePaula, Anabela, Mafalda Laranjo, Carlos Miguel Marto, Ana Margarida Abrantes, João Casalta-Lopes, Ana Cristina Gonçalves, Ana Bela Sarmento-Ribeiro, Manuel M. Ferreira, Maria Filomena Botelho, and Eunice Carrilho. 2019. "Biodentine™ Boosts, WhiteProRoot®MTA Increases and Life® Suppresses Odontoblast Activity" Materials 12, no. 7: 1184. https://doi.org/10.3390/ma12071184
APA StylePaula, A., Laranjo, M., Marto, C. M., Abrantes, A. M., Casalta-Lopes, J., Gonçalves, A. C., Sarmento-Ribeiro, A. B., Ferreira, M. M., Botelho, M. F., & Carrilho, E. (2019). Biodentine™ Boosts, WhiteProRoot®MTA Increases and Life® Suppresses Odontoblast Activity. Materials, 12(7), 1184. https://doi.org/10.3390/ma12071184